

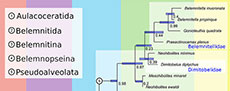
 Bayesian inference reveals a complex evolutionary history of belemnites
Bayesian inference reveals a complex evolutionary history of belemnites
Article number: 26.1.a13
https://doi.org/10.26879/1239
Copyright Paleontological Society, April 2023
Author biographies
Plain-language and multi-lingual abstracts
PDF version
Submission: 22 August 2022. Acceptance: 5 April 2023.
ABSTRACT
Belemnites are an extinct group of Mesozoic coleoid cephalopods, common in Jurassic and Cretaceous marine sedimentary rocks. Despite their significance, their total group phylogeny has rarely been considered in recent decades. In contrast, most researchers restricted the assignment of families to one of the two usually recognized subgroups, the Belemnitina and the Belemnopseina. As for many fossil cephalopods, researchers have been reluctant to employ modern phylogenetic methods to illuminate belemnites' evolutionary history.
To overcome the "dead end" of belemnite systematics, we performed the first tip-dated Bayesian analysis of belemnite phylogeny. In our analysis, the Aulacoceratida are found as the monophyletic sister group to belemnites. The Sinobelemnitidae are resolved as paraphyletic and fall outside the Belemnitina and Belemnopseina, which make up the remaining belemnites. Belemnitina is restricted to Jurassic species with generally no or apical furrows. Holcobelidae are sister group to other Belemnopseina. Cylindroteuthids sensu lato (including Oxyteuthidae) are nested within Belemnopseina, contrary to the common hypothesis placing them within the Belemnitina. Duvaliidae and Dicoelitidae are recovered as members of the Belemnopseina, but their precise relationship has to be evaluated based on more taxa and additional characters. We introduce the well-supported unranked clade Pseudoalveolata, which includes Dimitobelidae, Belemnitellidae, and members of the paraphyletic "Belemnopseidae".
The phylogeny presented here, based on reproducible and quantitative methods, contrasts with the usually applied authoritative "stratophenetic" approach to belemnite systematics, based on the overemphasis of single characters. This result is considered the basis for future studies on belemnite phylogeny, allowing for a rigorous testing of evolutionary hypotheses.
Kevin Stevens*. Institute for Geology, Mineralogy, and Geophysics, Ruhr University Bochum, Germany. Corresponding author. kevin.stevens@rub.de
Alexander Pohle*. Institute for Geology, Mineralogy, and Geophysics, Ruhr University Bochum, Germany and Palaeontological Institute and Museum, University of Zurich, Switzerland. alexander.pohle@rub.de
René Hoffmann. Institute for Geology, Mineralogy, and Geophysics, Ruhr University Bochum, Germany. rene.hoffmann@rub.de
Adrian Immenhauser. Institute for Geology, Mineralogy, and Geophysics, Ruhr University Bochum, Germany. adrian.immenhauser@rub.de
*both authors contributed equally to this study
Keywords: Belemnitida; phylogeny; Mesozoic; coleoid cephalopods
Final citation: Stevens, Kevin, Pohle, Alexander, Hoffmann, René, and Immenhauser, Adrian. 2023. Bayesian inference reveals a complex evolutionary history of belemnites. Palaeontologia Electronica, 26(1):a13.
https://doi.org/10.26879/1239
palaeo-electronica.org/content/2023/3819-belemnite-phylogeny
Copyright: April 2023 Paleontological Society.
This is an open access article distributed under the terms of Attribution-NonCommercial-ShareAlike 4.0 International (CC BY-NC-SA 4.0), which permits users to copy and redistribute the material in any medium or format, provided it is not used for commercial purposes and the original author and source are credited, with indications if any changes are made.
creativecommons.org/licenses/by-nc-sa/4.0/
INTRODUCTION
Belemnites (Belemnitida) are an extinct group of stem-decabrachian coleoids (e.g., Fuchs et al., 2013; Hoffmann and Stevens, 2020). They are characterized by a calcitic rostrum, which is by far the most commonly preserved part of their internal shell. Although exceptional cases of soft-tissue preservation are known from belemnites (e.g., Reitner and Urlichs, 1983; Klug et al., 2010), their preservation potential is lower than in early octobrachians, likely due to differences in buoyancy mechanisms (Clements et al., 2017). Thus, belemnite systematics is almost entirely based on rostrum morphology. In this paper, the terms belemnites and Belemnitida are used only for these calcite-rostrum-bearing species. This definition excludes groups like the Belemnoteuthida and Diplobelida, sometimes referred to as belemnites. The paraphyletic assemblage of Belemnitida, Aulacoceratida, Belemnoteuthida, and Diplobelida is referred to as "belemnoids" in lieu of a proper understanding of their interrelationships at present (Hoffmann and Stevens, 2020). Diplobelida probably represent close relatives of crown-Decabrachia (Fuchs et al., 2013; Fuchs, 2019). Fuchs et al. (2013) regarded belemnites as sister to a group consisting of crown Decabrachia, the stem-decabrachian Longibelus, and the Diplobelida. In a cladistic analysis, Sutton et al. (2016) found the belemnite Hibolithes closely related to the coleoid genera Phragmoteuthis and Belemnoteuthis, nested within the Decabrachia crown group.
The relationship of the rostrum-bearing Aulacoceratida with other "belemnoid" groups, is at present also unclear (Keupp and Fuchs, 2014). Aulacoceratids have aragonitic rostra (also called "telum"; see Jeletzky, 1966) and differ in other morphological aspects from belemnites (e.g., Jeletzky, 1966; Mariotti et al., 2021), but their aragonitic rostrum likely represents the ancestral rostrum structure of coleoids.
The internal phylogenetic relationships of belemnites are even less clear than their relationship to other coleoids. Early subdivisions of belemnites relied on general external characteristics, mostly on the number and position of furrows, e.g., the classification of Werner (1913). Abel (1916) subdivided all belemnites into the families "Clavirostridae" and "Conirostridae" due to their early ontogenetic development. The definitions used nowadays for belemnite families go back to Stolley's (1919) and Naef's (1922) classifications. Jeletzky's (1966) proposal of subdividing the calcite-rostrum bearing belemnites into apically furrowed Belemnitina and alveolar furrowed Belemnopseina has been largely followed by subsequent workers and has been virtually the only applied subdivision of belemnites higher than the family level since. Very few phylogenetic trees have been published that show internal phylogenetic relationships within the Belemnitida - all of them lack a quantitative framework, thus they remain speculative (e.g., Jeletzky, 1966; Schlegelmilch, 1998; Košťák, 2012).
Recognition of the alveolar furrowed Triassic-Early Jurassic Sinobelemnitidae as true belemnites (Zhu and Bian, 1984; Iba et al., 2012; Niko and Ehiro, 2022) has significantly altered views on belemnite phylogeny. Apart from the Sinobelemnitidae, belemnites are exclusively known from the earliest Jurassic onwards, which is considered their initial radiation (e.g., Dera et al., 2016; Neige et al., 2021). Earlier hypotheses of belemnite phylogeny focused on the well-known European fossil record of the group that suggested their origin during the Hettangian in the diminutive and relatively character-poor genera Schwegleria and Nannobelus, which both lack alveolar furrows. By the Early Jurassic, belemnites had reached a cosmopolitan distribution and relatively high diversity and abundance (e.g., Iba et al., 2014a, 2014b; Weis et al., 2015a). Although affected by second-order extinction events (e.g., Dera et al., 2016; De Baets et al., 2021; Neige et al., 2021), belemnites continued to be diverse during the Jurassic and early Early Cretaceous (e.g., Schlegelmilch, 1998; Mutterlose, 1988, 1998; Iba et al., 2011), with the two last occurring, disjunctively distributed families, the Boreal Belemnitellidae and the Austral Dimitobelidae, finally becoming extinct at the K/Pg-boundary (e.g., Doyle, 1992; Christensen, 1997; Iba et al., 2011).
