

Article Search
Volume 27.2
May–August 2024
Full table of contents
ISSN: 1094-8074, web version;
1935-3952, print version
Recent Research Articles
See all articles in 27.2 May-August 2024
See all articles in 27.1 January-April 2024
See all articles in 26.3 September-December 2023
 Anna K. Gillespie*, Palaeontology, Geobiology and Earth Archives (PANGEA) Research Centre, School of Biological, Earth and Environmental Sciences, University of New South Wales, Sydney 2052, Australia; a.gillespie@unsw.edu.au, *corresponding author
Anna K. Gillespie*, Palaeontology, Geobiology and Earth Archives (PANGEA) Research Centre, School of Biological, Earth and Environmental Sciences, University of New South Wales, Sydney 2052, Australia; a.gillespie@unsw.edu.au, *corresponding author
Anna Gillespie is a research assistant and senior preparator at the Vertebrate Palaeontology Laboratory at the University of New South Wales. She received her PhD from the University of New South Wales in 2007. Her doctorate focussed on the marsupial lions recovered from the Riversleigh World Heritage Area of Australia. She has been one of the chief preparators of the Riversleigh WHA fossil material since 1992. Her fields of interest include the evolution of Australian vertebrates and in particular, the evolution of marsupials, marsupial lion taxonomy and systematics, and functional morphology.

 Michael Archer, Palaeontology, Geobiology and Earth Archives (PANGEA) Research Centre, School of Biological, Earth and Environmental Sciences, University of New South Wales, Sydney 2052, Australia; m.archer@unsw.edu.au
Michael Archer, Palaeontology, Geobiology and Earth Archives (PANGEA) Research Centre, School of Biological, Earth and Environmental Sciences, University of New South Wales, Sydney 2052, Australia; m.archer@unsw.edu.au
Mike Archer was born in Sydney but lived in the USA until 1967 when he moved back to Australia. By age 11 he was addicted to fossil and living animals. Degrees: Princeton University BA, University of Western Australia PhD. Key positions: Professor, University of New South Wales; Director of the Australian Museum, Sydney; Dean of Science, University of New South Wales. Research foci (~16 books & >300 refereed journal articles) include: evolution of mammals, in particular monotremes and marsupials; evolution of Australia's terrestrial environments; biocorrelation of Australia's Cenozoic vertebrates; impacts of palaeoclimate; paleoconservation; innovative biofuel company; conservation through sustainable use of native resources; native animals as pets; battling Creationism; dental function and ontogeny; megafaunal extinctions; biogeographic history of New Zealand; early Cenozoic biotas of Argentina; megatsunamis; lacustrine and karst geology; etcetera. We started the Thylacine & Lazarus Projects to see how far we could get in efforts to recover, revitalize and enable ancient DNA to reconstruct living organisms. For more info: http://www.bees.unsw.edu.au/michael-archer, http://www.create.unsw.edu.au/team/marcher/.
 Suzanne J. Hand, Palaeontology, Geobiology and Earth Archives (PANGEA) Research Centre, School of Biological, Earth and Environmental Sciences, University of New South Wales, Sydney 2052, Australia; s.hand@unsw.edu.au
Suzanne J. Hand, Palaeontology, Geobiology and Earth Archives (PANGEA) Research Centre, School of Biological, Earth and Environmental Sciences, University of New South Wales, Sydney 2052, Australia; s.hand@unsw.edu.au
Suzanne Hand is a vertebrate palaeontologist and Professor at the University of New South Wales where she teaches zoology and geology. Her research interests are largely in the areas of evolutionary biology, functional morphology, phylogenetics, and biogeography, with a special interest in fossil and modern mammals, particularly bats. She co-leads research of the World Heritage-listed Riversleigh fossil deposits of northern Australia, and is also currently researching fossil mammal faunas in Europe, Asia, New Zealand and New Caledonia. She is the author of 150+ publications on Australian fossil mammals including several books.
FIGURE 1. Microleo attenboroughi n. gen. et sp., Holotype QM F41143: 1) right maxilla and 2) left maxilla in occlusal view, stereo images; 3) interpretive drawing of right maxilla; 4) interpretive drawing of left maxilla. 5) Paratype QM F42676, occlusal views of m3 (stereophotos). Abbreviations: aabc, accessory anterobuccal cusp; ac, anterior cusp; lb, longitudinal blade; mcl, metaconule; mcus, medial cusp; me, metacone; pa, paracone; pc, posterior cusp; pr, protocone.
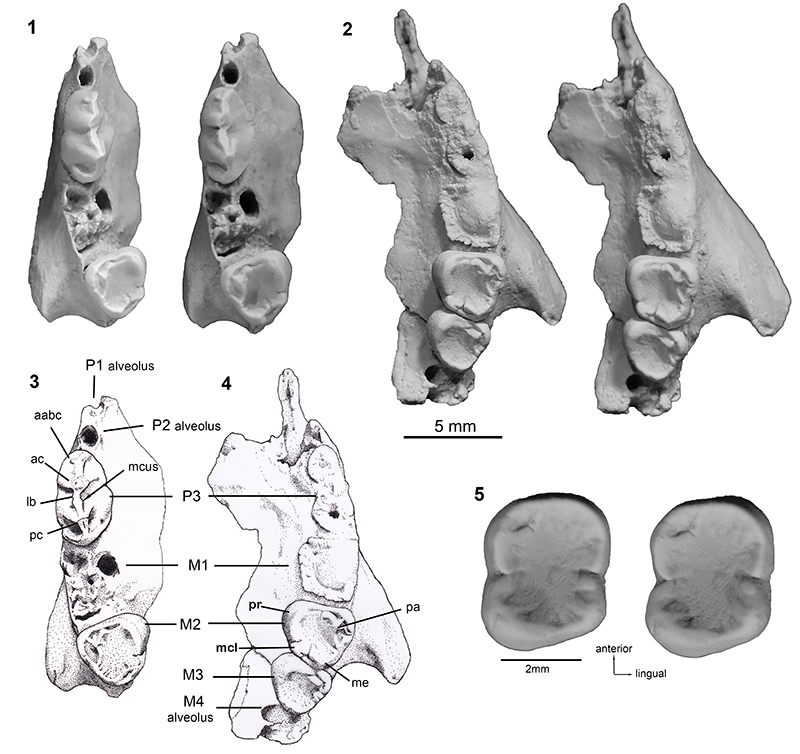
FIGURE 2. Microleo attenboroughi n. gen. et sp., Holotype QM F41143, right maxilla. 1) buccal view; 2) interpretive drawing in buccal view; 3) lingual view; 4 ) interpretive drawing in lingual view. Abbreviations: aabc, accessory anterobuccal cusp; abc, anterobuccal blade; ac, anterior cusp; alc, anterolingual crest; lb, longitudinal blade; mcl, metaconule; mcus, medial cusp; me, metacone; pa, paracone; pbb, posterobuccal basin; pbc, posterobuccal crest; pc, posterior cusp; plc, posterolingual crest; pr, protocone. Scale bar equals 5 mm.
