

 Review of the Pliocene and Pleistocene Talpidae (Soricomorpha, Mammalia) of Poland
Review of the Pliocene and Pleistocene Talpidae (Soricomorpha, Mammalia) of Poland
Article number: 17.2.26A
https://doi.org/10.26879/457
Copyright Palaeontological Association, July 2014
Author biography
Plain-language and multi-lingual abstracts
PDF version
Submission: 16 January 2014. Acceptance: 19 June 2014
{flike id=778}
ABSTRACT
Remains of Talpidae (Soricomorpha, Mammalia) from nine Pliocene and Early Pleistocene localities [Podlesice (MN14), Węże 1 (MN15) and 2 (MN15/MN16), Rębielice Królewskie 1A and 2 (MN16), Kielniki 3B, and Zamkowa Dolna Cave A (MN17), Kadzielnia 1 (MN17 or MN17/Q1) and Zamkowa Dolna Cave C (Q2)] located in Poland are reviewed. One new genus Skoczenia n. gen. and one new species Quyania europaea n. sp. are described. The list of Talpidae species from the Pliocene and Early Pleistocene of Poland now includes Condylura kowalskii, C. izaballae, Parascalops fossilis, ? Scalopoides sp., ? Neurotrichus polonicus, ? N. skoczeni, Quyania europaea n. sp., and Skoczenia copernici n. gen. Where necessary, previously described specimens were reevaluated and redescribed, and new illustrations were added. The systematic position, distribution, and number of species in each of the localities are discussed.
Barbara Rzebik-Kowalska. Institute of Systematics and Evolution of Animals, Polish Academy of Sciences, Sławkowska 17, 31-016 Kraków, Poland, rzebik@isez.pan.krakow.pl
Keywords: new genus; new species; fossil moles; Pliocene; Pleistocene; Poland
Final citation: Rzebik-Kowalska, Barbara. 2014. Review of the Pliocene and Pleistocene Talpidae (Soricomorpha, Mammalia) of Poland. Palaeontologia Electronica Vol. 17, Issue 2;26A; 26p. https://doi.org/10.26879/457
palaeo-electronica.org/content/2014/778-revision-of-talpidae
INTRODUCTION
Most of the Recent and fossil Polish Talpidae were studied by the late (1925-2007) Stanisław Skoczeń, professor of the University of Agriculture in Kraków. The first paper on fossil moles concerned the tribe Condylurini Gill, 1875 (Skoczeń 1976); the second Scaptonychini Van Valen, 1967, Urotrichini Dobson, 1883 and Scalopini Dobson, 1883 (Skoczeń 1980). The list of taxa described by Skoczeń includes: Condylura kowalskii Skoczeń, 1976 and C. izabellae Skoczeń, 1976, ? Geotrypus copernici Skoczeń, 1980, Scaptonyx (?) dolichochir (Gaillard, 1899), ? Neurotrichus polonicus Skoczeń, 1980, Scapanulus agrarius Skoczeń, 1980, Parascalops fossilis Skoczeń, 1993, and Neurotrichus minor Skoczeń, 1993.
The ascription of the Pliocene (MN 15 and MN16) remains from Węże 1 and 2 and Rębielice Królewskie 1A and 2 (Skoczeń, 1976) to Condylurini and the genus Condylura Illiger, 1811 was never questioned. On the other hand the assignment of the remaining mole specimens from the Pliocene and Pleistocene Polish localities (Skoczeń, 1980) was criticized by Storch and Qiu (1983). Most probably Skoczeń’s response was his paper of 1993 describing a new species ( Neurotrichus minor Skoczeń, 1993) and transferring specimens described in 1980 as Scapanulus agrarius Skoczeń, 1980 to the genus Parascalops and the new species, Parascalops fossilis Skoczeń, 1993.
However, this single transfer did not resolve all of the problems. The systematic position of Parascalops fossilis as well as other forms remained unclear and some authors (e.g., Dahlmann, 2001; Ziegler, 2003; Popov, 2004) still questioned taxonomic assignments and proposed different affiliations.
Until now, the author of the present paper never studied the mole material described by Skoczeń. In papers on the Pliocene and Pleistocene insectivore mammals from Poland (Rzebik-Kowalska, 1994, 2005, 2009) the names of moles were cited according to Skoczeń (1976, 1980, 1993) or as suggested by Storch and Qiu (1983). Now, for the first time, a revision of the Skoczeń material is undertaken.
The material originates from nine localities including Podlesice (MN14), Węże 1 (MN15), Węże 2 (MN15/MN16), Rębielice Królewskie 1A and 2 (MN16), Kielniki 3B (MN17), Zamkowa Dolna Cave A (MN17), Kadzielnia 1 (MN17 or MN17/Q1), and Zamkowa Dolna Cave C (Q2). The list of insectivore mammals found alongside the moles in these localities is given in Rzebik-Kowalska (2009).
MATERIAL AND METHODS
Unfortunately the material left by Skoczeń is incomplete. The number of specimens cited in his papers (Skoczeń, 1976, 1980, 1993) does not agree with the number of specimens found. These missing specimens are listed under “Material” concerning particular species. The detailed description of most specimens given by Skoczeń (1976, 1980, 1993) is good and is not repeated in the present paper except for situations in which new forms (new genera or new species) are created, or when new characters, not mentioned by Skoczeń (1976, 1980, 1993) were observed.
Measurements of specimens were taken according to Hutchison (1974). Homologous elements (e.g., the right first lower molar m1 or right humerus) were used to represent the minimum number of individuals (MNI). Talpidae teeth are measured on their occlusal surfaces. Abbreviations: n = number of specimens, L = maximum length, W = maximum width, DW = width of humerus diaphysis, sd = standard deviation, and cv = coefficient of variation.
The specimens discussed are housed in the collections of the Institute of Systematics and Evolution of Animals Polish Academy of Sciences (ISEAPAS) in Cracow and in the Institute of Paleobiology Polish Academy of Sciences (IPPAS) in Warsaw.
SYSTEMATIC PALAEONTOLOGY
Class MAMMALIA Linnaeus, 1758
Superorder INSECTIVORA sensu Novacek, 1986
Order SORICOMORPHA Gregory, 1910
Family TALPIDAE Fischer von Waldheim, 1814
Subfamily SCALOPINAE Gill, 1875
Tribe CONDYLURINI Gill, 1875
Genus CONDYLURA Illiger, 1811
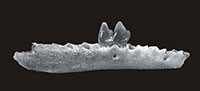
Condylura kowalskii Skoczeń, 1976
Figure 1.1-5, Figure 2.1-2
 1976 Condylura kowalskii Skoczeń; Skoczeń, p. 295, figs. 3, 4.
1976 Condylura kowalskii Skoczeń; Skoczeń, p. 295, figs. 3, 4.
1993 Condylura kowalskii Skoczeń; Skoczeń, p. 134, figs. 5, 6.
1994 Condylura kowalskii Skoczeń; Rzebik-Kowalska, p. 80, 86, 88.
2005 Condylura kowalskii Skoczeń; Rzebik-Kowalska, p. 126, 127, 128.
2009 Condylura kowalskii Skoczeń; Rzebik-Kowalska, p. 8, 19, 21, 22, 49, 86.
Material. Rębielice Królewskie 1A. Holotype - right humerus, no. MF/1006/16, left M1, left M2, right fragment of mandible with p1-p2, right fragment of mandible with m2, two left m1s, four left and two right m2s, as well as five right and seven left humeri. MNI = 7, catalogue number MF/1006. Rębielice Królewskie 2. Two right humeri. MNI = 2, catalogue number MF/1006b. Węże 1. Right M1, right fragment of mandible with m2, right and left humeri damaged in their proximal parts. MNI = 1, catalogue number MF/1005.
All material listed above and some postcranial bones (clavicle, ulna, radius, femur, astralagus, and calcaneus) not taken into account in this paper are listed and described in Skoczeń (1976) and are housed in the collection of the ISEAPAS in Kraków.
One m1, four mandibular fragments with m2 and m3 and 23 humeri of this species were also collected in Węże 2 and described by Skoczeń in his paper of 1993. The material of Węże 2 is housed in the IPPAS, Warsaw, catalogue number ZPAL/M-1.
In comparison with the description of Skoczeń (1976) one M1, one m1, one mandibular process, and one left humerus are lacking in the material. Furthermore in his paper of 1976, Skoczeń indicated a right humerus as the holotype of C. kowalskii (no. MF/1006/16) from Rębielice Krolewskie 1A, while in fact it is a left.
 Description. A description of detailed morphology of teeth and postcranial bones as well as measurements can be found in Skoczeń (1976, 1993). Here only the most characteristic tooth and mandible features are accentuated.
Description. A description of detailed morphology of teeth and postcranial bones as well as measurements can be found in Skoczeń (1976, 1993). Here only the most characteristic tooth and mandible features are accentuated.
The M1 has a long parastyle directed upwards and a high paracone and metacone. Its mesostyle is divided, and the ectoflexus and post-ectoflexus are deep. The lingual side of the tooth consists of a comparatively high protocon, and slightly lower paraconule and metaconule. They are separated by a shallow depression in the external and deeper depression in the internal sides of the crown. They form a continuous wall separated from the buccal part of the tooth by a deep valley. Cingula are lacking. The M2 is similar but its metastyle is shorter.
The p1 is laterally compressed and elongated. Its main cusp is situated in the anterior part of the crown. It is slightly convex on the buccal and flat on the lingual sides. Its anterocristid and posterocristid are rather blunt. A small parastylid is present. The posterior wall of the crown is slightly damaged. In general, the p2 is similar to the p1 but its parastylid is more distinct, the main cusp shorter and the talonid longer. Both teeth are devoid of cingula and they have two roots. In the mandible there is a space (diastema) between p1 and p2. It equals half of their (p1 and p2) length.
The m1 has a shorter and narrower trigonid than talonid. Its paraconid is usually rounded (in Figure 1.4 damaged), the hypoflexid very large and deep, and the crista obliqua ends near the tip of the metastylid, which is prominent. A big entostylid is also present. The m2 resembles m1 but its trigonid is longer and wider than the talonid and it has a parastylid. The teeth are devoid of cingula.
The mandible narrows in the anterior and posterior directions. It has a depression on the lingual side below p1 and p2. The mental foramen is situated below the anterior root of p2.
Systematic position and distribution. According to Skoczeń (1976) the remains listed above should be assigned to the genus Condylura Illiger, 1811. All tooth and mandible characters agree with those of extant Condylura cristata (Linnaeus, 1758), the only genus and species of the tribe Condylurini Gill, 1875 and one of two tribes of the subfamily Scalopinae Gill, 1875. The Recent species lives in marshy areas in the vicinity of the Great Lakes of the eastern United States and Canada.
On the other hand, 14 genera are included in the second tribe of this subfamily - Scalopini Gill, 1875 or directly to the Scalopinae. Recent forms include Scalopus Geoffroy Saint-Hilaire, 1803, Scapanus Pomel, 1848, Parascalops True, 1894 and Scapanulus Thomas, 1912. All live in the New World (Mexico, the USA, and Canada) with the exception of Scapanulus, which is known from China.
