

 A new approach to the systematics of Laeviprosopon (Brachyura: Homolidae), with remarks on molting process of the early brachyurans
A new approach to the systematics of Laeviprosopon (Brachyura: Homolidae), with remarks on molting process of the early brachyurans
Article number: 26.1.a10
https://doi.org/10.26879/1204
Copyright Paleontological Society, April 2023
Proceedings of the 8th Symposium on Fossil Decapod Crustaceans
Author biographies
Plain-language and multi-lingual abstracts
PDF version
Submission: 22 December 2021. Acceptance: 28 February 2023.
ABSTRACT
Our understanding of the molting process in the Jurassic brachyurans is very scarce because useful material is rare. New specimens from Poland and Germany, in addition to previously known specimens, let us further investigate the molting process. Specimens of Laeviprosopon laeve reveal characters not known before. On the dorsal carapace, molting lines are confirmed in three species of Laeviprosopon: the type species L. laeve, L. laculatum and L. fraasi. This brought questions about the systematic position of Laeviprosopon, which we here reassign to Homolidae within Homoloidea. Furthermore, two specimens ascribed previously to L. sublaeve are transferred to L. fraasi, and one specimen ascribed previously to Navarrahomola hispanica is transferred to Laeviprosopon hispanicum. Three new species of Laeviprosopon (L. joecollinsi n. sp. and L. lanceatum n. sp. from the Oxfordian of Poland, and L. ewakrzeminskae n. sp. from the Tithonian of Austria) are erected. Finally, we deliberate on Laeviprosopon as a possible phyletic link between homolodromioids and homoloids in the light of new data on the evolution of molting line.
Natalia Starzyk. Institute of Systematics and Evolution of Animals, Polish Academy of Sciences, ul. Sławkowska 17, 31-016 Kraków, Poland. Corresponding author. natalia_slaby@wp.pl
Barry W.M. Van Bakel. Department of Earth Sciences, Faculty of Geosciences, Utrecht University, Budapestlaan 4, 3584 CD Utrecht, the Netherlands and Oertijdmuseum, Bosscheweg 80, 5283 WB Boxtel, the Netherlands. barryvanbakel@gmail.com
Adiël A. Klompmaker. Department of Museum Research and Collections & Alabama Museum of Natural History, University of Alabama, Box 870340, Tuscaloosa, Alabama 35487, USA. adielklompmaker@gmail.com
Günter Schweigert. Staatliches Museum für Naturkunde, Rosenstein 1, 70191 Stuttgart, Germany. guenter.schweigert@smns-bw.de
René H.B. Fraaije. Oertijdmuseum, Bosscheweg 80, NL-5283 WB Boxtel, The Netherlands. info@oertijdmuseum.nl
Keywords: Brachyura; Homoloidea; Homolodromioidea; new species; molting line
Final citation: Starzyk, Natalia, Van Bakel, Barry W.M., Klompmaker, Adiël A., Schweigert, Günter, and Fraaije, René H.B. 2023. A new approach to the systematics of Laeviprosopon (Brachyura: Homolidae), with remarks on molting process of the early brachyurans. Palaeontologia Electronica, 26(1):a10.
https://doi.org/10.26879/1204
palaeo-electronica.org/content/2023/3788-systematics-of-laeviprosopon
Copyright: April 2023 Paleontological Society.
This is an open access article distributed under the terms of Attribution-NonCommercial-ShareAlike 4.0 International (CC BY-NC-SA 4.0), which permits users to copy and redistribute the material in any medium or format, provided it is not used for commercial purposes and the original author and source are credited, with indications if any changes are made.
creativecommons.org/licenses/by-nc-sa/4.0/
http://zoobank.org/C7630846-28CE-4F22-90FE-F970ACE300BA
INTRODUCTION
The systematic position of Laeviprosopon Glaessner, 1933, has changed several times due to the interpreted presence or absence of a molting line, often referred to as a linea homolica for crabs ascribed to Homoloidea. Glaessner (1933) initially placed this genus in the family Prosopidae von Meyer, 1860 (superfamily Homolodromioidea Alcock, 1900), but Patrulius (1966) transferred it to the superfamily Homoloidea due to the inferred presence of a linea homolica. His reassignment was followed by Collins and Wierzbowski (1985), Wright and Collins (1972) and Wehner (1988) (as also summarized in Klompmaker et al., 2013).
However, according to Schweitzer and Feldmann (2008), species of Laeviprosopon lack a true linea homolica and its presumed presence, noticed by Patrulius and others, might be a preservational artifact. Therefore, Laeviprosopon was placed in the Prosopidae again by them, followed by others subsequently (e.g., Franţescu, 2011; Klompmaker, 2013; Klompmaker et al., 2020; Starzyk, 2020).
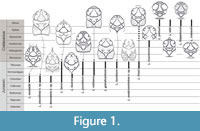 Six Late Jurassic species (Figure 1) from the Bachmayer collection housed in the Natural History Museum in Vienna were revised and described by Schweitzer and Feldmann (2008), including two new species: L. laeve (von Meyer, 1860), L. sublaeve (von Meyer, 1860), L. punctatum (von Meyer, 1860), L. fraasi (Moericke, 1889), L. laculatum Schweitzer and Feldmann, 2008, and L. grandicentrum Schweitzer and Feldmann, 2008. These authors also mentioned the Cretaceous species L. icaunensis (Van Straelen, 1936), which they treated as being specifically distinct from L. fraasi (opposite to Wehner, 1988). The arguments for that view were the geographic and age differences between L. icaunensis (Hauterivian of France - Van Straelen, 1936) and L. fraasi (Tithonian of Poland [Moericke, 1889] and Romania [Patrulius, 1966]). As Klompmaker (2013: table 2) noted, L. fraasi was also mentioned by Patrulius (1966) from the Urgonian (Barremian-Aptian) of Romania, but that specimen was not illustrated. Since there is a time gap of at least 15 Ma between the type specimen of L. fraasi and the specimen mentioned by Patrulius (1966), it is likely that the latter represents a different species.