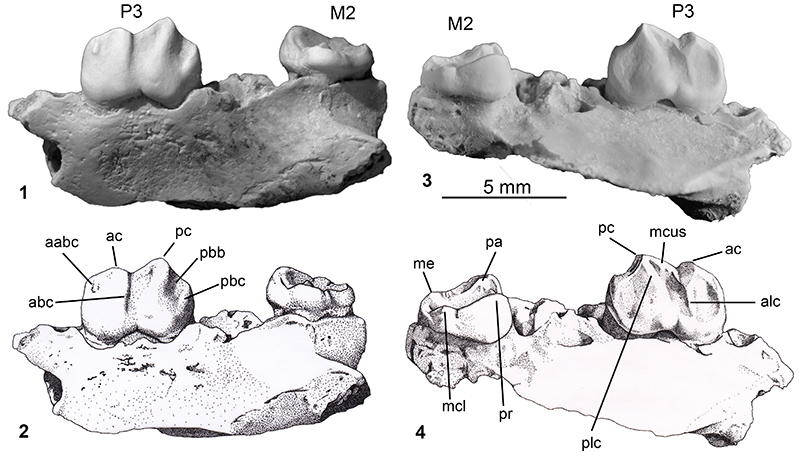
FIGURE 3. Cladistic relationships of Microleo attenboroughi within Thylacoleonidae and Vombatiformes: 1) strict consensus tree of nine most parsimonious trees obtained in the phylogenetic analysis (tree length = 272 steps; see Appendices 1, 2); 2) time-tree of thylacoleonid phylogeny.
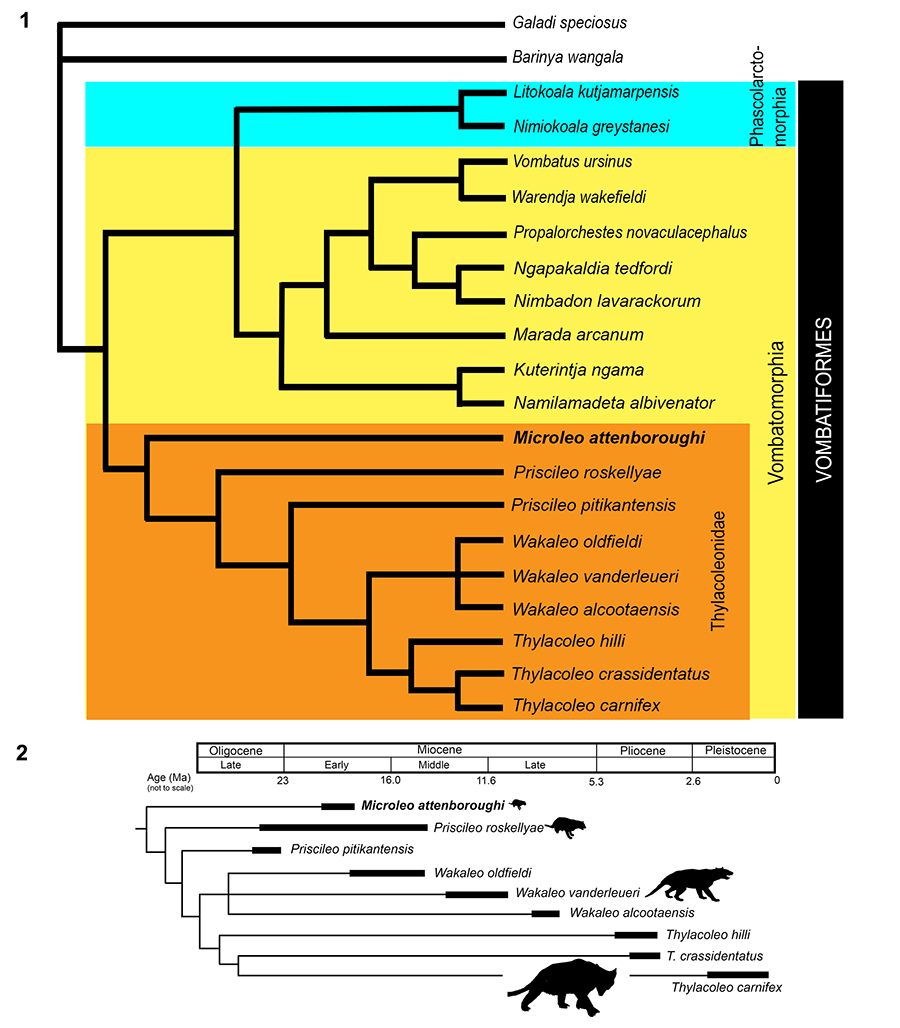
SUPPLEMENTARY FIGURE 1. Accessory anterobuccal cusp development on P3s of Thylacoleo carnifex ( 1-4) and Microleo attenboroughi ( 5, buccal view; 6, occlusal view). In a sample of 14 Thylacoleo carnifex P3s examined in the Australian Museum collection, an anterobuccal cusp was distinct in two specimens: AM F106836 (1, occlusal view; 3, buccal view); and AM F16609 (2, occlusal view; 4, buccal view). However, in T. carnifex premolars exhibiting relatively little wear, a short, broad, gently-rounded swelling was present in this region and in heavily worn specimens the entire anterobuccal surface was smooth which suggests that this feature may be relatively common but obliterated early by wear. Arrows indicate accessory anterobuccal crest (aabc).
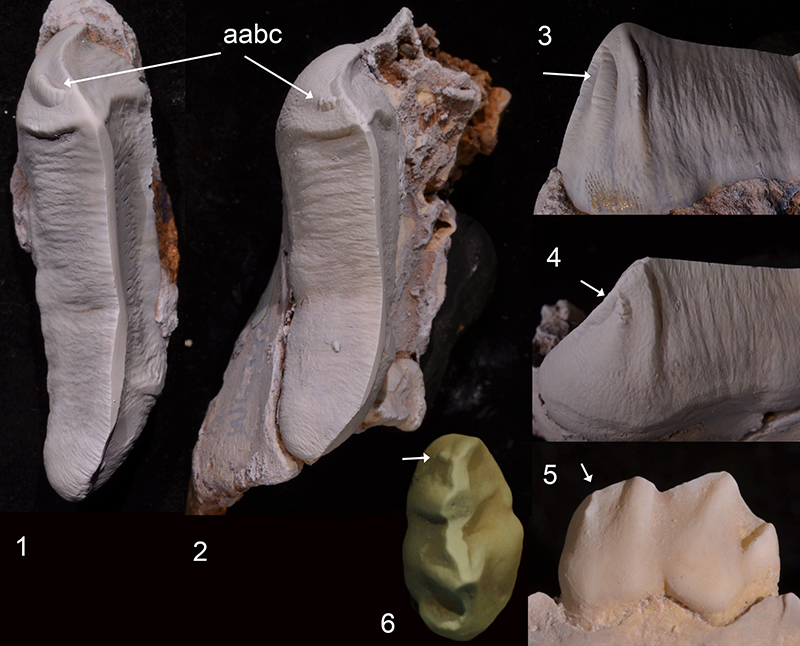
SUPPLEMENTARY FIGURE 2. Strict consensus tree of nine most-parsimonious trees obtained from the phylogenetic analysis. Tree length = 272 steps; CI = 0.580, RI = 0.750, RSI = 0.435. Unequivocal apomorphies for nodes (circled) are listed in Appendix 2; bootstrap support values for clades in the consensus tree are indicated in italics (50% majority rule).