Teeth of Recent species of Scalopini differ from teeth of Recent and fossil Condylura by a single-rooted p1, while the p1 of Condylura has two roots. Moreover, the lower molars of Scalopus and Scapanus are devoid of metastylids, which are present in molars of Condylura.
The fossil genera of the Scalopini are represented by Proscapanus Gaillard, 1899 known from many European localities dated from the Middle to the Late Miocene (MN4-MN 11), Leptoscaptor Ziegler, 2003 described from the Middle Miocene of Germany and Hugueneya Van den Hoek Ostende, 1989 also described from Germany and known from other localities of that country dated to the Late Oligocene and early Middle Miocene (MP30 - MN2 and MN4). An Asiatic form, Yanshuella Storch and Qiu, 1983, known from the terminal Late Miocene or Early Pliocene and Yunoscaptor Storch and Qiu, 1991 known from the Late Miocene, are both found in China. Scalopoides Wilson, 1960 was described from the Middle Miocene to the Middle Pliocene of the USA. It was also reported in several localities in Europe (MN6-MN10 and MN15 - MN17) but the taxonomic status of these specimens is rather unclear. Other fossil genera described from the New World encompass Proscalops Matthew, 1901 known from the Middle Oligocene to the late Early Miocene, Mydecodon Wilson, 1960 from the Early Miocene, Scapanoscapter Hutchison, 1968 from the Late Miocene and Domninoides Green, 1956 from the Early Pliocene. All of these are from the United States.
The most characteristic features of the dentition of fossil and Recent Scalopinae genera are presented in Table 1. This list shows that the set of characters of Condylura teeth cannot be mistaken for the remaining genera of the subfamily.
Two other moles often compared with Condylura , i.e., Scaptochirus Milne-Edwards, 1867 (Talpinae, Talpini) from China and Scaptonyx Milne-Edwards, 1872 (Talpinae, Scaptonychini) from China, Burma, and Vietnam have two-rooted p1s, but in both forms this tooth is enlarged, while in Condylura it is always small.
An especially characteristic feature of Condylura, so far unique among moles, is the arrangement of its canine and premolars. The spaces (diastemae) between these teeth in upper and lower jaws are very large and they equal, more or less, half of the length of p1 or p2. In the fossil specimen (in the fragment of mandible with p1-p2) of C. kowalskii the space between p1 and p2 is 0.39 mm while the L p1 = 0.81 mm and L p2 = 0.80 mm.
Unfortunately, the measurements given here do not agree with those of Skoczeń (1976). According to him the space (diastema) between p1 and p2 in the same specimen equals 2.00 mm. However, this is completely impossible because the whole fragment of mandible in which teeth p1 and p2 are present equals 2.26 mm. In his paper of 1976 Skoczeń did not mention the length of these premolars but the length of lower molars given by him is 1.65 mm (n=2) for m1 and 1.60-1.73 mm (n=7) for m2, and premolars are always much smaller.
Nevertheless a space between antemolars (i3 - p4) of Condylura is unique among Scalopinae moles. The fragment of mandible with p1 - p2 found in the fossil material is identical to the mandible of Recent Condylura. It supports this identification and demonstrates that moles belonging to this genus lived in Europe during the Early Pliocene.
The differences in teeth between C. kowalskii and C. cristata are visible in m1 and m2, which are somewhat shorter and wider in fossil specimens and their m2 have parastylids (Skoczeń, 1976).
The origin of Condylura is unknown. Hutchison (1968, 1984) suggested that it came from Achlyoscapter Hutchinson, 1968 (Talpinae incertae sedis). Found in the Middle and Late Miocene of Oregon and the Late Miocene of Nebraska, it was generalized enough to be a possible ancestor of Condylura, especially of cf. Condylura sp. found in the Late Miocene or Pliocene sediments of Oregon. Its remains are so far the first evidence for condylurines in western North America and the oldest record of this mole genus at all.
It is unknown whether Condylura originated in North America and later dispersed to Europe or vice versa, it colonized North America from Europe
The Pliocene relatives of Condylura disappeared from Europe most probably because of competition with desmans (Desmaninae) which were larger, had stronger dentition and already lived in Europe at that time, and likely occupied the same ecological niche.
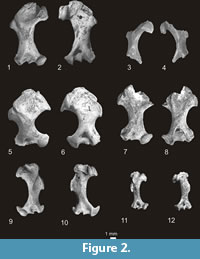
Condylura izabellae Skoczeń, 1976
Figure 2.3-4
1976 Condylura izabellae Skoczeń; Skoczeń, p. 305, fig. 5.
1994 Condylura izabellae Skoczeń; Rzebik-Kowalska, p. 80, 88.
2005 Condylura izabellae Skoczeń; Rzebik-Kowalska, p. 128.
2009 Condylura izabellae Skoczeń; Rzebik-Kowalska, p. 8, 22, 49.
Material. Rębielice Królewskie 1A. Holotype - one left humerus. MNI = 1, collection number MF/1007/1. It is housed in the collection of the ISEAPAS in Kraków.
In comparison with the description of Skoczeń (1976), four radii and one femur are missing in the material.
Description. A description of detailed morphology of the holotype and its measurements can be found in Skoczeń (1976).
Systematic position. C. izabellae is identical with C. kowalskii and differs from it and from C. cristata only in smaller size (Skoczeń, 1976). More material could confirm this identification.
Tribe SCALOPINI Gill, 1875
Genus PARASCALOPS True, 1894
Parascalops fossilis Skoczeń, 1993
Figure 2.5-6
1980 Scapanulus agrarius Skoczeń; Skoczeń, p. 440, fig. 5.
1993 Parascalops fossilis Skoczeń; Skoczeń, p. 127, fig. 3.
1994 Scapanulus agrarius Skoczeń; Rzebik-Kowalska, p. 80, 84, 86.
1994 Parascalops fossilis Skoczeń; Rzebik-Kowalska, p. 80, 84, 86.
2001 “ Scalopoides ” agrarius (Skoczeń); Dahlmann, p. 21, pl. 7, fig. 4.
2005 Parascalops fossilis Skoczeń; Rzebik-Kowalska, p. 123, 126.
2009 Parascalops agrarius (Skoczeń); Rzebik-Kowalska, p. 8, 16, 19.
2009 Scapanulus agrarius Skoczeń; Rzebik-Kowalska, p. 50.
Material. Podlesice. Holotype - a nearly complete left humerus no. MF/1018/24, paratype - the distal part of the right humerus, no. MF/1018/25. Remaining material: manubrium sterni and right ulna. MMI = 1, catalogue number MF/1018. Węże 1. Two incomplete humeri, left and right. MNI = 1, catalogue number MF/1019. They are housed in the collection of the ISEAPAS in Kraków.
According to Skoczeń (1993), a fragmentary right humerus, manubrium sterni, and ulna from Podlesice were previously identified as Scapanulus agrarius Skoczeń, 1980. The veracity of this statement is confirmed by the same catalogue number (MF/1018) of the specimens cited by Skoczeń in his papers of 1980 and 1993.
However, the material described by Skoczeń in 1980 as S. agrarius was more abundant. Besides the specimens mentioned above, the material also contained four astragali and 17 humeri. Their systematic position is now unclear because it is unknown whether Skoczeń (1993) left these specimens (four astragali and 17 humeri) to S. agrarius or whether he included them in P. fossilis. In the latter case specimens assigned to the mole known as S. agrarius cease to exist in the fossil fauna of Poland, and the fossil fauna in general and the name S. agrarius becomes a junior homonym of P. fossilis.
Unfortunately these questions cannot be resolved because in the latest paper of Skoczeń (1993) these remains (four astragali and 17 humeri) from Podlesice figure neither as S. agrarius nor as P. fossilis and they were not found in the material elaborated by Skoczeń (1980, 1993).
The holotype of S. agrarius was described as a “relatively less damaged” left humerus (Skoczeń, 1980) and that of P. fossilis as a “nearly complete” left humerus (Skoczeń, 1993). However, they cannot be the same specimens because their numbers are different, i.e., MF/1018/2 and MF/1018/24, respectively. These different numbers are not erroneous because the morphology and measurements of both holotypes are also different. The humerus length (L) of S. agrarius equals 8.52 mm and its proximal width (Wp) is 5.10 mm (Skoczeń, 1980), while the L humerus of P. fossilis equals 10.7 mm and its Wp is 7.7 mm (Skoczeń, 1993). A verification of measurements of the Podlesice material showed that the humerus indicated by Skoczeń in 1993 is the holotype of P. fossilis. The holotype of S. agrarius has not been found.
It is also unclear which humerus was chosen as the holotype for P. fossilis. It may have been one of the 17 humeri mentioned in the paper of 1980 (and now absent in the material) or a new one not cited earlier. Most probably the first supposition is correct because Skoczeń (1993) did not mention new material from Podlesice.
Description. The original description and measurements of Parascalops fossilis were given by Skoczeń (1993). Here the presence and the degree of development of the “scalopine ridge” on the humerus, characteristic for the Scalopinae family, is discussed.
Among 14 known Scalopinae genera, eight ( Proscapanus, Scapanulus, Scalopus, Scapanus, Yanshuella , Scalopoides, Yunascaptor, and Leptoscaptor) are characterized by a prominent or rather prominent “scalopine ridge” on their humeri. In one genus, Parascalops, this ridge is weak, fragmentary, or almost lacking. Known humeri of Domninoides and Scapanoscapter are incomplete and those of Mydecodon, Proscalops, and Hugueneya unknown. In this situation the presence or absence of a “scalopine ridge” is unknown. The humeri of fossil P. fossilis from Poland are characterized by a weak “scalopine ridge” similar to the ridge in the Recent Parascalops in which it is also faint.
Systematic position and distribution. The specimens mentioned above were previously described by Skoczeń (1980) as belonging to Scapanulus agrarius. However Storch and Qiu (1983) questioned its inclusion into this genus. According to them the Polish humeri differ from those of Recent Scapanulus oweni Thomas, 1912 from China, and they considered the generic status of Scapanulus agrarius undecided.
Most probably this critique caused Skoczeń (1993) to revise the systematic position of the S. agrarius remains and their inclusion into the genus Parascalops True, 1894. The only species of this genus, P. breweri (Bachman, 1842) (in Bachman and Charleston 1843-1844) now lives in the northeastern United States and southeastern Canada.
To confirm the new classification, Skoczeń (1993) listed 11 humeral characters common for Parascalops breweri and P. fossilis. Among them is a weak “scalopine ridge” present in both species. On the other hand he mentioned only five common characters of P. fossilis and Scapanulus oweni and among them also a weakly developed “scalopine ridge.” However, according to Storch and Qiu (1983) this ridge is not weak in Recent S. oweni and thus both forms have only four common features.
Only four characters discriminate between humeri of P. fossilis and Recent P. breweri and as many as eight delimit P. fossilis and Recent S. oweni (Skoczeń, 1993). This comparison shows that the specimens described earlier as Scapanulus agrarius are more similar to Parascalops than to Scapanulus (Skoczeń, 1980, 1993), and they should be included into the first genus ( Parascalops).