The evolutionary history of belemnites as a whole has rarely been studied since Jeletzky (1966). While several authors speculated about the interrelationships of belemnite families (e.g., Christensen, 1997; Iba et al., 2012; Weis et al., 2012), there has been no study of their phylogenetic relationships based on modern phylogenetic methods. This pattern reflects a general tendency of researchers studying fossil cephalopods in the past (Neige et al., 2007; Bardin et al., 2014; Pohle et al., 2022).
This paper presents the first quantitative approach towards belemnite phylogeny based on Bayesian inference. The dichotomous subdivision of all belemnites into Belemnitina and Belemnopseina, as these groups are usually defined, is not supported by these results. Our findings challenge usual assumptions about the evolution of belemnites and identify parts of the belemnite phylogenetic tree that still lack resolution.
METHODS
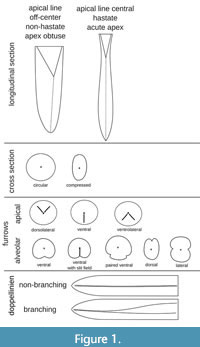 We selected 24 belemnite species, representative of the stratigraphic range, geographic distribution, and diversity of the whole group (Table 1) and scored them for 29 characters, mostly describing the rostrum (Figure 1; Table 2; Appendix 1). Three aulacoceratid genera (including one putative genus) were also included. Although other fossil "belemnoid" coleoid groups are likely more closely related to the Belemnitida than aulacoceratids (e.g., Diplobelida, Belemnoteuthida), these do not have proper rostra (sensu Fuchs, 2012) and so do not contribute to the resolution of internal relationships of belemnites, whose phylogeny is here inferred based mostly on rostrum characters. For the vast majority of belemnites, the rostrum is the only known part (e.g., Hoffmann and Stevens, 2020) mimicking the situation for conodonts, where inferences of their phylogenetic relationships must also be based on conodont element data with few characters only (e.g., Donoghue, 2001; Guenser et al., 2021; Bai et al., 2022). Morphological data comes from several published sources and our own observations (Table 1). The terminology of belemnite morphology follows Hoffmann and Stevens (2020) and Stevens et al. (2022). The character matrix was compiled with Mesquite version 3.7 (Maddison and Maddison, 2021). Coding practice follows suggestions by Brazeau (2011) for morphological character coding.
We selected 24 belemnite species, representative of the stratigraphic range, geographic distribution, and diversity of the whole group (Table 1) and scored them for 29 characters, mostly describing the rostrum (Figure 1; Table 2; Appendix 1). Three aulacoceratid genera (including one putative genus) were also included. Although other fossil "belemnoid" coleoid groups are likely more closely related to the Belemnitida than aulacoceratids (e.g., Diplobelida, Belemnoteuthida), these do not have proper rostra (sensu Fuchs, 2012) and so do not contribute to the resolution of internal relationships of belemnites, whose phylogeny is here inferred based mostly on rostrum characters. For the vast majority of belemnites, the rostrum is the only known part (e.g., Hoffmann and Stevens, 2020) mimicking the situation for conodonts, where inferences of their phylogenetic relationships must also be based on conodont element data with few characters only (e.g., Donoghue, 2001; Guenser et al., 2021; Bai et al., 2022). Morphological data comes from several published sources and our own observations (Table 1). The terminology of belemnite morphology follows Hoffmann and Stevens (2020) and Stevens et al. (2022). The character matrix was compiled with Mesquite version 3.7 (Maddison and Maddison, 2021). Coding practice follows suggestions by Brazeau (2011) for morphological character coding.
We used Bayesian tip-dating, which has become increasingly popular in recent years for phylogenetic inference from morphological data for diverse extinct groups of invertebrates, including cephalopods (e.g., Wright, 2017; Paterson, 2019; Pohle et al., 2022). The analyses were performed in BEAST 2.6.7 (Bouckaert et al., 2019) using the fossilized birth-death model as a tree prior (Stadler, 2010; Gavryushkina et al., 2014; Heath et al., 2014) and the parametrization of net diversification rate, turnover and sampling proportion (Heath et al., 2014). Morphological character evolution was modeled with the Mkv model, including invariant site correction (Lewis, 2001). Characters were partitioned according to their number of states; excepting character 13 ("Doppellinien" type), all characters were binary. The exchangeability rates were set to 1.0 for binary characters and 1.5 for the single three-state character to prevent the artificial upweighting of multistate characters (King et al., 2017), although this is naturally expected to have a minimal impact on the analyses. We furthermore accounted for heterogeneous rates across sites with two discretized gamma shape rate categories. Although usually four or more rate categories are employed in morphological datasets for this purpose (Harrison and Larsson, 2015), we used only two categories due to the small number of characters and states. Tip dates were fixed to the midpoint between the first and last occurrence date of the corresponding species. First and last occurrence dates are based on the literature and are calibrated to the ICS 2020 age model (Appendix 2; Gradstein et al., 2020). We used a strict morphological clock with a lognormally distributed prior (mean = 0.1, standard deviation = 1.25). We placed an exponential prior on the origin (mean = 10 my, offset = 253.1 my), limiting the youngest possible origin date to the age of the oldest taxon of the analysis. This approach avoids unrealistically old estimates while not imposing an overly informative prior. The prior on diversification rate was set to an exponential distribution (mean = 1.0), and the turnover prior to a uniform distribution between 0.0 and 1.0. For the sampling proportion, we used a uniform prior with an upper limit of 0.15, which we justify by a very rough estimate of the number of belemnite species in the Palaeobiology Database (PBDB), which resulted in c. 200 species.
Although this number likely underestimates the true number of belemnite species by some margin due to the incompleteness of both the fossil record and the PBDB, it represents a useful estimate to provide an absolute upper limit for the sampling rate, as it assigns zero probability to any values above 0.15 (corresponding to the ratio between taxa used in the analysis and approximate total number of known species). Lastly, we enforced a monophyletic constraint on the Belemnitida (without the Sinobelemnitidae). We justify this constraint by the strong prior expectation that this group is monophyletic, i.e., a polyphyletic belemnite origin within the Aulacoceratidae or Sinobelemnitidae has never been suggested. At the same time, aulacoceratids and sinobelemnites distinctly predate other belemnites, making an origin of either of these groups from within this constrained clade implausible (however, note that this does not preclude potential paraphyletic relationships with respect to other groups such as Diplobelida, Phragmoteuthida, or crown-Decabrachia). The analysis was run for two separate runs of the MCMC algorithm, each with 10 000 000 generations sampling every 10 000 generations and 10% of the samples discarded as burn-in. Convergence was checked using Tracer (Rambaut et al., 2018). The tree files were combined in LogCombiner (Bouckaert et al., 2019), and the Maximum Clade Credibility tree was generated with TreeAnnotator but using the older BEAST version 1.10.4 (Suchard et al., 2018). In contrast to the latter, TreeAnnotator in BEAST 2.6.7 treats sampled ancestors as not belonging to the same clade, which may result in underestimated posterior probabilities (Barido-Sottani et al., 2020). The xml script to run the analysis in BEAST, the resulting combined log and tree files and the annotated summary tree are contained in Appendix 3, Appendix 4, Appendix 5, Appendix 6.