Six Late Jurassic species (Figure 1) from the Bachmayer collection housed in the Natural History Museum in Vienna were revised and described by Schweitzer and Feldmann (2008), including two new species: L. laeve (von Meyer, 1860), L. sublaeve (von Meyer, 1860), L. punctatum (von Meyer, 1860), L. fraasi (Moericke, 1889), L. laculatum Schweitzer and Feldmann, 2008, and L. grandicentrum Schweitzer and Feldmann, 2008. These authors also mentioned the Cretaceous species L. icaunensis (Van Straelen, 1936), which they treated as being specifically distinct from L. fraasi (opposite to Wehner, 1988). The arguments for that view were the geographic and age differences between L. icaunensis (Hauterivian of France - Van Straelen, 1936) and L. fraasi (Tithonian of Poland [Moericke, 1889] and Romania [Patrulius, 1966]). As Klompmaker (2013: table 2) noted, L. fraasi was also mentioned by Patrulius (1966) from the Urgonian (Barremian-Aptian) of Romania, but that specimen was not illustrated. Since there is a time gap of at least 15 Ma between the type specimen of L. fraasi and the specimen mentioned by Patrulius (1966), it is likely that the latter represents a different species.
New specimens of Laeviprosopon punctatum were mentioned recently (Schweigert et al., 2016; Schweigert, 2019) from the late Kimmeridgian of Geislingen an der Steige (= Bad Überkingen-Oberböhringen) and Saal an der Donau, southern Germany, and one other specimen was illustrated by Klompmaker et al. (2020) from the Tithonian of Ernstbrunn in Austria.
In 2011, Franţescu described another species, L. lazarae Franţescu, 2011, from the Late Jurassic (late Oxfordian to early Kimmeridgian) of Romania (Figure 1).
Subsequently, Klompmaker (2013) described four new species: L. planum Klompmaker, 2013; L. hispanicum Klompmaker, 2013; L. edoi Klompmaker, 2013; and L. crassum Klompmaker, 2013. They originate from the mid-Cretaceous (late Albian) of Spain and are the stratigraphically youngest species of the genus (Figure 1). In 2020, Starzyk described the oldest species of Laeviprosopon: L. musialiki Starzyk, 2020, from the Late Jurassic (early Oxfordian) of Poland.
Schweitzer and Feldmann (2008) modified the diagnosis of Laeviprosopon created by Glaessner (1933), based on seven Jurassic and Cretaceous species: L. laeve, L. fraasi, L. grandicentrum, L. icaunensis, L. laculatum, L. punctatum and L. sublaeve. As four Cretaceous species described by Klompmaker (2013) differ from the diagnosis of Laeviprosopon by Schweitzer and Feldmann (2008), he expanded it to include also species with a wider carapace (Klompmaker, 2013). He also discussed the issue of the presence of a linea homolica in the Cretaceous representatives of the genus. All four species described by him have a symmetrical breakage on both sides of the carapace, which corresponds to the expected position of a linea homolica (Klompmaker, 2013). This observation leads to the question whether a linea homolica has evolved only once, within Homoloidea, or that similar structures evolved independently in different brachyuran groups.
The goals of this paper are to study the systematics and diversity of Jurassic and Cretaceous Prosopidae and Homolidae with a focus on Laeviprosopon, and also to study and hypothesize about modes of molting in basal crabs in the Mesozoic.
LOCALITIES AND STRATIGRAPHY
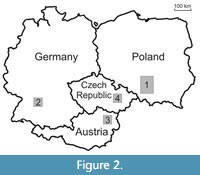 The new material described herein originates from five Oxfordian localities in the southern Polish Uplands, north-west of Kraków (Bzów - 50°28’04”N 19°31’12”E, Ogrodzieniec - 50°27’54”N 19°31’23”E, Niegowonice - 50°24’10”N 19°24’35”E, Szklary - 50°16‘16”N 19°70’08”E, Żarki - 50°37’42”N 19°22’03”E) and a Kimmeridgian locality in Bad Überkingen-Oberböhringen, Germany (48°37’15”N 9°47’58”E) (Figure 2). The first three localities and their stratigraphy have been reported in Fraaije et al. (2012a, b), Krzemińska et al. (2015), Starzyk et al. (2012) and Starzyk (2015a, b).
The new material described herein originates from five Oxfordian localities in the southern Polish Uplands, north-west of Kraków (Bzów - 50°28’04”N 19°31’12”E, Ogrodzieniec - 50°27’54”N 19°31’23”E, Niegowonice - 50°24’10”N 19°24’35”E, Szklary - 50°16‘16”N 19°70’08”E, Żarki - 50°37’42”N 19°22’03”E) and a Kimmeridgian locality in Bad Überkingen-Oberböhringen, Germany (48°37’15”N 9°47’58”E) (Figure 2). The first three localities and their stratigraphy have been reported in Fraaije et al. (2012a, b), Krzemińska et al. (2015), Starzyk et al. (2012) and Starzyk (2015a, b).