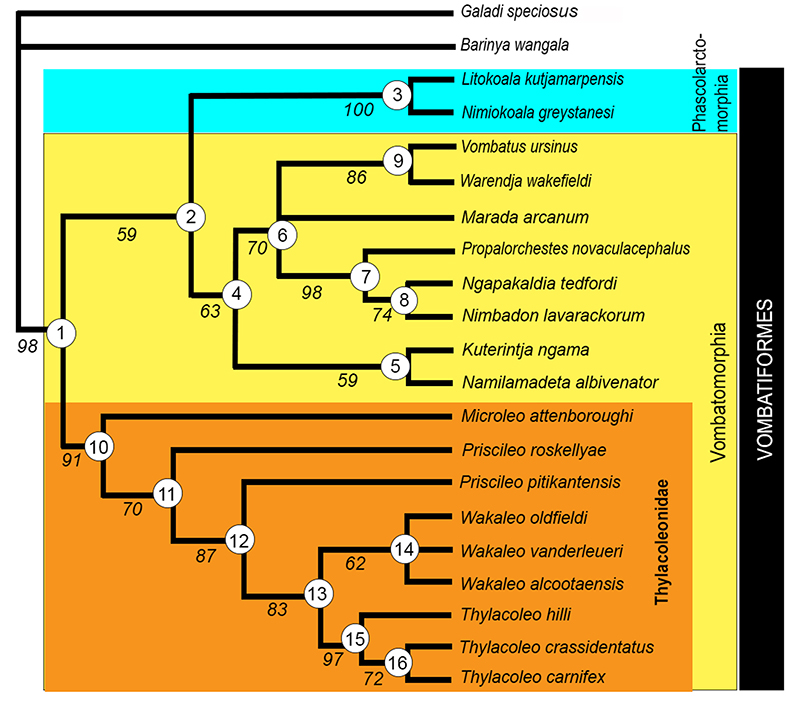
TABLE 1. Dental dimensions (mm) for Microleo attenboroughi QM F41143 and QM F42676. Abbreviations: l = length; w = width; a = alveolar measurement; [ ] = estimate. Measurements are of maximum dimensions.
| P3 | M1 | M2 | M3 | P3-M4 | m3 | ||||||
| Specimen # | l | w | l | w | l | w | l | w | l | l | w |
| QM F41143 | |||||||||||
| (left) | 4.7a | 2.0a | 3.5a | 2.9a | 3.3 | 3.6 | 2.8 | 3.0 | [16] | - | - |
| (right) | 4.6 | 2.7 | 3.6a | 2.8a | 3.3 | 3.6 | - | - | - | - | - |
| QM F42676 | - | - | - | - | - | - | - | - | - | 3.0 | 2.5 |
TABLE 2. Comparative measurements (mm) of P3 length of thylacoleonid species. Abbreviations: a = alveolus measure; ^ = P3/P3-M3; # =P3/P3+M1+M2; ^^ = P3/P3 +M1 (in Thylacoleo carnifex M1 lies anterior to the posterior end of P3 hence individual tooth lengths are summed for cheektooth row length); ** = measurements from Archer and Dawson (1982).
| Species | Specimen number | P3 length | P3:M1 | P3: cheektooth row length (P3-M4) |
| Microleo attenboroughi | QM F41143 | 4.6 | 1.2 | 0.29 |
| Priscileo roskellyae | QM F23453 | 8.2 | 1.4 | 0.35 |
| Priscileo pitikantensis | SAM P37719 | 11.2a | 1.8a | 0.36 |
| Wakaleo oldfieldi | QM F11852 | 14.1 | 1.3 | - |
| Wakaleo vanderleueri | CPC 26604 | 17.4 | 1.5 | 0.45^ |
| Wakaleo alcootaensis | NTM P1 | 23.3 | 1.6 | 0.49^ |
| Thylacoleo hilli | SAM P17621 | 24.4 | - | - |
| Thylacoleo crassidentatus | QM F10622** | 41.0 | 2.7 | 0.70# |
| Thylacoleo carnifex | AR 21609 | 55.4 | 3.9 | 0.78^^ |
APPENDIX 1
Phylogenetic Analysis of Microleo attenboroughi: Results and Description of Characters
The phylogenetic relationships of Microleo attenboroughi within the Thylacoleonidae were assessed as well as the relationships of thylacoleonids within the Suborder Vombatiformes, using a branch-and-bound search in PAUP * 4.0b10 (Swofford, 2002) based on a modified data matrix of Black et al. (2012) employing a total of 93 characters. Thirty characters were designated ordered. All characters were equally weighted and parsimony-informative. Five phascolarctid taxa in that matrix were not included in this analysis because many had high levels of missing data and the phascolarctids that were included were deemed informative for that taxon. The Oligo-Miocene peramelid Galadi speciosus and dasyurid Barinya wangala were used as outgroups. Within the thylacoleonid ingroup taxa, Priscileo pitikantensis, Wakaleo alcootaensis and Thylacoleo hilli have relatively high levels of missing data because they are only known from relatively incomplete specimens. The dentary specimen of Thylacoleo sp. cf. T. hilli (AM F63584) was used to code characters for the lower dentition of T. hilli on the basis of the assumption that it is likely to be representative of that species. Sixty five of the 71 characters of the original data matrix (Black et al., 2012) analysis were utilised. Six characters were removed because they either related to phascolarctid taxa not included in the analysis or were uninformative. One character relating to P3 morphology (ch.14) was modified to incorporate states in newly included taxa. A heuristic search was performed using branch and bound analysis. New characters relating to dental morphology (ch. 5, 6, 7, 8, 9, 10, 11, 12, 13, 15, 17 18, 27, 28, 30, 38, 39, 40, 41, 42, 43, 44, 48, 50, 53, 57, 58, 59) were included in the data matrix. Bootstrap values were calculated using 1000 replicates.