RESULTS
The parameter estimates of the tip-dated analyses are listed in Table 3. Although these parameters were not a focus of our current study, and more extensive model testing should be carried out before drawing any definite conclusions, they provide some insights. According to the estimated sampling rate (95% HPD interval between 0.015 and 0.15 with 27 included species), we would expect a total number of belemnite and aulacoceratid species that existed until the end of the Cretaceous to be somewhere between approximately 180 and 1 800. Although this is a rather large credible interval, it appears to be a reasonable estimate that could be refined by adding more species or occurrence data. The omission of the diplobelids and phragmoteuthids, which probably belong to the same clade, may also have caused slightly biased estimates. Furthermore, refined models with variable rates through time may also improve these estimates. The age of the last common ancestor (origin parameter) of belemnites and the aulacoceratids included here was estimated to lie within the Permian, which roughly agrees with previous hypotheses (e.g., Jeletzky, 1966; Kröger et al., 2011).
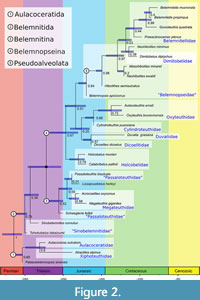 The topology of the maximum clade credibility tree (Figure 2) reveals several well-supported clades, although there are uncertainties in other areas of the tree. The placement of Palaeobelemnopsis sinensis within the Aulacoceratida is moderately well supported (posterior probability = PP = 0.69) as sister to the Atractites alpinus and Aulacoceras sulcatum clade. The Aulacoceratida were recovered as the sister group to the Belemnitida, with an estimated divergence date within the Permian. Furthermore, there is high support for including the sinobelemnitids within the Belemnitida (PP = 0.79), although the Sinobelemnitidae itself is paraphyletic. Within the Belemnitida, there is weak support for a group containing Jurassic taxa traditionally recognized as part of the Belemnitina (PP = 0.53), including Schwegleria feifeli, Passaloteuthis bisulcata, Acrocoelites oxyconus, Megateuthis gigantea, as well as Lissajousibelus harleyi, which was so far of uncertain placement inside Belemnitida. Within the Belemnitina, we recovered Schwegleria feifeli as sister group to the remainder of the Belemnitinae (PP = 0.59), with the Megateuthidae as monophyletic sister group (PP = 0.58) to Passaloteuthis #isulcate and Lissajousibelus harleyi (PP =0.31). Sister to this Belemnitina clade is a larger group containing the remaining belemnites, here referred to as the Belemnopseina (PP= 0.67). The uncertainty of the divergence date between those two clades essentially covers the entire Triassic. Within the Belemnopseina clade, several subclades displayed relatively high support: 1) the sister group relationship between Holcobelus munieri and Calabribelus pallinii (PP = 0.79), which diverged from the rest of the clade (PP = 0.51) between the Late Triassic and Early Jurassic. 2) A large clade containing belemnitids that share a pseudoalveolus (PP = 0.98) and appeared during the Late Jurassic. 3) The same clade but excluding Hibolithes semisulcatus, which was either sister group or sampled ancestor to the latter (PP = 0.97), diverging from the latter during the Late Jurassic. 4) The clade that included Gonioteuthis quadrata and the two species of Belemnitella (PP = 0.99) The clade that includes Belemnopsis apiciconus as sister group to the pseudoalveolus-bearing clade is moderately supported (PP = 0.64). The clade containing Praeactinocamax plenus, Gonioteuthis quadrata, Belemnitella propinqua, and B. mucronata (= Belemnitellidae) is only weakly supported (PP = 0.44), its origin estimated to the late Early Cretaceous. Other clades received low to moderate support. Among these, we recovered a monophyletic clade containing Cylindroteuthis puzosiana, Aulacoteuthis ernsti, and Oxyteuthis brunsvicensis (PP = 0.68), which formed a weakly supported monophyletic clade (PP = 0.25) together with Duvalia and Dicoelites (PP = 0.57). In our analysis, this clade diverged from the pseudoalveolus-bearing belemnites including Belemnopsis between the Early to Middle Jurassic. Furthermore, we recovered a polyphyletic Neohibolites with N. minimus forming a monophyletic clade with Dimitobelus diptychus (PP = 0.39) and N. ewaldi as sister to Mesohibolites minaret (PP = 0.2).
The topology of the maximum clade credibility tree (Figure 2) reveals several well-supported clades, although there are uncertainties in other areas of the tree. The placement of Palaeobelemnopsis sinensis within the Aulacoceratida is moderately well supported (posterior probability = PP = 0.69) as sister to the Atractites alpinus and Aulacoceras sulcatum clade. The Aulacoceratida were recovered as the sister group to the Belemnitida, with an estimated divergence date within the Permian. Furthermore, there is high support for including the sinobelemnitids within the Belemnitida (PP = 0.79), although the Sinobelemnitidae itself is paraphyletic. Within the Belemnitida, there is weak support for a group containing Jurassic taxa traditionally recognized as part of the Belemnitina (PP = 0.53), including Schwegleria feifeli, Passaloteuthis bisulcata, Acrocoelites oxyconus, Megateuthis gigantea, as well as Lissajousibelus harleyi, which was so far of uncertain placement inside Belemnitida. Within the Belemnitina, we recovered Schwegleria feifeli as sister group to the remainder of the Belemnitinae (PP = 0.59), with the Megateuthidae as monophyletic sister group (PP = 0.58) to Passaloteuthis #isulcate and Lissajousibelus harleyi (PP =0.31). Sister to this Belemnitina clade is a larger group containing the remaining belemnites, here referred to as the Belemnopseina (PP= 0.67). The uncertainty of the divergence date between those two clades essentially covers the entire Triassic. Within the Belemnopseina clade, several subclades displayed relatively high support: 1) the sister group relationship between Holcobelus munieri and Calabribelus pallinii (PP = 0.79), which diverged from the rest of the clade (PP = 0.51) between the Late Triassic and Early Jurassic. 2) A large clade containing belemnitids that share a pseudoalveolus (PP = 0.98) and appeared during the Late Jurassic. 3) The same clade but excluding Hibolithes semisulcatus, which was either sister group or sampled ancestor to the latter (PP = 0.97), diverging from the latter during the Late Jurassic. 4) The clade that included Gonioteuthis quadrata and the two species of Belemnitella (PP = 0.99) The clade that includes Belemnopsis apiciconus as sister group to the pseudoalveolus-bearing clade is moderately supported (PP = 0.64). The clade containing Praeactinocamax plenus, Gonioteuthis quadrata, Belemnitella propinqua, and B. mucronata (= Belemnitellidae) is only weakly supported (PP = 0.44), its origin estimated to the late Early Cretaceous. Other clades received low to moderate support. Among these, we recovered a monophyletic clade containing Cylindroteuthis puzosiana, Aulacoteuthis ernsti, and Oxyteuthis brunsvicensis (PP = 0.68), which formed a weakly supported monophyletic clade (PP = 0.25) together with Duvalia and Dicoelites (PP = 0.57). In our analysis, this clade diverged from the pseudoalveolus-bearing belemnites including Belemnopsis between the Early to Middle Jurassic. Furthermore, we recovered a polyphyletic Neohibolites with N. minimus forming a monophyletic clade with Dimitobelus diptychus (PP = 0.39) and N. ewaldi as sister to Mesohibolites minaret (PP = 0.2).