The locality in Szklary is dated as middle Oxfordian (Müller et al., 2000; Garassino and Krobicki, 2002). The age of the sediments in Żarki is dated as early Oxfordian costicardia Subzone of the cordatum Zone (Głowniak, 2012). The age of the natural outcrop in Oberböhringen, is dated as early late Kimmeridgian acanthicum Zone based on the ammonite fauna (Schweigert et al., 2016). The age of the Štramberk Limestone at Kotouč Quarry in the Czech Republic is dated as Tithonian to early Berriasian (Vašíček et al., 2018).
The crab-bearing Oxfordian localities of Poland and the Kimmeridgian locality in Germany are situated in deep-water environments characterized by exhaustive siliceous sponge-microbial buildups, which occurred in a belt along the northern Tethys shelf and span from the subsurface of present-day Atlantic to extra-Carpathian Romania (e.g., Leinfelder et al., 1994, 1996; Leinfelder and Keupp, 1995; Matyja and Wierzbowski, 1996). In the Oxfordian Polish localities, namely that of Szklary near Kraków, the crabs occur autochthonously within the reef mounds (Fraaije et al., 2022), whereas in the Germany locality of Kimmeridgian age the rock matrix is a thick-bedded biodetritic spongiolitic limestone pointing to episodical mudflows from adjacent reefs (G.S., pers. observation).
The Tithonian to Berriasian Štramberk Limestone in the Czech Republic has formed on an isolated carbonate platform of the Tethys similar to the coeval Ernstbrunn Limestone of Austria. Both are characterized by scleractinian coral patch reefs and predominantly associated biodetritic rudstones and grainstones. Microfacies as well as dasycladalean algae and typical microencrusters at both occurrences are indicative of shallow-water environments within the photic zone (e.g., Schneider et al., 2013, Hoffmann et al., 2017).
MATERIAL AND METHODS
The specimens studied herein come from collections of the following museums: Geological Museum of Warsaw University, Poland (IGPUW); Natural History Museum of Vienna, Austria (NHMW); Bayerische Staatssammlung für Paläontologie und Geologie, Munich, Germany (BSP, AS); Staatliches Museum für Naturkunde, Stuttgart, Germany (SMNS); Oertijdmuseum, Boxtel, The Netherlands (MAB); Museum of the Institute of Systematics and Evolution of Animals, Polish Academy of Sciences in Kraków, Poland (ISEA PAS, museal prefix - I-F/MP); Florida Museum of Natural History (Division of Invertebrate Paleontology), University of Florida, Gainesville, Florida, USA (UF).
 In total, there are 19 specimens described herein in the collection of ISEA PAS. Abundant material of fossil crabs from this collection has been described previously (Brachyura: Starzyk et al., 2011, 2012; Starzyk, 2013, 2015a, 2015b, 2016, 2020; Klompmaker et al., 2020; and Anomura: Fraaije et al., 2012a, 2012b, 2014; Krzemińska et al., 2015). Specimens from the collection of the Museum of ISEA PAS were collected by a family of amateur collectors: Iwona, Robert and Karolina Borek; two paratypes of L. joecollinsi n. sp. were collected by Adiël Klompmaker; 12 specimens of L. laeve described herein were collected by Horst Kuschel.
In total, there are 19 specimens described herein in the collection of ISEA PAS. Abundant material of fossil crabs from this collection has been described previously (Brachyura: Starzyk et al., 2011, 2012; Starzyk, 2013, 2015a, 2015b, 2016, 2020; Klompmaker et al., 2020; and Anomura: Fraaije et al., 2012a, 2012b, 2014; Krzemińska et al., 2015). Specimens from the collection of the Museum of ISEA PAS were collected by a family of amateur collectors: Iwona, Robert and Karolina Borek; two paratypes of L. joecollinsi n. sp. were collected by Adiël Klompmaker; 12 specimens of L. laeve described herein were collected by Horst Kuschel.
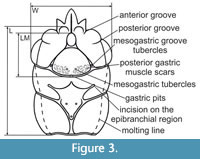
The length of the mesogastric region, width of the carapace at the hepatic region and the length of the carapace without the rostrum were measured (Figure 3). The total length was not measured because the rostrum in most of the carapaces is poorly preserved or broken.
SYSTEMATIC PALAEONTOLOGY
Order DECAPODA Latreille, 1802
Infraorder BRACHYURA Linnaeus, 1758
Superfamily HOMOLOIDEA de Haan, 1839
Family HOMOLIDAE de Haan, 1839
Genus LAEVIPROSOPON Glaessner, 1933
Type species. Prosopon laeve von Meyer, 1860, by original designation.
Other species included. Laeviprosopon crassum Klompmaker, 2013; L. edoi Klompmaker, 2013; L. ewakrzeminskae n. sp.; L. fraasi (Moericke, 1889) as Prosopon; L. grandicentrum Schweitzer and Feldmann, 2008; L. hispanicum Klompmaker, 2013; L. icaunensis (Van Straelen, 1936) as Prosopon; L. joecollinsi n. sp.; L. laculatum Schweitzer and Feldmann, 2008; L. lazarae Franţescu, 2011; L. lanceatum n. sp.; L. musialiki Starzyk, 2020; L. planum Klompmaker, 2013; L. punctatum (von Meyer, 1860) as Prosopon; L. sublaeve (von Meyer, 1860) as Prosopon.