Results
Our phylogenetic analysis recovered nine most-parsimonious trees (see consensus tree in Supplementary Figure 2: tree length = 272 steps, consistency index = 0.580, rescaled consistency index = 0.435, retention index = 0.750) in all of which Microleo attenboroughi is recovered as the sister taxon to all other known thylacoleonid taxa. Unequivocal thylacoleonid apomorphies (node 10) include bunodont molars and features of P3, including its length relative to M1 length (> 1.2), a longitudinal blade that has a long v-shaped (notched) occlusal edge and lack of a basal posterior cingulum. Autapomorphies distinguishing Microleo from all other thylacoleonids are features of the P3; its fusiform shape, medial cuspule, a posterolingual crest and steep posterobuccal crest at the rear of the crown. Bootstrap support was relatively strong for the thylacoleonid clade (91%) and for the Priscileo - Wakaleo - Thylacoleo clade (70%; see Supplementary Figure 2). Synapomorphies for the Priscileo - Wakaleo - Thylacoleo clade are: a posterior longitudinal blade on P3 that is gently-angled and M2 with confluent postprotocrista and premetaconulecrista. Priscileo roskellyae is recovered as the sister taxon to P. pitikantensis which in turn is recovered as the sister taxon to a Wakaleo/Thylacoleo clade. Unequivocal synapomorphies of the Wakaleo - Thylacoleo clade are increased P3 length relative to cheektooth row (> 0.4) and loss of M4/m4. Bootstrap support for this clade was moderate (83%). Species of Wakaleo form a clade (bootstrap support 62%) but their relationships are unresolved probably as a result of the limited data for both W. oldfieldi and W. alcootaensis. Synapomorphies for the Wakaleo clade include; reduced number of upper and lower premolars, development of an anterolingual cuspule and a moderately-developed posterobuccal crest on P3. Monophyly for species of Thylacoleo was well-supported (bootstrap frequency = 97%) and based on synapomorphies of P3 including: width greater anteriorly than posteriorly, a convex posterior longitudinal blade, a posterolingual crest, a significant increase in length, and reduction of the number of lower incisors to one.
Vombatiform Relationships. The strict consensus tree (Supplementary Figure 2) places thylacoleonids as the sister taxon to all other vombatiform families. This phylogeny differs from previous phylogenies (e.g., Aplin and Archer, 1987; Marshall et al., 1990; Gillespie, 2007; Black et al., 2012) which place thylacoleonids within the vombatomorphian clade. In our analysis, synapomorphies that united phascolarctomorphians and vombatomorphian families (excluding thylacoleonids, node 2) include presence of a single upper and lower premolar, presence of a lingual cusp on P3, a rectangular M2, lower incisors reduced to one, mandibular symphysis that extends to below m1 and a diastema that is < 30% the length of the horizontal ramus. Bootstrap support for this clade was weak (59%). Black et al. (2012) found phascolarctids were united by characters of the lower molars and were the sister group to all other vombatiform families. Bootstrap support for the phascolarctomorphian clade was strong (100%) but was weaker (63%) for the vombatomorphian clade (excluding thylacoleonids). With the exception of thylacoleonids and Marada arcanum, the relationships of all the other vombatomorphian families agree with those presented by Black et al. (2012). Our analysis was unable to resolve the position of Marada with respect to vombatids, diprotodontids and palorchestids.In previous analyses (Aplin and Archer, 1987; Black et al., 2012) thylacoleonids have been united with other vombatomorphian families predominantly on the basis of cranial characters and their repositioning in this analysis may be due to the large number of dental characters in the data matrix. The results of this analysis suggest that thylacoleonids possibly form a clade within Vombatiformes that has similar taxonomic status to phascolarctomorphians. However, in contrast to thylacoleonids, phascolarctomorphians show a greater diversity at the generic level and, at present, stronger support for the clade is required before erecting a possible new thylacoleonid infraorder.
Character Description
Description of 93 characters used in the phylogenetic analysis. Reference is given to characters used in previous analyses and whether they are modified.* Indicates ordered characters. Abbreviations for references: A84b, Archer (1984b); AD82, Archer and Dawson (1982); B07, Black (2007); B12, Black et al. (2012); BA97a, Black and Archer (1997a); BA97b, Black and Archer (1997b); G07, Gillespie (2007); H93, Hand et al. (1993); HSV03, Horovitz and Sanchez-Villagra (2003); HW82, Hope and Wilkinson (1982); L02, Luo et al. (2002); L04, Louys (2004); M86, Murray (1986); M90, Marshall et al. (1990); M94, Myers (1994); MA97, Myers and Archer (1997); R87, Reig et al. (1987), R98, Rougier et al. (1998); S97, Springer et al. (1997); SVW02, Sánchez-Villagra and Wible, (2002); T10, Travouillon et al. (2010); WD 87, Woodburne et al. (1987); W98, Wroe et al. (1998); W00, Wroe et al. (2000).
| 1.* | Number of upper incisors: five (0); four (1); three (2); two (3); one (4). (S97, R98, W00, HSV03, B12) |
| 2. | Size of I1/ relative to I2-3/: equal/similar length (0); large, elongate (1). (B12) |
| 3. | Upper canine: present (0); absent/vestigial (1). (Modified: S97, HSV03, B12) |
| 4.* | Number of upper premolars: three (0); two (1); one (2). (B12, AD82, B12) |
| 5. | Number of primary cusps on longitudinal blade: one (0); two (1); three (2); four (3). |
| 6. | P3/ occlusal shape: wider posteriorly than anteriorly (0); anterior and posterior widths similar, broadest centrally (1); wider anteriorly than posteriorly (2). (G07) |
| 7.* | Absolute size of P3: small (< 5mm) (0); moderate (5-10mm); (1); large (10-20mm) (2); very large (20-30mm) (3); extremely large (>30mm) (4). |
| 8.* | Size of P/p3 relative to cheektooth row length: very short (0); short (1); long (2); very long (3). (G07) |
| 9. | Curvature of the longitudinal blade of P3: straight (0); buccally convex (1); lingually convex (2). |
| 10.* | Development of the anterolingual crest on P3: absent (0); weak, long crest (1); moderate, long crest (2); cuspule (3). (G07) |
| 11. | Slope of the posterior end of P3 longitudinal blade: steep (0); gently-angled (1); gently convex (2). (G07) |
| 12. | Long v-shaped longitudinal blade on P3: absent (0); present (1). |
| 13. | P3 Posterolingual crest: absent (0); present (1). |