DISCUSSION
Due to the shared regeneration pattern and growth mode of Xiphoteuthidae (in the phylogeny represented by Atractites alpinus) and belemnites, Keupp and Fuchs (2014) suggested aulacoceratid paraphyly. On the other hand, Jeletzky (1966) argued for aulacoceratid monophyly, going so far as to view the group as an independent offshoot of bactritid cephalopods, leaving no descendants. Doyle et al. (1994) favored the derivation of belemnites from within the Aulacoceratida potentially via the Phragmoteuthida. Aulacoceratida are here recovered as a monophyletic group and sister to belemnites, with the Permian Palaeobelemnopsis sinensis confirmed as a member of the Aulacoceratida. However, since we only included a limited number of aulacoceratids in our analysis, we cannot rule out that the total group Aulacoceratida is paraphyletic, also with respect to other groups of early coleoids such as the Phragmoteuthida.
Since Jeletzky (1966), all belemnites were usually divided into two suborders; Belemnitina and Belemnopseina, with members of the former group considered ancestral to the latter. Under this traditional scheme, Belemnitina groups taxa with apical furrows and Belemnopseina taxa with alveolar furrows. Problematic under the Belemnitina/Belemnopseina scheme is that the earliest belemnites of the Sinobelemnitidae are considered to be Belemnopseina with a gap of ca. 25 Ma between the youngest Sinobelemnitida and remaining Belemnopseina (Iba et al., 2012). At least one of the two sinobelemnitids in the current analysis, on the other hand, forms the sister group to the remaining belemnites (Figure 2). This result is in better agreement with the fossil record than the earlier hypothesis. It casts doubt on the homologization of dorsal alveolar furrows in belemnites, which was discussed as a potential uniting character of the Sinobelemnitidae with the Duvaliidae or Dicoelitidae (Iba et al., 2012). Still, a well-resolved position of the Sinobelemnitidae and clarification of their mono- or paraphyly will require a detailed study of this still sparsely known group and a better sampled Triassic fossil record of belemnites in general.
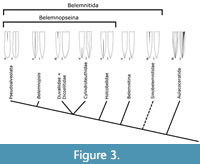 The species Lissajousibelus harleyi displays a ventral furrow in addition to dorsolateral apical furrows and was considered close to Belemnopseina by Weis et al. (2015b). In the present analysis, L. harleyi was found to be closely related to typical Belemnitina. Our Bayesian approach demonstrates how the relationship of belemnite taxa that do not easily fit the Belemnitina/Belemnopseina-scheme of Jeletzky (1966) can be resolved quantitatively by taking into account the maximally inclusive morphological and stratigraphic evidence. Belemnitina as defined herein is recovered as a monophyletic group with a likely origin in the Late Triassic but otherwise restricted to the Jurassic (Figure 2 and Figure 3). A close relationship between these taxa was already suggested by Stolley (1919), who grouped them into his family "Polyteuthidae". This new definition includes at least the likely paraphyletic "Passaloteuthidae" (following the definition of Schlegelmilch, 1998) and the Megateuthidae (Acrocoelitidae); it remains to be investigated in which way other Jurassic families not considered in the present analysis (e.g., Hastitidae, Salpingoteuthidae) are related to this group. Likewise, as taxonomic concepts differ for groups such as the “Passaloteuthidae”, including more species of these families may help clarifying their relationships further.
The species Lissajousibelus harleyi displays a ventral furrow in addition to dorsolateral apical furrows and was considered close to Belemnopseina by Weis et al. (2015b). In the present analysis, L. harleyi was found to be closely related to typical Belemnitina. Our Bayesian approach demonstrates how the relationship of belemnite taxa that do not easily fit the Belemnitina/Belemnopseina-scheme of Jeletzky (1966) can be resolved quantitatively by taking into account the maximally inclusive morphological and stratigraphic evidence. Belemnitina as defined herein is recovered as a monophyletic group with a likely origin in the Late Triassic but otherwise restricted to the Jurassic (Figure 2 and Figure 3). A close relationship between these taxa was already suggested by Stolley (1919), who grouped them into his family "Polyteuthidae". This new definition includes at least the likely paraphyletic "Passaloteuthidae" (following the definition of Schlegelmilch, 1998) and the Megateuthidae (Acrocoelitidae); it remains to be investigated in which way other Jurassic families not considered in the present analysis (e.g., Hastitidae, Salpingoteuthidae) are related to this group. Likewise, as taxonomic concepts differ for groups such as the “Passaloteuthidae”, including more species of these families may help clarifying their relationships further.
The Holcobelidae, Duvaliidae, and Dicoelitidae have an uncertain phylogenetic placement (e.g., Stolley, 1911a, 1911b; Stevens, 1964; Jeletzky, 1966; Combémorel, 1973; Weis et al., 2012). Our results place the Holcobelidae as the sister group to the rest of the Belemnopseina. This leaves it ambiguous whether their ventral furrow that does not reach the alveolus represents the ancestral state of the Belemnopseina or if this character represents a secondary development from an alveolar furrow reaching the alveolus as seen in typical "Belemnopseidae". In our tree, the placement of Duvaliidae as sister to Dicoelitidae finds moderate support, but the placement of this clade itself has only weak support (Figure 2). The Duvaliidae share with the Dicoelitidae the presence of a dorsal alveolar furrow. However, a belemnopsein ventral alveolar furrow is also developed in the Dicoelitidae. Duvalia grasiana has recently been shown to display more organic-rich primary rostrum calcite than other belemnites (Stevens et al., 2022). A similar detailed description of the microstructure of other duvaliids and potentially related forms might reveal the phylogenetic position of this enigmatic group in future studies with more certainty.
The monophyletic clade containing the Oxyteuthidae as sister to Cylindroteuthis puzosiana confirms earlier thoughts on their phylogenetic relationships (e.g., Mutterlose, 1983). However, in contrast to these earlier hypotheses, we recovered the Cylindroteuthidae sensu lato clade within the Belemnopseina instead of within the Belemnitina. Still, the problem of the phylogenetic placement and evolution of Cylindroteuthidae and Oxyteuthidae needs further focused analyses, especially for Boreal belemnites of the Jurassic-Cretaceous transition, which will also have to include closely related species of the Pachyteuthidae. Our analysis suggests homology of the ventral furrow present in many Cylindroteuthidae sensu lato, not with the apical ventral furrow as is typical for the Megateuthidae/Acrocoelitidae, but with the belemnopsein ventral furrow. This hypothesis contrasts with the "stratophenetic" reasoning of the independent evolution of a ventral furrow in the genus Aulacoteuthis (Mutterlose and Baraboshkin, 2003; Baraboshkin and Mutterlose, 2004).
A name is suggested here for a newly identified and well-supported clade inside the Belemnopseina, the unranked Pseudoalveolata (see Appendix 7; PP=0.98; Figure 2). The Pseudoalveolata is characterized by the synapomorphy of pseudoalveolus formation, a secondary alveolus-deepening, developing by dissolution/erosion of anterior organic-rich rostrum sections (Stevens et al., 2022). The pseudoalveolus is further characterized by a well-mineralized ‘“spike” projecting anteriorly toward the protoconch (Nadelspitze of Stolley, 1911a). Hibolithes has here been recovered as the earliest pseudoalveolate belemnite. Pseudoalveolus types have been considered of great importance in the phylogeny of the Belemnitellidae (e.g., Košťák, 2012), but contrary to suggestions by Dauphin et al. (2007) and Košťák and Wiese (2008), there is no conclusive evidence for the anterior rostrum of pseudoalveolate belemnites being of primarily aragonitic composition, as it likely consisted of calcite with primarily high organic contents (see Stevens et al., 2017; 2022). Irrespective of its composition, it still seems to be a different structure, which is consistently recovered.
