

 The Sumidero Quarry, an Albian fossil fish site in San Cristóbal de las Casas, Chiapas, southeastern Mexico
The Sumidero Quarry, an Albian fossil fish site in San Cristóbal de las Casas, Chiapas, southeastern Mexico
Article number: 28.2.a23
https://doi.org/10.26879/1437
Copyright Palaeontological Association, June 2025
Author biographies
Plain-language and multi-lingual abstracts
PDF version
Submission: 9 August 2024. Acceptance: 16 May 2025
ABSTRACT
The Sumidero Quarry is a new Albian paleontological site of the Cantelhá Formation at the escarped southwestern edge of the San Cristóbal de las Casas town, State of Chiapas, southeastern Mexico. The fossil assemblage recovered at this site includes ray-finned fishes as the predominant element, along with mollusk shells, plants, microfossils, and ichnofossils. To date, no cartilaginous fish remains have been recovered at this site. The fishes identified here include Teoichthys brevipina, Notagogus novomundi, indeterminate specimens of the family Pycnodontidae, and the genera Belenostomus and Thorectichthys. The Sumidero Quarry is the second fossil locality preserving shallow marine fishes of Albian age (113—100.5 Ma) found in Mexico and one of the few in the Americas. Comparisons of the Sumidero ichthyofauna to those of coeval and geographically nearby sites yield contrasting results. The Albian assemblage of the Sumidero shares some genera with that of the middle-late Albian from the Tlayúa Quarry, but it is entirely different from the Muhi Quarry assemblage of the Albian-Cenomanian boundary. Assessing the taxonomic diversity of the Sumidero fossils complements our current understanding of Albian marine fishes within North America and their affinities to other coeval deposits globally, allowing a more complete picture of marine fish diversity trends across the Albian-Cenomanian boundary.
Jesús Alvarado-Ortega. Instituto de Geología, Universidad Nacional Autónoma de México, Circuito de la Investigación S/N, Ciudad Universitaria, Delegación Coyoacán, D.F., 04510 México. Correspondence author. alvarado@geologia.unam.mx
Stephanie Pacheco-Ordaz. Posgrado en Ciencias Químicobiológicas, Escuela Nacional de Ciencias Biológicas, Instituto Politécnico Nacional, Unidad Profesional Lázaro Cárdenas. Prolongación de Carpio y Plan de Ayala S/N, Col. Santo Tomás, Alcaldía Miguel Hidalgo, Ciudad de México, 11340 México. spachecoo2200@alumno.ipn.mx, tephy.pacheco@gmail.com
Bruno Andrés Than-Marchese. Programa de Doctorado en Ciencias en Biodiversidad y Conservación de Ecosistemas Tropicales, Instituto de Ciencias Biológicas, Universidad de Ciencias y Artes de Chiapas, Libramiento Norte No. 1150, Colonia Lajas Maciel, Tuxtla Gutiérrez, Chiapas, 29039, México. bruthmar@gmail.com
Nayeli Lupercio-Espericueta. Posgrado en Ciencias de la Tierra, Instituto de Geología, Universidad Nacional Autónoma de México, Circuito de la Investigación S/N, Ciudad Universitaria, Delegación Coyoacán, Ciudad de México, 04510 México. nayelupercio@gmail.com
Braulio Renato de Jesus Capistrán-Romero. Posgrado en Biociencias, Escuela Nacional de Ciencias Biológicas, Instituto Politécnico Nacional, Unidad Profesional Lázaro Cárdenas. Prolongación de Carpio y Plan de Ayala S/N, Col. Santo Tomás, Alcaldía Miguel Hidalgo, Ciudad de México, 11340 México. renatodetauro@gmail.com
Jesús Alberto Díaz-Cruz. Colección Nacional de Peces, Departamento de Zoología, Instituto de Biología, Universidad Autónoma de México, Tercer Circuito Exterior S/N. CP 045110, México City. México. vertebrata.j@gmail.com
Keywords. New paleontological site; Osteichthyes; Albian-Cenomanian Boundary; Diversity; Cantelhá Formation
Final citation: Alvarado-Ortega, Jesús, Pacheco-Ordaz, Stephanie, Than-Marchese, Bruno Andrés, Lupercio-Espericueta, Nayeli, Renato de Jesus Capistrán-Romero, Braulio, and Díaz-Cruz, Jesús Alberto. 2025. The Sumidero Quarry, an Albian fossil fish site in San Cristóbal de las Casas, Chiapas, southeastern Mexico. Palaeontologia Electronica, 28(2):a23.
https://doi.org/10.26879/1437
palaeo-electronica.org/content/2025/5541-sumidero-quarry
Copyright: June 2025 Palaeontological Association
This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
creativecommons.org/licenses/by/4.0
INTRODUCTION
In 2008, paleontologists of the Instituto de Geología of the Universidad Nacional Autónoma de México (Igl, UNAM) and archaeologists of the Instituto Nacional de Antropología e Historia (INAH) launched a joint research project to identify the geological source of the fossil fishes recovered within the Mayan archeological site of Palenque (Cuevas-García and Alvarado-Ortega, 2009). Since then, this team has explored different sedimentary sequences within Chiapas, southeastern Mexico. So far, the achievements have been extraordinary; we discovered numerous fossiliferous localities, including the geological origin of the fossils that the ancient Maya incorporated into their ritual acts, offerings, and architectural structures, as well as numerous new fossil fish species (e.g., Alvarado-Ortega et al., 2015, 2018, 2020; Cantalice and Alvarado-Ortega, 2016; Cantalice et al., 2018, 2020).
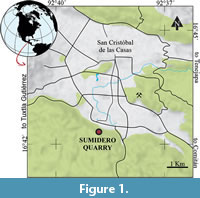 The Sumidero Quarry (Figure 1, Figure 2) is one of the fossiliferous sites identified in the aforementioned project. This site is a small outcrop of laminated marly limestones on the outskirts of the San Cristóbal de las Casas town, Chiapas, discovered about 20 years ago and intermittently exploited by the local people. The fossil assemblage found in this site includes marine organisms, mainly bony fishes and invertebrates. Here, such fossils overlie the interstratification planes of the extractable limestones of the small ledges or steps present along the gorge at the southeastern limit of San Cristóbal de las Casas, on the northern edge of the rugged hills known as the El Gavilán. We report the discovery of this new site, providing a general description of its geology and an overview of its ichthyofauna, which includes Ophipsiformes, Macrosemiformes, Pycnodontiformes, Aspidorhynchiformes, and Ellimmichthyiformes.
The Sumidero Quarry (Figure 1, Figure 2) is one of the fossiliferous sites identified in the aforementioned project. This site is a small outcrop of laminated marly limestones on the outskirts of the San Cristóbal de las Casas town, Chiapas, discovered about 20 years ago and intermittently exploited by the local people. The fossil assemblage found in this site includes marine organisms, mainly bony fishes and invertebrates. Here, such fossils overlie the interstratification planes of the extractable limestones of the small ledges or steps present along the gorge at the southeastern limit of San Cristóbal de las Casas, on the northern edge of the rugged hills known as the El Gavilán. We report the discovery of this new site, providing a general description of its geology and an overview of its ichthyofauna, which includes Ophipsiformes, Macrosemiformes, Pycnodontiformes, Aspidorhynchiformes, and Ellimmichthyiformes.
 The Sumidero Quarry is on the southwestern border of the town of San Cristóbal de las Casas, Chiapas, southeastern Mexico, at the coordinates 16°42'33.90" N and 92°39'9.60" W (Figure 1). The limestone sequence exposed in this small commercial quarry belongs to the Sierra Madre Group. Here, the locals often extract beige-to-brown slabs of thin, laminar, and parallel marly limestones used to make floors or wall coverings. Massive yellowish limestone strata bound these thin, marly layers. The Sierra Madre Group is a Cretaceous (Aptian-Santonian) marine carbonate succession of at least 2575 m thickness, present in the central and western parts of the State of Chiapas, as well as in the north of Guatemala and the east of Belize (Steele, 1986; Waite, 1986). According to Salas (1949), in 1929, Nuttall W.L.F. identified this sequence in an unpublished work about the stratigraphy of southeastern Mexico, which he named the Sierra Madre Limestones. Subsequently, these were recognized as a tripartite Group involving the Albian-Cenomanian Cantelhá Formation plus the Turonian-Santonian Jolpabuchil and Cintalapa Formations (see Quezada-Muñetón, 1987). Steele (1986) and Waite (1986) studied the Sierra Madre limestones through 21 and 11 levels (named by them as units), respectively, concluding that units 1-2 are of Albian age, while the upper units are Cenomanian-Santonian. Recently, Eguiluz de Antuñano et al. (2023) suggested the separation of this group into three sequences: the Albian Cantelhá Sucession, the Cenomanian El Chango Sucession, and the Turonian-Santonian Cintalapa Sucession.
The Sumidero Quarry is on the southwestern border of the town of San Cristóbal de las Casas, Chiapas, southeastern Mexico, at the coordinates 16°42'33.90" N and 92°39'9.60" W (Figure 1). The limestone sequence exposed in this small commercial quarry belongs to the Sierra Madre Group. Here, the locals often extract beige-to-brown slabs of thin, laminar, and parallel marly limestones used to make floors or wall coverings. Massive yellowish limestone strata bound these thin, marly layers. The Sierra Madre Group is a Cretaceous (Aptian-Santonian) marine carbonate succession of at least 2575 m thickness, present in the central and western parts of the State of Chiapas, as well as in the north of Guatemala and the east of Belize (Steele, 1986; Waite, 1986). According to Salas (1949), in 1929, Nuttall W.L.F. identified this sequence in an unpublished work about the stratigraphy of southeastern Mexico, which he named the Sierra Madre Limestones. Subsequently, these were recognized as a tripartite Group involving the Albian-Cenomanian Cantelhá Formation plus the Turonian-Santonian Jolpabuchil and Cintalapa Formations (see Quezada-Muñetón, 1987). Steele (1986) and Waite (1986) studied the Sierra Madre limestones through 21 and 11 levels (named by them as units), respectively, concluding that units 1-2 are of Albian age, while the upper units are Cenomanian-Santonian. Recently, Eguiluz de Antuñano et al. (2023) suggested the separation of this group into three sequences: the Albian Cantelhá Sucession, the Cenomanian El Chango Sucession, and the Turonian-Santonian Cintalapa Sucession.
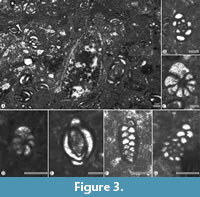 The preliminary review of the petrology and microfossils of the Sumidero Quarry fossiliferous strata reveals that these are of the Albian age and represent part of the Cantelhá Succession of the Sierra Madre limestones. These packstone strata have abundant miliolids (Figure 3), though also contain ostracods, pellets, small gastropods, and a diverse benthonic foraminiferal assemblage that includes specimens attributable to the following taxa: Anomalidae; Globorotalites Brotzen, 1942; Nezzazata Omara, 1956; Nummoloculina Steinmann, 1881; Quinqueloculina d'Orbigny, 1826; Spiroloculina d'Orbigny, 1826; Spiroplectammina Cushman, 1926; Textularia Defrance, 1924; Triloculina d'Orbigny, 1826; Valvulammina Cushman, 1933; and Valvulineria Cushman, 1926.
The preliminary review of the petrology and microfossils of the Sumidero Quarry fossiliferous strata reveals that these are of the Albian age and represent part of the Cantelhá Succession of the Sierra Madre limestones. These packstone strata have abundant miliolids (Figure 3), though also contain ostracods, pellets, small gastropods, and a diverse benthonic foraminiferal assemblage that includes specimens attributable to the following taxa: Anomalidae; Globorotalites Brotzen, 1942; Nezzazata Omara, 1956; Nummoloculina Steinmann, 1881; Quinqueloculina d'Orbigny, 1826; Spiroloculina d'Orbigny, 1826; Spiroplectammina Cushman, 1926; Textularia Defrance, 1924; Triloculina d'Orbigny, 1826; Valvulammina Cushman, 1933; and Valvulineria Cushman, 1926.
Similar lithology and microfossil content are present in Unit 2, documented by Steele (1986, p. 25) and Waite (1986, chert of biostratigraphic zones, p. 40-141), as well as in the Cantelhá Formation (e.g., Castro-Mora et al., 1975; Eguiluz de Antuñano et al., 2023). Such lithology and microfossil assemblage suggest that the fossiliferous strata of the Sumidero Quarry were deposited in a shallow marine environment, within a euphotic zone with moderate to high energy, in an open marine platform or an open lagoon (Quezada-Muñetón, 1987; Eguiluz de Antuñano et al., 2023).
MATERIALS AND METHODS
Preparation methods. Fossils were prepared mechanically with needles and dental excavators under a stereoscopic microscope. When necessary, the fossils were glued with cyanoacrylate and hardened with a weak solution of plexigum and methacrylate acetate applied with a fine brush. The specimen IGM 14038 is an impression; therefore, we prepared the fossils using the resin-transference and cleaning procedures with acetic acid, as described by Toombs and Rixon (1959). We studied all the specimens under the naked eye, stereomicroscope, and high-resolution photographs obtained under white light and UV light, with the specimens non-coated and coated with magnesium.
Institutional abbreviations. All the fossils reported in this work are deposited and cataloged into Colección Nacional de Paleontología, housed in the Instituto de Geología of the Universidad Nacional Autónoma de México, cataloged under the acronym IGLM. Microfossils illustrated here form part of three sedimentological thin sections of the Sumidero Quarry cataloged with the acronym IGM-mi.
Anatomical nomenclature and abbreviations. The taxonomical descriptions of the fossils studied here follow the anatomical nomenclature and abbreviations used in the publications cited in the Systematic Paleontology section.
SYSTEMATIC PALEONTOLOGY
Subclass ACTINOPTERYGII Cope, 1887
Order OPHIOPSIFORMES Ebert, 2018
Family OPHIOPSIELLIDAE Ebert, 2018
Genus TEOICHTHYS Applegate, 1988
TEOICHTHYS BREVIPINA Machado, Alvarado-Ortega, Machado, and Brito, 2013.
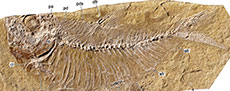
Specimen referred. IGM 14037, an incomplete specimen showing the left side of the head well preserved and the anterior part of the trunk somewhat twisted, with a partial length of approximately 51 mm (Figure 4).
 Occurrence. Albian marine deposits of the Cantelhá Formation exposed in the Sumidero Quarry, San Cristóbal de las Casas, Chiapas, southeastern Mexico.
Occurrence. Albian marine deposits of the Cantelhá Formation exposed in the Sumidero Quarry, San Cristóbal de las Casas, Chiapas, southeastern Mexico.