In 2001 Dahlmann named a mole from the Early Pliocene locality Wölfersheim in Germany as “ Scalopoides ” agrarius (Skoczeń, 1980). According to him these remains resemble Scapanulus agrarius Skoczeń, 1980 described from Poland and also compared by Skoczeń (1980, p. 444) to Scalopoides Wilson, 1960 because of some common characters in both forms. However, the fossil Scalopoides from the USA (Early Miocene, Colorado) is characterized by a prominent “scalopine ridge” which in P. fossilis (previously Scapanulus agrarius) is only very weak. The five humeri of “ Scalopoides ” agrarius from Germany are damaged. In this situation the degree of formation of their “scalopine ridge” is unknown and their inclusion into the genus Scalopoides is rather open. On the other hand, despite the absence of teeth among P. fossilis specimens, the close similarity of their ( P. fossilis andrecent P. breweri ) humeri, ulna, and manubrium shows that it represents a Parascalops -like mole and can be ascribed to the genus Parascalops.
The name of the species remains problematic. Previously (Rzebik-Kowalska, 1994, 2005, 2009) it seemed certain that P. fossilis was described on the grounds of the same material as S. agrarius , all specimens belonging to S. agrarius were included into the genus Parascalops and the new form should be named Parascalops agrarius (Skoczeń, 1993). Now, after a revision of the material (in which the holotypes of both forms are not the same specimens and most remains of Scapanulus agrarius have disappeared) the name P. fossilis Skoczeń, 1993 has been maintained. At the same time the species Scapanulus agrarius Skoczeń, 1980 and its name disappear from the Polish fossil fauna as well as from the fossil fauna in general.
Apart from Poland, fossil remains of Parascalops were found in Pleistocene localities of North America (Van Zyll de Jong, 1983).
Genus ? SCALOPOIDES Wilson, 1960
? Scalopoides sp.
Figure 2.7-8
1980 ? Geotrypus copernici Skoczeń; (partim), Skoczeń, p. 414, figs. 2, 3.
1994 “ Geotrypuscopernici ” Skoczeń; (partim), Rzebik-Kowalska, p. 80, 90, 91, 93.
2005 Geotrypus ? copernici Skoczeń; (partim), Rzebik-Kowalska, p. 129, 131.
2009 Scalopoides sp. =? G. copernici Skoczeń; (partim), Rzebik-Kowalska, p. 8, 24, 50.
Material. Kadzielnia 1. Four fragmentary humeri (two right and two left). MNI = 2, catalogue number MF/1008. Zamkowa Dolna Cave A. Three humeri (two left). MNI = 2, catalogue number Mf/1010. They are housed in the collection of the ISEAPAS in Kraków.
In comparison to the material of Skoczeń (1980), two humeri are lacking in Zamkowa Dolna Cave A.
Description. Done by Skoczeń (1980, p. 418) in the chapter on ? Geotrypus copernici nov. sp. The author noticed the elongated and narrow shaft of the humerus and the proximal position of its teres tubercle.
Systematic position and distribution. The abovementioned humeri together with mandibles and upper and lower teeth were described by Skoczeń (1980) as belonging to ? Geotrypuys copernici. However, Storch and Qiu (1983) and Dahlmann (2001) questioned the inclusion of this material to the genus Geotrypus Lavocat, 1951. They wrote that the humeri rather resembled the bones of Scalopoides described by Wilson (1960) from the Early Miocene of the USA and of Yanshuella found in China (the Late Miocene or Early Pliocene) than the European Geotrypus.
In general, however, they seem to be somewhat more slender than humeri of Yanshuella. The width of their shaft (DW) equals 2.24 - 2.45 mm (n=5) compared to Scalopoides, 2.01 - 2.33 mm (n=14), and Yanshuella, 2.50 - 2.90 mm (n=7). As their morphology points to close affinities with the genus Scalopoides (see Skoczeń, 1980, p. 421) the specimens cited above have been tentatively ascribed to ? Scalopoides sp.
It is, however, clear that these humeri cannot be united with teeth and mandibles described below as Skoczenia copernici and previously by Skoczeń (1980) under the name of “ G.” copernici. Their morphology does not resemble the morphology of the humerus in Talpini moles, while the number and the morphology of teeth certainly places them in the Talpini. Moreover the humeri described above cannot be united with teeth of Skoczenia copernici because they are too small. They are even smaller than humeri of Talpa minor Freudenberg, 1914 present in Kadzielnia 1and Zamkowa Dolna Cave A, while the teeth are as big as teeth of Talpa europaea Linnaeus, 1758 and also much more robust. As the humeri are incomplete (the heads and “scalopine ridge” are missing) they are only tentatively included in Scalopoides.
It is, however, very strange that as in the case of Skoczenia copernici in which humeri are lacking there are no teeth in the material which could be united with humeri of ? Scalopoides sp. In the studied material there are only teeth and humeri of much smaller forms such as Quyania and Neurotrichus and of the genus Talpa - Talpaminor and Talpa fossilis Pétenyi, 1864.
In Europe moles of the genus Scalopoides were reported in older localities in France (La Grive Saint-Alban, MN7/8, Hutchison,1974) and in Spain (Hostalets de Pierola Inferior, MN7/8, Hostalets de Pierola Superior, MN9, and Trinchera norte Autopista, MN10 (Van den Hoek Ostende and Furió, 2005).
Subfamily TALPINAE Fischer von Waldheim, 1814
Tribe NEUROTRICHINI Hutterer, 2005
? NEUROTRICHUS Günther, 1880
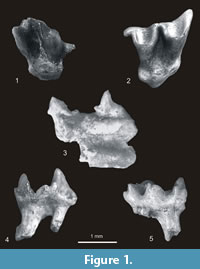
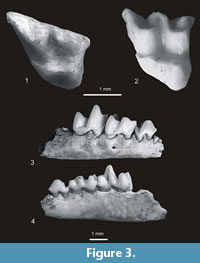
? Neurotrichus polonicus Skoczeń, 1980
Figure 2.9-10, Figure 3.1-4
 1980 ? Neurotrichus polonicus Skoczeń; Skoczeń, p. 427, pls. 5, 6.
1980 ? Neurotrichus polonicus Skoczeń; Skoczeń, p. 427, pls. 5, 6.
1993 Neurotrichus polonicus Skoczeń; Skoczeń, p. 133, fig. 4.
1994 “? Neurotrichus polonicus” Skoczeń; Rzebik-Kowalska, p. 80, 89, 90, 91.
2004 Quyaniapolonica (Skoczeń); Popov, p. 71, figs. 6, 7, 8.
2005 Neurotrichus ? polonicus Skoczeń; Rzebik-Kowalska, p. 128, 128, 130, 131.
2009 Neurotrichus polonicus Skoczeń; Rzebik-Kowalska, p. 9, 22, 24, 25, 26, 51.
Material. Rębielice Królewskie 1A. One right P4, two right and one left M1, one right M2, one right and one left m2, one right and one left m3, four fragments of mandibles (three left) with m1 - m3, three right and three left clavicles, eight right and five left humeri and right fragment of ulna. MNI = 8, catalogue number MF/ 1015. Kielniki 3B. One right humerus. MNI = 1, catalogue number MF/1020. Kadzielnia 1. Holotype - right fragment of mandible with roots of p3 and p4 - m2, no. MF/1016/1, right fragment of mandible with m1-m2, one right and one left humeri. MNI = 2, catalogue number MF/1016. Zamkowa Dolna Cave C. Two right and one left M1, one left M2, one right m2, one right and one left m3, one right humerus. Minimal number of individuals = 2, catalogue number MF/ 1017.
They are housed in the collection of the ISEAPAS in Kraków.
In comparison with the description of Skoczeń (1980) one right mandible with p3, one right m1, one right m2, and one radius are lacking in the materials from Rębielice Królewskie 1A and one m1 from Zamkowa Dolna C. Also two molars in Rębielice Królewskie 1A were incorrectly determined.
Description. A detailed description and measurements of the material are given in Skoczeń (1980, 1993). In general it is compatible with the present examination. There are, however, some differences. According to Skoczeń (1980) the upper molars of ? N. polonicus have protoconules while in reality they are absent except for one tooth from Rębielice Królewskie 1A in which it is vestigial. Skoczeń (1980, 1993) did not mention that some lower m1s from Rębielice Królewskie 1A have traces of mesoconids and small notches between the end of the crista obliqua and the posterior wall of their trigonid. He also did not point out the “scalopine ridge” in the humerus of ? Neurotrichus polonicus, which is quite prominent.
Measurements. See Table 2.
Systematic position and distribution. In his work of 1980, Skoczeń included these specimens in the genus ? Neurotrichus Günther, 1880 and the new species ? N. polonicus n. sp. Storch and Qiu (1983) indicated several characters of ? N. polonicus common with Recent American Neurotrichus gibbsi (Baird, 1858) and fossil Quyania chowi from the Late Miocene and the Early Pliocene of China described by them in the same paper. The dental formula of both genera ( Neurotrichus and Quyania) is, however, different. Neurotrichus has 36 teeth (I3/3; C1/1; P2/2; M3/3) and Quyania ?38-40 (I3/3; C1/1; P?3/3; M3/3).
Unfortunately the lack of antemolars anterior to P4 and p3 in ? N. polonicus complicate its correct identification and Storch and Qiu (1983, p. 101) “provisionally leave the taxonomic status (of this species) as given by Skoczeń unchanged.”
On the other hand Popov (2004) described a mole similar to ? N. polonicus from the Early Pleistocene (former MN17) locality of Varshets in Bulgaria and named it Quyania aff. polonica. He was of the opinion that some characters of ? N. polonicus (e.g., double rooted lower premolars and broad posterior wall of the protoconid in p3) mentioned by Storch and Qiu (1983) in Quyania from China speak in favor for its inclusion into the genus Quyania .
The revision of the specimens from Poland shows that the species ? N. polonicus combines characters of both genera, Neurotrichus and Quyania (Table 3) . It is as large as Neurotrichus and larger than Quyania (both teeth and humeri) and intermediate in morphology or closer to Quyania. Its first mental foramen in the mandible is situated anteriorly before p3 as in Quyania or below the first root pf p3 as in Neurotrichus. Some lower molars of specimens from Rębielice Królewskie 1A are characterized by vestigial mesoconids and small notches between the end of crista obliqua and the posterior wall of the trigonid as in Quyania. In the younger teeth from Zamkowa Dolna Cave C and Kadzielnia 1 the lower molars are devoid of mesoconids and notches as in Neurotrichus. On the other hand, the upper and lower teeth (Table 2) of ? N. polonicus are wider than the teeth of Neurotrichus, and similar to those of Quyania.
This comparison shows that, as mentioned by Storch and Qiu (1983), ? N. polonicus fits well in an ancestor - descendant relationship with Q. chowi and together with Q. showi belongs to one phyletic lineage of the Old World moles. However, its affiliation with Neurotrichus or Quyania is still open, and it is left tentatively as given by Skoczeń (1980, 1993).