| 14. | P3 Lingual cusp: absent (0); present (1). (Modified: MA97, B12) |
| 15. | P3 posterobuccal crest: absent (0); well developed (1); moderately developed (2); weakly developed (3). (G07) |
| 16. | P3 transverse parametacone crest: absent (0); present (1). (Modified: H93, BA97a, B12) |
| 17. | P3 medial cuspule: absent (0); present (1). |
| 18. | Posterior cingulum development on P3: buccal and lingual (0); lingual (1); absent (2). |
| 19.* | Size of P3/ relative to M1/: P3L/M1L ≤ 0.8 (0); 0.9 < P3L/M1L < 1.1 (1); 1.1< P3L/ M1L≤ 1.2 (2); 1.2 < P3L/M1L≤ 1.4 (3); 1.4< P3L/ M1L≤ 1.5 (4); 1.5 < P3L/ M1L≤ 1.6 (5) P3L/ M1L > 1.6 (6). (Modified: BA97a, B12) |
| 20.* | Molar morphology: tribosphenic (0); selenodont (1); semi-lophodont (2); lophodont but stylar cusps evident on lophs (3); fully lophodont (4); bunodont (5). (Modified: B12) |
| 21. | Anteriorly concave lower/ anteriorly convex upper molar lophs: absent (0); present (1). (B12) |
| 22. | Ever-growing unrooted cheekteeth: absent (0); present (1). (B12) |
| 23. | Enamel crenulations: small (0); strong, crest-like (1). (Modified: S97, B12) |
| 24.* | Stylar cusp development: strong (0); moderate (1); weak/absent (2). (Modified: BA97b, B12) |
| 25.* | Parastyle development on M1: absent/small (0); moderately developed with expansion of anterobuccal tooth corner (1); large, cuspate and pyramid-like (2). (BA97b, B12) |
| 26. | Separation of stylar cusps C and D: close together (0); separated by large trough (1). (MA97, B12) |
| 27.* | M1 occlusal outline: triangular (0); subtriangular/subsquare (1); square (2); rectangular (3). (G07) |
| 28. | M1 metacone: moderate (0); large (1). |
| 29.* | M1 paraconule: absent/weak (0); moderate/strong, linear (1); strong, crescentic (2). (Modified: BA9, B12) |
| 30. | M1 anteroposterior buccal gradient: paracone much shorter than metacone (0); paracone slightly shorter than metacone (1); paracone and metacone similar height (2); paracone slightly taller than metacone (3); paracone much taller than metacone (4). (G07) |
| 31.* | M1 neometaconule: absent (0); weak/small (1); moderate/well developed (2). (Modified: WD87, BA97b, S97) |
| 32. | Protostyle: absent (0); present (1). (WD87, BA97b, B12) |
| 33. | Paracone and metacone placement: medial (0); buccal (1). (S97, HSV03, B12) |
| 34. | Paracone buccal basin on M1 deep, enclosed: absent (0); present (1). (B12) |
| 35. | Posterolingual paracristae: absent/weak (0); strongly developed (1). (BA97b, B12) |
| 36. | Postprotocrista: present (0); absent (1). (A84b, B12) |
| 37.* | M2 occlusal shape: triangular (0); subtriangular (1); rectangular i.e., longer than wide (2); square i.e., width ≥ length (3). (Modified: S97, HSV03, B12) |
| 38.* | M2 buccal height vs lingual height: similar (0); slightly taller (1); moderately taller (2); much taller (3). (G07) |
| 39.* | M2 buccal inflation of crown below paracone: absent (0); present (1). (G07) |
| 40.* | M2 postprotocrista and premetaconule crista: confluent (0); not confluent, separated (1). |
| 41. | M2 postparacrista direction: posterobuccal (0); posterior (1). |
| 42. | M2 width of trigon basin relative to crown width: broad (0); narrow (1). |
| 43. | m3: present (0); absent (1). (AD82, G07) |
| 44. | M/m4: present (0); absent (1). (AD82, G07) |
| 45. | M4 metaconule: absent or significantly reduced and retracted towards posterior cingulum (0); distinct, cuspate (1). (B12) |
| 46.* | Number of lower incisors: three (0); two (1); one (2). (S97, R98, W00, L02, HSV03, B12) |
| 47. | Inclination angle of i1: high, ≥ 30 degrees (0); low < 30 degrees (1). (M90, B07, B12) |
| 48. | Number of lower premolars: three (0); two (1); one (2). |
| 49. | p3 morphology: bicuspid, simple (0); multicusped/bladed (1). (B07, B12) |
| 50. | p3 anterolingual crest: well-developed (0); weakly-developed: absent (1); absent (2). |
| 51. | Posterior cingulum on p3: absent/weak (0); present (1). (M94, B12) |
| 52. | Molar gradient: ratio m4/m1 < 1 (0); m4/m1 ≥ 1 (1). (B07, B12) |
| 53. | m2 talonid height relative to paraconid and trigonid: slightly shorter (0); similar (1); much shorter (2). |
| 54. | Development of paraconid and paracristid on m1: paraconid present and paracristid well-developed (0); paraconid weak or absent, paracristid present but low (1); paraconid and paracristid absent (2). (B0, B12) |
| 55. | Position of protoconid on m1: buccal half of trigonid (0); lingual third of trigonid (1). (BA97b, B12) |
| 56.* | Protostylid on m1: absent (0); small (1); moderate (2); large (3). (WD87, S97, BA97b, MA97, B0, B12) |
| 57.* | m1 talonid basin width between entocristid and hypocristid: broad (0); slightly narrowed (1); narrow (2). (G07) |
| 58.* | m1 talonid width relative to trigonid width: broader (0); slightly narrower (1); much narrower (2). (G07) |
| 59.* | m3 talonid basin: broad (0); narrow (1); absent/lost (2). |
| 60.* | Metastylid development: absent (0); present, cuspate (1); present, metastylid fold (2). (Modified: BA97b, B12) |
| 61. | Entostylid ridge on m1: absent (0); present (1). (BA97b, B12) |
| 62. | Cristid obliqua: well-developed, does not meet postprotocristid (0); well-developed, meets postprotocristid lingual to horizontal tooth midline (1); well-developed, meets postprotocristid at or buccal to horizontal tooth midline (2); weak/absent (3). (B12) |
| 63. | Internal ribs on conids of lower molars: absent (0); present, meet in longitudinal valley (1); present, do not meet (2). (B12) |
| 64.* | Nasal aperture retracted beyond incisor arcade: absent (0); retracted to above diastema (1); retracted to above cheek tooth row (2). (B12) |
| 65.* | Masseteric process: absent/weak- dorsal to molar row (0); at level of molar row (1); elongate- extends ventral to molar row (2). (W98, B12) |
| 66. | Lacrimal tuberosity: absent (0); present (1). (R98; HSV03, B12) |
| 67. | Infraorbital shelf: well-developed (0); weak (1). (B12) |
| 68.* | Posterior palatal vacuities: anteriorly extensive to opposite or anterior to M1 (0); extends anteriorly to opposite M2 (1); confined within palatine, opposite M3-4 (2); absent (3). (A84b, R87, R98, W00, HSV03, B12) |
| 69. | Frontal/squamosal contact: absent, alisphenoid-parietal contact (0); present (1). (S97, W98, HSV03, B12) |
| 70. | Infratemporal crests: weak/absent (0); well-developed (1). (B12) |
| 71. | Postglenoid constriction: absent (0); present (1). (B12) |
| 72. | Glenoid fossa: flat articular eminence, shallow mandibular fossa (0); flat articular eminence, deep mandibular fossa (1); prominent articular eminence, mandibular fossa absent (2). (B12) |