Diagnosis. As in Klompmaker (2013). “Carapace rectangular, typically longer than wide, often narrowing slightly anteriorly; regions well-defined by deep grooves; rostrum variable, projected well beyond orbits; orbits shallow, rimmed, directed forward, located at base of rostrum; augenrest poorly formed to nearly absent; subhepatic region inflated, sometimes greatly; epigastric region separated from protogastric and hepatic regions by transverse groove; hepatic and protogastric regions confluent or partly divided by groove on lateralmost part of dorsal carapace; cervical groove deep, strongly concave forward to sinuous; branchiocardiac groove usually shallower than cervical groove; post-cervical groove usually short; posterolateral portions of carapace often broken as if less calcified than other portions of carapace.”
Remarks. As was mentioned in the introduction, the systematic position of Laeviprosopon has changed several times. Authors that did not confirm the presence of linea homolica placed it in the family Prosopidae (superfamily: Homolodromioidea), while authors that confirmed the presence of this structure in L. laeve placed the genus in the family Homolidae (superfamily: Homoloidea). In this paper, we show unequivocal evidence of the presence of linea homolica in three Late Jurassic representatives of Laeviprosopon. Eleven new specimens of L. laeve represent molds with clearly visible molting lines similar to a linea homolica, which is also present on the carapaces of L. fraasi and L. laculatum. Specimens of four Cretaceous species of Laeviprosopon (L. hispanicum, L. planum, L. edoi and L. crassum) are also molds presenting the same type of molting line. These observations are sufficient to reassign Laeviprosopon to the family Homolidae as it includes the oldest known brachyurans with a linea homolica (see chapter Discussion for more details).
Laeviprosopon laeve (von Meyer, 1860)
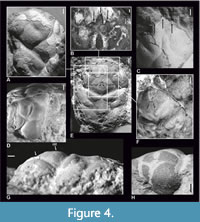
Figure 1, Figure 3, Figure 4, Figure 5
1857 Prosopon laeve von Meyer, p. 556.
1860 Prosopon laeve von Meyer, 1857. - von Meyer, p. 209, pl. 23, figure 21.
1895 Prosopon laeve von Meyer, 1857. - Remeš, p. 202, pl. 12, figure 14.
1966 Laeviprosopon laeve (von Meyer, 1857). - Patrulius, p. 506, figure 2a, 4, pl. 30, figure 7.
1985 Laeviprosopon laeve (von Meyer, 1857). - Collins in Collins and Wierzbowski, p. 86, pl. 4, figure 3-3.
2008 Laeviprosopon laeve (von Meyer, 1857). - Schweitzer and Feldmann, p. 277, figure 1.1-1.5.
2018 Laeviprosopon sublaeve (von Meyer, 1857). - Schweigert and Kuschel, p. 19, figure 8.
Diagnosis. (After Schweitzer and Feldmann, 2008, emended). Species reaching relatively large size (length of mesogastric region of largest specimen is 16.1 mm). Carapace longer than wide, rectangular, strongly vaulted transversally and moderately vaulted longitudinally; cuticle smooth or covered with small granules. Rostrum trifid. Orbit flattened, short, distinctly bordered, with a suborbital spine. Mesogastric region with two gastric spines and posterior gastric muscle scars posterior to these spines. Mesogastric groove tubercles present. Anterior part of carapace and epibranchial region convex, branchial region flatter. Molting lines similar to linea homolica present.
Type material. Laeviprosopon laeve was described by von Meyer in 1860 based on two specimens which are both lost (Schweitzer and Feldmann, 2008). They have been collected from the Kimmeridgian locality Oerlinger Tal in Germany. Herein we describe new material from the upper Kimmeridgian locality Bad Überkingen-Oberböhringen, which is stratigraphically close to the Oerlinger Tal locality. Therefore, we assign a neotype: specimen SMNS 70527/1.
Material Examined
Austria. Ernstbrunn (Tithonian of Waschberg Zone): NHMW 1990/0041/3309; NHMW 1990/0041/3834; NHMW 1990/0041/4915; NHMW 2007z0149/0020; NHMW 2007z0149/0021; NHMW 2007z0149/0022.
Poland. Rudniki (lower Kimmeridgian, planula Zone): IGPUW/C/1/128, Niwiska Dolne (lower Kimmeridgian, planula Zone): IGPUW/C/1/129.
Germany. Bad Überkingen-Oberböhringen (upper Kimmeridgian): SMNS 70527/1 - neotype, SMNS 70527/2, SMNS 70527/3, SMNS 70527/4, SMNS 70527/5, SMNS 70527/6, SMNS 70527/7, SMNS 70527/8, SMNS 70527/9, SMNS 70527/10, SMNS 70527/11.
Poland. Bzów (middle Oxfordian): I-F/MP/3663/1533/08; I-F/MP/3914/1533/08; I-F/MP/4627/1534/08.
Niegowonice (middle - upper Oxfordian): I-F/MP/915/1507/08; I-F/MP/3245/1532/08; I-F/MP/3295/1532/08; I-F/MP/4810/1534/08; I-F/MP/5467/1543/09; I-F/MP/6275/1599/12.
Ogrodzieniec (lower - middle Oxfordian): I-F/MP/172/1489/08.