Similar to the Recent Neurotrichus gibbsi, ? N. polonicus together with Q. showi and Q. europaea n. sp. discussed below should be included into the tribe Neurotrichini. In his first paper Skoczeń (1980) described specimens from Poland as ? Neurotrichus polonicus. In the second paper Skoczeń (1993) abandoned the question mark which was repeated by Rzebik- Kowalska in 2009. Now, after detailed analysis of the material and lack of teeth before p3, it seems clear that the question mark is still necessary and an inclusion of the Polish specimens into the genus Quyania by Popov (2004) is premature (see Q. europaea n. sp.). Assumed remains of fossil Neurotrichus (? N. columbianus) were described by Hutchison (1968) from Oregon.
? Neurotrichus skoczeni Zijlstra, 2010
1993 Neurotrichus minor Skoczeń; Skoczeń, p. 130, fig. 4.
1994 Neurotrichus minor Skoczeń; Rzebik-Kowalska, p. 80, 88.
2005 Neurotrichus minor Skoczeń; Rzebik-Kowalska, p. 127.
2009 Neurotrichus minor Skoczeń; Rzebik-Kowalska, p. 9, 21.
2010 Neurotrichus skoczeni Zijlstra; Zijlstra, p. 1903.
Material. Węże 2. Holotype - right humerus, no. ZPAL/M-2/2 and isolated left M1. MNI = 1, catalogue number ZPAL/M-2. The material is housed in the IPPAS in Warsaw.
Description. A detailed description, measurements, and photos of specimens are given in Skoczeń (1993).
Systematic position. The M1 and humerus described by Skoczeń (1993) as N. minor from the Polish locality of Węże 2 dated to the Early/Late Pliocene boundary (Ruscinian/Villanyan, MN15/16) are very similar to ? Neurotrichus polonicus but smaller.
In 2010 Zijlstra found that Dalquest and Burgner (1941) had already described a new subspecies of Recent Neurotrichus gibbsi as N. g. minor from western Washington in the US. In this situation, Skoczeń’s name, Neurotrichus minor,became a primary homonym and is invalid. In the place of Neurotrichus minor, Zijlstra (2010) proposed a new name, Neurotrichus skoczeni, which is accepted in this paper.
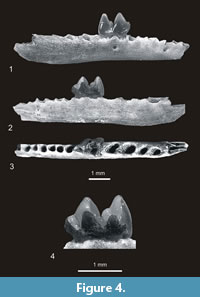
Genus QUYANIA Storch and Qiu, 1983
Quyania europaea n. sp.
Figure 2.11-12, Figure 4.1-4
zoobank.org/1C4F95AD-A2B4-479E-A158-ED243D48E58D
 1980 Scaptonyx (?) dolichochir (Gaillard); Skoczeń, p. 422, pl. 5.
1980 Scaptonyx (?) dolichochir (Gaillard); Skoczeń, p. 422, pl. 5.
1994 Scaptonyx (?) dolichochir (Gaillard); Rzebik-Kowalska, p. 80, 83, 86, 89, 92.
2005 Urotrichus ? dolichochir (?) (Gaillard); Rzebik-Kowalska, p. 123, 126, 127, 128, 129.
2009 Urotrichus sp.; Rzebik-Kowalska, p. 16, 19, 22, 26, 52.
Etymology. The species name “ europaea ” was given because it is the first species of Quyania found in Europe.
Holotype. Left mandibular fragment with m1 and alveoli of c-p4 and m2-m3, broken (and stuck) between p3 and p2, no. MF/1013/1. It is housed in the collection of the ISEAPAS in Kraków.
Type locality. Rębielice Królewskie 1A, Late Pliocene, Early Villanyian, MN16.
Studied localities. Podlesice, Early Pliocene (Early Ruscinian, MN14), Węże 1, Early Pliocene (Late Ruscinian, MN15), Kadzielnia 1, Early Pleistocene (Late Villanyan, MN17), or Pliocene/Pleistocene boundary.
Material. Podlesice. One left humerus damaged in its proximal part. MNI = 1, catalogue number MF/1011. Węże 1. Two fragments of left mandibles with m2-m3 and m1- m2. MNI = 2, catalogue number MF/1012. Rębielice Królewskie 1A. Three fragments of left mandibles with m1 and m2, two fragments of right mandibles with m1 and m2 and six (three left, three right) humeri, four fragmentary and two almost complete. MNI = 3, catalogue number MF/1013. Kadzielnia 1. One fragment of right mandible with m1-m2. MNI = 1, catalogue number MF/1014. All material is housed in the collection of the ISEAPAS in Kraków.
In comparison to the material of Skoczeń (1980), one isolated M2 from Rębielice Królewskie 1A and one m2 from Kadzielnia 1 are lacking. The mandible no. MF/1013/1 from Rębielice Królewskie 1A has 7 alveoli, not 8 as mentioned by Skoczeń (1980).
Diagnosis. A very small shrew-mole. Its mandible is tapered anteriorly. The first mental foramen is situated below p2. Lower molars are relatively high. Their crista obliqua terminates rather labially and is separated by a notch from the posterior wall of the trigonid. The mesoconid in m1 is present and metastylids on lower molars are lacking. Their talonid valley is closed by a well-developed ridge connecting the metaconid with the entoconid.
Differential diagnosis. Q. europaea differs from Q. chowi by a more slender shaft of the humerus, more delicate “scalopine ridge” as well as by a narrower precingulid in m1 and talonids of m1 and m2 closed by a well-developed ridge connecting the metaconid with the entoconid.
Description of the material. The description of the specimens cited as Scaptonyx (?) dolichochir are given in Skoczeń (1980). However, considering the description of a new species, the specimens mentioned above were measured once more.
Skoczeń (1980) did not mention that the mesoconids are present in all m1s and that small notches which separate crista obliqua from the posterior wall of trigonids occur on the lower molars. He also did not mention the “scalopine ridge” present in one humerus from Rębielice Królewskie 1A. The ridge is slightly damaged, but very clear. It is less visible in the second humerus from the same locality because this area of the specimen is destroyed. The remaining 11 humeri are incomplete, and the presence or absence of the “scalopine ridge” cannot be documented.
Measurements. See Table 2. Holotype: m1 - L=1.51 mm, W=0.92 mm.
Systematic position and distribution. The remains cited above were assigned by Skoczeń (1980) to Scaptonyx (?) dolichochir (Gaillard, 1899), a species originally described on the basis of a humerus from La Grive Saint Alban in France and dated to the Middle Miocene (MN7+8).
Because of the striking resemblance of the Scaptonyx dolichochir holotype (humerus) to the Recent forms, Urotrichus talpoides Temminck, 1841 from Japan, Hutchison (1974) tentatively transferred the species “ dolichochir ” to the genus Urotrichus Temminck, 1841.
Storch and Qiu (1983) called in question the assignment of the fossil Polish mole Scaptonyx (?) dolichochir to Scaptonyx because in comparison with the Recent form ( S. fusicaudatus Milne-Edwards, 1872) from Asia, the Polish specimens lack metastylids on the lower molars and the crista obliqua (in its m2 and m3) terminates more buccally than in Recent species. Storch and Qiu (1983, p. 105) wrote: “as long as there are no dentitions of the Miocene species dolichochir available we consider the inclusion of the Polish Pliocene form by Skoczeń as questionable.” Taking the above statement under consideration, Rzebik-Kowalska (2009) listed these specimens as Urotrichus sp.
However, after detailed revision of the material its association to the genus Urotrichus seems questionable. It is generally known that the mandible of Urotrichus is not tapered anteriorly, and its first mental foramen is situated below p3. On the other hand, mandibles from Poland tapered to the front, and the first mental foramen is situated below the posterior root of p2. Moreover, Recent Urotrichus has eight ( U. talpoides) or nine ( U. pilirostris True, 1886) teeth in half of the mandible, and among antemolars only p4 is two-rooted. In this situation, U. talpoides has 12 alveoli and six before m1, and U. pilirostris has 13 and seven before m1 (Abe, 1967; Storch and Qiu, 1983). One mandible (no. MF/1013/1) from Rębielice Królewskie 1A has seven alveoli preserved before m1 but it is broken in its anterior part and is visibly incomplete. It is thus quite clear that more alveoli were present in its broken anterior part, more than in both Recent species of Urotrichus ( U. talpoides and U. pilirostris) and in Neurotrichus gibbsii.
According to Storch and Qiu, 1983 nine teeth (3i, 1c, 2p, 3m) and 14 alveoli (eight in antemolars in which p3 and p4 are double-rooted) are present in the Recent species N. gibbsii (Baird, 1858). This is only one more than in the mandible from Rębielice Królewskie 1A (no. MF/1013/1), which must have been longer and thus must have also had more alveoli in its anterior part.
Taking these facts under consideration, specimen no. 1 from Rębielice Królewskie 1A could belong to Quyania, the mandible of which has 10 teeth (3i, 1c, 3p, 3m) and 16 alveoli (10 before m1) in its row. Although mandibles of Q uyania and Neurotrichus taper anteriorly, Quyania has more teeth (and alveoli) before m1 and its first mental foramen is situated below p2, not below p3 as in the mandible of Neurotrichus. Moreover, two other characteristic features distinguish Quyania from Neurotrichus. The m1 of Quyania is characterized by a clear mesoconid. This character was not mentioned by Skoczeń (1980) or by Storch and Qiu (1983). It is, however, very well visible in the m1 of Quyania in the paper of Storch and Qiu (1983, table 1, no. 12) and in all (five) m1 present in the Polish material assigned by Skoczeń (1980) to Scaptonyx (?) dolichochir. Also the crista obliqua on the m1 as well as on the remaining lower molars from Poland [ascribed previously to Scaptonyx (?) dolichochir ] is separated from the posterior wall of trigonid by a small but clear notch that is also characteristic of the Quyania molars (and rather not of Neurotrichus).
The remaining characters of the mandible, teeth, and humerus [e.g., presence of the “scalopine ridge” not mentioned by Skoczeń (1980)] as well as their size also agree with the description and size given by Storch and Qiu (1983) for Quyania. In this situation the remains from Polish localities described by Skoczeń (1980) as S. (?) dolichochir are now included into the genus Quyania Storch and Qiu, 1983.
Unfortunately the species name “ dolichochir ” discussed by Storch and Qiu (1983) cannot be used in the case of the specimens (especially teeth) from Poland (see above). Because they slightly differ from those of Quyania chowi (see differential diagnosis) and there is a large gap in time and ranges between both forms, Quyania from Poland cannot be ascribed to Q. chowi. It is thus recognized as belonging to a new species and named Q. europaea.
Popov (2004) described Quyania aff. polonica (Skoczeń, 1980) from the Early Pleistocene (previously Late Pliocene, MN17) locality at Varshets in Bulgaria based on the description of ? Neurotrichus polonicus from Poland (Skoczeń, 1980). He also decided that Polish specimens are more similar to Quyania than to Neurotrichus and their characters “speak in favor of its inclusion in Quyania. ”
However, specimens from Varshets are larger (Table 2) than the specimens of Q. chowi and Q. europaea and in opposite to these two species, its first mental foramen is placed under p3 (in Quyania under p2) and its lower molars are devoid of mesoconids, which are present in the molars of Quyania. Thus, these size and morphological differences make the species designation of the Varshets specimens to Quyania problematic.