Description. The head is triangular and elongated; its length up to the posterior opercular edge is nearly 18 mm, and its maximum height at the nape is 11 mm. The skull represents three-quarters of the head length; the postorbital region is short and extends in the posterior third of the skull length, while the ethmoid and orbital areas are similar and occupy the remaining two-thirds. The mandibular joint is below the anterior third of the orbit. The rostral bone is roughly V-shaped in the ethmoid region and carries the anterior sensory commissure. The oblong nasal bone is expanded and slightly longer than wide.
In the skull, the frontal, parietal, dermosphenotic, and dermopterotic bones are sutured firmly to each other. Above, below, and behind the orbit, these and other bones are ornamented intensely with irregular tubercles and ridges of ganoine that reach the dorsal opercle tip and pectoral girdle bones. Notably, the parietals are rectangular, elongated, narrow, have a deep sinuous interparietal suture, and cover practically the entire postorbital skull region. Below the parietal, the small dermosphenotic bone forms the posterior orbital border. Posteriorly, the narrow dermopterotic bone is as long as the respective parietal. A large triangular extrascapular bone covers the occiput.
The circumorbital series is composed of at least three supraorbital bones, the dermosphenotic sutured to the skull, a mass of numerous suborbital bones covering most of the cheek, at least one high and smooth infraorbital, and an acicular anterorbital extending to the rostral. The upper jaw involves a small triangular premaxilla, an elongated triangular maxilla with a long and straight anterodorsal projection, a notched posterior edge, and a single long supramaxilla. The triangular lower jaw shows a shallow and long alveolar part. The jaw teeth are conical, broad-based, shallow, and acute-ended.
The opercular series includes four laminar bones; the opercle is kidney-shaped and higher than long; the preopercle is very narrow and crescent-shaped; the subopercle is elongated and semicircular; and the infraopercle is a small triangular bone. IGM 14037 has at least eight elongated and thin branchiostegal rays.
The vertebral column is incomplete; however, it is notable that the exposed centra are well ossified and laterally perforated by a pair of elongated cavities, and possibly, those behind the dorsal fin are diplospondylous. The pectoral girdle has a narrow, crescent-shaped cleithrum, a high oblong supracleithrum, and a triangular posttemporal. At least two large postcleithra are present. The pectoral fin is triangular, arises in the ventral edge of the trunk, involves nearly 23 rays, and covers a little less than the anterior half of the predorsal length of the trunk.
IGM 14037 only preserves the anterior 11 rays of the dorsal fin, far from the head. The predorsal distance of the trunk is slightly less than the length of the head; there are 19 scale rows before this fin, and this distance is at least twice the length of the pectoral fin. Thick rhomboid ganoine scales cover the trunk. These scales are smooth and not serrated. The scales are higher than long behind the head; however, these are square-shaped in the rest of the body.
Remarks. Teoichthys is an Albian Mexican representative of the order Ophiopsiformes, a clade of marine halecomorph fishes erected by Ebert (2018), which includes the Sinemurian (Middle Triassic)-Cenomanian (Middle Cretaceous) relatives of Ophiopsis Agassiz, 1834a. These fish represent 14 or 15 genera from Europe, America, Africa, and the Middle East (Ebert, 2019, 2020, 2021; Taverne, 2014; Taverne and Capasso, 2017). Among the diagnostic features of the order, the most easily observable include the presence of massive ganoine scales covering the trunk, the dermosphenotic incorporated into the skull, the maxilla notched posteriorly, narrow crescent-shaped preopercle, and the roughly V-shaped rostral bone with lateral horns carrying the anterior sensory commissure. IGM 14037 is an undoubted member of this order because it presents all these features.
The family Ophiopsiellidae, established by Ebert (2018), is the most derived clade within the Ophiosiformes. This family includes Middle Triassic-Late Jurassic species of Ophiopsiella Lane and Ebert, 2015, from Europe and Africa, initially classified under the genus Ophiopsis. It also encompasses Aptian-Cenomanian species related to Ophiopsiella from America, Africa, and the Middle East. The diagnostic features of this family present in IGM 14037 include the presence of one extrascapular on each side of the body and vertebral centra solid, perichordally ossified, and diplospondylous in the posterior part of the trunk.
Among ophiopsiellids, Ophiopsiella shows putative primitive features, such as the presence of ganoine rhomboidal scales equally longer than wide and serrated caudally; dorsal fin single, relatively small, and rising in the anterior half of the standard length (SL); skull bones unornamented; and cheek covered with scarce suborbital bones. Other ophiopsiellids show alternative conditions of these features (Table 1).
While the scales on Macrepistius and Agoultichthys are generally higher than they are long (Schaeffer, 1960; Murray and Wilson, 2009), those on Petalopterix, Teoichthys, and Placidichthys are higher than long at least in the predorsal trunk region (Pictet, 1850; Applegate, 1988; Brito, 2000; Brito and Alvarado-Ortega, 2008; Machado et al. 2013). Ophiopsiellids have scales with serrated posterior edges, with the exception of Teoichthys brevipina, which has scales that are not serrated. Teoichthys kallistos is an ophiopsiellid with a long and undivided dorsal fin (Applegate, 1988); however, in its sister species, T. brevipina, this is short and rises far from the head, at the beginning of the posterior half of the SL. In other ophiopsiellids (Petalopterix, Placidichthys, and Agoultichthys), the long dorsal fin occupies a large part of the anterior half of the SL and consists of two lobes. In Macrespistius, the dorsal fin is incomplete, but it is probably like in T. kallistos. Additionally, Teoichthys show two outstanding diagnostic features, including numerous ganoine tubercles and irregular ridges ornamenting the posterodorsal part of the skull and numerous small suborbitals covering the cheek. Other ophiopsiellids have an unornamented skull and scarce larger suborbitals, like those of Ophiopsiella. Here, we identify IGM 14037 as Teoichthys because it shares this genus’s aforementioned features.
The genus Teoichthys includes two species — T. kallistos and T. brevipina —described with specimens from the Albian marine deposits of Tlayúa Quarry (Applegate, 1988; Machado et al., 2013). As noted above, the dorsal fin is long in the first, whereas this fin is noticeably shorter in the last. Additionally, T. brevipina has a condition that differs from the other ophiopsiellids; its dorsal fin is far from the head, rises slightly behind the 50% of the SL after 23 scale rows, and its predorsal distance in the trunk is somewhat longer than the head length and at least two times the pectoral fin length (Machado et al., 2013, figure 1A). In other ophiopsiellids, regardless of whether this fin is long or short, it originates more anteriorly between the head and the middle of the SL. Notably, in T. kallistos, this fin increases by about 37% of the S.L. at a distance 1.4 times greater than the head length. Although the dorsal fin of IGM 14037 is incomplete, this specimen is identified as T. brevipinna because its dorsal fin arises far from the head, at a point on the body that at least doubles the length of the head and the pectoral fin length, and behind 19 lines of scales. Finally, although the skull ornamentation of IGM 14037 resembles the holotype of T. kallistos (IGM 3460), we dismiss its possible diagnostic value because this seems to be a widely variable feature in the type series of T. brevipina (IGM 6741-IGM 6747, and IGM 6604), in which the skull is practically smooth in some specimens and exhibits more conspicuous ornamentation in others.
Order MACROSEMIFORMES Grande, 2010
Family MACROSEMIIDAE Thiolière 1858
Genus NOTAGOGUS Agassiz, 1833
Notagogus novomundi González-Rodríguez and Reynoso, 2004
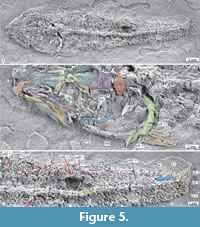 Specimen referred. IGM 14038, a complete specimen with and SL of about 78.5 mm and a somewhat twisted specimen transferred to resin, exposing the skull dorsal surface and the trunk left-dorsal surface (Figure 5).
Specimen referred. IGM 14038, a complete specimen with and SL of about 78.5 mm and a somewhat twisted specimen transferred to resin, exposing the skull dorsal surface and the trunk left-dorsal surface (Figure 5).
Occurrence. Albian marine deposits of the Cantelhá Formation exposed in the Sumidero Quarry, San Cristóbal de las Casas, Chiapas, southeastern Mexico.
Description. Given the preservation of this specimen, it is impossible to describe its body shape; however, it seems to be somewhat elongated, with the trunk covered by rhomboid scales. Its elongated head occupies about 30% of SL. The pectoral fin is broad, rounded, and probably restricted to the predorsal trunk region. A pair of possible rounded dorsal fins separated by a small hiatus occupy about 49% of the SL, between 38.5 and 84% of this length. The caudal peduncle is relatively high, and the caudal fin consists of 14 rays.
The rostral is a small acicular bone projected laterally in the skull ethmoid region. The nasal is a flat, smooth, drop-shaped bone covering most of this ethmoid region. The frontal bone is an elongated triangular bone that occupies nearly 80% of the skull length. This bone is smooth and has a straight interfrontal suture in its preorbital part. In contrast, the postorbital region shows sinuous sutures and tiny superficial ridges. Each frontal posteriorly joins a rectangular parietal with ornaments like the frontal. In the specimen, the skull bones are largely disarticulated and covered, making it difficult to describe.
The opercular series seems to be reduced to three bones because the interopercle is not discernible. The opercle is a broad and kidney-shaped bone superficially ornamented with tiny ridges. Below, the subopercle is an elongated ventrally-curving bone ornamented like the opercle. The preopercle is narrow and crescent-shaped, with large pores as openings of the preopercular-mandibular sensory canal. The circumorbital series is not well preserved; however, it is possible to recognize that the upper half of the orbit shows a mosaic of small supraorbital bones arranged in at least two rows. The dermosphenotic is crescent-shaped and sutured to the skull. The circumorbitals preserved below and behind the orbit are tiny scroll-like structures with no laminar component.
The jaw bones are not well-preserved. Nonetheless, the supramaxilla appears to be absent; the premaxilla is small and triangular; and the maxilla is elongated, posteriorly expanded, and bears a longitudinal maxillary sensory canal laterally exposed. The mandibular joint is below the middle of the orbit. The upper and lower jaws show robust and sharp conical teeth fragments.
Bones of the axial skeleton are mostly covered by the scales; however, some partially exposed vertebrae along the body show that probably all of them, including those of the caudal skeleton, are well ossified, at least perichordally. In the pectoral girdle, the cleithrum is narrow and inverted-C shaped in lateral view, and posteriorly, it has at least three large-scale-like postcleithra. At the dorsal end of the pectoral girdle, the posttemporal bone is triangular, expanded ventrally, with rounded edges, and ornamented with reticular ridges. The pectoral fin consists of 12 rays with no preceding fulcra. The anal and pelvic fins are not exposed. Two dorsal fins divided by a small hiatus occupy most of the trunk length; these are similarly sized and have 13 rays each. The caudal fin is bent and rests below the body; however, its base reveals that it consists of 14 rays dorsally preceded by at least three large basal and six fringing fulcra.
Rhomboid scales cover the whole trunk. These scales are evenly sized, have peg-and-socket joints, and are finely serrated along the posterior edges (observed in well-preserved scales). These lack or have a weak ganoine layer that exhibits the growth rings. The scales do not carry the lateral line.
Remarks. Macrosemiidae Thiollière, 1858 (=Macrosemiiformes, Grande (2010, p. 860), but see López-Arbarello (2019)) is an extinct clade of ginglymodian fishes that includes 8-10 marine genera of Norian (Late Triassic)-Cenomanian (Late Cretaceous) deposits of Europe, northwestern Africa, and southern North America (Mexico). According to different authors, the members of this group share some diagnostic features; these are long-dorsal fin fishes with small interopercle located far from the mandible, pectoral fin with no fulcra, and nine scroll-like infraorbitals bones with two placed behind the orbit (e.g., Bartram 1977; González-Rodríguez and Reynoso, 2004; Arratia and Schultze, 2012). IGM 14038 is identified as an unmistakable member of this group because it exhibits all these features. Although it does not preserve all the infraorbitals, those present are scroll-like behind and below the orbit (Figure 5).
Among macrosemiiforms, only Notagogus Agassiz, 1833, has the following distinctive diagnostic features: scales cover the whole of the trunk; a short hiatus separates two dorsal fins; and all centra are perichordally well ossified (Bartram, 1977; Saint-Seine, 1949, figure 86). Other macrosemiiforms have a scale-less area below the dorsal fin, one long dorsal fin or two dorsal fins separated by a comparative long inter-dorsal hiatus. In the vertebral column, only the most anterior centra show poor ossification. In addition, according to González-Rodríguez and Reynoso (2004), the monophyly of Notagogus is supported by three synapomorphies, including the medium depth of the body, infraorbitals without laminar component and a reduced ganoine layer in the scales. Here again, we identified IGM 14038 as part of Notagogus because it shows all the distinctive and synapomorphic features, except for that referred to the body shape, which is undefinable in this specimen.
Among macrosemiiform, Notagogus is the most diverse genus and includes eight nominal species, seven from European Late Jurassic sites and one from the Albian limestones of the Tlayúa Quarry, Mexico (e.g., Bartram, 1977; Bravi, 1994). Although it is a homoplastic feature, González-Rodríguez and Reynoso (2004) discovered that the presence of a mosaic of small supraorbital bones ordered in two rows is present in only three Notagogus species, including their N. novomundi from the Tlayúa Quarry, N. helenae (Thiollière 1850) from Cerin, France, and N. pentlandi Agassiz, 1834b, from the Albian deposits of Torre d'Orlando, Italy. This character is also in Macrosemiocotzus americanus González-Rodríguez, Applegate, and Espinosa-Arrubarrena (2004) from Tlayúa Quarry. IGM 14038 is identified here as N. novomundi because its anterior and posterior dorsal fins have 13 rays each, and its maxilla shows an open sensory canal along the maxilla. In contrast, N. helenae and N. pentlandi have no exposed sensory canal in the maxilla, and their anterior and posterior dorsal fins have 15+10-12 and 14+10 rays, respectively (Bartram, 1977).
Order PYCNODONTIFORMES Berg, 1937
Family PYCNODONTIDAE Agassiz, 1833 sensu Nursall 1996a
Indeterminate genus and species
 Specimen referred. IGM 14039, an incomplete specimen with 55 mm partial length, exposing the left side of the body, lacking the snout, jaws, and unpaired fins (Figure 6).
Specimen referred. IGM 14039, an incomplete specimen with 55 mm partial length, exposing the left side of the body, lacking the snout, jaws, and unpaired fins (Figure 6).
Occurrence. Albian marine deposits of the Cantelhá Formation exposed in the Sumidero Quarry, San Cristóbal de las Casas, Chiapas, southeastern Mexico.