The two species of Neohibolites analyzed herein (N. ewaldi and N. minimus) are not recovered as sister species. This indicates the possibly polyphyletic or paraphyletic nature of Neohibolites, a genus which has long been seen as ancestral to the two only belemnite families left after the Cenomanian, the Belemnitellidae and Dimitobelidae, (e.g., Mutterlose, 1998). Future studies focusing on the origin of Belemnitellidae and Dimitobelidae might shed more light on the exact origins of these two last surviving belemnite groups and their origins in the paraphyletic "Belemnopseidae". "Belemnopseidae" has long been recognized as paraphyletic with respect to the Belemnitellidae and Dimitobelidae (e.g., Jeletzky, 1966; Mutterlose, 1988). However, a new phylogenetic definition of the family containing all belemnites closer to Belemnopsis than to the Pseudoalveolata seems possible. This would require a more thorough sampling of the diverse "belemnopseid" taxa of the earliest Cretaceous and a thorough revision of not only the genus Belemnopsis but also Hibolithes, encompassing detailed revision of the diverse Belemnopsis species of the Austral Realm (e.g., Stevens, 1965; Challinor, 1990). Based on the unclear and absent types for both genera (e.g., Combémorel and Howlett, 1993; Mitchell, 2015), the assignment of species to either genus was often based on superficial morphological assessment. We here confirm that there is no pseudoalveolus formation in the type species of Belemnopsis, B. apiciconus. Pseudoalveolus formation had already been suggested as a differentiating character of the genera Belemnopsis and Hibolithes by Stolley (1911a) but was unfortunately not followed on by later authors.
The tip-dated Bayesian analysis confirms earlier ideas that the epirostrum, a "tertiary" rostrum formation (Fuchs, 2012), which is developed only in some belemnites, represents a parallelism and does not indicate a close relationship (Bandel and Spaeth, 1988; Arkhipkin et al., 2015; Stevens et al., 2017). Epirostra are present in the analyzed species Megateuthis gigantea, Holcobelus munieri, Calabribelus pallinii, and Neohibolites minimus, found on disparate parts of our tree (Figure 2). However, it is unclear how preservation, ontogeny and producing polished sections may affect whether an epirostrum is recovered or not. According to Arkhipkin et al. (2015), it forms rather late in ontogeny and similar structures evolved repeatedly in different coleoid lineages. Thus, it is possible that they have been overlooked in some cases, but it likely presents a highly homoplastic character.
In our proposed systematic framework (Figure 3, Table 4), we regard the potentially paraphyletic Sinobelemnitidae as the earliest belemnites. The remainder of the belemnites still falls into two large monophyletic clades, the Belemnitina and Belemnopseina. To align with the current taxonomy as far as possible, we retain those two clades as suborders but slightly adjust the included families. We consider the Cylindroteuthidae sensu lato, Duvaliidae, and Dicoelitidae more derived than the Holcobelidae inside the Belemnopseina but otherwise of uncertain position with regard to the Belemnopsis + Pseudoalveolata clade.
The presented topology represents only a first step towards a well-resolved phylogeny of all belemnites. To achieve further resolution, it will likely be necessary to detect and evaluate further microstructural and geochemical data of several belemnite taxa and incorporate more taxa and characters into the analysis. This includes the use of continuous characters, which are currently not straightforward to implement in Bayesian analyses. Morphometric analyses show their potential in this regard, as belemnite morphospace appears to reflect taxonomic units (Dera et al., 2016). Furthermore, well-preserved specimens from Konservat-Lagerstätten may provide additional valuable insights into the variability of soft-part anatomy, statoliths, radulae, hooks, or jaws within belemnites (e.g., Klug et al., 2010; Fuchs and Hoffmann, 2017), potentially adding other characters to include into phylogenetic analysis. Unfortunately, many of these characters will probably remain unknown for the majority of belemnite species. However, they would be potentially informative for a more inclusive phylogenetic analysis involving non-rostrum bearing "belemnoids" to resolve the decabrachian crown and stem groups.
CONCLUSIONS
The first tip-dated analysis of belemnite (Belemnitida) phylogeny is presented. Our results suggest that the usually applied dichotomous subdivision of all belemnites into Belemnitina and Belemnopseina based only on the presence of apical versus alveolar furrows needs some adjustment. We consequently suggest the subdivision of all belemnites, except the early and potentially paraphyletic Sinobelemnitidae, into newly phylogenetically defined Belemnitina and Belemnopseina. Holcobelidae are the sister group to all other Belemnopseina, and Duvaliidae and Dicoelitidae are confirmed as Belemnopseina but are still of uncertain placement inside this group. A major change involves the transfer of the Cylindroteuthidae (including Oxyteuthidae) from the Belemnitina to the Belemnopseina. A new well-supported subgroup of the Belemnopseina, the unranked Pseudoalveolata, is suggested here, including a phylogenetic definition and the suggestion of a potential synapomorphy.
Because of their high fossilization potential and often high abundance in the marine fossil record, belemnites are particularly important in tracking faunal changes of the Jurassic and Cretaceous pelagic realms. The present study is only a first step; further analyses based on more taxa and characters, including continuous characters and detailed microstructural analyses of the rostra, are needed to further resolve more details of the belemnites’ evolutionary history. Applying quantitative and reproducible phylogenetic methodology in contrast to earlier approaches relying on authoritative overemphasis of single characters will lay a solid foundation for the future study of belemnites.
ACKNOWLEDGMENTS
This research was funded by DFG project no. 507867999. We thank two anonymous reviewers and Matus Hyzny for comments and suggestions that significantly improved this paper. We further thank handling editor Joshua Samuels.
REFERENCES
Abel, O. 1916. Paläobiologie der Cephalopoden aus der Gruppe der Dibranchiaten. Gustav Fischer Verlag, Jena.
Arkhipkin, A., Weis, R., Mariotti, N., and Shcherbich, Z. 2015. ‘Tailed’ cephalopods. Journal of Molluscan Studies, 81:345-355. https://doi.org/10.1093/mollus/eyu094
Bai, R., Song, H., Benton, M.J., and Tian, L. 2022. Phylogenetic classification and evolution of Early Triassic conodonts. Palaeogeography, Palaeoclimatology, Palaeoecology 585:110731. https://doi.org/10.1016/j.palaeo.2021.110731
Bandel, K. and Spaeth, C. 1988. Structural differences in the ontogeny of some belemnite rostra, p. 247-271. In Wiedmann, J. and Kullmann, J. (eds.), Cephalopods Present and Past. Schweitzerbartsche Verlagsbuchhandlung, Stuttgart, Germany.
Baraboshkin, E.J. and Mutterlose, J. 2004. Correlation of the Barremian belemnite successions of northwest Europe and the Ulyanovsk - Saratov area (Russian Platform). Acta Geologica Polonica, 54:499-510.
Bardin, J., Rouget, I., and Cecca, F. 2014. Cladistics in ammonoids: back to the future. Neues Jahrbuch für Geologie und Paläontologie, 274:239-253. https://doi.org/10.1127/njgpa/2014/0449
Barido-Sottani, J., van Tiel, N.M.A., Hopkins, M.J., Wright, D.F., Stadler, T., and Warnock, R.C.M. 2020. Ignoring fossil age uncertainty leads to inaccurate topology and divergence time estimates in time calibrated tree inference. Frontiers in Ecology and Evolution, 8:183. https://doi.org/10.3389/fevo.2020.00183
Bather, F.A. 1888. Shell-growth in Cephalopoda (Siphonopoda). Journal of Natural History, 1:298-309.
Blainville, M.H. 1827. Mémoire sur les Bélemnites, considérés zoologiquement et géologiquement. Levrault, Strasbourg.
Boehm, G. 1906. Geologische Mitteilungen aus dem Indo-australischen Archipel. 1: Neues aus dem Indo-australischen Archipel. Neues Jahrbuch für Mineralogie, Geologie und Paläontologie, 22:385-412.