Tribe TALPINI Fischer von Waldheim, 1814
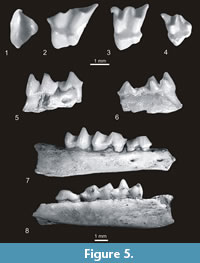
Genus SKOCZENIAn. g.
Skoczenia copernici (Skoczeń, 1980)
Figure 5.1-8
zoobank.org/EE3533BF-8577-4EC4-AADD-1643CAB3944D
 1980 ? Geotrypus copernici Skoczeń; (partim), Skoczeń, p. 414, pls. 3, 4, figs. 2, 3.
1980 ? Geotrypus copernici Skoczeń; (partim), Skoczeń, p. 414, pls. 3, 4, figs. 2, 3.
1994 “ Geotrypuscopernici ” Skoczeń; (partim), Rzebik-Kowalska, p. 80, 90, 91, 93.
2005 Geotrypus ? copernici Skoczeń; (partim), Rzebik-Kowalska, p. 129, 131.
2009 Scalopoides sp. =? G. copernici Skoczeń; (partim), Rzebik-Kowalska, p. 8, 24.
Etymology. The genus is named in honour of late professor Stanisław Skoczeń from Kraków (Poland) who passed away in 2007.
Holotype. Fragment of right mandible with p4-m2, last alveolus of p2 and two alveoli of p3 and of m3 (the same specimen as in the species “ copernici ”, no. MF/1008/17). It is housed in the collection of the ISEAPAS in Kraków.
Type locality. Kadzielnia 1.
Studied localities. Kadzielnia 1, Early Pleistocene (Late Villanyan, MN17 or Pliocene/Pleistocene boundary); Zamkowa Dolna Cave A, Early Pleistocene (Late Villanyan, MN17).
Material. Kadzielnia 1. Two fragments of left mandible - one with m1-m2, and one with p4, two toothless fragments of mandibles (left and right), and isolated teeth - two right P4, and two left M1. MNI = 3, catalogue number MF/1008. Zamkowa Dolna Cave A. Six (two right and four left) fragments of mandibles with p4-m2, isolated teeth - five P4 (three right), ten M1 (seven left), seven M2 (four left), five M3 (left), four p4 (three right), ten m1 (eight left), five m3 (four left). MNI = 8, catalogue number MF/1010. All material is housed in the collection of the ISEAPAS in Kraków.
In comparison to Skoczeń (1980) the material in Kadzielnia 1 lacks isolated teeth: one C1, one M2, one p4, four m1 and four m2. In Zamkowa Dolna Cave A one C1, three M3, one m1 and four m3 are missing. Teeth assigned as C1 are present in material from both localities but they are not the canines of moles.
Humeri from Kadzielnia 1 and Zamkowa Dolna Cave A cited by Skoczeń (1980) as “ G ”. copernici cannot be linked together with teeth of “ G ”. copernici as proposed by Skoczeń (1980) and they are tentatively assigned to ? Scalopoides sp. (see above).The mandible from Kamyk (Skoczeń 1980) also cited as belonging to “ G ”. copernici represents T. fossilis Petényi, 1864 (Rzebik-Kowalska, 2009).
Diagnosis. A large species of Talpini mole with massive teeth, very well-developed parastyles, and clearly divided mesostyles in upper molars. Its dental formula of the mandible is 3i, 1c, 4p, and 3m. The lower canine is small and one-rooted, all lower premolars double-rooted, the p1 enlarged, and p2 and p3 small, almost equal in size, lying slightly obliquely in the mandible with their anterior roots situated more lingually. The lower p4 is long and lower molars diminish in size from m1 to m3 (m1>m2>m3). The horizontal ramus of the mandible tapers anteriorly, and its two mental foramina are situated below the p2/p3 junction and below the posterior root of p4.
Differential diagnosis for the genus. In spite of an unreduced (complete) dental formula in the mandible (22 teeth) and large size as in most Pliocene and Pleistocene European Desmaninae Thomas, 1912 Skoczenia n. g. differs from these by the morphology of upper molars which are devoid of a protoconule (present in Desmaninae), by a large p1 (small in desmans), by the p4 devoid of a metaconid (present in desmans) and by the oblique cristid of the lower molars, which ends buccally (in Desmaninae situated either lingually against the tip of the metaconid or against the protoconid-metaconid crest) (Rümke, 1985).
Gaillardinae Hutchison, 1968 - aquatic desman-like moles from North America - also differ from Skoczenia n. g. by morphology of their mandibles in which a posterior mental formen is situated below the posterior root of m1 (in Skoczenia n. g. it is situated below the posterior root of p4), and by lower molars in which the oblique cristid reaches the lingual side and the metastylids (Hutchison, 1968). The teeth of Skoczenia n. g. are devoid of metastylids and the oblique cristid is short and only reaches the buccal (m1) or middle sides of the teeth (m2, m3).
Large fossil Suleimaninae Van den Hoek Ostende, 2001 from Anatolia (Turkey) with reduced dental formulae (only two upper molars present in jaw, Van den Hoek Ostende, 2001) cannot be taken into consideration because of the several very prominent M3s present among the specimens of Skoczenia n. g. from Kadzielnia 1 and Zamkowa Dolna Cave A.
The large size, complete lower dentition (22) and the hypertrophy of p1 with two roots (which can be concluded from mandible no. MF/1010/56) exclude Skoczenia n. g. from the Scalopinae [both Condylurini ( Condylura) and Scalopini ( Parascalops, Scalopus, Scapanus, Scapanulus, Hugueneya, Proscapanus, Leptoscaptor, Yanshuella, Yunoscaptor, Scalopoides, Mydecodon, Scaptonoscaptor, Proscalops, and ? Domninoides)]. Scalopinae, known from Eurasia and North America, are generally small, they have a reduced dental formula, their p1 is small with one root or this tooth is absent. If the dental formula of some genera of Scalopinae is unreduced and p1 has two roots as in Condylura or in Scaptonoscaptor , the first form is much smaller than Skoczenia n. g., its p1 is small and the lower molars have metastylids, and the second, Scaptonoscaptor, although large, also has a small p1 and small metastylids absent in Skoczenia n. g. Hugueneya has a two-rooted p1 as in Skoczenia n. g., but it is not very large and moreover its m1 is smaller than the m2 whereas in Skoczenia n. g. the m1 is visibly larger than the m2 (Van Zyll de Jong, 1983; Wilson, 1960; Hutchison, 1968; Storch and Qiu, 1983, 1991; Van den Hoek Ostende, 1989; Ziegler, 2003).
The small shrew-moles - Uropsilinae Dobson, 1883 from Eurasia and North America include genera such as Uropsilus Milne-Edwards, 1871, Mystipterus Hall, 1930, Desmanella Engesser, 1972, Theratiskos Van den Hoek Ostende, 2001, Mygatalpa Schreuder, 1940, Nasillus Thomas, 1911, and Rhynchonax Thomas, 1912, with small or absent p1s are also quite different from Skoczenia n. g. and can be easily excluded as its relatives.
Skoczenia n. g. also differs from the old (Oligocene - early Miocene) extinct American moles of the subfamily Proscalopinae K.M. Reed, 1961 raised by Barnosky (1981) to family (Proscalopidae K.M. Reed, 1961) level by having an unreduced dental formula (in Proscalopidae only one species, Proscalops tertius K.M. Reed, 1961, has an unreduced formula, Barnosky, 1981), by the presence of bifid mesostyles in upper molars, by the absence of metastylids in lower molars, by enlarged p1 and by m1>m2.
The subfamily Talpinae consists of tribes Urotrichini Dobson, 1883, Scaptonychini Van Valen, 1967, Neurotrichini, and Talpini.
Eurasiatic Urotrichini [( Urotrichus Temminck, 1841 = Dymecodon True, 1886), Myxomygale Filhol, 1890, Paratalpa Lavocat, 1951, Tenuibrachiatum Ziegler, 2003, Urotrichus ( Scaptonyx) dolichochir (Gaillard, 1899)] differ from Skoczenia n. g. by small size and reduced dental formula, i.e., 36 - 42 teeth. Only Myxomygale hasa fullset of teeth but its p1 is small, one-rooted, its m2>m1, and upper and lower molars have distinct cingula (Ziegler, 1990). The enigmatic Urotrichus (? Scaptonyx) dolichochir Gaillard, 1899 described on the grounds of the humerus is also smaller and quite different in morphology.
Asiatic Scaptonychini with one genus Scaptonyx Milne-Edwards, 1872 (Recent form - S. fusicaudatus Milne-Edwards, 1872) can also be excluded. This form is large and it has a double-rooted p1 as Skoczenia n. g., but the dental formula of its mandible is reduced (3,1,3,3, Gureev, 1979), and its lower molars have metastylids which are absent in Skoczenia n. g.
Although this character (the presence or absence of metastylids) seems unstable because in 14 genera of Scalopini seven ( Parascalops, Scapanulus, Proscapanus, Yunoscaptor, Domninoides, Scalopoides, Scapanoscapter) have metastylids and seven do not ( Scalopus, Scapanus, Hugueneya , Leptoscaptor, Yanshuella, Mydecodon, and Proscalops), Scaptonyx additionally differs from Skoczenia n. g. by the lack of a tooth cingulum, a different position of crista obliqua in the lower molars and by a larger m2 than m1.
The fossil “ Scaptonyx ” edwardsi Gaillard, 1899 has teeth and mandible similar to the Recent Scaptonyx (Gureev, 1979) and it is also different from Skoczenia n. g.
The North American and Eurasian Neurotrichini ( Neurotrichus, Quyania) are also small shrew-like moles with reduced teeth (mandible dental formula = 3-1-2-3 in Neurotrichus and 3-1-3-3 in Quyania), and thus they also cannot be united with Skoczenia n. g.
Skoczenia n. g. is most similar to representatives of the tribe Talpini. The tribe comprises such genera as Euroscaptor Miller, 1940, Mogera Pomel, 1848 (= Nesoscaptor Abe, Shiraishi and Arai, 1991), Parascaptor Gill, 1875, Scaptochirus Milne-Edwards, 1867, Geotrypus Lavocat, 1951, and Talpa Linnaeus, 1758. All Recent forms are known from Asia with the exception of Talpa living in Eurasia. They are characterized by a large or very large p1 with two roots as in Skoczenia n. g. They have, however, a reduced dental formula ( Parascaptor and Mogera 42, Scaptochirus and Geotrypus, e.g., G. ehrensteinensis 40 and Nesoscaptor (now included into Mogera) 38, with the exception of the Recent Euroscaptor (included by some authors in Talpa) and Talpa which have 44 teeth. However, in comparison to Skoczenia n. g., Euroscaptor is smaller and has a very small p2, much smaller than p3 (Abe et al., 1991).
Representatives of the genus Mogera have the mental foramen of the mandible situated between the roots of m1 and its m2>m1 which differentiates them from Skoczenia n. gen. Scaptochirus has large molars and lower molars diminishing in size from m1>m2>m3 as in Skoczenia n. g., but contrary to the latter, a very small, one rooted p3 and very narrow pointed coronoid process (Abe et al., 1991). In contrast to Skoczenia n. g., Parascaptor is rather small, it has p2 and p3 in one line parallel to the long axis of the mandible and its p2 is very small, considerably smaller than p3.