Description. IGM 14039 preserves almost the entire trunk and perhaps only lacks the arcocentra of the caudal peduncle. The preserved trunk suggests the specimen is a nearly round-bodied fish, slightly longer than high (body depth ratio 55/50 mm). Here, the insertion point of the pelvic fin is in the middle of the trunk, opposing the end of the first third of the dorsal fin base. The cloacal notch is equidistant between the bases of the anal and pelvic fins. The positions of the dorsal and anal pterygiophores reveal that the long dorsal fin extends in the posterior trunk half, and the short anal fin opposes the posterior half of the dorsal fin.
On the skull roof, the dermosupraoccipital rotated in its position and now shows its ventral surface. The left preopercle and the right parietal and opercle are displaced upwards, so the supraoccipital exposes its laminar dorsal process. In this part of the skull, the sutures between the frontal, parietal, dermopterotic, and dermosphenotic bones are intensely zigzagging. The parietal is an X-shaped bone with wide arms; the anterior limb sutures the frontal; the dorsal and ventral ones reach the dermosupraoccipital and dermopterotic, respectively; and the posterior one represents the posterior parietal process that is distally branched. The parietal and dermosupraoccipital have confluent notches that form the posterior part of the temporal fenestra, which must have been closed by the frontal. The dermopterotic is rectangular, occupies most of the postorbital skull region, and posteriorly sutures with a small extrascapular. All these bones are strongly ornamented with anastomosed reticulated ridges and numerous pores, which have a radial order from the central part of each bone. The dermosphenotic and the contiguous infraorbital are preserved and poorly ornamented.
Dermohyomandibular and preopercle occupy most of the cheek. The first is an elongated rectangular bone scarcely ornamented in the posterior part and ending on an elongated and curved dorsal articular head. The preopercle is rectangular, as long as the dermohyomandibular but at least three times higher, strongly ornamented like the skull bones, and bears a stout and long anterodorsal process. The opercle is a long bone, concave anteriorly, and expanded ventrally. The subopercle and infraopercle are not present.
The vertebral column consists of at least 26 pairs of arcocentra, including 14 abdominals and at least 12 caudals, which do not invade the notochordal canal that remains empty from the central arch 5. Anteriorly, the synarcual occupies this space. All neural and hemal arches are laterally triangular, small, autogenous, and show a broad notochordal border. The contact between adjacent arcocentra is simple through their pre- and postzygapophyses, which are short spine-like structures that are projected longitudinally and similar in shape and size throughout the trunk. All the neural and caudal-hemal arcocentra have long spines with anterior sagittal flanges or triangular laminar wings that rise from the prezygapophyses and border most of the anterior edge of each spine. An indeterminate number of ribs articulate with the abdominal-hemal arcocentra. These ribs are long, slightly curved, and possess wing-like structures from the head of the element.
All the fins are not preserved except for parts of their internal supports. In the pectoral girdle, the cleithrum is a short and high crescent bone, anteriorly concave and located between the ventral edge of the body and the vertebral column. Additionally, the cleithrum is dorsally acute, ventrally expanded, and with its ventral third surface strongly ornamented with ridges and pores. The point of insertion of the pelvic fin is present between scales just above the abdominal edge. The dorsal fin is wholly disarticulated and separated from the trunk; only a few scattered remains of dorsal rays are near the fossil. The fossil preserves eight rays of the anal fin. There are 34 dorsal pterygiophores extended in the posterior half of the dorsal body edge, while only 20 anal pterygiophores are present in the posterior third of the ventral body edge. In both series, the pterygiophores are long, straight, and flat.
In this fish, the scale pattern is peltate. The trunk has no scales in the caudal region between the unpaired fins. In contrast, two types of scales cover only the most anterior part of the trunk. The predorsal trunk region shows 12 or 13 rows of modified bar-like scales, each row with three to six scales, anterodorsally tilted and overlay the neural spines. Broad and imbricate scales cover the abdominal trunk region and form 18 to 20 scale rows.
Although in life, this fish must have had a complete series of predorsal and preanal bordering scutes, only two dorsal ridge scutes are disarticulated and scattered near the trunk. The dorsal ridge scutes are stout, triangular, and non-serrated. Nineteen ventral keel scutes from a complete series between the isthmus and the anal fin; these scutes include 17 precloacals (the posterior one is behind the pelvic fin) and two postcloacals. The ventral keel scutes are triangular and ventrally thick, have blunt ends contacting the adjacent scutes, and show tiny, thin lateral wings. These scutes are unserrated, except for the postpelvic and both postcloacals, which have four or five medial spines, uniformly small and evenly spaced. Small, unmodified scales border the cloacal vestibule. The posterior postcloacal scute and last abdominal scales cover most of the postcoelomic; however, this is a high triangular bone, laterally flat, smooth, and extends up to the half of the abdominal cavity, far from the vertebral column.
Remarks. Pycnodontiformes is a globally distributed actinopterygian clade with controversial phylogenetic relationships. This Norian (Late Triassic)-Eocene (Paleogene) clade includes Pycnodus Agassiz, 1833, and relatives (Berg, 1937; Nursall, 1996b; Blot, 1987). These highly specialized, primarily marine fishes also had repetitive invasions in freshwater environments (Szabó et al., 2016; Cawley and Kriwet, 2024).
Early phylogenetic studies of Pycnodontiformes identified the features that support this order and its subordinal clades (Nursall, 1996a; Poyato-Ariza and Wenz, 2002). Among the multiple diagnostic features of this order listed by these authors, IGM 14039 shows a high and rounded body that is compressed laterally, the notochordal canal empty because the vertebrae are not ossified, and only ventral and hemal arcocentra occupy this part of the skeletal axis. The skull roof shows a single dermosupraoccipital. The subopercle and interoperculum are missing. The opercle is crescent-shaped and small, and the preopercle is extremely large. The ribs are winged. Finally, the fish has a dorsal ridge and ventral keel scutes bordering its predorsal and preanal trunk regions.
Within Pycnodontiformes, the family Pycnodontidae is strongly supported by several diagnostic features (Payato-Ariza and Wenz, 2002). IGM 14039 is identified as part of this family because it shows the following diagnostic features: the posterior parietal process and dermohyomandibular are present; the scales of the predorsal trunk region are bar-like, and there are six or fewer postcloacal keel scutes.
Pycnodontidae is the most successful pycnodontiform clade. This family contains about 80 species representing 40 genera, and its classification still faces problems. In the latest phylogenetic analyses including this family, the existence of the contrasting traits of three subfamilies has been confirmed, including Pycnodontinae, Nursallinae, and Turboscinetinae (e.g., Nursall, 1996a; Ebert, 2016). It is worth noting three issues: 1) the monophyly of Pycnodontinae has recently been questioned because its members seem to represent different stem taxa located at the base of the nursallines (now reclassified as tribe Nursallini); 2) Turboscinetinae includes the members of Proscinetinae (now re-ranked as the tribe as Proscinetini); and 3) Ebert (2016) recognized his Group B as a monophyletic clade containing the members of the subfamily Pycnodontinae (see, Poyato-Ariza and Wenz, 2002, Poyato-Ariza, 2020; Ebert, 2016; Taverne and Capasso, 2018). In this scenario, considering only four features of GM 14039, including the presence of the dermocranial fenestra, the peltated scale pattern, the posterior part of the postorbital skull region well ossified, and the lack of free dorsal pterygiophores allow us to identify it as a non-pycnodontine pycnodontid fish belonging to the Group B (Table 2).
Group B members and IGM 14039 have dermocranial fenestra, but this cavity does not occur in Turboprocinetinae and Nursallinae. Considering the kind of scales (modified bar-like scales and unmodified broad scales) and their distributions in the predorsal, abdominal, and caudal trunk regions, Nursall (1996a, figure 19) recognized distinctive scale patterns in different pycnodontiform groups. The scale pattern of IGM 14039 and Turboprocinetinae share a peltated pattern, with bar-like scales on the predorsal region, broad scales covering the abdomen, and the caudal region being nude. Nursallinae members show the imbricated pattern (broad scales cover the whole trunk) and a peculiar scale pattern that here is named peltated-expanded, in which the bar-like scales are on the predorsal and caudal regions of the trunk while broad scales cover the abdomen. Members of Group B show three patterns: non-Pycnodontinae are peltated as in Turboprocinetinae, some Pycnodontinae are calthrated (the caudal region is nude, with broad scales covering a small part of the abdomen close to the ventral trunk edge and with bar-like scales covering the rest of the abdomen and all the predorsal trunk region, as in Pycnodus). Some Pycnodontinae also show clathrate-reduced morphology, in which the distributions of both kinds of scales are comparatively reduced, the broad scales covering a small area near the abdominal trunk edge, and the bar-like scales are only near the predorsal trunk edge, as in Pseudopycnodus Taverne, 1997. Within Group B, Pycnodontinae have reduced bones in the postorbital part of the skull and most of the supraoccipital bone in lateral view; in IGM 14039 and other pycnodontids, these bones are well ossified and cover the supraoccipital bone. Finally, in non-procinetine members of Turboscinetinae, most anterior dorsal pterygiophores are free or do not meet with any dorsal ray; however, other pycnodontids have no free dorsal pterygiophores. IGM 14039 has no dorsal fin rays; therefore, it is impossible to recognize the presence or absence of the last condition.
Order ASPIDORHYNCHIFORMES Bleeker, 1859
Family ASPIDORHYNCHIDAE Nicholson and Lydekker, 1889
Geneus BELONOSTOMUS Agassiz, 1834b
Belonostomus sp.
 Specimen referred. IGM 14040, an incomplete serpentiform and longirostrine fish exposed on the right side of the body, in which the head is impressed, and the tail and terminal part of the snout are missing (Figure 7). The length of the specimen is 175 mm.
Specimen referred. IGM 14040, an incomplete serpentiform and longirostrine fish exposed on the right side of the body, in which the head is impressed, and the tail and terminal part of the snout are missing (Figure 7). The length of the specimen is 175 mm.
Occurrence. Albian marine deposits of the Cantelhá Formation exposed in the Sumidero Quarry, San Cristóbal de las Casas, Chiapas, southeastern Mexico.
Description. The body of this fish is bent, with the tips of the snout and caudal region inclined ventrally. The specimen only preserves 175 mm of its total length. On the trunk length, the pelvic bone (= base of the pelvic fin) is at 97 mm, and the anal and dorsal fins (opposed to each other) are far in the back, at 152 mm. The height of the head is 22 mm, the most anterior part of the trunk shows the same size; however, this becomes shallower backward and reaches 12 mm at the base of the anal/dorsal fin (Figure 7).
Overall, the skull is incompletely impressed; however, the orbital and postorbital lengths are similar. The lower and upper jaws are elongated, almost uniformly high through their length, and have straight alveolar borders. Below the ethmoid region, the part of the lower jaw corresponding to the dentary shows that the alveolar and ventral borders are parallel. The lower jaw articulation is below and near the posterior limit of the orbit.
The impression of the bones of the opercular series and pectoral girdle are imperfect. Here, small parts of the opercle, subopercle, and anterior limb of the cleithrum are present. A fragment of the posttemporal lays horizontally dorsal to the nape. Short, curved, and shallow ganoine ridges ornament these bones. The pectoral fin is trapezoidal, arises in the ventral edge of the body, and consists of 12 short rays, of which the first two are remarkably broad.
At least 53 vertical scale rows entirely cover the preserved part of the trunk, each composed of two dorsal, at least two ventral, and three flank scales. The dorsal flank scales are somewhat hexagonal with rounded edges. The ventral flank scales are rectangular and slightly longer than high. The three flank scales have different shapes; the lower and middle scales show a vertical alignment, and the upper scale is inclined anterodorsally. The lower flank scales are rectangular, slightly higher than long. The middle flank scales are rectangular, 3-4 times higher than long. In each flank scale, the lateral line runs at the base of the upper quarter, which is inflected anteriorly. The upper flank scales are rhomboidal, anteriorly tilted, and about 1.5 times higher than long. The dorsal edges of the scales of dorsal, ventral, and lower flank rows have a triangular dorsal process, revealing their peg-and-socket joints. The middle and upper flank scales are poorly imbricated, and their joint is nearly straight. The joint between the upper flank and dorsal scales is unknown in the specimen referred. These scales do not have posterior dentitions or serrae. Short, curved, and shallow ganoine ridges cover the surfaces of all these scales, though the ornamentation is more conspicuous in the dorsal members than those on the flanks and ventral portions of the body.
An unknown number of pelvic rays and bones of the pelvic girdle are disarticulated but lie within the posterior half of the abdomen, below the scale rows 21 and 29, respectively. Here, the pelvic bone is rectangular and anteriorly acute, and the impression of the pelvic rays shows that the first one is broad, like that of the pectoral fin. The anal and dorsal fins are placed far in the back of the body, and the anal and dorsal fin are below and above the scale rows 43 and 45, respectively. Given the specimen’s poor preservation, the number of dorsal rays is unknown, but the anal fin is composed of at least seven rays.
Remarks. Aspidorhynchiformes (= Aspidorhynchidae) is an extinct single-family order of marine and highly specialized serpentiform fishes with elongated snouts (Van Vranken et al., 2019) that has a global distribution spanning the Middle Jurassic (Bathonian)-Late Paleocene (Thanetian) clade. IGM 14040 is assigned to this family because it shows some of its diagnostic features (Brito, 1997); the body is serpentiform, the anal and dorsal fins are placed far in the back of the body, and there are three longitudinal rows of flank scales.
Currently, Aspidorhynchidae comprises six genera that show noticeable differences in the bones of both jaws, opercular series, and proportions of the flank scales (Brito, 1997; Bartholomai, 2004; Gouiric-Cavalli, 2015, figures 6, 8; Arratia, 2015). In his comprehensive review of Aspidorhynchidae, Brito (1997) amended the diagnoses of Belonostomus and Vinctifer Jordan, 1919, pointing out that these share the presence of a longitudinal row of high rectangular flank scales (= middle flank scales in this work) that differ noticeably from the less high scales placed above and below. Later, Jonoichthys Gouiric-Cavalli, 2015, was revealed to have such flank-scale condition. However, in this Argentinian Late Jurassic fish, the articulation observed between the middle and upper flank scales is deeply curved, whereas, in the first two genera, this articulation is relatively straight.