Dimensions. The state of preservation of the material from the ISEA collection is not good enough to take measurements of all specimens. The length of the mesogastric region of the smallest specimen is 3.6 mm and that of the largest one 8.8 mm. The length of the mesogastric region of specimen SMNS 70527/1 is 5.4 mm, 4.0 mm for SMNS 70527/4, and 4.5 mm for SMNS 70527/2. Total length without rostrum is 12.0 mm in the specimen SMNS 70527/1, 9.1 in the specimen SMNS 70527/4 and 10.3 mm in the specimen SMNS 70627/2. The width at the hepatic region is 9.2 mm in the specimen SMNS 70527/1.
 Emended description. As a large number of new specimens show characters not known before, a redescription is needed.
Emended description. As a large number of new specimens show characters not known before, a redescription is needed.
The carapace is longer than wide, strongly vaulted transversally and moderately vaulted longitudinally, widest in the hepatic region. The rostrum is well preserved only for 2007z0149/0022, showing three spines, of which the middle one is the longest and directed downwards. There is only a forked base of the rostrum visible in SMNS 70527/8 (Figure 4C marked with black arrows). There are distinct spines situated laterally from the rostrum (Figure 4D, H).
The anterior groove is visible in the intersection of the rostrum and the upper orbital margin. The epigastric regions are rounded. The mesogastric region is distinctly bordered by mesogastric grooves; the length of its anterior (narrow) part is about 0.59 of the posterior (wide) part; the separation between these parts is marked by an incision. The posterior gastric muscle scars are present on the posterior mesogastric region, and in front of them there are two tubercles (Figure 4E-G). Mesogastric groove tubercles are present (Figure 4E).
The gastric pits are rounded. The urogastric region is vaulted, with an incision in the center dividing it into two parts (Figure 4E). A tubercle on the urogastric region is very small. Lateral parts of the postcervical groove are connected with the branchiocardiac groove. These grooves bound narrow surfaces anterior to the cardiac region (Figure 4E; Figure 5A, D-E).
The cardiac region is distinctly delimited by the branchiocardiac groove. There are two tubercles in the anterior part of this region and one in the posterior part (Figure 5A-B, D-E).
The anterior part of the carapace and the epibranchial region are convex, and the branchial region is more flattened. The highest point of the carapace lies in the posterior part of the mesogastric region (Figure 5G-H). The grooves are deep and wide, and the cervical groove is the widest. The branchiocardiac groove reaches the posterior margin of the carapace. Few of the specimens from ISEA (I-F/MP/915/1507/08, I-F/MP/54671543/09) have almost smooth carapaces with only few tubercles on the steinkern; the cuticle is not preserved. Others from ISEA (I-F/MP/3663/1533/08, I-F/MP/3914/1533/08 and I-F/MP/6275/1599/12) have carapaces covered with tubercles (Figure 4A). Specimens from Oberböhringen and Ernstbrunn are also covered with tubercles. The cuticle of these specimens is relatively thick, and its surface is covered with tiny pits, each of which corresponds to a tubercle on the steinkern. Pits on the cuticle and corresponding tubercles on the steinkern are also visible on specimen I-F/MP/3663/1533/08 (Figure 4A). The posterior margin of the carapace is preserved only partially or not at all.
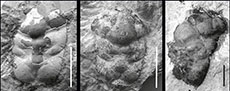
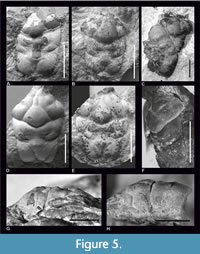 Most specimens from the Polish localities are damaged in the posterior part, but specimens I-F/MP/39141533/08, I-F/MP/6275/1599/12 (Figure 5C), IGPUW/C/1/128 and IGPUW/C/1/129 are showing a loss of lateral parts of the branchial regions. Most specimens from the German locality Oberböhringen are also molts with a molting line similar to a linea homolica (Figure 5A-B, D-G). Only one specimen shows an entire, unbroken carapace and the molting line is not visible on it (Figure 5H). Molting lines are laying on the dorsal surface of the branchial and epibranchial regions and crossing on the ventrolateral side of the hepatic region (Figure 5).
Most specimens from the Polish localities are damaged in the posterior part, but specimens I-F/MP/39141533/08, I-F/MP/6275/1599/12 (Figure 5C), IGPUW/C/1/128 and IGPUW/C/1/129 are showing a loss of lateral parts of the branchial regions. Most specimens from the German locality Oberböhringen are also molts with a molting line similar to a linea homolica (Figure 5A-B, D-G). Only one specimen shows an entire, unbroken carapace and the molting line is not visible on it (Figure 5H). Molting lines are laying on the dorsal surface of the branchial and epibranchial regions and crossing on the ventrolateral side of the hepatic region (Figure 5).
Discussion. The new specimens of the type species from the German locality Oberböhringen are crucial for reassessing the systematic placement of Laeviprosopon. These specimens are ascribed to the type species due to their characters which correspond to the original illustration of L. laeve. The overall shape and convexity of the carapace and each region and groove, the presence of gastric muscle scars and the mesogastric groove tubercles are consistent with L. laeve.
There is intraspecific variation visible in the ornamentation of the carapace of specimens from different localities, apparently not related to preservation. There are specimens with and without tubercles on the steinkern in the collection of ISEA PAS. Specimens from Oberböhringen and Ernstbrunn are also covered with tubercles on the steinkern. These tubercles correspond with pits on the cuticle where preserved.
These pits in the cuticle and corresponding tubercles on the steinkern are also present on specimen I-F/MP/3663/1533/08. Because all other characters correspond well with Laeviprosopon laeve, the presence of tubercles is considered to represent intraspecific variation.