| 73.* | Postglenoid process: elongate (0); short (1); absent (2). (Modified: S97, HSV03, B12) |
| 74. | Medial glenoid process: absent (0); present (1). (M86, B12) |
| 75. | Well-developed postglenoid cavity: absent (0); present (1). (L04, B12) |
| 76. | Position of postglenoid foramen: posterior to PGP and bounded medially by petrosal (0); anteromedial to or in line with postglenoid process (1); posteromedial to postglenoid process within squamosal (2); within epitympanic fenestra, surrounded by bony septum (3). (Modified: S97, W98, HSV03, B12) |
| 77. | Tympanic cavity roof elements: alisphenoid and petrosal (0); alisphenoid and squamosal (1); squamosal (2). (Modified: T10, B12) |
| 78. | Tympanic floor elements: alisphenoid (0); alisphenoid and squamosal (1); squamosal (2). (B12) |
| 79.* | Alisphenoid tympanic wing: absent (0); short (1); moderate, extends under periotic (2); elongate, completely floors middle ear (3). (S97, W98, HSV03, B12) |
| 80. | Epitympanic fenestra: absent (0); present (1). (B12) |
| 81. | Non auditory sinuses: absent (0); present (1). (M86, B12) |
| 82.* | Posterior epitympanic fossa: absent (0); present, shallow (1); deep, perforating squamosal and mastoid (2) (M86, B12) |
| 83. | Rostral tympanic process of periotic: strong (0); absent/weak (1). (R98, SVW02, HSV03, B12) |
| 84. | Posterior parietal width: broad (0); narrow (1). (B12) |
| 85. | Interparietal: present (0); absent (1). (L04, B12) |
| 86. | Narrow mastoid strip on occiput: absent (0); present (1). (M86, B12) |
| 87. | Ventrolaterally flared mastoid process on occiput: absent (0); present (1). (M86, B12) |
| 88. | Angle of the anterior border of the ascending ramus: < 70 degrees (0); ≥ 70 degrees (1). (HW82, B07, B12) |
| 89.* | Posterior extent of mandibular symphysis: anterior to p3 (0); below p3 (1); below m1 (2); below m2-3 (3). (B07, B12) |
| 90.* | Diastema (between i1 and p3): absent (0); present, length < 30% of horizontal ramus length (1); present, length > 30% of horizontal ramus length (2). (B07, B12) |
| 91. | Tooth row length (p3-m4) relative to horizontal ramus length: > 60% (0); ≤ 60% (1). (B12) |
| 92. | Masseteric foramen: absent (0); present (1). (B07, B12) |
| 93. | Flared masseteric eminences: absent/weak (0); moderately to strongly flared (1). (M98, B12) |
APPENDIX 2
Data Matrix
Data matrix used for phylogenetic analysis of Vombatiformes. Abbreviations: '?' signifies 'missing data'; '-' signifies 'inapplicable'. Polymorphic states indicated by: A, (0,1).
| 1 0 |
2 0 |
3 0 |
4 0 |
5 0 |
6 0 |
7 0 |
|
| Barinya wangala | 1000000000 | 0000000010 | 00-00-0100 | 0000000101 | 010000?002 | 011000000- | -20100?110 |
| Galadi speciosus | 0000000000 | 0000000000 | -0-0001000 | 00000012-0 | 010000?002 | 0000000000 | 0200000000 |
| Litokoala kutjamarpensis | ??? 2201012 | 0011001101 | -012202121 | 21011032-1 | 00000???1? | 00?0130002 | 112?1?0100 |
| Nimiokoala greystanesi | 2002200011 | 2002200011 | -012202121 | 21011031-1 | 0000020212 | 0000130001 | 1120000?00 |
| Vombatus ursinus | 4 -121010-0 | -000000- 02 | 01-2003102 | 00100020-- | - -00121202 | 00210000-0 | 02-1010211 |
| Warendja wakefieldi | 4 -12- 000-0 | - 000000212 | 01-2003- 02 | 00100020-- | - -00121202 | 00210000-0 | 02-?00020? |
| Kuterintja ngama | ??? 22010 -3 | 0001000011 | 0000112101 | 00010021-1 | 0000121212 | 1021000000 | 01-?2?0??? |
| Namilamadeta albivenator | 2002302001 | 0011001022 | 00-0112102 | 00000021-0 | 0000121210 | 1021000000 | 02-1210111 |
| Marada arcanum | ???2?0?0?? | ?????????2 | 00-??????2 | ??????? ??? | ??00?21202 | 1001000000 | 02-??????? |
| Propalorchestes novaculacephalus | ??12022?20 | 0011000003 | 10-1013102 | 0010012001 | - -00121?0? | 1102000000 | 02-2??1301 |
| Ngapakaldia tedfordi | 2112001000 | 0011010004 | 10-2013102 | 0010012001 | 1-00121202 | 1102000000 | 03-1111311 |
| Nimbadon lavarackorum | 2112002000 | 0011010014 | 10-2013102 | 0010012001 | --0012120- | 1101000000 | 03-0211311 |
| Microleo attenboroughi | ??? 0110111 | 0110101235 | - 002002??? | ?0?? 001001 | 0000 ?????? | ???????? 0? | ?? 0??????? |
| Priscileo roskellyae | 2100101122 | 1100300235 | -002001003 | 0000001000 | 0000?10000 | 0011100000 | 02- 0000001 |
| Priscileo pitikantensis | ? 100? 021?? | ????????? 5 | -00 ????? ?? | ????0002? 0 | ?100?????? | ?????? ?? ?? | ?????? ?? ?? |
| Wakaleo oldfieldi | 210A102223 | 1100200235 | -002000103 | 0000002110 | 1101-10201 | 0011102110 | 020 ??????? |
| Wakaleo vanderleueri | 210A102223 | 1100200235 | -002000104 | 0000003110 | 1101-10202 | 0011102120 | 0200000101 |
| Wakaleo alcootaensis | ?10 2103223 | 1100200235 | -002?00??? | ?????? ?? ?? | ??11-??2?? | ??????2?-? | ??0??????? |
| Thylacoleo hilli | ???? 123? 21 | 21100002 ?? | -0???????? | ?????? ?? ?? | ?????20000 | 0???0????? | ?????? ???? |
| Thylacoleo crassidentatus | 2100124321 | 2110000245 | -002100104 | 000000???? | ?111-20?00 | 00111022-0 | 020????0?? |
| Thylacoleo carnifex | 2100124321 | 2110000245 | -002103104 | 000000???? | ??11-20001 | 00111022 -0 | 0200010011 |
| 8 0 |
9 0 |
9 3 |
|||||
| Barinya wangala | 0100000020 | 00000?0000 | 100 | ||||
| Galadi speciosus | 0100000020 | 0000100?00 | 100 | ||||
| Litokoala kutjamarpensis | 0100111020 | 0000000??? | ??? | ||||
| Nimiokoala greystanesi | ????1?1??0 | 0?000??021 | 010 | ||||
| Vombatus ursinus | 1221122200 | 0110111132 | 111 | ||||
| Warendja wakefieldi | ???11?2200 | 0??01??022 | 111 | ||||
| Kuterintja ngama | ????? ?? ?? | ?????? ?12? | ??? | ||||
| Namilamadeta albivenator | 1001122120 | 0110111021 | 010 | ||||
| Marada arcanum | ?????? ?? ?? | ?????? ?012 | 110 | ||||
| Propalorchestes novaculacephal | 11111?21?1 | 1211111122 | 111 | ||||
| Ngapakaldia tedfordi | 1111132201 | 1211110122 | 110 | ||||
| Nimbadon lavarackorum | 1111132111 | 1211110121 | 1A0 | ||||
| Microleo attenboroughi | ?????? ?? ?? | ?????? ?? ?? | ??? | ||||
| Priscileo roskellyae | 0001122120 | 0010100?10 | 010 | ||||
| Priscileo pitikantensis | ?????? ?? ?? | ?????? ?? ?? | ??? | ||||
| Wakaleo oldfieldi | ?????? ?? ?? | ?????? ?010 | 010 | ||||
| Wakaleo vanderleueri | 0001122120 | 0001110010 | 010 | ||||
| Wakaleo alcootaensis | ?????? ?? ?? | ?????? ?? 1? | ??? | ||||
| Thylacoleo hilli | ?????? ?? ?? | ?????? ?? ?0 | ??? | ||||
| Thylacoleo crassidentatus | ?????? ?? ?? | ?????? ?? 10 | 010 | ||||
| Thylacoleo carnifex | 0001022200 | 0011110010 | 010 |
List of Synapomorphies
List of nodal apomorphies resulting from the phylogenetic analysis. Node numbers refer to Supplementary Figure 2.