Vinctifer, Aspidorhynchus Agassiz, 1833, and Richmondichthys Bartholomai, 2004, have short snout heads. These taxa and Pseudovinctifer Arratia, 2015, show a triangular lower mandible, including a small anterior predentary and a trapezoidal dentary that tapers anteriorly and is high posteriorly. Consequently, in these four fishes, the alveolar and ventral edges in the posterior part of the lower jaw (mainly formed by the dentary) are inclined to each other. In addition, at least in Vinctifer and Richmondichthys, the upper jaw consists of a triangular maxilla significantly expanded posteriorly plus a thick, saber-like premaxilla that is elongated, straight, and anteriorly projected far from the lower jaw tip (Brito, 1997, figures 6, 13). On the other hand, Belonostomus and Jonoichthys have noticeably elongated snouts, in which the alveolar and ventral edges of the lower jaw are parallel, plus an elongated triangular laminar premaxilla. These last two genera and Aspidorhynchus have a narrow and elongated maxilla (Brito, 1997, figures 42, 43).
Despite the lack of a preserved tail and anterior tip of the snout, IGM 14040 is identifiable as Belenostomus. In its head, the preorbital region has no space to accommodate an expanded bone like the maxilla in Vinctifer and Richmondichthys. Additionally, as Belenostomus and Jonoichthys, the proximal part of the lower jaw of this Mexican aspidorhynchid has parallel alveolar and ventral edges, and its middle flank scales are notably highest at their joint with the upper adjacent scales and are straight as in other aspidorhynchids except for Jonoichthys (in which these scales have peg-and-socket joints).
Among aspidorhynchids, the Belonostomus is the most diverse and widely distributed geographically and temporally. This genus contains at least 15 nominal species from Kimmeridgiano (Late Jurassic)-Thanetian (Late Paleocene) sites of Europe, Northern Africa, North and South America, and Asia (e.g., Taverne and Ross, 1973; Brito, 1997; Bogan et al., 2011; Ebert, 2014). These species differ in pterygial formula, body and scale proportions, ornamentation, and features of jaws, teeth, and skull bones. Given the incomplete preservation, we identify IGM 14040 as an indeterminate species of this genus.
Superorder CLUPEOMORPHA Greenwood et al., 1966
Order ELLIMMICHTHYIFORMES Grande, 1982
Family UNNAMED
Genus THORECTICHTHYS Murray and Wilson, 2013
Thorectichthys sp.
 Specimen referred. IGM 14041, a nearly complete fish, exposed on the left side, with an estimated SL of 58.91 mm, and without the bones in front of the orbit (Figure 8).
Specimen referred. IGM 14041, a nearly complete fish, exposed on the left side, with an estimated SL of 58.91 mm, and without the bones in front of the orbit (Figure 8).
Occurrence. Albian marine deposits of the Cantelhá Formation exposed in the Sumidero Quarry, San Cristóbal de las Casas, Chiapas, southeastern Mexico.
Description. The specimen lacks the anterior part of the head, while its rostral and caudal ends are somewhat curved dorsally; thus, all the following measurements are estimated. It is 79 and 62 mm in total and SL, respectively. Expressed as the percentage of the SL, the head length is 30% (18.6 mm), the predorsal length is 46% (28.8), the prepelvic length is 57% (35.7 mm), and the preanal length is 68.2% (42.3 mm). The height of the head is 33% (20.5 mm), the maximum height of the body is 37.5% (23.3 mm), with the height of the caudal peduncle being 9.6% (6 mm). The dorsal fin is shorter than the anal fin. Their respective lengths are 12.5% (7.8 mm) and 22.9% (15 mm).
Although the head is not well-preserved, it exposes some valuable features. The large parietals are rectangular, and probably both meet at the midline. The orbit is entirely circular; its diameter equals the postocular length of the skull, and two thin, semi-circular sclerotic bones occupy it. The jaw-quadrate joint is below the posterior half of the orbit. The preopercle and subopercle are not well preserved in the opercular series. The interopercle is an oblong and sharp-ended bone. The operculum is a kidney-shaped bone with straight vertical striations ornamenting its ventral part.
The vertebral column involves 35 total centra, including 18 abdominals, 15 caudals, and two urals. There are 16 pairs of long curved ribs enclosing almost the entire abdomen. The eight posterior ribs articulate long parapophyses, and the anterior ones do this directly with the lateroventral cavities of the centra. There are seven predorsal bones. The most anterior neural spines are distally bifid; long epineurals are present along the trunk up to the end of the anal fin; those in the abdominal trunk are long and thread-like, and those behind become shorted and triradiated. On the caudal region, the epipleurals are similar and symmetrically opposed to the epineurals.
The cleithrum is S-shaped in lateral view, with a small descending anterior limb. Sixteen rays form a triangular pectoral fin rising slightly above the ventral trunk edge. The pelvic fin is small, opposed to the posterior half of the dorsal fin, and involves at least six rays. The dorsal fin is relatively short and consists of two procurrent and 14 long rays that extend between 46% and 58.5% of the SL. The anal fin is long, opposed to the postdorsal part of the trunk, and consists of two procurrent and 25 long anal rays extended between 68.2% and 91.1% of the SL. The caudal fin is deeply forked and consists of two long triangular lobes symmetrically opposed. Its caudal formula is viii+I+9—8+I+vi.
Oval-shaped cycloid oval scales cover the entire trunk. Numerous concentric sinuous circuli ornament the scales superficially. Thick scutes border the predorsal and preanal trunk edges. Nearly 11 predorsal scutes form a complete series between the occiput and the dorsal fin; these are oval, slightly expanded anterolaterally, and superficially ornamented with straight and parallel shallow ridges. A small middle posterior spine protrudes in the posterior edge of the most posterior predorsal scutes. The abdominal keel of this fish involves a complete series of at least 22 ventral scutes, including about 15 prepelvic and seven postpelvic. These scutes have a thick ventral edge and lateral triangular wings covering the ventral third of the abdomen. In the postpelvic ventral scutes, the medial thick bar is projected posteriorly as a stout spine.
Remarks. Grande (1982, 1985) erected Ellimmichthyiformes to include the putative primitive and extinct clupeomorph fishes resembling Ellimmichthys Jordan and Gilbert, 1919. Today, this clade includes nearly 50 nominal species, gathered in 22 genera, recovered in Valanginian (Early Cretaceous)-Eocene (Paleocene) marine and freshwater deposits worldwide. The different phylogenetic hypotheses of Ellimmichthyiformes recently proposed show contradictory details (Alvarado-Ortega et al., 2020). Nevertheless, as with all clupeomorph fish, the trunk of IGM 14041 shows a ventral keel. Additionally, this fossil is identified as part of Ellimmichthyiformes because it shows large parietals that contact one another medially, along with ornamented predorsal scutes expanded anterolaterally and bearing a posterior spine.
Murray and Wilson (2013) erected Thorectichthys for T. marocensis and T. rhadinus, two Moroccan species recovered in the marine Cenomanian-Turonian deposits of the Akrabou Formation. Recently, a North American species of this genus was named T. fideli Than-Marchese, Alvarado-Ortega, Murray, Velázquez-Velázquez, and Domínguez Domínguez, 2024, based on nicely preserved specimens from the early Cenomanian deposits of the El Chango Quarry, Chiapas, Mexico. IGM 14041 is identified as Thorectichthys because it shows the most conspicuous diagnostic features of the genus: straight longitudinal parallel ridges ornamenting the predorsal scutes.
A brief comparison of Thorectichthys species and IGM 14041 reveals some differences in the proportions of the body (see Than-Marchese et al., 2024, tbl. 2). This fossil is the slenderest of Thorectichthys, with its maximum body height (= 37.5% of the SL) surpassed by those observed in the three nominal species (=40-65% of the SL). It is also the only Thorectichthys species with a dorsal fin so far on the body (46% of the SL versus 36-44% of the SL in nominal Thorectichthys species). Nonetheless, IGM 14041 does not refer to any of these species or a new one simply because its body proportions are estimated.
DISCUSSION
According to Guinot and Cavin (2016), ray-finned fishes experienced the second-largest diversification event in gnathostome evolutionary history during the Cenomanian. At this time, a dramatic taxonomic turnover occurred and led to the establishment of the Late Cretaceous ichthyofauna, which later led to the emergence of the dominant modern groups during the end-Cretaceous and early Cenozoic (Friedman et al., 2023). Among other factors, Guinot and Cavin (2016) also noted that the rise in sea levels and the fragmentation of the continents allowed the development of large epicontinental seas that favored the diversification processes of coastal fishes. Given the importance of such a biological event, considering some new questions to understand this holistically is valid. In this scenario, the review of the diversity of Albian-Cenomanian bony fishes of both marine contours and continental waters is essential to recognizing the possible regional or local components of the Cenomanian Diversification Event of Bony Fishes (CDEBF), as well as the potential effect of the different biological-environmental factors that triggered it.
In recent years, the diversity and paleontological sites of Cretaceous fossil fishes in North America have been reviewed on continental, regional, or local scopes by different authors (e.g., Applegate 1996; Wilson and Bruner, 2004; Alvarado-Ortega et al., 2006; Everhart, 2009; González-Rodríguez et al., 2013a, 2016; Murray and Cook, 2016). Without being copious yet, the accumulated data is beginning to reveal some traits of the CDEBF on the North American subcontinent. With this in mind, and to assess the importance of the Albian ichthyofauna from the Sumidero Quarry, we compiled data on the North American marine bony fishes from the Albian-early Cenomanian, only including nominal species and reliable generic identifications. This work does not include materials of a more extended temporal range because the fossil fish record from Mexico is deficient between the Kimmeridgian and Early Albian, and the material recovered in two middle Cenomanian sites, El Tanque and La Mula, in the north of Coahuila, near the Mexico-U.S.A. border, lack a strict stratigraphic control within sequences that extend up to the Santonian (Giersch et al., 2008; Giersch, 2014).
Table 3 and Figure 9 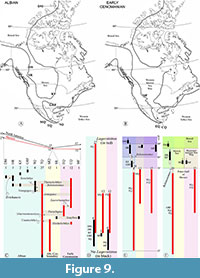 summarize the results of our analysis of the North American Albian-early Cenomanian fishes. Our study reveals that North America has only 11 Albian-early Cenomanian localities carrying well-preserved osteichthyans, where the quality of the fossils allows their accurate alpha taxonomic identifications. These well-preserved localities are essential for studying taxonomic diversification processes, as in the case of CDEBF. These localities have a remarkable unbalanced geographic distribution; three of them are in the west and north of Canada; other three are in the central, southeastern, and western regions of the USA, and with the addition of the Sumidero Quarry, the remaining five are in the central and southern portions of Mexico (Figure 9A and B). These sites include:
summarize the results of our analysis of the North American Albian-early Cenomanian fishes. Our study reveals that North America has only 11 Albian-early Cenomanian localities carrying well-preserved osteichthyans, where the quality of the fossils allows their accurate alpha taxonomic identifications. These well-preserved localities are essential for studying taxonomic diversification processes, as in the case of CDEBF. These localities have a remarkable unbalanced geographic distribution; three of them are in the west and north of Canada; other three are in the central, southeastern, and western regions of the USA, and with the addition of the Sumidero Quarry, the remaining five are in the central and southern portions of Mexico (Figure 9A and B). These sites include:
1) Dragon Mountain—Middle Albian marine sequence belonging to the Macdougall Point Member of the Christopher Formation, exposed on Dragon Mountain at the western part of Axel Heiberg Island, Nunavut, Canada (see Hermus et al., 2004; Murray and Cook, 2016).
2) Hay River—Middle-late? Albian marine deposits of the Loon River Formation exposed alongside the Hay River, Northwest Territories, Canada. (Forey, 1975; Wilson and Brunner, 2004; Murray and Cook, 2016; Maxwell and Caldwell, 2006; Vernygora et al., 2016).
3) Kiowa Formation—Outcrops of the late Albian Champion Shell Bed at the base of the Kiowa Formation, exposed in the vicinity of Belvidere in Kiowa County, as well as in the Kanopolis Reservoir in Ellsworth County, the area around the Clark County State lake in Clark County, and an unreported point in McPherson County, in Kansas, USA (Williston, 1894; Cragin, 1894; Scott, 1970; Schultze and Stewart, 1982; Russell, 1988; Beamon, 1999; Everhart, 2004, 2009, 2011).
4) Glen Rose Formation—The single valid nominal species as far known in this formation is Macrepistius arenatus Cope, 1894, described based on a fragmented specimen recovered in the marine deposits of the middle Albian along the Peluxi River that belong to the lower part of the Glen Rose Formation, near Glen Rose town, Somervell County, Texas. The deposit of this formation occurred in shallow marine conditions on the northern edge of the Paleo-Gulf of Mexico (Martha et al., 2019). Additional unidentified fish remains (a possible clupeiform and a holotean) of this formation are from the area near Granbury town, Hood County, Texas (Schaeffer, 1960).
5) Sumidero Quarry—Described in this work.
6) Tlayúa Quarry—Middle-late Albian limestone strata belonging to the Middle Member of the Tlayúa Formation (Pantoja-Alor, 1992), exposed in the Tlayúa Ravine, outside of Tepexi de Rodríguez, Puebla, central Mexico (e.g., Applegate and Espinosa-Arrubarrena, 1982; Applegate et al., 2006; Alvarado-Ortega et al., 2007, 2009; Espinosa-Arrubarrena and Alvarado-Ortega, 2010).
7) Muhi Quarry—Laminated calcareous sequence of the Albian-Cenomanian boundary belonging to the La Negra Facies of the El Doctor Formation, quarry in the eastern side of the Zimapán, Hidalgo, central Mexico (Bravo-Cuevas et al., 2012; López-Palomino et al., 2021)
8) Smoky River—According to Wilson and Murray (1996), this site is immediately west of the town of Judah Station, on the east bank of the Smoky River, Alberta, Canada. The site is an outcrop of the Fish Scale Zone of the Albian/Cenomanian boundary, which forms part of the Shaftesbury Formation.
9) El Espinal Quarry—The locality represents a small group of small artisanal quarries located west of the El Espinal de Morelos, Municipality of Ocozocoautla de Espinosa, Chiapas, southeastern Mexico (e.g., Alvarado-Ortega and Ovalles-Damián, 2008). Initially, the age of these fossiliferous outcrops was though Albian; however, recent works have confirmed that this belongs to the early Cenomanian sequence of the Cintalapa Formation (Alvarado-Ortega et al., 2009; Alvarado-Ortega and Than-Marchese, 2012, 2013; Moreno-Bedmar et al., 2014; Eguiluz de Antuñano et al., 2023).
10) El Chango Quarry—Early Cenomanian laminar limestones of the Cintalapa Formation, exposed near the town Pluma de Oro, south of the Municipality of Ocozocoautla de Espinosa, Chiapas, southeastern Mexico (Alvarado-Ortega et al., 2009; Alvarado-Ortega and Than-Marchese, 2012, 2013; Moreno-Bedmar et al., 2014; Eguiluz de Antuñano et al., 2023).