Laeviprosopon sublaeve (von Meyer, 1860)
Figure 1
1857 Prosopon sublaeve von Meyer, p. 556.
1860 Prosopon sublaeve von Meyer. - von Meyer, p. 209, pl. 23, figure 23.
1925 Prosopon sublaeve von Meyer. - Beurlen, p. 484, 485.
1929 Prosopon laeve var. sublaevis von Meyer. - Glaessner, p. 344.
1933 Laeviprosopon laeve var. sublaevis (von Meyer). - Glaessner, p. 180.
1988 Laeviprosopon sublaeve (von Meyer). - Wehner, p. 119, text-figure 32, pl. 8, figure 3.
Diagnosis. (After Schweitzer and Feldmann 2008, emended). Carapace longer than wide, rectangular; front broadly convex; orbits well-developed for genus; regions ornamented with tubercles and larger nodes, especially posteriorly, five tubercles on each side of mesogastric region.
Discussion and type designation. Laeviprosopon sublaeve was described by von Meyer (1860), who illustrated a drawing of a specimen from Oerlinger Thal near Ulm (late Kimmeridgian). He drew transverse grooves dividing areas laying on both sides of the mesogastric region on epigastric, protogastric, hepatic and subhepatic regions and five tubercles laying on each side of the mesogastric region. Both on the drawing and in the description (translated in Schweitzer and Feldmann, 2008), the transverse grooves are present. Unfortunately, all of the six syntypes of von Meyer are lost, so that there is no possibility left to select a lectotype.
Wehner (1988) illustrated a specimen (GPIT-PV-51741) of Laeviprosopon sublaeve from Oerlinger Thal housed in the Museum of the University in Tübingen. Wehner cited the label with the wrong information: the mentioned specimen is not one of the specimens illustrated by Quenstedt (1857, pl. 95) and probably not illustrated in later books of Quenstedt (see Quenstedt 1885, pl. 31, fig. 21). As this information was erroneous, we cannot be sure if the locality described as Oerlinger Thal is right. However, as all six syntypes are lost, we here designate the specimen described by Wehner as a neotype. Its current number in the paleontological collection of Tübingen University is GPIT-PV-51741.
The specimen has all the diagnostic characters described and drawn by von Meyer. On the photo published by Wehner, two tubercles seem to be visible on each side of the mesogastric region, while the drawing made by Wehner (who studied the actual specimen) shows only one tubercle on each side (Wehner, 1988, figure 32B, C; pl. 8, figure 3).
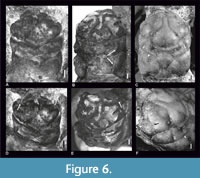
The specimens assigned to L. sublaeve by Schweitzer and Feldmann (2008, figure 1.10 and 1.11, herein Figure 6) differ. Both specimens have transverse grooves between the cervical and branchial groove, which are absent in the specimens described by von Meyer (1860) and Wehner (1988). From the neotype specimen we consider it is barely possible that both von Meyer and Wehner could have overlooked the presence of these grooves on their drawings and descriptions. As specimens NHMW 1990/0041/4051 and NHMW 1990/0041/3295 described by Schweitzer and Feldmann (2008) are most similar to Laeviprosopon fraasi, we consider them to represent this species (see text below and Figure 6).
Laeviprosopon fraasi (Moericke, 1889)
Figure 1, Figure 6
1889 Prosopon fraasi Moericke, p. 63, pl. 6, figure 17.
 1897 Prosopon fraasi Moericke, p. 63, pl. 6, figure 17.
1897 Prosopon fraasi Moericke, p. 63, pl. 6, figure 17.
1895 Prosopon fraasi Moericke. - Remeš, p. 203, pl. 3, figure 15.
1905 Prosopon fraasi Moericke. - Remeš, p. 35.
1925 Avihomola fraasi (Moericke). - Van Straelen, p. 347.
1929 Nodoprosopon fraasi (Moericke). - Glaessner, p. 271.
1933 Laeviprosopon fraasi (Moericke). - Glaessner, p. 180.
1966 Laeviprosopon fraasi (Moericke). - Patrulius, p. 507.
1988 Laeviprosopon fraasi (Moericke). - Wehner, p. 119, text-figure 32, pl. 8, figure 4a, 4b.
2008 Laeviprosopon fraasi (Moericke). - Schweitzer and Feldmann, p. 281, pl. 1.6.
2008 Laeviprosopon sublaeve (von Meyer). - Schweitzer and Feldmann, p. 278-280, pl. 1.10, 1.11.
Diagnosis. (After Schweitzer and Feldmann, 2008, emended.) Carapace longer than wide, regions well defined, smooth on anterior part, with tubercles on posterior part. Grooves deep; carapace flattened longitudinally and moderately vaulted transversely. Orbits flattened, short, distinctly bordered, narrowing posteriorly. Mesogastric region with posterior gastric muscle scars. Mesogastric groove tubercles present. Molting lines similar to linea homolica present.
Material Examined
Czech Republic. Štramberk (Tithonian): holotype BSP AS III 309.
Austria. Ernstbrunn (Tithonian of Waschberg Zone): NHMW 1990/0041/3295; NHMW 1990/0041/4051.
Dimensions. Dimensions of specimens are given in Schweitzer and Feldmann (2008, p. 279, table 1).