Besides the dental formula, which in some species is unreduced, Geotrypus differs from Skoczenia n. g. by the lack of a protoconule, metaconule, and metastyle in upper teeth and by its mesostyle only slightly divided or undivided. In addition, its lower molar m1 is always smaller or equal to m2 and its crista obliqua is situated more lingually (Crochet, 1995; Van den Hoek Ostende, 2001).
Skoczenia n. g. also differs from Recent and fossil species of Talpa. Its teeth are much stronger and contain a cingulum below the hypoflexid of the lower molars and in the posterior walls of the p4 and P4 as well as on the buccal side of M1 above the metacone. Its upper teeth have deeply fissured mesostyles, distinct metaconules, its lower p4 is elongated, lower molars with mesoconids and grooves situated between the oblique cristids and the posterior wall of the trigonids, posterior mental foramen situated below the posterior root of p4 and the tip of the coronoid process narrow and curved posteriorly.
Fossil Talpinae incertae sedis, e.g., Desmanodon Engesser, 1980 known from the Miocene of Europe and Asiatic Turkey, the Early Miocene Nuragha de Bruijn and Rümke, 1974 - endemic to Sardinia and only known from isolated teeth as well as Eotalpa anglica Sigé, Crochet and Insole, 1977 from the Middle - Late Eocene of Wight Island, apart from their different morphology, are much smaller.
Description of holotype. It is represented by a fragment of mandible with p4-m2 and alveoli of m3 as well as two alveoli of p3 and second alveolus of p2. The horizontal ramus of the mandible is heavy and tapered anteriorly. There are two mental foramina, one beneathp2/p3 and the other below the posterior root of p4. The alveoli of the p3 are slightly oblique and its anterior roots are situated slightly lingually.
Teeth are massive but unfortunately very worn. The p4 is very long and narrowed in the middle. Its protoconid is situated more or less in the middle of the tooth, the antero-labial side is convex, the postero-labial side is slightly basined, and the lingual side is almost straight. The anterocrista (paracristid) is sharp, the posterocrista very worn so it is unknown if the metaconid was present. The talonid is very short. The parastyle and cingula are lacking.
The m1 is also very long and its trigonid is longer than the talonid. The crista obliqua terminates bucally, below the protoconid, the hypoflexid is shallow and a trace of the cingulum is visible below it, the entoconid is wide, the entostylid rather small, and metastylid and remaining cingula absent.
The m2 is smaller than the m1, and its trigonid and talonid are more or less of the same length. Crista obliqua terminates more lingually, in the middle of the posterior wall of the trigonid (at the level of the protocristid notch), and the hypoflexid is deeper and cingulum below it more prominent. The parastylid and entostylid are present, and the metastylid and cingula are absent.
Description of the remaining material. A detailed description is in Skoczeń (1980). Here some of the most characteristic features are repeated and considering the description of the new genus the specimens were measured once more.
P4 is robust, its paracone is high, rounded anteriorly. The sharp posterior ridge extends from its tip to the posterior cingulum and is terminated by a small cuspule. The protocone is well developed, the parastyle in some specimens is also well developed but in others less so. A feeble cingulum surrounds the tooth with the exception of the protocone. It is more protruding in the buccal than in the lingual side.
The M1 has a very large parastyle, a deeply divided mesostyle, and large protocone and metaconule. A protoconule is lacking. The cingulum is also absent with the exception of the fragment on the buccal side beyond the metacone. The M2 has a large parastyle, large divided mesostyle, large metaconule, and very small protoconule.The M3 is similar to M1 and M2 with a strongly divided mesostyle, large parastyle, and a trace of a protoconule, but the cingulum is absent.
In the other mandible with unworn teeth (p4 - m2), the p4 is also long but its anterocrista is less sharp, talonid slightly longer, and a parastyle or a fragment of anterior cingulum is visible. In m1 a small mesoconid and a notch (between crista obliqua and posterior wall of the trigonid) are present, and the cingulum below the hypoflexid is more developed. Mesoconids and grooves are also visible in the m1 of the mandibles from Zamkowa Dolna Cave A (MF/1010/, 58 - 60).
The most important is mandible no. MN/1010/56. It bears a p4, the first alveolus of m1 and alveoli before p4. There are four alveoli of p3 and p2 more or less equal in size and placed slightly obliquely to the long axis of the mandible, with the first root of both teeth deflected to the lingual side. Before them are two large alveoli of p1, and subsequently four for c (the largest of these four) - i1.
Measurements. See Table 4.
Systematic position. Mandible No. 56 from Zamkowa Dolna Cave A and the remaining material from this locality as well as from Kadzielnia 1 are homogeneous and represent the same mole species as the holotype from Kadzielnia 1. The long p4 and the posterior mental foramen of the mandible situated below the posterior root of p4, characters unknown in other genera, discriminate this form (as shown above) from other moles known so far. The large size, 22 teeth in the mandible, double-rooted lower premolars, small lower c, and enlarged p1 allow for an unambiguous assignment to the tribe Talpini. However, the remaining characters do not allow its inclusion into Recent and fossil genera of Talpini.
Within the Talpini an unreduced tooth formula is present only in Euroscaptor and Talpa ; however, the differential diagnosis shows that both forms also differ in morphology from Skoczenia n.g. In this situation the massive teeth and mandibles from Kadzielnia 1 and Zamkowa Dolna Cave A were described as a new genus, and the species “ copernici ” as established by Skoczeń (1980). The humeri previously associated with these teeth by Skoczeń (1980) were transferred to the genus ? Scalopoides. It is strange, however, that humeri corresponding in size to the Skoczenia teeth were not found in the material.
DISCUSSION AND CONCLUSION
European moles (Talpidae) are considered as newcomers from Asia in the course of the major terrestrial Eurasiatic faunal turnover called the “Grande Coupure” and associated with the overall climate shift at the Eocene/Oligocene (MP18-MP21) transition. Previously uncommon in Europe they began to diversify during the end of the Middle and the Late Oligocene, most probably together with changes in the environment, e.g., with advancing aridity and steppe zones with gramineous plants and with the spread of species of open areas (Sigé et al., 1977).
A great diversity of mole species is known from Miocene and Pliocene localities of Europe. For example, eight species were found in German localities Ulm-Westtangente (MN2), seven in Petersbuch 2 (MN4) and Sandelzhausen (MN5), and as many as 11 in Wölfersheim (MN15) (Ziegler et al., 2005). A large number of Talpidae are also known from Schernham b. Haag in Austria (MN10), where 10 species were found (Ziegler and Daxner-Höck, 2005), from La Grive Saint-Alban in France (MN7/8, Guerin and Mein, 1971), Anwil in Switherlands (MN7/8, Engesser, 2005), Dolnice in the Czech Republic (MN4), and Devínska Nová Ves in Slovakia (MN6) (Fejfar and Sabol, 2005) where six species were found. Five species of moles were found in Tegelen in The Netherlands (MN16/17, Reumer, 2005), Františkove Láznĕ in The Czech Republic (MN5, Fejfar and Sabol, 2005), Mikhailovka 2 in Ukraine (MN11, Rzebik-Kowalska and Nesin, 2010) and Betfia IX and VII/1 in Romania (Early Pleistocene, Rzebik-Kowalska, 2000). Moreover, during Miocene and Pliocene many other European localities (from Spain to Russia) yielded three to four moles as well.
In Poland during the Miocene only four mole species were described in Bełchatów B (MN 5 or MN5/6) and three in Bełchatów A (MN7/8 or MN9) (Rzebik-Kowalska, 2009) but as seen from the Table 5 moles in the Pliocene Polish localities were more diverse.
Seven forms were excavated in Węże 1 and Rębielice Królewskie 1A and six in Podlesice and Kadzielnia 1. On the other hand, Zamkowa Dolna Cave C (Q2) is the youngest Polish locality where other genus (? Neurotrichus) than Talpa Linnaeus, 1758 (the only mole living today in Poland) was present, although there are many older other sites (the Early Pleistocene - MN17, Q1) from which only Talpa is known.
Similar to Recent moles, the fossil Talpidae likely included “true” moles adapted to deep and intense burrowing (Talpinae), forms less adapted to fossorial habitats (Uropsilinae) and those adapted to an aquatic or semi-aquatic life (Desmaninae). However, from the ecological point of view such a great number of mole species in one locality seems unprobable.
An analysis of the ecological requirements made on the grounds of the actualistic and morphological approaches shows that several moles could probably have lived in a mosaic terrain not far one from another if they were different in size and inhabited different biotopes. For example, in Węże 1 the following species may have existed side by side: Parascalopsfossilis - a large and probably intense burrowing mole as its descendant, Parascalops breveri, living today in eastern North America as well as two other fossorial moles of the genus Talpa, smaller than P. fossilis, T. Fossilis, and the smallest T. minor. Furthermore, in Węże 1 Quyania europaea, Condylura kowalskii, Galemys sulimskii Rümke, 1985, and Desmana nehringi Kormos, 1913 were excavated. According to Storch and Qiu (1983), the small Quyania chowi was related to the Recent shrew-mole Neurotrichus gibbsii, the smallest mole of the New World and adapted to a semi-fossorial mode of life. It is quite probable that the similar Quyania ( Q. europaea) also lived on forested hillsides and valley bottoms with moist loose soil and abundant leaf litter and constructed runways between the surface litter and the soil. On the other hand, the remaining three species were adapted to an aquatic or semi-aquatic lifestyle. They differ in size; Condylura kowalskii was the smallest, Galemys sulimskii was larger, and Desmana nehringi wasthe largest. Today, species of these genera live in different parts of the World: Condylura in North America, and Galemys Kaup, 1829 and Desmana Gueldenstaedt, 1777 at the western and eastern ends of Europe. They prefer slightly different biotopes so possibly they could live together. Condylura cristata is most abundant along small streams but it is also found in wet fields. The Recent Pyrenean desman Galemys pyrenaicus (Geoffroy Saint Hilaire, 1811) is found along swift-flowing streams, and Desmana moschata (Linnaeus, 1758) prefers quiet freshwater streams, lakes, and ponds of the Volga, Don, Oka, and Ural drainage areas.
In Rębielice Królewskie 1A we found four species associated with water bodies - two small Condylura (the larger C. kowalskii and smaller C. izabellae ), the middle sized Galemys kormosi,and large Desmana nehringi. Besides, there were also two? semi-fossorial species - the small Quyania europaea and larger ? Neurotrichus polonicus and one typical digger, the small Talpa minor.
In Podlesice there was one species less (six) which included the small fossorial Talpa minor and larger Parascalops fossilis, two aquatic or semi-aquatic species - the small Roumkelia aff. gettica (Terzea, 1980) and larger Desmana nehringi, one semi-fossorial shrew-mole, Quyania europaea, and one more shrew-mole Desmanella aff. dubia Rümke, 1976. The last form is assigned to the most primitive Talpidae subfamily, to Uropsilinae (Rümke, 1976; Engesser, 1980; Van den Hoek Ostende, 2001). Recently only one genus, Uropsilus (Asiatic shrew-mole) with four gracile species, lives in China and northern Burma. They inhabit forests and the alpine zone at elevations of 1250 - 4500 metres. Possibly fossil Desmanella was also connected with higher parts of the area.