11) Mowry Formation—Earliest Cenomanian deposits of the Mowry Formation, exposed in Vernal County, Utah, U.S.A. (Dunkle, D., 1971; Newbrey et al., 2013; Stewart and Hakel, 2006).
The known taxonomic diversity of the North American marine osteichthyans during the Albian-early Cenomanian is low, partly due to the low number of paleontological localities (11) discovered in this continental region with well-preserved fossils (Figure 9A and B, Table 3). Nevertheless, the known taxonomic diversity includes 45 nominal species that could reach a maximum of 67 if we consider that all indeterminate generic reports represent at least one species (Table 3). The same record shows 55 genera, of which 32 are in the Albian (including 27 exclusively Albian, Unamichthys that reach the Albian-early Cenomanian boundary and Armigatus, Thorectichthys, Paraclupea, and Macrosemiocotzus also present in the early Cenomanian), 13 represent the Albian-Cenomanian boundary (including 10 exclusively present in this boundary, Unamichthys also present in the Albian, plus Xenyllion and Heckelichthys that reache the early Cenomanian (Figure 9C); this count does not include Armigatus, Thorectichthys, Paraclupea, and Macrosemiocotzus that could potentially be present in this boundary because their distributions range between the Albian to the early Cenomanian), 17 are early Cenomanian (including 11 exclusively of this age, Xenyllion and Heckelichthys present in the Albian-early Cenomanian boundary, plus Armigatus, Thorectichthys, Paraclupea, and Macrosemiocotzus also present from the Albian to early Cenomanian). At this point, it is essential to highlight the relevant contribution of Mexico to fossil record because together, its Lagerstätten (the Tlayúa, Muhi, and El Chango quarries) and its non-lagerstäten sites (the Sumidero and El Espinal quarries) contribute most of the generic diversity of North American Albian-early Cenomanian bony fishes. In comparing the fossil contributions within the same geological periods, the Mexican sites include 27 of 32 known genera for the Albian, 12 of 13 genera for the Albian-Cenomanian boundary, and 15 of 17 genera for the early Cenomanian. However, such generic diversity barely changes if only the Mexican Lagerstätten are considered; this diversity is 27 of 32 for the Albian, 12 of 13 for the Albian-Cenomanian boundary, and 12 of 17 for the early Cenomanian (Figure 9C).
Recently, Arratia and González-Rodríguez (2024) noted that the early-middle Cretaceous fish localities in Mexico show high apparent endemism. This work confirms such observation, which also applies to the Albian-early Cenomanian localities in North America's northern, central, and southern regions. That is, except for the record of Belenostomus (in the Albian deposits of Hay River, Kiowa Formation, and the Sumidero Tlayúa quarries), there is no other genus common between the Albian sites of Canada, Albian-early Cenomanian of Mexico, and Albian Cenomanian boundary, and early Cenomanian of Kansas and Utah. In this scenario, the other three taxa reported here from the Sumidero Quarry are important because of their geographical and temporal distribution to other localities (Notagogus and Teoichthys are also present the middle-late Albian of Tlayúa, and Thorectichthys in the early Cenomanian of El Chango), complementing the biogeographic patterns in these fishes into the Paleo-Gulf of Mexico region. The other genera present in more than one locality in this southern North American region include Unamichthys (in Tlayúa and Muhi), Armigatus (in Tlayúa and El Chango), Saurorhamphus (in the El Espinal and El Chango quarries), Heckelichthys (in Muhi and El Chango), and Paraclupea and Macrosemiocotzus (in Tlayúa and El Espinal). The other genera already described in the Paleo-Gulf region of Mexico are endemic because these are unknown in more than one locality.
Three crucial factors help to explain the geographical-temporal pattern of the Albian-early Cenomanian bony fishes from North America described above and the remarkably high generic diversity observed in Mexico. There is a strong Lagerstätten effect in favor of Mexico because the three North American Lagerstätten -type localities (Tlayúa, Muhi, and El Chango quarries) known for Albian-early Cenomanian range are concentrated in this country (Figure 9D). Since the Cretaceous, the latitudinal position of North America has not changed significantly, so there is also a robust latitudinal effect directly linked to the climate that is also in favor of Mexico; all the Albian-early Cenomanian localities of Mexico are below 30° N and within the tropical region, where high biodiversity is common (Figure 9E).
The relatively high diversity of bony fishes of the Albian-early Cenomanian of Mexico may have been triggered by the interconnectivity effect of the North American sea bodies (Figure 9F). During the Albian period, the north-central region of North America, between Canada and the USA, was occupied by the Mowry Sea, which connected only to the Boreal Sea. In the Cenomanian, this intercontinental sea opened to the south, becoming the Western Interior Seaway of North America. The history of Mexico-adjacent seas is contrastingly more active. The Gulf of Mexico originated at the end of the Jurassic, while the Tethys Sea made its way between North and South America through the Hispanic Corridor; thus, Mexico housed marine taxa in the epicontinental regions of the Paleo-Gulf of Mexico, whose descendants survived in Mexican sites of the Cretaceous. The west opening of the Hispanic Corridor also made it possible for taxa from the Paleo-Pacific Ocean to access Mexico. During the early Cretaceous, while North America, North Africa, and South America were still nearby, Mexico also received fish from the Paleo-Atlantic regions of Africa and South America (Maisey, 2000). Finally, the opening of the North American Inland Sea toward the Paleo-Gulf of Mexico, at least since the late Albian (Blakey, 2014; Slattery et al., 2015; Eldrett et al., 2017), allowed the exchange of marine fauna between these marine regions.
In the scenario described above, all the localities of Canada and the USA considered here naturally include exclusive taxa of the Boreal and Mowry seas, except for the presence of Belonostomus and probably Lepidotes, which is a genus of wide distribution that appeared in the Jurassic (Figure 9G). On the contrary, the Albian-early Cenomanian localities of the Paleo-Gulf of Mexico region share a significative number of genera present in the other areas; this has taxa with cosmopolitan distribution (Belonostomus, Megalops, Enchodus, and Pachyrhizodus), share genera with the eastern Tethys Sea (e.g., Heckelicthys, Armigatus, Saurorhamphus; Scombroclupea; Thorectichthys, and Notagogus), the epicontinetal seas of the Early Cretaceous South America (e.g., Neoproscinetes, Araripichthys, Axelrodichthys, and Brannerion). Finally, all the taxa described in the localities of Mexico, without a wider distribution, form the center of a typical association of the Paleo-Gulf of Mexico (e.g., Unamichthys, Michin, and Macrosemiocotzus).
The Colección Nacional de Paleontología (Igl-UNAM) houses nearly 30 additional fossil fish specimens from the Sumidero Quarry. Given their preservation mode, the entire account for the diversity of fishes from this site requires considerably more preparation time. Thus, the contributions of this locality to understanding the diversity of North American bony fishes will surely increase in the coming future.
CONCLUSION
This paper provides a general report on the fossil fish found at the Sumidero Quarry. This site is the second Albian fish site in Mexico. The lithostratigraphic position within the Sierra Madre Group and the affinities of its microfossils and fishes, which include specimens of Notagogus novomundi and Teoichthys brevipina previously described from specimens of the Tlayúa Quarry, it is determined that the Sumidero Quarry is of Albian age. This finding provides valuable information that complements the knowledge of the Albian/Cenomanian ichthyofauna of North America and its contribution to the Cenomanian Diversification Event of Bony Fishes. Like other Albian-early Cenomanian fossil fish sites of Mexico, the fish assemblage of this site is eclectic because it contains taxa with cosmopolitan distribution (Belonostomus), genera with Tethys Sea affinities (Thorecthichthys), and taxa exclusive of the Paleo-Gulf of Mexico region (Notagogus and Teoichthys).
ACKNOWLEDGEMENTS
We extend our sincere gratitude to the local workers of the Sumidero Quarry for their invaluable support and friendship during fieldwork seasons since 2008. This manuscript has been enhanced by the insightful observations of L.E. Arrubarrena, M. Everhart, J.V. Andrews, and D. Grossnickle. J.M. Contreras took the photographs for this work. G. Alvarez Reyes assisted us with the preparation of the fossils referenced in this research. Financial support for this project was provided by UNAM through grants DGAPA-PAPIIT IN225008 and IN115223. Additionally, the authors SPO, BATM, NLS, and BRJCR were awarded postgraduate scholarships by the Consejo Nacional de Ciencias, Humanidades y Tecnología (CONACHYT, Mexico).
REFERENCES
Agassiz, L. 1833. Recherches sur les poisons fossils, volume 1, 188 p. (preface dated in 1833), Neuchâtel.
Agassiz. L. 1834a. Abgerissene Bemerkungen über fossile Fische. Neues Jahrbuch für Mineralogie, Geognosie, Geologie und Petrefaktenkunde 1834:379-390.
Agassiz, L. 1834b. Recherches sur les poissons fossiles, volume 2 (336 p. + 75 tabs in the supplementary Atlas), Neuchâtel.
Alvarado-Ortega, J. 2004. Description and relationships of a new ichthyodectiform fish from the Tlayúa Formation (Early Cretaceous: Albian), Puebla, Mexico. Journal of Vertebrate Paleontology, 24:802-813.
https://doi.org/10.1671/0272-4634(2004)024[0802:DAROAN]2.0.CO;2
Alvarado-Ortega, J. 2005. Sistemática de los peces Ichthyodectiformes de la cantera Tlayúa, Puebla, México. Unpublished PhD Thesis, Universidad Nacional Autónoma de México, Distrito Federal, México.
Alvarado-Ortega, J. 2014. Ancient herring from the Tlayúa Quarry (Cretaceous, Albian) near Tepexi de Rodíguez, Puebla State, central Mexico, closing the gap in the early diversification of Clupeomorpha. Cretaceous Research, 50:171-186.
https: //doi.org/10.1016/j.cretres.2014.02.022
Alvarado-Ortega, J. and Alves, Y.M. 2022. Nusaviichthys nerivelai gen. et sp. nov., an Albian crossognathiform fish from the Tlayúa Lagerstätte, Mexico. Cretaceous Research, 135:105189.
https://doi.org/10.1016/j.cretres.2022.105189
Alvarado-Ortega, J. and Brito, P.M. 2011. A new species of Araripichthys (Teleostei, Elopocephala) from the Tlayúa Formation (Cretaceous, Albian), Mexico. Journal of Vertebrate Paleontology, 31:1376-1381.
https://www.jstor.org/stable/41407692
Alvarado-Ortega, J. and Espinosa-Arrubarrena, L. 2008. A new genus of ionoscopiform fish (Halecomorphi) from the Lower Cretaceous (Albian) lithographic limestones of the Tlayúa quarry, Puebla, México. Journal of Paleontology, 82:163-175.
https://doi.org/10.1666/04-152.1
Alvarado-Ortega, J. and Melgarejo-Damián, M.P. 2017. Paraclupea seilacheri sp. nov., a double armored herring (Clupeomorpha, Ellimmichthyiformes) from the Albian limestones of Tlayúa quarry, Puebla, Mexico. Revista Mexicana de Ciencias Geológicas, 34:234-249.
Alvarado-Ortega, J. and Ovalles-Damián, E. 2008. Triplomystus applegatei, sp. nov. (Teleostei: Ellimmichthyiformes), a rare “triple armored herring” from El Espinal Quarry (Early Cretaceous), Chiapas, southeastern Mexico. Journal of Vertebrate Paleontology, 28:53-60.
https://doi.org/10.1671/0272-4634(2008)28[53:TASNTE]2.0.CO;2
Alvarado-Ortega, J. and Than-Marchese, B.A. 2012. A Cenomanian aipichthyoid fish (Teleostei, Acanthomorpha) from America, Zoqueichthys carolinae gen. and sp. nov. from El Chango quarry (Cintalapa Member, Sierra Madre Formation), Chiapas, Mexico. Revista Mexicana de Ciencias Geológicas, 29:735-748.
Alvarado-Ortega, J. and Than-Marchese, B.A. 2013. The first record of a North American Cenomanian Trachichthyidae fish (Acanthomorpha, Acanthopterygii), Pepemkay maya, gen. et sp. nov., from El Chango quarry (Sierra Madre Formation), Chiapas, Mexico. Journal of Vertebrate Paleontology, 33:48-57.
https://doi.org/10.1080/02724634.2012.712585
Alvarado-Ortega, J., González-Rodríguez, K.A., Blanco-Piñón, A., Espinosa-Arrubarrena, L., and Ovalles-Damián, E. 2006. Mesozoic osteichthyans of Mexico, p. 169-207. In Vega, F.J., Nyborg, T., Perrilliat, M.C., Montellano-Ballesteros, M., Cevallos-Ferriz, S.R.S., and Quiroz-Barroso, S.A. (eds.), Studies on Mexican Paleontology. Studies on Mexican Paleontology, Topics on Geobiology, vol. 24. Springer, Dordrecht, The Netherland.
https://doi.org/10.1007/1-4020-3985-9_9
Alvarado-Ortega, J., Espinosa-Arrubarrena, L., Blanco, A., Vega, F.J., Benammi, M., and Briggs, D.E. 2007. Exceptional preservation of soft tissues in Cretaceous fishes from the Tlayúa Quarry, central Mexico. Palaios, 22:682-685.
Alvarado-Ortega, J., Mayrinck, D., and Brito, P.M. 2008. A basal pachyrhizodontid fish (Actinopterygii, Teleostei) from the Lower Cretaceous of the Tlayúa Quarry, central Mexico. Comptes Rendus Palevol, 7:269-275.
https://doi.org/10.1016/j.crpv.2008.03.006
Alvarado-Ortega, J., Ovalles-Damián, E., and Arratia, G. 2008. A review of the interrelationships of the order Ellimmichthyiformes (Teleostei: Clupeomorpha). p. 257-278. In Arratia, G., Schultze H.-P., and Wilson, M.V.H. (eds.), Mesozoic Fishes IV -Homology and Phylogeny-. Verlag Dr. Friedrich Pfiel, München, Germany.