Discussion. Schweitzer and Feldmann (2008) assigned two specimens from Ernstbrunn to L. sublaeve, although their characters distinctly differ from the holotype. Specimens NHMW 1990/0041/4051 and NHMW 1990/0041/3295 (Figure 6; Schweitzer and Feldmann, 2008, figure 1.10, 1.11) are very similar to L. fraasi because their groove pattern on both sides of the mesogastric region and transverse grooves between the cervical and branchiocardiac grooves are alike. They have a short rostrum, shortened epigastric regions, and the ornamentation on the posterior part of the carapace is similar.
There is a difference between the holotype of L. fraasi (Figure 6C, F) and one of the specimens from Ernstbrunn; specimen NHMW 1990/0041/3295 (Figure 6A, D) has a smaller posterior part of the mesogastric region, opposite to specimen NHMW 1990/0041/4051 (Figure 6B, E), but given all other matching characters we consider both specimens to represent L. fraasi.
There is a molting line similar to a linea homolica visible on specimen NHMW 1990/0041/4051 (Figure 6B, E). This specimen presents a complete carapace and the molting line is shaped similar as in L. leave; on the dorsal part of the branchial and epibranchial regions, it is not reaching the anterior margin, but it is directed to the ventral surface of the carapace on the level of the cervical groove. Patrulius (1966) also mentioned the presence of a linea homolica on the specimens of L. laeve and L. fraasi. This view was supported by Wright and Collins (1972) and Collins and Wierzbowski (1985).
Laeviprosopon ewakrzeminskae n. sp.
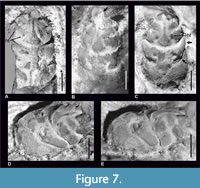
Figure 1, Figure 7
zoobank.org/004C6A33-3FC4-45B7-8C5A-D3002EC6B540
 Diagnosis. Carapace very elongate, narrowing anteriorly, strongly vaulted transversally and moderately vaulted longitudinally. Orbits not differentiated. Very long and narrow mesogastric region. Two tubercles on each side of mesogastric region present. Branchial region ornamented with very small tubercles.
Diagnosis. Carapace very elongate, narrowing anteriorly, strongly vaulted transversally and moderately vaulted longitudinally. Orbits not differentiated. Very long and narrow mesogastric region. Two tubercles on each side of mesogastric region present. Branchial region ornamented with very small tubercles.
Etymology. The species name is dedicated to Ewa Krzemińska (ISEA PAS), with a deep gratitude for being an advisor and a mentor of Natalia Starzyk. Ewa is a renowned specialist of recent and fossil Diptera, also working on fossil Brachyura and Anomura.
Comparisons. Specimens of Laeviprosopon ewakrzeminskae n. sp. are narrowing anteriorly as in L. joecollinsi, L. lazarae and L. grandicentrum. The carapace is elongated unlike in L. planum and L. crassum. It has a long and narrow mesogastric region unlike in L. joecollinsi, L. musialiki, L. lanceatum, L. laeve, L. punctatum, L. sublaeve, L. laculatum, L. fraasi, L. icaunensis, L. hispanicum and L. crassum. Symmetric tubercles on the mesogastric region present in L. laeve are absent in L. ewakrzeminskae n. sp., but there are two tubercles on each side of the mesogastric region which are absent in L. joecollinsi, L. laeve, L. punctatum, L. laculatum, L. fraasi, L. icaunensis, L. edoi, L. planum, L. hispanicum and L. crassum. Epigastric, protogastric and hepatic regions are separated by transverse grooves as in L. laeve.
Material Examined
Type material. Holotype: NHMW 1990/0041/2744a; type locality: Ernstbrunn; age: Tithonian.
Paratypes. NHMW 1990/0041/5179, NHMW 2014/0194/0978; locality: Ernstbrunn; age: Tithonian.
Additional material. NHMW 1990/0041/3959b; locality: Ernstbrunn; age: Tithonian.
Dimensions. The state of preservation of the material from the NHMW collection is not good enough to take measurements of all specimens. The length of the mesogastric region of the holotype is 1.4 mm, and that of the paratypes are 1.7 and 1.4 mm, respectively. The width of the hepatic region of the holotype is 2.4 mm. The width at the branchial region of the holotype is 2.3 mm. As the posterior margin is not preserved on any of the specimens, it is not possible to measure the length of the carapace.
Description. The carapace is longer than wide, widest on the branchial region, strongly vaulted transversally and moderately vaulted longitudinally (Figure 7).
Only the base of the rostrum is preserved in specimen NHMW 2014/0194/0978 (Figure 7B). The anterior groove is deep, visible at the intersection of the rostrum and the upper orbital margin (Figure 7). The epigastric regions are rounded. The mesogastric region is very long, flattened with a very narrow anterior part. There is no separation between the anterior and posterior part of this region (Figure 7). The posterior gastric muscle scars are absent.
Gastric pits and mesogastric groove tubercles are absent. There are two hepatic tubercles on each side of the mesogastric region (Figure 7A, C). The epigastric, protogastric and hepatic regions are separated by transverse grooves (Figure 7B, D-E).
The urogastric region is undivided (Figure 7C). The cardiac region is distinctly bordered by the branchiocardiac groove, narrowing posteriorly, with two tubercles in the anterior part and a tubercle in the posterior part (Figure 7C).
The carapace is covered with granules only in its posterior part. They are visible on the internal mold and in places with the cuticle preserved. The grooves are wide and deep. The lateral parts of the postcervical groove are connected with the branchial groove. The branchiocardiac groove reaches the posterior margin of the carapace. The posterior margin of the carapace is not preserved in any of the specimens.