| Node | Character | Change |
| 1 | 1 | 1→2 |
| 10 | 0→1 | |
| 20 | 0→2 | |
| 24 | 0→2 | |
| 42 | 1→0 | |
| 46 | 0→1 | |
| 73 | 0→1 | |
| 75 | 0→1 | |
| 76 | 0→2 | |
| 77 | 0→2 | |
| 89 | 0→1 | |
| 91 | 1→0 | |
| 92 | 0→1 | |
| 2 | 4 | 0→2 |
| 14 | 0→1 | |
| 37 | 1→2 | |
| 46 | 1→2 | |
| 48 | 0→2 | |
| 89 | 1→2 | |
| 90 | 0→1 | |
| 3 | 17 | 0→1 |
| 18 | 0→1 | |
| 23 | 0→1 | |
| 25 | 1→2 | |
| 29 | 0→2 | |
| 31 | 0→2 | |
| 32 | 0→1 | |
| 34 | 0→1 | |
| 35 | 0→1 | |
| 37 | 2→3 | |
| 56 | 0→3 | |
| 60 | 0→1 | |
| 61 | 0→1 | |
| 62 | 2→1 | |
| 77 | 2→1 | |
| 85 | 1→0 | |
| 4 | 45 | 0→1 |
| 47 | 0→1 | |
| 51 | 0→1 | |
| 64 | 0→1 | |
| 66 | 0→1 | |
| 71 | 0→1 | |
| 82 | 0→1 | |
| 86 | 0→1 | |
| 87 | 0→1 | |
| 5 | 24 | 2→0 |
| 53 | 0→2 | |
| 65 | 1→2 | |
| 6 | 3 | 0→1 |
| 10 | 1→0 | |
| 27 | 2→3 | |
| 33 | 0→1 | |
| 38 | 1→0 | |
| 68 | 1→2 | |
| 73 | 0→1 | |
| 79 | 2→0 | |
| 90 | 1→2 | |
| 91 | 0→1 | |
| 7 | 13 | 0→1 |
| 20 | 2→3 | |
| 21 | 0→1 | |
| 36 | 0→1 | |
| 52 | 0→1 | |
| 67 | 0→1 | |
| 68 | 2→3 | |
| 80 | 0→1 | |
| 81 | 0→1 | |
| 82 | 1→2 | |
| 84 | 0→1 | |
| 8 | 16 | 0→1 |
| 20 | 3→4 | |
| 62 | 2→3 | |
| 9 | 1 | 2→4 |
| 14 | 1→0 | |
| 22 | 0→1 | |
| 51 | 1→0 | |
| 53 | 0→2 | |
| 78 | 1→2 | |
| 93 | 0→1 | |
| 10 | 12 | 0→1 |
| 18 | 0→2 | |
| 19 | 0→3 | |
| 20 | 2→5 | |
| 11 | 11 | 0→1 |
| 40 | 1→0 | |
| 12 | 7 | 1→2 |
| 37 | 1→0 | |
| 38 | 0→2 | |
| 42 | 0→1 | |
| 13 | 8 | 1→2 |
| 44 | 0→1 | |
| 14 | 4 | 0→2 |
| 10 | 2→3 | |
| 15 | 0→2 | |
| 48 | 0→2 | |
| 15 | 6 | 0→2 |
| 7 | 2→3 | |
| 11 | 1→2 | |
| 13 | 0→1 | |
| 46 | 1→2 | |
| 16 | 7 | 3→4 |
APPENDIX 3
Body Mass Estimate
An estimate of the body mass of Microleo attenboroughi, Priscileo roskellyae and Wakaleo sp. nov. was made using a regression equation formulated by Myers (2001) from correlations of marsupial body mass with cranio-dental measurements. The equation used for the analysis was the highest possible ranked regression from the ‘Diprotodontians data set’ of Myers (2001, table 5, p.106) and employed the measurement for upper molar row length (UMRL, i.e., M1-M4). This equation (log y = -0.418 + 3.011 [log x], where x equals UMRL) was chosen because it was based on alveolar measurements rather than molar measurements; the posterior teeth are missing in both fossil taxa. The UMRL measurements (average of the left and right lengths) for M. attenboroughi (11.3 mm) P. roskellyae (QM F23453, 16.4 mm) and Wakaleo sp. nov. (QM F45200, 23.3 mm) resulted in body weight estimates of 590 g, 1813 g, and 5221 g, respectively. As recommended by Myers (2001), a smearing estimate (4.4%) was applied to the equation. Myers (2001) indicates that the regression equations are not appropriate for the larger species of marsupial lions (based on their unusual reduced molar numbers that result in large underestimates of body mass), hence, an estimate was not calculated for Wakaleo vanderleueri.