Alvarado-Ortega, J., Ovalles-Damián, E., and Blanco-Piñón, A. 2009. The fossil fishes from the Sierra Madre Formation, Ocozocoautla, Chiapas, Southern Mexico. Palaeontologia Electronica, 12.2.4A:1-22.
https://palaeo-electronica.org/2009_2/168/index.html
Alvarado-Ortega, J., Cuevas-García, M., Melgarejo-Damián, M.P., Cantalice, K.M., Alaniz-Galvan, A., Solano-Templos, G., and Than-Marchese, B.A. 2015. Paleocene fishes from Palenque, Chiapas, southeastern Mexico. Palaeontologia Electronica 18.2.39A:1-22.
https://doi.org/10.26879/536
Alvarado-Ortega, J., Cuevas-García, M., and Cantalice, K. 2018. The fossil fishes of the archaeological site of Palenque, Chiapas, southeastern Mexico. Journal of Archaeological Science: Reports, 17:462-476.
https://doi.org/10.1016/j.jasrep.2017.11.029
Alvarado-Ortega, J., Cantalice, K.M., Martínez-Melo, A., García-Barrera, P., Than-Marchese, B.A., Díaz-Cruz, J.A., and Barrientos-Lara, J.I. 2020. Tzimol, a Campanian marine paleontological site of the Angostura Formation near Comitán, Chiapas, southeastern Mexico. Cretaceous Research, 107:104279.
https://doi.org/10.1016/j.cretres.2019.104279
Alvarado-Ortega, J., Than-Marchese, B.A., and Melgarejo-Damián, M.P. 2020. On the Albian occurrence of Armigatus (Teleostei, Clupeomorpha) in America, a new species from the Tlayúa Lagerstätte, Mexico. Palaeontologia Electronica, 23:a52.
https://doi.org/10.26879/1107
Alves, Y.M., Alvarado-Ortega, J., and Brito, P.M. 2020. † Epaelops martinezi gen. and sp. nov. from the Albian limestone deposits of the Tlayúa quarry, Mexico -A new late Mesozoic record of Elopiformes of the western Tethys. Cretaceous Research, 110:104260.
https://doi.org/10.1016/j.cretres.2019.104260
Amaral, C. R., Alvarado-Ortega, J., and Brito, P.M. 2013. Sapperichthys gen. nov., a new gonorynchid from the Cenomanian of Chiapas, Mexico, p. 305-323. In Arratia, G., Schultze, H.-P., and Wilson, M.V.H. (eds.), Mesozoic Fishes V -Global Diversity and Evolution-. Verlag Dr. Friedrich Pfiel, München, Germany.
Applegate, S. P. 1988. A new genus and species of a holostean belonging to the family Ophiopsidae, Teoichthys kallistos, from the Cretaceous, near Tepexi de Rodríguez, Puebla. Revista Mexicana de Ciencias Geológicas, 7:200-205.
Applegate, S.P. 1992. A new genus and species of pycnodont from the Cretaceous (Albian) of central Mexico, Tepexi de Rodríguez, Puebla. Revista Mexicana de Ciencias Geológicas, 10:64-178.
Applegate, S.P. 1996. An overview of the Cretaceous fishes of the quarries near Tepexi de Rodríguez, Puebla, México, p. 529-538. In Arratia, G. and Viohl, G. (eds.), Mesozoic Fishes -Systematics and Paleoecology-. Verlag, Dr. Friedrich Pfeil. München, Germany.
Applegate, S.P. 2001. The Pycnodont fauna from the Lower Cretaceous of Tepexi de Rodríguez (Puebla, México), p. 3. In Tintori, A. (ed.), Abstract book, III International Meeting on Mesozoic Fishes -Systematics, Paleoenvironments and Biodiversity-. University of Milan, Serpiano-Monte San Giorgio. Italy.
Applegate, S.P. and Espinosa-Arrubarrena, L. 1982. Lithographic limestone-like deposits in Tepexi de Rodríguez, Puebla. Field guide for the excursion, Vertebrate Paleontology, Annual Meeting 42. Instituto de Geología and Society of Vertebrate Paleontology, Mexico.
Applegate, S.P., Espinosa-Arrubarrena, L., Alvarado-Ortega, J., and Benammi, M., 2006. Revision of recent investigations in the Tlayúa Quarry, p. 275-304. In Vega, F.J., Nyborg, T., Perrilliat, M.-C., Montellano-Ballesteros, M., Cevallos-Ferriz, S.R.S., and Quiroz-Barroso, S.A. (eds.), Studies on Mexican Paleontology. Topics on Geobiology, vol. 24. Springer, Netherlands.
https://doi.org/10.1007/1-4020-3985-9_9
Arambourg, C. 1950. Nouvelles observations sur les Halecostomes et Torigine des Clupeidae. Comptes Rendus de l'Académie des Sciences, 231:416-418.
Arratia, G. 2015. Los peces osteíctios fósiles de Chile y su importancia en los contextos paleobiogeográfico y evolutivo, p. 31-79. In Rubilar-Rogers, D., Otero, R., Vargas, A., and Sallaberry, M. (eds.), Vertebrados fósiles de Chile. Publicación Ocasional, 63. Museo Nacional de Historia Natural, C hile.
Arratia, G. and Schultze, H.-P. 2012. The macrosemiiform fish companion of the Late Jurassic theropod Juravenator from Schamhaupten, Bavaria, Germany. Fossil Record, 15(1):5-25.
https://doi.org/10.1002/mmng.201200001
Arratia, G., González-Rodríguez, K.A., and Hernández-Guerrero, C. 2018. A new pachyrhizodontid fish (Actinopterygii, Teleostei) from the Muhi Quarry (Albian-Cenomanian), Hidalgo, Mexico. Fossil Record, 21:93-107.
https://doi.org/10.5194/fr-21-93-2018, 2018
Arratia, G. and González-Rodríguez, K.A. 2024. A New Intriguing Teleost from the Albian Muhi Quarry, Central Mexico, and Early Euteleostean Diversification. Diversity, 2024,16:414.
https:// doi.org/10.3390/d16070414
Baños-Rodríguez, R.E., González-Rodríguez, K.A., Wilson, M.V., and González-Martínez, J.A. 2020. A new species of Heckelichthys fr om the Muhi Quarry (Albian-Cenomanian) of central Mexico. Cretaceous Research, 110:104415.
https://doi.org/10.1016/j.cretres.2020.104415
Bartholomai, A. 2004. The large aspidorhynchid fish, Richmondichthys sweeti (Etheridge Jnr and Smith Woodward, 1891) from Albian marine deposits of Queensland, Australia. Memoirs of the Queensland Museum, 49:521-536.
Bartram, A.W.H. 1977. The Macrosemiidae, a Mesozoic family of holostean fishes. Bulletin of the British Museum (Natural History) Geology series, 29:137-234.
Beamon, J.C. 1999. Depositional environment and fossil biota of a thin clastic unit of the Kiowa Formation, Lower Cretaceous (Albian), McPherson County, Kansas. Unpublished Masters Thesis, Fort Hays State University, Hays, Kansas, U.S.A.
Berg, L.S. 1937. A classification of fish-like vertebrates. Izvestiya Akademii Nauk USSR, Seriya Biologischeskaya, 4:1277-1280.
Blakey, R.C. 2014. Paleogeography and paleotectonics of the Western Interior Seaway, Jurassic-Cretaceous of North America. AAPG Search and Discovery, Article 30392, p. 72.
Bleeker, P. 1859. Enumeratio specierum piscium hucusque in Archipelago Indico observatarum, adjectis habitationibus citationibusque, ubidescriptiones earum recentiores reperiuntur, nec non speciebus Musei Bleekeriani Bengalensibus, Japonicis, Capensibus Tasmanicisque. Acta Societatis Regiae Scientiarum Indo-Neêrlandicae, 6:1-276.
Blot, J. 1987. L’Ordre des Pycnodontiformes. Museo Civico di Storia Naturale di Verona, Studi e ricerche sui giacimenti terziari di Bolca, 5:1-211.
Bogan, S., Taverne, L., and Agnolin, F.L. 2011. Description of a new aspidorhynchid fish, Belonostomus lamarquensis sp. nov.(Halecostomi, Aspidorhynchiformes), from the continental Upper Cretaceous of Patagonia, Argentina. Bulletin de l'Institut royal des Sciences naturelles de Belgique, Sciences de la Terre, 81:235-245.
Bravo-Cuevas, V.M., González-Rodríguez, K.A., Baños-Rodríguez, R., and Hernández-Guerrero, C. 2012. The Muhi Quarry: A Fossil-Lagerstätte from the Mid-Cretaceous (Albian-Cenomanian) of Hidalgo, Central México, p. 107-122. In Elitok, Ö. (ed.), Stratigraphic Analysis of Layered Deposits. IntechOpen, Rijeka, Croatia.
Bravi, S. 1994. New observations on the Lower Cretaceous fish Notagogus pentlandi Agassiz (Actinopterygii, Halecostomi, Macrosemiidae). Bollettino della Società Paleontologica Italiana, 33(1):51-70.
Brito, P.M. 1997. Révision des Aspidorhynchidae (Pisces, Actinopterygii) du Mésozoïque: ostéologie, relations phylogénétiques, données environnementales et biogéographiques. Geodiversitas, 19:681-772.
Brito, P.M. 2000. A new halecomorph with two dorsal fins, Placidichthys bidorsalis ng, n. sp. (Actinopterygii: Halecomorphi) from the Lower Cretaceous of the Araripe Basin, northeast Brazil. Comptes Rendus de l'Académie des Sciences-Series IIA-Earth and Planetary Science, 331:749-754.
https://doi.org/10.1016/S1251-8050(00)01477-4
Brito, P.M. and Alvarado-Ortega, J. 2008. A new species of Placidichthys (Halecomorphi: Ionoscopiformes) from the Lower Cretaceous Marizal Formation, northeastern Brazil, with a review of the biogeographical distribution of the Ophiopsidae. Geological Society, London, Special Publications, 295:145-154.
https://doi.org/10.1144/SP295.9
Brito, P.M. and Alvarado-Ortega, J. 2013. Cipactlichthys scutatus, gen. nov., sp. nov. a new halecomorph (Neopterygii, Holostei) from the Lower Cretaceous Tlayúa Formation of Mexico. PLoS One, 8(9):e73551.
https://doi.org/10.1371/journal.pone.0073551
Brotzen, F. 1942. Die Foraminiferengattung Gavelinella nov. gen. und die Systematik der Rotaliiformes. Sveriges Geologiska Undersökning, Ser. C, Avhandlingar och uppsatser. Årsbok, 36:1-60.
Cantalice, K.M. and Alvarado-Ortega, J. 2016. Eekaulostomus cuevasae gen. and sp. nov., an ancient armored trumpetfish (Aulostomoidea) from Danian (Paleocene) marine deposits of Belisario Domínguez, Chiapas, southeastern Mexico. Palaeontologia Electronica 19.3.53A:1-24.
https://doi.org/10.26879/682
Cantalice, K.M., Alvarado-Ortega, J., and Alaniz-Galvan, A. 2018. Paleoserranus lakamhae gen. et sp. nov., a Paleocene seabass (Perciformes: Serranidae) from Palenque, Chiapas, southeastern Mexico. Journal of South American Earth Sciences, 83:137-146.
https://doi.org/10.1016/j.jsames.2018.01.010
Cantalice, K.M., Alvarado-Ortega, J., and Bellwood, D.R. 2020. † Chaychanus gonzalezorum g en. et sp. nov.: A damselfish fossil (Percomorphaceae; Pomacentridae), from the Early Paleocene outcrop of Chiapas, Southeastern Mexico. Journal of South American Earth Sciences, 98:102322.
https://doi.org/10.1016/j.jsames.2019.102322
Cantalice, K.M., Than‐Marchese, B.A., and Villalobos‐Segura, E. 2021. A new Cenomanian acanthomorph fish from the El Chango quarry (Chiapas, south‐eastern Mexico) and its implications for the early diversification and evolutionary trends of acanthopterygians. Papers in Palaeontology, 7:1699-1726.
https://doi.org/10.1002/spp2.1359
Castro-Mora, J., Schlaepfer, C.J., and Martínez-Rodríguez, E. 1975. Estratigrafía y Microfacies del Mesozoico de la Sierra Madre del sur de Chiapas. Boletín de la Asociación Mexicana de Geólogos Petroleros, 27:1-103.
Cawley, J.J. and Kriwet, J. 2024. The Fossil Record and Diversity of Pycnodontiform Fishes in Non-Marine Environments. Diversity, 16:225.
https://doi.org/10.3390/d16040225
Cope, E.D. 1887. Zittel’s manual of palaeontology. American Naturalist, 21:1014-1019.
Cope, E.D. 1894. New and little known Paleozoic and Mesozoic fishes. Journal of the academy of Natural Sciences of Philadelphia, Series 2, 9:427-448.
Costa, O.G. 1857. Su di un nuovo genere di pesce fossile. Memorie della Reale Accademia delle Scienze, Napoli, 2:235-236.
Cragin, F.W. 1894. Vertebrata from the Neocomian of Kansas. Colorado College Studies, 5:69-73 + pl. I-II.
Cuevas-García, M. and Alvarado-Ortega, J. 2009. Estudio arqueológico y paleontológico de los fósiles marinos que proceden del sitio de Palenque, Chiapas. Informe de la primera temporada de campo 2008. Reporte Inédito, Instituto Nacional de Antropología e Historia, México.
Cushman, J.A. 1926. Foraminifera of the genera Siphogenerina and Pavonina. Proceedings of the United States National Museum, 67(25):1-24+6 plts.
Cushman, J. A. 1933. Some new foraminiferal genera. Contributions from the Cushman Laboratory for Foraminiferal Research, 9(2):32-38.
Defrance J.L.M. 1924. Mineralogie et Geologie, vol. 30-33. In Cuvier, F. (ed.), Dictionnaire des Sciences Naturelles dans lequel on traite méthodiquement des différents êtres de la nature (par plusieurs professeurs du Jardin du Roi), Paris.
Díaz-Cruz, J.A., Alvarado-Ortega, J., and Carbot-Chanona, G. 2016. The Cenomanian short snout enchodontid fishes (Aulopifomes, Enchodontidae) from Sierra Madre Formation, Chiapas, southeastern Mexico. Cretaceous Research, 61:136-150.
https://doi.org/10.1016/j.cretres.2015.12.026
Díaz-Cruz, J.A., Alvarado-Ortega J., and Carbot-Chanona, G. 2019a. Dagon avendanoi gen. and sp. nov., an Early Cenomanian Enchodontidae (Aulopiformes) fish from the El Chango quarry, Chiapas, southeastern Mexico. Journal of South American Earth Sciences, 91:272-284.
https://doi.org/doi:10.1016/j.jsames.2019.01.014
Díaz-Cruz, J.A., Alvarado-Ortega J., and Carbot-Chanona, G., 2019b. Corrigendum to "Dagon avendanoi gen. and sp. nov., an Early Cenomanian Enchodontidae (Aulopiformes) fish from the El Chango quarry, Chiapas, southeastern Mexico [J. South Am. Earth Sci. 91 (2019) 272-284]". Journal of South American Earth Sciences, 95:102314.
https://doi.org/10.1016/j.jsames.2019.102314
Díaz-Cruz, J.A., Alvarado-Ortega, J., and Giles, S. 2020. A long snout enchodontid fish (Aulopiformes: Enchodontidae) from the Early Cretaceous deposits at the El Chango quarry, Chiapas, southeastern Mexico: A multi-approach study. Palaeontologia Electronica, 23(2):a30.
https://doi.org/10.26879/1065
Dixon, F. 1850. Geology and Fossils of the Tertiary and Cretaceous Formations of Sussex. Longman, Brown, Green & Longmans, London.
d´Orbigny, A. D. 1826. Tableau méthodique de la classe des Céphalopodes. Annales des Sciences Naturelles, 7(96-169):245-314.