Bouckaert, R., Vaughan, T.G., Barido-Sottani, J., Duchêne, S., Fourment, M., Gavryushkina, A., Heled, J., Jones, G., Kühnert, D., De Maio, N., Matschiner, M., Mendes, F.K., Müller, N.F., Ogilvie, H.A., du Plessis, L., Popinga, A., Rambaut, A., Rasmussen, D., Siveroni, I., Suchard, M.A., Wu, C.-H., Xie, D., Zhang, C., Stadler, T., and Drummond, A.J. 2019. BEAST 2.5: An advanced software platform for Bayesian evolutionary analysis. PLoS Computational Biology, 15:e1006650. https://doi.org/10.1371/journal.pcbi.1006650
Brazeau, M.D. 2011. Problematic character coding methods in morphology and their effects. Biological Journal of the Linnean Society, 104:489-498. https://doi.org/10.1111/j.1095-8312.2011.01755.x
Challinor, A.B. 1990. A belemnite biozonation for the Jurassic-Cretaceous of Papua New Guinea and a faunal comparison with eastern Indonesia. BMR Journal of Australian Geology & Geophysics, 11:429-447.
Chen, T. 1982. Mesozoic Coleoidea fauna from Xizang. Palaeontology of Xizang, 4:282-325. [Chinese]
Christensen, W.K. 1971. Belemnitella propinqua propinqua (Moberg, 1885) from Scandinavia. Bulletin of the Geological Society of Denmark, 20:369-389.
Christensen, W.K. 1997. The Late Cretaceous belemnite family Belemnitellidae: Taxonomy and evolutionary history. Bulletin of the Geological Society of Denmark, 44:59-88.
Clements, T., Colleary, C., De Baets, K., and Vinther, J. 2017. Buoyancy mechanisms limit preservation of coleoid cephalopod soft tissues in Mesozoic Lagerstätten. Palaeontology, 60:1-14. https://doi.org/ 10.1111/pala.12267
Combémorel, R. 1973. Les Duvaliidae (Pavlow) du cretace inferieur francais. Document des Laboratoires de Geology de la Faculte des Sciences de Lyon, 57:131-186.
Combémorel, R. and Howlett, P. 1993. Le genre Hibolites: Hibolites Mayer-Eymar, 1883, non Hibolithes Montfort, 1808, belemnite du Jurassique. Geobios, 15:67-72. https://doi.org/10.1016/S0016-6995(06)80362-1
Cuvier, G. 1797. Tableau élémentaire de l´histoire naturelle des animaux. Baudoin, Paris.
Dauphin, Y., Williams, C.T., and Barskov, I.S. 2007. Aragonitic rostra of the Turonian belemnitid Goniocamax: Arguments from diagenesis. Acta Palaeontologica Polonica, 52:85-97.
De Baets, K., Nätscher, P.S., Rita, P., Fara, E., Neige, P., Bardin, J., Dera, G., Duarte, L.V., Hughes, Z., Laschinger, P., García-Ramos, J.C., Piñuela, L., Übelacker, C., and Weis, R. 2021. The impact of the Pliensbachian-Toarcian crisis on belemnite assemblages and size distribution. Swiss Journal of Palaeontology, 140:25. https://doi.org/10.1186/s13358-021-00242-y
Dera, G., Toumoulin, A., and De Baets, K. 2016. Diversity and morphological evolution of Jurassic belemnites from South Germany. Palaeogeography, Palaeoclimatology, Palaeoecology, 457:80-97. https://doi.org/10.1016/j.palaeo.2016.05.029
Donoghue, P.C.J. 2001. Conodonts Meet Cladistics: Recovering Relationships and Assessing the Completeness of the Conodont Fossil Record. Palaeontology, 44:65-93. https://doi.org/10.1111/1475-4983.00170
Doyle, P. 1990. The British Toarcian (Lower Jurassic) Belemnites. Part 1. Monograph of the Palaeontographical Society Publication, 584:1-49. The Palaeontographical Society, London, UK.
Doyle, P. 1992. A review of the biogeography of Cretaceous belemnites. Palaeogeography, Palaeoclimatology, Palaeoecology, 92:207-216. https://doi.org/10.1016/0031-0182(92)90082-G
Doyle, P., Donovan, D.T., and Nixon, M. 1994. Phylogeny and systematics of the Coleoidea. The University of Kansas, Paleontological Contributions, 5:1-15.
d'Orbigny, A. 1842. Paléontologie française: description zoologique et géologique de tous les animaux mollusques et rayonnés fossiles de France. Terrains crétacés. Les céphalopodes 1. A. Bertrand, Paris, France.
Duval-Jouve, J. 1841. Bélemnites des terrains crétacés inférieures des environs de Castellane (Basses-Alpes), considérées géologiquement et zoologiquement, avec la description de ces terrains. Académie des Sciences Naturelles, Paris, France.
Ernst, G. 1964. Ontogenie, Phylogenie und Stratigraphie der Belemnitengattung Gonioteuthis Bayle aus dem nordwestdeutschen Santon/Campan. Fortschritte der Geologie in Rheinland und Westfalen, 7:113-174.
Eudes-Deslongchamps, E. 1878. Le Jura Normand. 2ième livraison, monographies 4. Assises supérieures des marnes infra-oolithiques. Savy, Paris; Blanc-Hardel, Caen.
Fuchs, D. 2012. The “rostrum”-problem in coleoid terminology - an attempt to clarify inconsistencies. Geobios, 45:29-39. https://doi.org/10.1016/j.geobios.2011.11.014
Fuchs, D. 2019. Part M, Coleoidea, Chapter 23E: Systematic descriptions: Diplobelida. Treatise Online, 118.
Fuchs, D. and Hoffmann, R. 2017. Part M, Chapter 10: Arm armature in belemnoid coleoids. Treatise Online, 91:1-20.
Fuchs, D., Iba, Y., Ifrim, C., Nishimura, T., Kennedy, W.J., Keupp, H., Stinnesbeck, W., and Tanabe, K. 2013. Longibelus gen. nov., a new Cretaceous coleoid genus linking Belemnoidea and early Decabrachia. Palaeontology, 56:1081-1106.
https://doi.org/10.1111/pala.12036
Gavryushkina, A., Welch, D., Stadler, T., and Drummond, A.J. 2014. Bayesian inference of sampled ancestor trees for epidemiology and fossil calibration. PLoS Computational Biology, 10:e1003919. https://doi.org/10.1371/journal.pcbi.1003919
Gradstein, F.M., Ogg, J.G., Schmitz, M.D., and Ogg, G.M. 2020. Geologic time scale 2020. Elsevier.