At Kadzielnia 1 six moles were found including three species of fossorial moles (the largest and most robust Skoczenia copernici, the slightly smaller and gracile Talpa fossilis, and the smallest T. minor). There were also two semi-fossorial (larger? Neurotrichus polonicus and smaller Quyania europaea) and one semi-fossorial or semi-aquatic mole, ? Scalopoides sp. The last form was originally described from the Early Miocene of USA as a Scalopinae mole, Scalopoides isodens Wilson, 1960. According to Wilson (1960, p. 46) it “might be described as a Condylura -like mole with higher-crowned teeth and with antemolar dentition specialized toward Neurotrichus, or perhaps better as a Neurotrichus -like mole, somewhat more specialized in molar dentition and more fossorial than the living species. It could have been semiaquatic as well.”
The moles cited above belong to eleven genera. Among them four represent fossil or Recent American forms ( Condylura, Parascalops, Scalopoides, Neurotrichus) one Asiatic ( Quyania), two Euroasiatic ( Desmanella, Talpa), three European ( Roumkelia , Galemys, Desmana) and one of unknown origin ( Skoczenia).
As mentioned above, although the taxonomic richness of Talpidae in the Miocene and Pliocene is generally known, ecologically these six to seven mole species found in one locality seems excessive, especially since the differences in size between them and in biotopes in which they lived are often insignificant.
In this situation 11 species ( Desmanella woelfersheimensis Dahlmann, 2001, Talpa fossilis, Talpa gilothi Storch, 1978, Talpa minor, Archaeodesmana acies Dahlmann, 2001, Desmana cf. amutriensis Rădulescu, Samson, Štiucă, 1989, Desmana nehringi, Storchia wedrevis Dahlmann, 2001, “ Scalopoides ” agrarius (Skoczeń, 1980), “ Scalopodes ” cf. copernici (Skoczeń, 1980), and Talpinae gen. et sp. indet.) found in Wolfersheim are unlikely to have been sympatric.
Rich fossil remains (taphocoenoses) reflect ancient biocoenoses but most probably in the case of Wolfersheim or Węże 1, the biocoenoses comprised a larger area than only the vicinity of the localities in which they were found. In general, three possibilities regarding the accumulation of small vertebrate remains in sediments are considered. The first are vertical caves, the second transport and accumulation of remains by water, and the third points to predators (mainly owls) as the main factor leading to bone accumulation. Of course, these factors are not mutually exclusive, because owl pellets or carnivore scats can later be transported by water and be found in caves or in other types of localities. Vertical caves (as in Węże 1) could function as pit-falls in which the remains of animals of all sizes are accumulated (Sulimski, 1959).
In Wolfersheim, according to Dahlmann (2001), fragmentary material in the form of isolated teeth, single bones, and bone-fragments as well as the presence of aquatic vertebrates suggests a fluvio-lacustrine origin of the locality. However, the main source of microvertebrates in the sediments of many fossil localities is generally recognized as owl activity. As the hunting territory of some owls can be large [about 60 hectares for a pair of Barn owls according to Webster (1973)], some species of owls migrate more or less regularly or are nomadic in some seasons, therefore the distance that the prey items are transported to the place of regurgitation and subsequent deposition in sediments can also be great. According to Goszczyński (1976) the average time of pellet formation in the Barn owl ( Tyto alba) is about 15 ½ hours. Love (1980) using stained food showed that in Tawny owl ( Strix aluco) some bones were retained for 24 hours and a few as long as two days. There is therefore the possibility that the bones and teeth of prey taken at one place will be deposited in owl pellets in a remote region, sometimes outside the range of the particular prey species. This is particularly likely in the case when the number of remains found in a locality is small (one humerus or some isolated teeth), and indicates that either the prey species was very rare in the hunting territory or it was brought from a distant site. This shows that we should be careful in using small mammals in the reconstruction of past environments because it is probable that not all species were caught by owls or transported by water from the nearest vicinity.
ACKNOWLEDGEMENTS
I am grateful to Mr. A. Pereswiet-Soltan for preparing the illustrations and to Dr. M. Sabol and an anonymous referee for their insightful comments that improved the manuscript.
REFERENCES
Abe, H. 1967. Classification and biology of Japanese Insectivora (Mammalia). I. Studies on variation and classification. Journal of the Faculty of Agriculture Hokkaido University, 55:191-265.
Abe, H., Shiraishi, S., and Arai, S. 1991. A new mole from Uotsuri-jima, Ryukyu Islands. Journal of Mammalogical Society of Japan, 15:47-60.
Bachman, J. and Charleston, D.D. 1843-1844. Observation on the genus Scalops (shrew moles) with descriptions of the species found in North America. Boston Journal of Natural History, 4:26-35.
Baird, S.F. 1858. Mammals: general report upon the zoology of several Pacific railroad Routes. Reports of explorations to ascertain the most practicable and economical route for railroad from the Mississippi River to the Pacific Ocean. Washington D.C., Senate Executive Document 78, 8(part 1):1-757.
Barnosky, A.D. 1981. A skeleton of Mesoscalops (Mammalia, Insectivora) from the Miocene Deep River Dormation, Montana, and a review of the proscalopid moles: evolutionary, functional, and stratigraphic relationships. Journal of Vertebrate Paleontology, 1:285-339.
Crochet, J.-Y. 1995. Le Garouillas et les sites contemporains (Oligocène, MP25) des phosphorites du Quercy (Lot, Tarn-Garonne, France) et leurs faunes de vertébrés. Palaeontographica Abteilungen A, 236:39-75.
Dahlmann, T. 2001. Die Kleinsäuger der unter-pliozänen Fundstelle Wölfersheim in der Wetterau (Mammalia: Lipotyphla, Chiroptera, Rodentia). Courier Forschungsinstitut Senckenberg , 227:1-129.
Dalquest, W.W. and Burgner, R.L. 1941. The shrew-mole of western Washington. The Murrelet, 22:12-14.
de Bruijn, H. and Rümke, C.G. 1974. On a peculiar mammalian association from the Miocene of Oschiri (Sardinia). I and II. Proceedings of the Koninklijke Nederlandse Akademie van Wetenschappen, Section B, 77:46-79.
Dobson, G.E. 1883. A Monograph of the Insectivora, Systematic and Anatomical , Part II. Including the Families Potamogalidae, Chrysochloridae, and Talpidae. John Van Voorst, London.
Engesser, B. 1972. Die obermiozäne Säugetierfauna von Anwil (Baselland). Tätigkeitsberichte der Naturforschchenden Gesellschaft Baselland, 28:35-364.
Engesser, B. 1980. Insectivora und Chiroptera (Mammalia) aus dem Neogen der Türkei. Schweizerische Paläontologische Abhandlungen, 102:47-149.
Engesser, B. 2005. Switzerland, p. 285-295. In Van den Hoek Ostende, L.W., Doukas, C.S., and Reumer, J.W.F. (eds.), The Fossil Record of the Eurasian Neogene Insectivores (Erinaceomorpha, Soricomorpha, Mammalia), part I.Scripta Geologica Special Issue5. National Museum of Natural History, Leiden.
Fejfar, O. and Sabol, M. 2005. Czech Republic and Slovak Republic, p. 51-60. In Van den Hoek Ostende, L.W., Doukas, C.S., and Reumer, J.W.F. (eds.), The Fossil Record of the Eurasian Neogene Insectivores (Erinaceomorpha, Soricomorpha, Mammalia), part I.Scripta Geologica Special Issue5 . National Museum of Natural History, Leiden.
Filhol, H. 1890. Description d’un nouveau genre d’insectivore provenant des dépôts de phosphate de chaux du Quercy. Bulletin de la Société philomatique de Paris, serie II, 8:176-177.
Fischer von Waldheim, G. 1813-1814. Zoognosia Tabulis Synopticis llustrata. Nicolai Sergeidis Vsevolozsky, Moscow.
Freudenberg, W. 1914. Die Säugetiere des älteren Quartärs von Mitteleuropa mit besonderer Berücksichtigung der Fauna von Hundsheim und Deutschaltenburg in Niederösterreich etc. Geologische und palaeontologische Abhandlungen (Neue Folge), 12:1-219.
Gaillard, C. 1899. Mammifères miocènes nouveaux ou peu connus de La Grive-St.-Alban. Archives du Muséum d’Histoire Naturelle de Lyon, 7:1-79.
Geoffroy Saint-Hilaire, É. 1803. Catalogue des mammifères du Muséum National d’Histoire Naturelle. Muséum National d’Histoire Naturelle, Paris.
Geoffroy Saint-Hilaire, É. 1811. Mémoire Sur les éspèces des Genres Musaraigne et Mygale. Annales du Muséum National d’Histoire Naturelle, 17:169-194.
Gill, T. 1875. Synopsis of insectivorous mammals. Bulletin of the United States Geological and Geographical Survey of the Territories (U.S.), 1(Bulletin 2):91-120.
Goszczyński, J. 1976. Estimation of daily food ratio of Tyto alba Scopoli under natural conditions. Polish Ecological Studies, 2:95-102.
Green, M. 1956. The lower Pliocene Ogallala-Wolf Creek vertebrate fauna, South Dakota. Journal of Paleontology, 30:146-159.
Gregory, W.K. 1910. The orders of mammals. Bulletin of the American Museum of Natural History , 37:1-524.
Gueldenstaedt, D. 1777. Beschreibung des Dsmans. Beschäftigungen der Berlinischen Gesellschaft Naturforschender Freunde 3:107-137.
Guerin, C. and Mein, P. 1971. Les principaux gisements de mammifères Miocènes et Pliocènes du domaine rhodanien. Documents du Laboratoire de Géologie de la Faculté des Sciences de Lyon 1:131-170.
Günther, A. 1880. Notes on some Japanese Mammalia. Proceedings of the Zoological Society of London, 48:440-443.
Gureev, A.A. 1979. Nasekomoyadnye (Mammalia, Insectivora - Erinaceidae, Talpidae, Soricidae). Fauna SSSR, Mlekopitayushchie, 4:1-501 [in Russian].
Hall, E.R. 1930. A new genus of bat from the later Tertiary of Nevada: University of California Publications, Bulletin of the Department of Geological Sciences, 19:319-320.
Hutchison, J.H. 1968. Fossil Talpidae (Insectivora, Mammalia) from the Late Tertiary of Oregon. Bulletin of the Museum of Natural History University of Oregon , 11:1-117.
Hutchison, J.H. 1974. Notes on type specimens of European Miocene Talpidae and a tentative classification of Old World Tertiary Talpidae (Insectivora: Mammalia). Geobios, 7:211-256.
Hutchison, J.H. 1984. Cf. Condylura (Mammalia, Talpidae) from the Late Tertiary of Oregon. Journal ofVertebrate Paleontology, 4:600-601.
Hutterer, R. 2005. Order Soricomorpha, p. 220-311. In Wilson, D.E. and Reeder, D.A.M. (eds.), Mammal Species of the World. The Johns Hopkins University Press, Baltimore.