Dunkle, D. 1971. Notes on an addition to the fish fauna of the Mowry Shale (Cretaceous) of Wyoming. Kirtlandia, 14:1-8.
Eastman, C.R. 1917. Fossil fishes in the collection of the United States National Museum. Proceedings of the United States National Museum, 52:235-304.
Ebert, M. 2014. The genus Belonostomus Agassiz, 1834 (Neopterygii, Aspidorhynchiformes) in the late Jurassic of the Solnhofen Archipelago, with a focus on Belonostomus kochii Münster, 1836 from Ettling (Germany). Archaeopteryx, 32:15-43.
Ebert, M. 2016. The Pycnodontidae (Actinopterygii) in the Late Jurassic: 2) Turboscinetes gen. nov. in the Solnhofen Archipelago (Germany) and Cerin (France). Archaeoptery, 33:12-53.
Ebert, M. 2018. Cerinichthys koelblae, gen. et sp. nov., from the Upper Jurassic of Cerin, France, and its phylogenetic setting, leading to a reassessment of the phylogenetic relationships of Halecomorphi (Actinopterygii). Journal of Vertebrate Paleontology, 38:e1420071.
https://doi.org/10.1080/02724634.2017.1420071
Ebert, M. 2019. Zandtfuro and Schernfeldfuro, new genera of Halecomorphi (Actinopterygii) from the Upper Jurassic Solnhofen-Archipelago. Journal of Vertebrate Paleontology, 39:e1592759.
https://doi.org/10.1080/02724634.2019.1592759
Ebert, M. 2020. Elongofuro and Altmuehlfuro, new genera of Halecomorphi (Neopterygii) from the Upper Jurassic of the Solnhofen Archipelago and Nusplingen (Germany). Neues Jahrbuch für Geologie und Paläontologie-Abhandlungen, 295:185-210.
https://doi.org/10.1127/njgpa/2020/0883
Ebert, M. 2021. A new Ophiopsiformes (Halecomorphi, Neopterygii) from the Upper Jurassic of Nusplingen (Germany) and a comparison of Kimmeridgian Ophiopsiformes from Nusplingen with Tithonian taxa from the Solnhofen Archipelago. Neues Jahrbuch für Geologie und Paläontologie-Abhandlungen, 302:299-329.
https://doi.org/10.1127/njgpa/2021/1033
Eguiluz de Antuñano, S., Andrés Than-Marchese, B.A., Camacho Ortegón, L.F., López-Martínez, R., and Enrique Gómez-Pérez, L. 2023. Estratigrafía del Grupo Sierra Madre en Chiapas y su hipotético potencial petrolero. Boletín de la Sociedad Geológica Mexicana, 75:A301023.
https://doi.org/10.18268/ BSGM2023v75n3a301023
Eldrett, J.S., Dodsworth, P., Bergman, S.C., Wright, M., and Minisini, D. 2017. Water-mass evolution in the Cretaceous Western Interior Seaway of North America and equatorial Atlantic. Climate of the Past, 13(7);855-878.
https://doi.org/10.5194/cp-13-855-2017
Espinosa-Arrubarrena, L., Applegate, S.P., and González-Rodríguez, K.A. 1996. The First Mexican Record of a Coelacanth (Osteichthyes: Sarcopterygii) from the Tlayua Quarries Near Tepexi De Rodriguez, Puebla, with a Discussion on the Importance of this Fossil. The Paleontological Society Special Publications, 8:116-116.
Espinosa-Arrubarrena, L. and Alvarado-Ortega, J. 2010. Field trip to the Tlayúa Quarry, p. 89-113, In González-Rodríguez, K. A. and Arratia, G. (eds.), Abstract Book and Field Guides, Fifth International Meeting on Mesozoic Fishes -Global Diversity and Evolution-. Ciencia al Día 19, Universidad Autónoma del Estado de Hidalgo, Pachuca de Soto, Mexico.
Everhart, M.J. 2004. First record of the hybodont shark genus, “ Polyacrodus ” sp., (Chondrichthyes; Polyacrodontidae) from the Kiowa Formation (Lower Cretaceous) of McPherson County, Kansas. Kansas Academy of Science, Transactions 107(1-2):39-43.
https://doi.org/10.1660/0022-8443(2004)107[0083:FROTHS]2.0.CO;2
Everhart, M.J. 2009. First Occurrence of Marine Vertebrates in the Early Cretaceous of Kansas: Champion Shell Bed, Basal Kiowa Formation. Transactions of the Kansas Academy of Science, 112:201-210.
https://doi.org/10.1660/062.112.0408
Everhart, M.J. 2011. Occurrence of the hybodont shark genus Meristodonoides (Chondrichthyes; Hybodontiformes) in the Cretaceous of Kansas. Kansas Academy of Science, Transactions, 114(1-2):33-46.
https://doi.org/10.1660/062.114.0103
Fielitz, C. and González-Rodríguez, K. 2008. A new species of Ichthyotringa from the El Doctor Formation (Cretaceous), Hidalgo, Mexico, p. 373-388. In Arratia, G., Schultze, H.-P., and Wilson M.V.H. (eds.), Mesozoic Fishes 4 -Homology and Phylogeny-. Verlag Dr. Friedrich Pfeil, München, Germany.
Fielitz, C., and González-Rodríguez. K. A. 2010. A new species of Enchodus (Aulopiformes: Enchodontidae) from the Cretaceous (Albian to Cenomanian) of Zimapán, Hidalgo, México. Journal of Vertebrate Paleontology, 30:1343-1351.
https://doi.org/10.1080/02724634.2010.501438
Figueiredo, F.J. and Silva Santos, R. 1987. Considerações taxinômicas dos picnodontidos da Formaçaõ Gramame (camada de Fostato), Pe. Anais X congresso brasileiro de Paleontologia, Rio de Janeiro, p. 25-31.
Forey, P.L. 1975. A fossil clupeomorph fish from the Albian of the Northwest Territories of Canada, with notes on cladistic relationships of clupeomorphs. Journal of Zoology, 175:151-177.
https://doi.org/10.1111/j.1469-7998.1975.tb01394.x
Friedman, M., Andrews J.V., Saad, H., El-Sayed, S. 2023. The Cretaceous-Paleogene transition in spiny-rayed fishes: surveying “Patterson’s Gap” in the acanthomorph skeletal record. Geologica Belgica, 26:1-23.
https://doi.org/10.20341/gb.2023.002
Giersch, S. 2014. Die Knochenfische der Oberkreidezeit in Nordost-Mexiko: Beschreibung, Systematik, Vergesellschaftung, Paläoökologie und Paläobiogeographie. Unpublished PhD Thesis. Ruprecht-Karl-University Heidelberg, Heidelberg, Germany.
Giersch, S., Frey, E., Stinnesbeck, W., and Gonzalez Gonzalez, A.H. 2008. Fossil fish assemblages of northeastern Mexico: New evidence of Mid Cretaceous actinopterygian radiation, p. 43-45. In Krempaskà, Z (ed.), Volume of abstracts, 6th Meeting of the European Association of Vertebrate Palaeontologists, Spiŝská Nová Ves, Slovak Republic.
González-Rodríguez, K.A., Applegate, S.P., and Espinosa-Arrubarrena, L. 2004. A New World macrosemiid (Pisces: Neopterygii—Halecostomi) from the Albian of Mexico. Journal of Vertebrate Paleontology, 24:281-289.
https://doi.org/10.1671/1862
González-Rodríguez, K.A. and Reynoso, V.H. 2004. A new Notagogus (Macrosemiidae, Halecostomi) species from the Albian Tlayúa Quarry, Central Mexico, p. 265-278. In Arratia, G. and Tintori, A. (eds.), Mesozoic Fishes 3 -Systematics, Paleoenvironments and Biodiversity-. Verlag Dr. Friedrich Pfeil, München, Germany.
González-Rodríguez, K.A. and Fielitz, C. 2008. A new species of acanthomorph fish from the Upper Cretaceous Muhi Quarry, Hidalgo, Central Mexico, p. 399-411. In Arratia, G., Schultze, H.-P., and Wilson M. V. H. (eds.), Mesozoic fishes 4 -Homology and Phylogeny-. Verlag Dr. Friedrich Pfeil, München, Germany.
González-Rodríguez, K.A., Applegate, S.P., and Espinosa-Arrubarrena, L., and González-Barba, G. 2013a. An overview of the Mexican fossil fish record. p. 9-24. In Arratia, G., Schultze, H.-P., and Wilson M.V.H. (eds.), Mesozoic fishes 5 -Global Diversity and Evolution and Phylogeny-. Verlag Dr. Friedrich Pfeil, München, Germany.
González-Rodríguez, K.A., Schultze, H.-P., and Arratia, G. 2013b. Miniature armored acanthomorph teleosts from the Albian/Cenomanian (Cretaceous) of Mexico, p. 457-487. In Arratia, G., Schultze, H.-P., and Wilson M.V.H. (eds.), Mesozoic fishes 5 -Global Diversity and Evolution and Phylogeny-. Verlag Dr. Friedrich Pfeil, München, Germany.
González-Rodríguez, K.A., Fielitz, C., Bravo-Cuevas, V.M., and Baños-Rodríguez, R.E. 2016. Cretaceous osteichthyan fish assemblages from Mexico, p. 1-14. In Khosla, A. and Lucas, S.G. (eds.), Cretaceous Period: Biotic Diversity and Biogeography. New Mexico Museum of Natural History and Science Bulletin 71. Alburquerque, New Mexico.
Gouiric-Cavalli, S. 2015. Jonoichthys challwa gen. et sp. nov., a new Aspidorhynchiform (Osteichthyes, Neopterygii, Teleosteomorpha) from the marine Upper Jurassic sediments of Argentina, with comments about paleobiogeography of Jurassic aspidorhynchids. Comptes Rendus Palevol, 14:291-304.
https://doi.org/10.1016/j.crpv.2015.03.007
Grande, L. 1982. A revision of the fossil genus Diplomystus: with comments on the interrelationships of clupeomorph fishes. American Museum Novitates, 2728:1-34.
Grande, L. 1985. Recent and fossil Clupeomorph fishes with materials for revision of the subgroups of Clupeoids. Bulletin of the American Museum of Natural History, 181:231-372.
Grande, L. 2010. An empirical synthetic pattern study of gars (Lepisosteiformes) and closely related species, based mostly on skeletal anatomy. The resurrection of Holostei. American Society of Ichthyology and Herpetology, Copeia, 10(2A), (suppl), Special Publication 6:1-87 + x.
Grande, L. and Bemis, W. E. 1998. A comprehensive phylogenetic study of amiid fishes (Amiidae) based on comparative skeletal anatomy. An empirical search for interconnected patterns of natural history. Journal of Vertebrate Paleontology, 18 (suppl 1):1-696.
https://doi.org/10.1080/02724634.1998.10011114
Greenwood, P.H., Rosen, D.E., Weitzman, S.H., and Myers, G.S. 1966. Phyletic studies of teleostean fishes, with a provisional classification of living forms. Bulletin of the American Museum of Natural History, 131:139-456.
Guinot, G. and Cavin, L. 2016. ‘Fish’ (Actinopterygii and Elasmobranchii) diversification patterns through deep time. Biological Reviews, 91:950-981.
https://doi.org/10.1111/brv.12203
Heckel, J. 1850. Beiträge zur Kenntniss der fossilen Fische Osterreichs Abhandlungen I. Denkschriften des Kaiserlichen Akademie der Wissenschaften, Mathematisch-Naturwissenschaftliche Klasse 1:201-242+7 pls.
Hermus, C.R., Wilson, M.V., and Macrae, A. 2004. A new species of the Cretaceous teleostean fish Erichalcis from Arctic Canada, with a revised diagnosis of the genus., p. 449-462. In Arratia, G. and Tintori, A. (eds.), Mesozoic Fishes 3 -Systematics, Paleoenvironments and Biodiversity-. Verlag Dr. Friedrich Pfeil, München, Germany.
Hernández-Guerrero, C. 2015. Revisión taxonómica y análisis filogenético de la familia Pachyrhizodontidae en México. Unpublished Masters Thesis. Universidad Autónoma del Estado de Hidalgo, Pachuca, México.
Hernández-Guerrero, C., Cantalice, K.M., González-Rodríguez, K.A., and Bravo-Cuevas, V.M. 2021. The first record of a pterothrissin (Albuliformes, Albulidae) from the Muhi Quarry, mid-Cretaceous (Albian-Cenomanian) of Hidalgo, central Mexico. Journal of South American Earth Sciences, 107:103032.
https://doi.org/10.1016/j.jsames.2020.103032
Jordan, D.S. 1919. New genera of fossil fishes from Brazil. Proceedings of the Academy of Natural Sciences of Philadelphia, 71:208-210.
Jordan, D.S. and Gilbert, J.Z. 1919. Fossil Fishes of the Miocene (Monterey) Formations, p. 13-69. In Fossils of Southern California. Leland Stanford Junior University Publications, University Series.
Lacépède, B.G.E. 1803. Histoire naturelle des poisons, Tome 5. Chez Plassan, Imprimeur-Libraire. Paris.
Lane, J.A. and Ebert, M. 2015. A taxonomic reassessment of Ophiopsis (Halecomorphi, Ionoscopiformes), with a revision of Upper Jurassic species from the Solnhofen Archipelago, and a new genus of Ophiopsidae. Journal of Vertebrate Paleontology, 35:e883238.
https://doi.org/10.1080/02724634.2014.883238
López-Arbarello, A. and Alvarado-Ortega, J. 2011. New semionotiform (Neopterygii) from the Tlayúa Quarry (Early Cretaceous, Albian), Mexico. Zootaxa, 2749:1-24.
https://doi.org/10.11646/ZOOTAXA.2749.1.1
López-Arbarello, A., Bürgin, T., Furrer, H., and Stockar, R. 2019. Taxonomy and phylogeny of Eosemionotus Stolley, 1920 (Neopterygii: Ginglymodi) from the Middle Triassic of Europe. Palaeontologia Electronica, 22,1.10A:1-64.
https://doi.org/10.26879/904
López-Palomino, I., González-Rodríguez, K.A., Schultze, H.-P., Palma-Ramírez, A., and Contreras-Cruz, D. 2021. Ammonites from the La Negra Facies (El Doctor Formation, late Albian) of the Muhi Quarry, Hidalgo, central Mexico. Journal of South American Earth Sciences, 111:103400.
https://doi.org/10.1016/j.jsames.2021.103400
Machado, G.P., Alvarado-Ortega, J., Machado, L.P., and Brito, P.M. 2013. Teoichthys brevipina, sp. nov., a new ophiopsid fish (Halecomorphi, Ionoscopiformes) from the Lower Cretaceous Tlayúa Formation, Central Mexico. Journal of Vertebrate Paleontology, 33:482-487.
https://doi.org/10.1080/02724634.2013.729962
Maisey, J.G. 1986. Coelacanths from the Lower Cretaceous of Brazil. American Museum Novitates 2866:1-30.