Guenser, P., Warnock, R.C., Pett, W., Donoghue, P.C., and Jarochowska, E. 2021. Does time matter in phylogeny? A perspective from the fossil record. bioRxiv. https://doi.org/10.1101/2021.06.11.445746
Gustomesov, V.A. 1977. K revizii yurskikh belemnitov [On a revision of Jurassic belemnites]. Bjulleten Moskovskogo Obshchestva Ispytateley Prirody (Novaya Serija, Geologicheskiye nauki), 52:103-117. [Russian]
Harrison, L.B. and Larsson, H.C.E. 2015. Among-character rate variation distributions in phylogenetic analysis of discrete morphological characters. Systematic Biology, 64:307-324. https://doi.org/10.1093/sysbio/syu098
Heath, T.A., Huelsenbeck, J.P., and Stadler, T. 2014. The fossilized birth-death process for coherent calibration of divergence-time estimates. Proceedings of the National Academy of Sciences, 111:E2957-2966. https://doi.org/10.1073/pnas.1319091111
Hoffmann, R. and Stevens, K. 2020. The palaeobiology of belemnites - foundation for the interpretation of rostrum geochemistry. Biological Reviews, 95:94-123. https://doi.org/10.1111/brv.12557
Iba, Y., Mutterlose, J., Tanabe, K., Sano, S., Misaki, A., and Terabe, K. 2011. Belemnite extinction and the origin of modern cephalopods 35 m.y. prior to the Cretaceous−Paleogene event. Geology, 39:483-486. https://doi.org/10.1130/G31724.1
Iba, Y., Sano, S., Mutterlose, J., and Kondo, Y. 2012. Belemnites originated in the Triassic - A new look at an old group. Geology, 40:911-914. https://doi.org/10.1130/G33402.1
Iba, Y., Sano, S., and Mutterlose, J. 2014. The Early Evolutionary History of Belemnites: New Data from Japan. PLoS ONE, 9:e95632. https://doi.org/10.1371/journal.pone.0095632
Iba, Y., Sano, S., Rao, X., Fuchs, D., Chen, T., Weis, R., and Sha, J. 2015. Early Jurassic belemnites from the Gondwana margin of the Southern Hemisphere-Sinemurian record from South Tibet. Gondwana Research, 28(2):882-887.
https://doi.org/.10.1016/j.gr.2014.06.007
Jeletzky, J.A. 1965. Taxonomy and phylogeny of fossil Coleoidea (=Dibranchiata). Geological Survey, Canada, Paper 65:72-76.
Jeletzky, J.A. 1966. Comparative morphology, phylogeny, and classification of fossil Coleoidea. The University of Kansas Paleontological Contributions 7:1-162.
Keupp, H. and Fuchs, D. 2014. Different regeneration mechanisms in the rostra of aulacocerids (Coleoidea) and their phylogenetic implications. Göttingen Contributions to Geosciences, 77:13-20.
King, B., Qiao, T., Lee, M.S.Y., Zhu, M., and Long, J.A., 2017. Bayesian morphological clock methods resurrect placoderm monophyly and reveal rapid early evolution in jawed vertebrates. Systematic Biology, 66(4):499-516. https://doi.org/10.1093/sysbio/syw107
Klug, C., Schweigert, G., Fuchs, D., and Dietl, G. 2010. First record of a belemnite preserved with beaks, arms and ink sac from the Nusplingen Lithographic Limestone (Kimmeridgian, SW Germany): Belemnite with beaks, arms and ink sac. Lethaia, 43:445-456. https://doi.org/10.1111/j.1502-3931.2009.00203.x
Košťák, M. 2012. On the Turonian origin of the Goniocamax - Belemnitella stock (Cephalopoda, Coleoidea). Geobios, 45:79-85. https://doi.org/10.1016/j.geobios.2011.11.004
Košťák, M. and Wiese, F. 2008. Lower Turonian record of belemnite Praeactinocamax from NW Siberia and its palaeogeographic significance. Acta Palaeontologica Polonica, 53:669-678. https://doi.org/10.4202/app.2008.0411
Kröger, B., Vinther, J., and Fuchs, D. 2011. Cephalopod origin and evolution: a congruent picture emerging from fossils, development and molecules. BioEssays, 33:602-613. https://doi.org/10.1002/bies.201100001
Lewis, P.O. 2001. A likelihood approach to estimating phylogeny from discrete morphological character data. Systematic Biology, 50:913-925. https://doi.org/10.1080/106351501753462876
Maddison, W. P. and Maddison, D.R. 2021. Mesquite: a modular system for evolutionary analysis. Version 3.70. http://www.mesquiteproject.org
Mariotti, N., Pignatti, J., and Riegraf, W. 2021. Part M, Coleoidea, Chapter 23E: Systematic descriptions: Aulacoceratida. Treatise Online, 148.
Mayer, K. 1866. Diagnoses de bélemnites nouvelles. Journal de Conchylogie, 3:358-369.
McCoy, F. 1867. On the occurrence of Ichthyosaurus and Plesiosaurus in Australia. Annuals and Magazine of Natural History, 19:355-356.
Miller, J.S. 1826. Observations on Belemnites V. Transactions of the Geological Society of London, second series, 1:45-62.
Mitchell, S.F. 2015. A reassessment of the validity and affinities of Belemnites sulcatus Miller, 1826, Belemnopsis Edwards in Gray, 1849, and Belemnopsis Bayle, 1878. Carnets de Geologie, 15:31-39.
Moberg, J.C. 1885. Cephalopoderne i Sveriges Kritsystem. II. Artsbeskrifning. Sveriges geologiska undersökning. Afhandlingar och upsatser C. 73.
Mutterlose, J. 1983. Phylogenie und Biostratigraphie der Unterfamilie Oxyteuthinae (Belemnitida) aus dem Barreme (Unter-Kreide) NW-Europas. Palaeontographica Abteilung A, 180:1-90.
Mutterlose, J. 1988. Migration and evolution patterns in Upper Jurassic and Lower Cretaceous belemnites, p. 525-537. In: Wiedmann, J. and Kullmann, J. (eds.), Cephalopods - Present and Past. Schweizerbart’sche Verlagsbuchhandlung, Stuttgart.
Mutterlose, J. 1998. The Barremian-Aptian turnover of biota in northwestern Europe: evidence from belemnites. Palaeogeography, Palaeoclimatology, Palaeoecology, 144:161-173. https://doi.org/10.1016/S0031-0182(98)00081-9
Mutterlose, J. and Baraboshkin, E.J. 2003. Taxonomy of the Early Cretaceous belemnite species Aulacoteuthis absolutiformis (Sinzow, 1877) and its type status. Berliner Paläobiologische Abhandlungen, 3:179-187.
Naef, A. 1922. Die fossilen Tintenfische. Verlag Gustav Fischer, Jena.
Neige, P., Rouget, I., and Moyne, S. 2007. Phylogenetic Practices Among Scholars of Fossil Cephalopods, with Special Reference to Cladistics, p. 3-14. In Landman, N.H., Davis, R.A., and Mapes, R.H. (eds.), Cephalopods Present and Past: New Insights and Fresh Perspectives. Springer Netherlands, Dordrecht.
Neige, P., Weis, R., and Fara, E. 2021. Ups and downs of belemnite diversity in the Early Jurassic of Western Tethys. Palaeontology, 64:263-283. https://doi.org/10.1111/pala.12522
Niko, S. and Ehiro, M. 2022. Tohokubelus gen. nov., the Oldest Belemnite from the Olenekian (Lower Triassic) of Northeast Japan. Paleontological Research, 26:115-123. https://doi.org/10.2517/PR200036
Paterson, J.R., Edgecombe, G.D., and Lee, M.S.Y. 2019. Trilobite evolutionary rates constrain the duration of the Cambrian explosion. Proceedings of the National Academy of Sciences, 116:4394-4399. https://doi.org/10.1073/pnas.1819366116
Pavlow, A.P. 1914. Jurassic and lower Cretaceous Cephalopoda of North Siberia. Zapiski Imperatorskoj Akademii Nauk, 8:1-68. [Russian]
Pohle, A., Kröger, B., Warnock, R.C.M., King, A.H., Evans, D.H., Aubrechtová, M., Cichowolski, M., Fang, X., and Klug, C. 2022. Early cephalopod evolution clarified through Bayesian phylogenetic inference. BMC Biology, 20:88.
https://doi.org/10.1186/s12915-022-01284-5
Rambaut, A., Drummond, A.J., Xie, D., Baele, G., and Suchard, M.A. 2018. Posterior summarization in Bayesian phylogenetics using Tracer 1.7. Systematic Biology, 67:901-904. https://doi.org/10.1093/sysbio/syy032
Raspail, F.V. 1829. Histoire naturelle des belemnites, accompagnee de la description et de la
classification des especes, que M. Emeric de Castellane a recuillies dans les Basses-Alpes de Provence. Annales de science d’observation, 1:271-331.