Illiger, C.D. 1811. Prodromus Systematis Mammalium et Avium Additis Erminis Zoographicis Uttriusque Classis. Salfeld, Berlin.
Kaup J.J. 1829. Skizzierte Entwicklungsgeschichte und Natürliches System der Europäischen Thierwelt. Commission bei Carl Wilhelm Leske, Darmstadt und Leipzig.
Kormos, T. 1913. Trois nouvelles espèces fossiles des desmans en Hongrie. Annales Musei Nationales Hungarici, 11:135-146.
Lavocat, R. 1951. Révision de la Faune de Mammifères Oligocènes d’Auvergne et du Velay. Sciences et Avenir, Paris.
Linnaeus, C. 1758. Systema naturae pre regna tria naturae, secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis. Editio decima, reformata. Laurentius Salvus,Stockholm.
Love, V.P.W. 1980. Variation in the digestion of prey in the Tawny owl ( Srix aluco L.). Journal of Zoology, 192:283-293.
Matthew, W.D. 1901. Fossil mammals of the Tertiary of northeastern Colorado. Memoirs of the American Museum of Natural History, 1:353-447.
Miller, G.S. 1940. Notes of some moles from southeastern Asia. Journal of Mammalogy, 21:442- 444.
Milne-Edwards, A. 1867. Observations sur quelques mammifères du nord de la Chine. Annales des sciences naturelles, Zoologie et biologie animale, 7:375-377.
Milne-Edwards, A. 1871. In David, A. (ed.), Catalogue des oiseaux de Chine. Nouvelles Archives du Muséum d’Histoire Naturelle de Paris, 7:92.
Milne-Edwards, A. 1872. In David, A. (ed.), Rapport adressé a M.M. Les professeurs-administrateurs du Muséums d’Histoire Naturelle. Nouvelles Archives du Muséums d’histoire naturelle de Paris , 7:92.
Novacek, M.J. 1986. The skull of leptictid insectivorans and the higher-level classification of eutherian mammals. Bulletin of the American Museum of Natural History, 183:1-112.
Petényi, S.J. 1864. Hátrahagyott Munkái. F. Eggenberger, Budapest.
Pomel, N.A. 1848. Études sur les carnassiers insectivores. I. Insectivores fossiles. II. Classification des insectivores. Archives des sciences physiques et naturelles , 9:195-165, 244-251.
Popov, V.V. 2004. Late Pliocene Erinaceidae and Talpidae (Mammalia: Insectivora) from Varshets (North Bulgaria). Acta zoologica cracoviensia, 47:61-80.
Rădulescu, C., Samson, P. and Ştiucă, E. 1989. Pliocene (Lower Romanian) micromammals in the Dacic Basin. Miscellanea Speologica Romanica, 1:313-326.
Reed, K.M. 1961. The Proscalopinae, a new subfamily of talpid insectivores. Bulletin of the Museum of Comparative Zoology, 125:473-494.
Reumer, J.W.F. 2005. The Netherlands, p. 113-118. In Van den Hoek Ostende, L.W., Doukas, C.S., and Reumer, J.W.F. (eds.), The Fossil Record of the Eurasian Neogene Insectivores (Erinaceomorpha, Soricomorpha, Mammalia), part I.Scripta Geologica Special Issue5 . National Museum of Natural History, Leiden.
Rümke, C.A. 1976. Insectivora from Pikermi and Biodrak (Greece). Proceedings of the Koninklijke Nederlandse Akademie van Wetenshappen, Serie B, 79:256-270.
Rümke, C.A. 1985. A review of fossil and recent Desmaninae (Talpidae, Insectivora). Utrecht micropaleontplogical bulletins. Special publication, 4:1-264.
Rzebik-Kowalska, B. 1994. Pliocene and Quaternary Insectivora (Mammalia) of Poland. Acta zoologica cracoviensia, 37:77-136.
Rzebik-Kowalska, B. 2000. Insectivora (Mammalia) from the Early and early Middle Pleistocene of Betfia in Romania. II. Erinaceidae Bonaparte, 1838 and Talpidae Gray, 1825. Acta zoological cracoviensia, 43:55-77.
Rzebik-Kowalska, B. 2005. Poland, p. 119-134. In Van den Hoek Ostende, L.W., Doukas, C.S., and Reumer, J.W.F. (eds.), The Fossil Record of the Eurasian Neogene Insectivores (Erinaceomorpha, Soricomorpha, Mammalia), part I.Scripta Geologica Special Issue5. National Museum of Natural History, Leiden.
Rzebik-Kowalska, B. 2009. Biodiversity of Polish Fossil Insectivores (Erinaceomorpha, Soricomorpha, Insectivora, Mammalia) Compared to the European and Global Faunas . Institute of Systematics and Evolution of Animals Polish Academy of Sciences, Kraków.
Rzebik-Kowalska, B. 2013. Sorex bifidus n. sp. and the rich insectivore mammal fauna (Erinaceomorpha, Soricomorpha, Mammalia) from the Early Pleistocene of Żabia Cave in Poland. Palaeontologia Electronica, 16.2.12A:35pp, 1.96MB;
palaeo-electronica.org/content/2013/428-a-new-shrew-species
Rzebik-Kowalska, B. and Nesin, V.A. 2010. Erinaceomorpha and Soricomorpha (Insectivora, Mammalia) from the Late Miocene of Ukraine. Institute of Systematics and Evolution of Animals Polish Academy of Sciences, Kraków.
Schreuder, A. 1940. A revision of the fossil water-moles (Desmaninae). Archives Néerlandaises de Zoologie, 4:201-333.
Sigé, B., Crochet, J.-Y., and Insole, A. 1977. Les plus vieilles taupes. Geobios, 10(1):141-157.
Skoczeń, S. 1976. Condylurini Dobson, 1883 (Insectivora, Mammalia) in the Pliocene of Poland. Acta zooloéica cracoviensia, 21:291-313.
Skoczeń, S. 1980. Scaptonychini Van Valen, 1967, Urotrichini and Scalopini Dobson, 1883 (Insectivora, Mammalia) in the Pliocene and Pleistocene of Poland. Acta zoologica cracoviensia, 24:411-448.
Skoczeń, S. 1993. New records of Parascalops, Neurotrichus and Condylura (Talpinae, Insectivora) from the Pliocene of Poland. Acta Theriologica, 38:125-137.
Storch, G. 1978. Die turolische Wirbeltierfauna von Dorn-Dürkheim, Rheinhessen (SW-Deutschland). 2. Mammalia: Insectivora. Senckenbergiana Lethaea 58:421-449.
Storch, G. and Qiu, Z. 1983. The Neogene mammalian faunas of Ertemte and Harr Obo in Inner Mongolia (Nei Mongol), China. 2. Moles - Insectivora: Talpidae. Senckenbergiana lethaea, 64:89-127.
Storch, G. and Qiu, Z. 1991. Insectivores (Mammalia: Erinaceidae,Soricidae, Talpidae) from the Lufeng hominoid locality, Late Miocene of China. Geobios, 24:601-621.
Sulimski, A. 1959. Pliocene insectivores from Węże. Acta Palaeontologica Polonica, 4:119-173.
Temminck, C.J. 1841. Quatorzième monographie. Sur les genres taphien - queue en fourreau-queue-cache - et queue-bivalve,Monographies de Mammalogie ou Description de quelques Genres de Mammifères dont les Espèces ont été observées dans les Différents Musées de l’Europe. Van der Hock, Leiden.
Terzea, E. 1980. Deux Micromammifères du Pliocène de Roumanie. Travaux de l'Institut de Spéologie "Émile Racovitza", 19:191-201.
Thomas, O. 1911. New mammals from Central and Western Asia, mostly collected by Mr. Douglas Carruthers. Annals and Magazine of Natural History: Series 8, 8(48):758-762.
Thomas, O. 1912. On a collection of small mammals from the Tsin-ling Mountains, central China, presented by Mr. G. Fenwick Owen to the National Museum. Annals and Magazine of Natural History:Series 8, 10(58):395-403.
True, F.W. 1886. Description of a new genus and species of mole, Dymecodon pilirostris, from Japan. Proceedings of the United States National Museum , 86:97-98.
True, F.W. 1894. Notes on mammals of Baltistan and the Vale of Kashmir, presented to the National Museum by Dr W. L. Abbott. Proceedings of the United States National Museum, 17:1-16.
Van den Hoek Ostende, L.W. 1989. The Talpidae (Insectivora, Mammalia) of Eggingen-Mittelhart (Baden-Württenberg, F.R.G.) with special reference to the Paratalpa - Desmanodon lineage. Stuttgarter Beiträge zur Naturkunde, Serie B ( Geologie und Paläontologie), 152:1-29.
Van den Hoek Ostende, L.W. 2001. Insectivore faunas from the Lower Miocene of Anatolia - Part 5: Talpidae. Scripta Geologica, 122:1-45.
Van den Hoek Ostende, L.W. and Furió, M. 2005. Spain, p. 148-284. In Van den Hoek Ostende, L.W., Doukas, C.S., and Reumer, J.W.F. (eds.), The Fossil Record of the Eurasian Neogene Insectivores (Erinaceomorpha, Soricomorpha, Mammalia), part I.Scripta Geologica Special Issue5 . National Museum of Natural History, Leiden.
Van Valen, 1967. New Paleocene Insectivores and insectivore classification. Bulletin of American Museum of Natural History, 132:1-126.
Van Zyll de Jong, C.G. 1983. Handbook of Canadian mammals. Marsupials and Insectivores. National Museum of Canada, 1:1-210.
Webster, J.A. 1973. Seasonal variation in mammal contents of Barn owl castings. Bird study , 20:185-196.
Wilson, R.W. 1960. Early Miocene rodents and insectivores from northeastern Colorado. The University of Kansas Paleontological Contribution, 7:1-92.
Ziegler, R. 1990. Talpidae (Insectivora, Mammalia) aus dem Oberoligozän und Untermiozän Süddeutschlands. Stuttgarter Beiträge zur Naturkunde, Serie B, 176:1-81.
Ziegler, R. 2003. Moles (Talpidae) from the late Middle Miocene of South Germany. Acta Palaeontologica Polonica, 48:617-648.
Ziegler, R., Dahlmann, T., Reumer, J.W.F., and Storch, G. 2005. Germany, 61-90. In Van den Hoek Ostende, L.W., Doukas, C.S., and Reumer, J.W.F. (eds.), The Fossil Record of the Eurasian Neogene Insectivores (Erinaceomorpha, Soricomorpha, Mammalia), part I.Scripta Geologica Special Issue5 . National Museum of Natural History, Leiden.
Ziegler, R. and Daxner-Hock, G. 2005. Austria, p. 11-29. In Van den Hoek Ostende, L.W., Doukas, C.S., and Reumer, J.W.F. (eds.), The Fossil Record of the Eurasian Neogene Insectivores (Erinaceomorpha, Soricomorpha, Mammalia), part I.Scripta Geologica Special Issue5 . National Museum of Natural History, Leiden.
Zijlstra, J.S. 2010. Neurotrichus skoczeni, new name for Neurotrichus minor Skoczeń, 1993, preoccupied. Journal of Vertebrate Paleontology, 30:1903.