Maisey, J.G. 2000. Continental break up and the distribution of fishes of Western Gondwana during the Early Cretaceous. Cretaceous Research, 21:281-314.
https://doi.org/10.1006/cres.1999.0195
Martha, S.O., Taylor, P.D., and Rader, W.L. 2019. Early Cretaceous cyclostome bryozoans from the early to middle Albian of the Glen Rose and Walnut formations of Texas, USA. Journal of Paleontology, 93(2):244-259.
Maxwell, E.E. and Caldwell, M.W. 2006. A new genus of ichthyosaur from the Lower Cretaceous of Western Canada. Palaeontology, 49:1043-1052.
https://doi.org/10. 1111/j.1475-4983.2006.00589.x
Moreno-Bedmar, J.A., Latil, J.L., Villanueva-Amadoz, U., Calvillo-Canadell, L., and Cevallos-Ferriz, S.R.S. 2014. Ammonite age-calibration of the El Chango fossil-lagerstätte, Chiapas state (SE Mexico). Journal of South American Earth Sciences, 56:447-453.
https://doi.org/10.1016/j.jsames.2014.09.022
Murray, A.M. and Cook, T.D. 2016. Overview of the Late Cretaceous fishes of the northern Western Interior Seaway, p. 255-261. In Khosla, A. and Lucas, S.G. (eds.), Cretaceous Period: Biotic Diversity and Biogeography. New Mexico Museum of Natural History and Science Bulletin 71. Alburquerque, New Mexico.
Murray A.M. and Wilson, M.V.H. 2009. A new Late Cretaceous macrosemiid fish (Neopterygii, Halecostomi) from Morocco, with temporal and geographical range extensions for the family. Palaeontology, 52:429-440.
https://doi.org/10.1111/j.1475-4983.2009.00851.x
Murray A.M. and Wilson, M.V.H. 2013. Two new paraclupeid fishes (Clupeomorpha: Ellimmichthyiformes) from the Upper Cretaceous of Morocco, p. 267-290. In Arratia, G., Schultze, H.-P., and Wilson M.V.H. (eds.), Mesozoic fishes 5 -Global Diversity and Evolution and Phylogeny-. Verlag Dr. Friedrich Pfeil, München, Germany.
Murray, A.M. and Wilson, M.V.H. 2014. Four new basal acanthomorph fishes from the Late Cretaceous of Morocco. Journal of Vertebrate Paleontology, 34:34-48.
https://doi.org/10.1080/02724634.2013.791693
Newbrey, M.G., Murray, A.M., Wilson, M.V.H., Brinkman, D.B., and Neuman, A.G. 2013. A new species of the paracanthopterygian Xenyllion (Sphenocephaliformes) from the Mowry Formation (Cenomanian) of Utah, USA, p. 363-384. In Arratia, G., Schultze, H.-P., and Wilson M. V. H. (eds.), Mesozoic fishes 5 -Global Diversity and Evolution and Phylogeny-. Verlag Dr. Friedrich Pfeil, München, Germany.
Nicholson, H.A. and Lydekker, R. 1889. A Manual of Palćontology for the Use of Students with a General Introduction on the Principles of Paleontology, Volume. 1. W. Blackwood and Sons. Edinburgh.
Nursall, J.R. 1996a. The phylogeny of pycnodont fishes, p. 125-152. In Arratia, G. and Viohl, G. (eds.), Mesozoic fishes -Systematics and Paleoecology-. Verlag Dr. F. Pfeil, München, Germany.
Nursall, J.R. 1996b, Distribution and ecology of pycnodont fishes, p. 115-124. In Arratia, G. and Viohl, G. (eds), Mesozoic fishes -Systematics and Paleoecology-. Verlag Dr. F. Pfeil, München, Germany.
Omara, S. 1956. New foraminifera from the Cenomanian of Sinai, Egypt. Journal of Paleontology, 30:883-890.
Pacheco-Ordaz, S., Mejía, O., and Alvarado-Ortega, J. 2025. Tahnaichthys magnuserrata gen. and sp. nov., a double-hump pycnodontid (Actinopterygii, Pycnodontiformes) fish from the Albian limestones of the Tlayúa Quarry, Puebla, Mexico. Journal of South American Earth Sciences, 152:105277.
https://doi.org/10.1016/j.jsames.2024.105277
Pantoja-Alor, J. 1992. Geología y paleoambiente de la cantera Tlayúa, Tepexi de Rodríguez, Estado de Puebla. Revista Mexicana deCiencias Geológicas, 9:156-169.
Pictet, F.-J. 1850. Description de quelques poissons fossiles du Mont Liban. Jules-Guillaume Fick, Genève.
Poyato-Ariza, F.J. 2020. Studies on pycnodont fishes (II): revision of the subfamily Pycnodontinae, with special reference to Italian forms. Rivista italiana di Paleontologia e Stratigrafia, 126:447-473.
https://doi.org/10.13130/2039-4942/13514
Poyato-Ariza, F.J. and Wenz, S. 2002. A new insight into pycnodontiform fishes. Geodiversitas, 24: 139-248.
Quezada-Muñetón, J.M. 1987. El Cretácico Medio-superior, y el límite Cretácico superior-Terciario Inferior en la Sierra de Chiapas. Boletín de la Asociación Mexicana de Geólogos Petroleros, 34:3-97.
Rabosky, D.L., Chang, J., Title, P.O., Cowman, P.F., Sallan, L., Friedman, M., Kaschner, K., Garilao, C., Near, T.J., Coll, M., and Alfaro, M.E. 2018. An inverse latitudinal gradient in speciation rate for marine fishes. Nature, 559:392-395.
https://doi.org/10.1038/s41586-018-0273-1
Russell, D.A. 1988. A check list of North American marine Cretaceous vertebrates including fresh water fishes. Tyrrell Museum Palaeontology, Occasional Paper 4:1-57.
Saint-Seine, P. 1949. Les Poissons des Calcaires lithographiques de Cerin (Ain). Nouvelles Archives du Muséum d'Histoire Naturelle de Lyon, Fascicule II:1-357.
Salas, G.P. 1949. El Cretácico de la Cuenca de Macuspana y su correlación. Boletín de la Sociedad Geológica Mexicana, 14:47-65.
Schaeffer, B. 1960. The Cretaceous holostean fish Macrepistius. American Museum Novitates, 2011:1-18.
Schultze, H.-P., Stewart, J.D., Neuner, A.M., and Coldiron, R.W. 1982. Type and figured specimens of fossil vertebrates in the collection of the University of Kansas Museum of Natural History. Part I. Fossil fishes. University Kansas Museum Natural History, Miscellaneous Publication, 73:1-53.
Scott, R.W. 1970. Paleoecology and paleontology of the Lower Cretaceous Kiowa Formation, Kansas. The University of Kansas Paleontological Contributions, 52:1-94.
Silva Santos, R. 1971. Nouveau genre et espèce d'Elopidae du bassin sédimentaire de la Chapada do Araripe. Anais da Academia Brasileira de Ciências, 43:439-442.
Slattery, J.S., Cobban, W.A., McKinney, K.C., Harries, P.J., and Sandness, A.L. 2015. Early Cretaceous to Paleocene paleogeography of the Western Interior Seaway: the interaction of eustasy and tectonism. Wyoming Geological Association Guidebook, 2015:22-60.
Steele, D.R. 1986. Contributions to the Stratigraphy of the Sierra Madre Limestone (Cretaceous) of Chiapas. Part 1. Physical stratigraphy and petrology of the Cretaceous Sierra Madre Limestone, west-central Chiapas. Universidad Nacional Autónoma México, Instituto de Geología, Boletin 102:1-101.
Steinmann, G. 1881. Die Foraminiferengattung Nummoloculina n.g.. Neues Jahrbuch für Mineralogie, Geologie und Paläontologie, 1:31-43.
Stewart, J.D. and Hakel, M. 2006. Ichthyofauna of the Mowrly shale (Early Cenomanian) Wyoming, p. 161-163. In Khosla, A. and Lucas, S.G. (eds.), Cretaceous Period: Biotic Diversity and Biogeography. New Mexico Museum of Natural History and Science Bulletin 71. Alburquerque, New Mexico.
Sun, A.-L. 1956. Paraclupea -A genus of double-armoured herrings from Chekiang. Acta Palaeontologica Sinica, 4:413-418.
Szabó, M., Gulyás, P., and Ősi, A. 2016. Late Cretaceous (Santonian) pycnodontid (Actinopterygii, Pycnodontidae) remains from the freshwater deposits of the Csehbánya Formation (Iharkút, Bakony Mountains, Hungary). Annales de Paléontologie: 102:123-134.
Taverne L. 1997. Les poissons crétacés de Nardò. 5°. Pycnodus nardoensis sp. nov. et considérations sur l’ostéologie du genre Pycnodus (Actinopterygii, Halecostomi, Pycnodontiformes). Bollettino del Museo Civico di Storia Naturale di Verona, 21:437-454.
Taverne, L. 2008. Considerations about the Late Cretaceous genus Chirocentrites and erection of the new genus Heckelichthys (Teleostei, Ichthyodectiformes)-A new visit inside the ichthyodectid phylogeny. Bulletin de l’Institut royal des Sciences naturelles de Belgique (Sciences de la Terre), 78:209-228.
Taverne, L. 2014. Osteology and phylogenetic relationships of Congophiopsis lepersonnei gen. nov. (Halecomorphi, Ionoscopiformes) from the Songa Limestones (Middle Jurassic, Stanleyville Formation), Democratic Republic of Congo. Geo-Eco-Trop, 38: 223-240.
Taverne, L. and Capasso, L. 2017. Italophiopsis derasmoi gen. and sp. nov. (Ionoscopiformes, Italophiopsidae fam. nov.) from the cretaceous of Pietraroja (Italy). Thalassia Salentina, 39:9-24.
Taverne, L. and Capasso, L. 2018. Osteology and relationships of Libanopycnodus wenzi gen. et sp. nov. and Sigmapycnodus giganteus gen. et sp. nov. (Pycnodontiformes) from the Late Cretaceous of Lebanon. European Journal of Taxonomy, 420:1-29.
https://doi.org/10.5852/ejt.2018.420
Taverne, L.P. and Ross, P.H. 1973. Fischreste aus dem ToÈck (Unter-Aptien) von Helgoland. Meyniana, 23:99-111.
Than-Marchese, B.A., Alvarado-Ortega, J., Matamoros, W.A., and Velázquez-Velázquez, E. 2020. Scombroclupea javieri sp. nov., an enigmatic Cenomanian clupeomorph fish (Teleostei, Clupeomorpha) from the marine deposits of the Cintalapa Formation, Ocozocoautla, Chiapas, southeastern Mexico. Cretaceous Research, 112:104448.
https://doi.org/10.1016/j.cretres.2020.104448
Than-Marchese, B.A. and Alvarado-Ortega, J. 2022. Armigatus felixi sp. nov. An Albian double armored herring (Clupeomorpha, Ellimmichthyiformes) from the Tlayúa Lagerstätte, Mexico. Journal of South American Earth Sciences, 118:103905.
https://doi.org/10.1016/j.jsames.2022.103905
Than-Marchese, B.A., Alvarado-Ortega, J., Murray, A.M., Velázquez-Velázquez, E., and Domínguez-Domínguez, O.D. 2024. First report of the genus Thorectichthys (Ellimmichthyiformes) in America, from the Cenomanian deposits of the Cintalapa Formation, Chiapas, southeastern Mexico. Cretaceous Research, 154:105739.
https://doi.org/10.1016/j.cretres.2023.105739
Thiollière, V. 1850. Seconde notice sur le gisement et les fossiles d'origine organique des calcaires lithographiques, dans le Jura du Département de l'Ain...(&c.). Annales des sciences physiques et naturelles, d'agriculture et d'industrie, Lyon, (2)3:111-184.
Thiollière, V. 1858. Note sur les poissons fossiles du Bugey, et sur l’application de la méthode de Cuvier à leur classement. Bulletin de la Société géologique de France, 15:782-794.
Toombs, H.A. and Rixon, A.E. 1959. The use of acids in the preparation of vertebrate fossils. Curator, 2:304-312.
https://doi.org/10.1111/j.2151-6952.1959.tb00514.x
Van Vranken, N.E., Fielitz, Ch., and Ebersole, J.A. 2019. New occurrences of Belonostomus (Teleostomorpha: Aspidorhynchidae) from the Late Cretaceous of the North American Gulf Coastal Plain, USA. Palaeontologia Electronica 22.3.58:1-11.
https://doi.org/10.26879/983
Vernygora, O., Murray, A.M., and Wilson, M.V.H. 2016. A primitive clupeomorph from the Albian Loon River Formation (Northwest Territories, Canada). Canadian Journal of Earth Sciences, 53:331-342.
https://doi.org/10.1139/cjes-2015-0172
Waite, L.E. 1986. Contributions to the stratigraphy of the Sierra Madre Limestone (Cretaceous) of Chiapas. Boletín del Instituto de Geología Universidad Nacional Autónoma de México, 102:103-193.
Williston, S.W. 1894. On various vertebrate remains from the lowermost Cretaceous of Kansas. Kansas University Quarterly, 3(1):1-4, pl. I.
Wilson, M.V.H. and Murray, A.M. 1996. Early Cenomanian acanthomorph teleost in the Cretaceous Fish Scale Zone, Albian/Cenomanian boundary, Alberta, Canada, p. 369-382. In Arratia, G. and Viohl, G. (eds.), Mesozoic fishes -Systematics and Paleoecology-. Verlag Dr. Friedrich Pfeil, München, Germany.
Wilson, M.V.H. and Bruner, J.C. 2004. Mesozoic fish Assemblages of North America, p. 575-595. In Arratia, G. and Tintori, A. (eds.), Mesozoic Fishes 3 -Systematics, Paleoenvironments and Biodiversity-. Verlag Dr. Friedrich Pfeil, München, Germany.