Reitner, J. and Urlichs, M. 1983. Echte Weichteilbelemniten aus dem Untertoarcium (Posidonienschiefer) Südwestdeutschlands. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen, 165:450-465.
Sachs, V.N. and Nalnjaeva, T.I. 1967. Recognition of the superfamily Passaloteuthaceae in the suborder Belemnoidea (Cephalopoda, Dibranchia, Decapoda). Doklady Akademii Nauk SSSR, 173:438-441. [Russian]
Schlegelmilch, R. 1998. Die Belemniten des suddeutschen Jura. Ein Bestimmungsbuch für Geowissenschaftler und Fossiliensammler. Gustav Fischer Verlag, Stuttgart, Germany.
Schwegler, E. 1939. Belemniten aus dem Psilonotenton Schwabens. Zentralblatt für Mineralogie, Geologie und Paläontologie, Abteilung B, 5:200-208.
Spaeth, C. 1871. Untersuchungen an Belemniten des Formenkreises um Neohibolites minimus (Miller, 1826) aus dem Mittel- und Ober-Alb Nordwestdeutschlands. Beihefte zum Geologischen Jahrbuch, 100:1-127.
Stadler, T. 2010. Sampling-through-time in birth-death trees. Journal of Theoretical Biology, 267:396-404. https://doi.org/10.1016/j.jtbi.2010.09.010
Stevens, G.R. 1964. The belemnite genera Dicoelites Boehm and Prodicoelites Stolley. Palaeontology, 7:606-620.
Stevens, G.R. 1965. The Jurassic and Cretaceous of New Zealand and a Review of the Jurassic and Cretaceous belemnites of the Indo-Pacific Region. New Zealand Geological Survey Paleontological Bulletin, 36:1-238.
Stevens, K., Griesshaber, E., Schmahl, W., Casella, L.A., Iba, Y., and Mutterlose, J. 2017. Belemnite biomineralization, development, and geochemistry: The complex rostrum of Neohibolites minimus. Palaeogeography, Palaeoclimatology, Palaeoecology, 468:388-402. https://doi.org/10.1016/j.palaeo.2016.12.022
Stevens, K., Mutterlose, J., Ohnemus, B., Idakieva, V., and Ivanov, M. 2022. Microstructures of Early Cretaceous belemnites and their diagenesis. Cretaceous Research, 137:105259. https://doi.org/10.1016/j.cretres.2022.105259
Stolley, E. 1911a. Beitrage zur Kenntnis der Cephalopoden der norddeutschen unteren Kreide, I. Die Belemnitiden der norddeutschen unteren Kreide, 1. Die Belemniten des norddeutschen Gaults (Aptiens und Albiens). Geologische und Palaontologische Abhandlungen, 10:3.
Stolley, E. 1911b. Studien an den Belemniten der unteren Kreide Norddeutschlands. Jahresberichte des niedersächsischen geologischen Vereins, 4:174-191.
Stolley, E. 1919. Die Systematik der Belemniten. Jahresbericht des Niedersächsischen Geologischen Vereins, 11:1-59.
Stoyanova-Vergilova, M. 1970. Les Fossiles de Bulgarie, IVa Cretace inferieur, Belemnitida. Academie Bulgare des Sciences. [Bulgarian]
Strombeck, A.V. 1861. Über den Gault und besondere die Gargas-Mergel (Aptien d'Orb.) im nordwestlichen Deutschland nächst dem Harze. Zeitschrift der deutschen geologischen Gesellschaft, 13:20-60.
Suchard, M.A., Lemey, P., Baele, G., Ayres, D.L., Drummond, A.J., and Rambaut, A. 2018. Bayesian phylogenetic and phylodynamic data integration using BEAST 1.10. Virus Evolution, 4:vey016. https://doi.org/10.1093/ve/vey016
Sutton, M., Perales-Raya, C., and Gilbert, I. 2016. A phylogeny of fossil and living neocoleoid cephalopods. Cladistics, 32:297-307. https://doi.org/10.1111/cla.12131
von Gümbel, C.W. 1861. Geognostische Beschreibung des bayerischen Alpengebirges und seines Vorlandes 1. J. Perthes, Gotha.
von Hauer, F. 1860. Nachträge zur Kenntniss der Cephalopoden-Fauna der Hallstätter Schichten. Sitzungsberichte der kaiserlichen Akademie der Wissenschaften, mathematisch-naturwissenschaftliche Classe, 41:113-148.
von Schlotheim, E.F. 1813. Beiträge zur Naturgeschichte der Versteinerungen in geognostischer Hinsicht. Leonhardʼs Taschenbuch fur die Gesammte Mineralogie, 7:3-134.
von Schlotheim, E.F. 1820. Die Petrefaktenkunde auf ihrem jetzigen Standpunkte, durch die Beschreibung seiner Sammlung versteinerter und fossiler Überreste des Thier- und Pflanzenreichs der Vorwelt erläutert. Becker, Gotha.
von Zieten, K.H.V. 1830-1833. Die Versteinerungen Württembergs. Schweizerbart, Stuttgart.
von Zittel, K.A. 1895. Grundzüge der Palaeontologie (Palaeozoologie). R. Oldenbourg, Munich.
Weis, R., Mariotti, N., and Di Cencio, A. 2015a. Systematics and evolutionary implications of Early Jurassic belemnites from the Peri-Mediterranean Tethys. Paläontologische Zeitschrift, 89:729-747. https://doi.org/10.1007/s12542-015-0265-5
Weis, R., Dzyuba, O.S., Mariotti, N., and Chesnier, M. 2015b. Lissajousibelus nov. gen., an Early Jurassic canaliculate belemnite from Normandy, France. Swiss Journal of Palaeontology, 134:289-300. https://doi.org/10.1007/s13358-015-0086-x
Weis, R., Mariotti, N., Riegraf, W. 2012. The belemnite family Holcobelidae (Coleoidea) in the European Jurassic: systematics, biostratigraphy, palaeobiogeography and evolutionary trends. Palaeodiversity, 5:13-49.
Werner, E. 1913. Über die Belemniten des schwäbischen Lias und die mit ihnen verwandten Formen des Braunen Jura (Acoeli). Palaeontographica, 59:103-146.
Whitehouse, F.W. 1924. Dimitobelidae - A new family of Cretaceous belemnites. Geological Magazine, 61:410-416.
Williamson, T. 2006. Systematics and biostratigraphy of Australian early cretaceous belemnites with contributions to the timescale and palaeoenvironmental assessment of the early Australian early cretaceous system derived from stable isotope proxies. Unpublished PhD thesis, James Cook University, Cairns, North Queensland, Australia.
Wright, D.F. 2017. Bayesian estimation of fossil phylogenies and the evolution of early to middle Palaeozoic crinoids (Echinodermata). Journal of Paleontology, 91:799-814. https://doi.org/10.1017/jpa.2016.141
Zhu, K.-Y. and Bian, Z.-X. 1984. Sinobelemnitidae, a new family of Belemnitida from the Upper Triassic of Longmenshan, Sichuan. Acta Palaeontologica Sinica, 23:300-325.
zu Münster, G. 1830. Bemerkungen zur näheren Kenntnis der Belemniten. Birner, Bayreuth.