A tiny new marsupial lion (Marsupialia, Thylacoleonidae) from the early Miocene of Australia
Plain Language Abstract
Marsupial lions (Thylacoleonidae) are an extinct family of marsupials that are found in late Oligocene to Pleistocene deposits of Australia. Previously, this family consisted of three genera and eight species, ranging from domestic cat to lion in size. The paper describes a new genus and species of marsupial lion, Microleo attenboroughi, recovered from early Miocene freshwater limestones of the Riversleigh World Heritage Area of Australia. This possum-sized new species is strikingly diminutive compared to other marsupial lions. It exhibits the elongated, trenchant, upper third premolar (P3) and subtriangular, bunodont molars that are characteristic of the family, but other features of its P3 are distinctively different. Analysis of its relationships suggest that it is the most primitive member of its family and on a broader scale, and in contrast to previous hypotheses, that marsupial lions are the sister-taxon to a diverse yet entirely herbivorous group that contains all wombat-like marsupials and koalas.
Resumen en Español
Un nuevo pequeño león marsupial (Marsupialia, Thylacoleonidae) del Mioceno temprano de Australia
Se describe a Microleo attenboroughi, un nuevo género y especie de un diminuto león marsupial (Marsupialia: Thylacoleonidae) proveniente de calizas dulceacuícolas del Mioceno temprano del Área Patrimonio de la Humanidad Riversleigh, noroeste de Queensland, Australia. Un paladar roto que retiene filas de postcaninos incompletas demuestra que este nuevo león marsupial muy pequeño poseía el P3 agudo y elongado y molares superiores predominantemente subtriangulares característicos de los thilacoleónidos, mientras que otras características del premolar apoyan su inclusión en un género nuevo. Los análisis filogenéticos sugieren que Microleo attenboroughi es el taxón hermano de todos los otros thilacoleónidos, y que Thylacoleonidae podría situarse por fuera de Vombatomorphia como el taxón hermano de todos los marsupiales vombátidos, incluyendo koalas. Sin embargo, dada la información limitada sobre la morfología craneal de M. attenboroughi, conservadoramente se concluye que Thylacoleonidae es parte del clado vombatomorpho. Este nuevo thilacoleónido aumenta a tres el número de especies de leones marsupiales que se han recuperado de los depósitos del Mioceno temprano de Riversleigh e indica un nivel de diversidad no observado previamente para este grupo. Es probable que el tamaño y morfología diferentes de los tres taxones simpátricos refleje una partición de nicho y una consecuente reducción en la competencia. Los thilacoleónidos pueden haber sido los predadores arbóreos dominantes del Cenozoico de Australia.
Palabras clave: Thylacoleonidae; león marsupial; género nuevo; especie nueva; Mioceno temprano; Riversleigh
Traducción: Diana Elizabeth Fernández
Résumé en Français
Un nouveau lion marsupial (Marsupialia, Thylacoleonidae) de très petite taille du Miocène ancien d'Australie
Microleo attenboroughi gen. nov., sp. nov., un lion marsupial (Marsupialia : Thylacoleonidae) de très petite taille, est décrit à partir de restes provenant de calcaires d'eau douce du Miocène ancien de la "Riversleigh World Heritage Area", nord-ouest du Queensland, Australie. Un palais cassé préservant les rangées dentaires jugales incomplètes démontre que ce nouveau lion marsupial de très petite taille possède la P3 allongée et tranchante et les molaires supérieures majoritairement subtriangulaires caractéristiques des thylacoléonidés, alors que d'autres caractères de la P3 soutiennent son placement dans un nouveau genre. Une analyse phylogénétique suggère que Microleo attenboroughi gen. nov., sp. nov., est le groupe-frère de tous les autres thylacoléonidés, et que Thylacoleonidae pourrait être placé à l'extérieur de Vombatomorphia comme groupe-frère de tous les autres marsupiaux ressemblant à des wombats, dont les koalas. Cependant, étant donné les données limitées sur la morphologie crânienne de M. attenboroughi gen. nov., sp. nov., il est conclu provisoirement ici que Thylacoleonidae fait partie du clade Vombatomorphia. Ce nouveau thylacoléonidé porte à trois le nombre d'espèces de lions marsupiaux qui ont été trouvées dans les dépôts du Miocène ancien à Riversleigh, et indique un niveau de diversité non observé jusqu'à présent dans ce groupe. Il est probable que les différences de taille et de morphologie de ces trois taxons sympatriques reflètent un partage des niches, et donc une compétition réduite. Les thylacoléonidés étaient peut-être les prédateurs arboricoles dominants en Australie pendant le Cénozoïque.
Mots-clés : Thylacoleonidae ; lion marsupial ; nouveau genre ; nouvelle espèce ; Miocène ancien ; Riversleigh
Translator: Antoine Souron
Deutsche Zusammenfassung
Ein kleiner neuer Beutellöwe (Marsupialia, Thylacoleonidae) aus dem frühen Miozän von Australien
Microleo attenboroughi, eine neue Gattung und Art kleiner Beutellöwen (Marsupialia: Thylacoleonidae), wird aus den frühmiozänen Frischwasser-Kalken des Riversleigh Welterbestätten Gebietes (nordwestliches Queensland, Australien) beschrieben. Ein zerbrochener Gaumen mit lückenhaften Backenzahnreihen zeigt, dass dieser neue, sehr kleine Beutellöwe die länglichen, scharfen P3 und die überwiegend subtriangularen oberen Molaren, die für die Thylacoleoniden typisch waren, besaß, während andere Merkmale des Premolaren die Einordnung in einer neuen Gattung unterstützen. Phylogenetische Analyse legt nahe, dass Microleo attenboroughi das Schwestertaxon zu allen anderen Thylacoleoniden ist, und dass die Thylacoleonidae möglicherweise außerhalb der Vombatomorphia als das Schwestertaxon zu allen anderen Wombat-ähnlichen Beuteltieren inklusive der Koalas liegen. Wegen der begrenzten Datenmenge zur Schädelanatomie von M. attenboroughi, werden die Thylacoleonidae hier konservativ miteingeschlossen als Teil der vombatomorphen Klade.
Dieser neue Thylacoleonide erhöht die Anzahl der Beutellöwen auf drei Arten aus den frühmiozänen Ablagerungen von Riversleigh und weist auf ein Diversitätslevel hin, wie man es für diese Gruppe bis jetzt nicht angenommen hatte. Es ist wahrscheinlich, dass die unterschiedliche Größe und Morphologie der drei sympatrischen Taxa Nischenaufteilung abbildet und damit verringerte Konkurrenz. Thylacoleoniden waren möglicherweise die vorherrschenden baumbewohnenden Raubtiere des känozoischen Australien.
Schlüsselwörter: Thylacoleonidae; Beutellöwe; neue Gattung; neue Art; frühes Miozän; Riversleigh
Translator: Eva Gebauer
Arabic

Translator: Ashraf M.T. Elewa
-
-
-
Review: The Princeton Field Guide to Mesozoic Sea Reptiles
 The Princeton Field Guide to Mesozoic Sea Reptiles
The Princeton Field Guide to Mesozoic Sea ReptilesArticle number: 26.1.1R
April 2023

 Poster Winners 2024
Poster Winners 2024