All of the specimens are almost complete carapaces not showing any traces of molting lines.
Laeviprosopon lanceatum n. sp.
Figure 1, Figure 8
zoobank.org/4A54E1D0-43A6-4786-A202-005D820F9D4D
Diagnosis. Carapace longer than wide. Rostrum with a very long central spine. Orbital margin round, surface posterior to orbits convex. Epigastric regions strongly vaulted. Large tubercle on each hepatic region. Epigastric, protogastric and hepatic regions separated by transverse grooves.
Etymology. The name lanceatum derives from the Latin word “lancea” - spear. It refers to the prominent rostrum.
Comparisons. The holotype of Laeviprosopon lanceatum n. sp. is much smaller than specimens of L. laeve, but there is not enough material to ascertain whether the small size is a true character of this species or the specimens are just juveniles. Its carapace is widest on the hepatic region, as in most species of the genus except L. joecollinsi n. sp., L. musialiki, L. lazarae and L. ewakrzeminskae n. sp. The carapace is elongated unlike in L. planum, L. crassum and L. hispanicum. The anterior and posterior part of the mesogastric region of L. lanceatum n. sp. is more or less the same length unlike in L. musialiki, L. laeve, L. lazarae, L. sublaeve, L. grandicentrum, L. laculatum, L. fraasi, L. planum, L. hispanicum and L. crassum. Symmetrical tubercles on the mesogastric region present in L. laeve are absent in L. lanceatum n. sp. A single large tubercle on each side of the posterior part of the mesogastric region present in L. lanceatum n. sp. is also present only in L. lazarae and L. grandicentrum. Epigastric, protogastric and hepatic regions are separated by transverse grooves as in L. laeve, L. joecollinsi, L. lazarae, L. sublaeve, L. ewakrzeminskae n. sp. and L. fraasi. Epigastric regions are rounded in L. lanceatum n. sp., not short and wide as in L. lazarae and L. sublaeve. Laeviprosopon lanceatum n. sp. has a rostrum with a very long central spine not noticed in any other species of the genus.
Material Examined
Type material. Holotype: I-F/MP/736/1502/08; type locality: Ogrodzieniec; age: early-middle Oxfordian.
Paratype. I-F/MP/342/1497/08; locality: Niegowonice; age: middle - late Oxfordian (upper elizabethae Subzone - upper wartae Subzone).
Dimensions. Specimens are not well preserved, and thus most measurements cannot be taken accurately. Length of the mesogastric region of the holotype is 0.9 mm, of the paratype 1.4 mm. Carapace width at the level of the hepatic region is 1.4 mm for the holotype and 1.6 mm for the paratype.
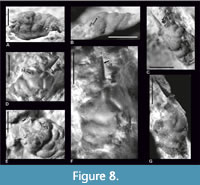 Description. The carapace is longer than wide, widest on the hepatic region, strongly vaulted transversally and moderately vaulted longitudinally (Figure 8A-B, F). The anterior part of the carapace in front of the cervical groove is relatively long.
Description. The carapace is longer than wide, widest on the hepatic region, strongly vaulted transversally and moderately vaulted longitudinally (Figure 8A-B, F). The anterior part of the carapace in front of the cervical groove is relatively long.
The rostrum is only preserved as an impression with a very long central spine preserved in both specimens. The base of rostrum is very wide, and the central spine is located between damaged bases of the lateral parts of rostrum (Figure 8C-F). The orbital margin is rounded. There is a groove separating the base of the rostrum and the orbital margin. The surface behind the orbits is slightly convex (Figure 8A, C, E).
The epigastric regions are rounded, strongly vaulted, partially preserved on both specimens. The mesogastric region is distinctly bounded by grooves. The anterior (narrow) and posterior (wide) part of this region are more or less the same length. The posterior part is the highest point of the carapace. There is no incision between the anterior and posterior part of this region (Figure 8).
A pair of small, rounded gastric pits is present in the cervical groove. There is a pair of large hepatic tubercles on both sides of the posterior part of the mesogastric region (one on each side) (Figure 8A-B, D, G). The epigastric, protogastric and hepatic regions are separated by transverse grooves (Figure 8A).
The urogastric region is convex and relatively broad, delimited by the postcervical groove. There are very narrow grooves laying parallel to the lateral parts of the branchiocardiac groove (Figure 8A). The cardiac region is not distinctly bordered, narrowing posteriorly, with two tubercles in the posterior part.
The grooves are very wide and deep. The cervical groove is the deepest. The internal mold of the carapace is covered with small tubercles. The cuticle is not preserved.
Laeviprosopon joecollinsi n. sp.
Figure 1, Figure 9
zoobank.org/C569B457-C53E-4130-88E9-2DD874756F88
 Diagnosis. Carapace longer than wide, widest on branchial region (length of holotype from epigastric region to posterior margin: 6.7 mm), strongly vaulted transversally and moderately vaulted longitudinally. Epigastric, protogastric and hepatic regions separated by transverse grooves. Internal mold of carapace densely covered with granules posteriorly, sparsely so in anterior part; granules also visible on cuticle. Shallow grooves laying parallel to branchial groove.
Diagnosis. Carapace longer than wide, widest on branchial region (length of holotype from epigastric region to posterior margin: 6.7 mm), strongly vaulted transversally and moderately vaulted longitudinally. Epigastric, protogastric and hepatic regions separated by transverse grooves. Internal mold of carapace densely covered with granules posteriorly, sparsely so in anterior part; granules also visible on cuticle. Shallow grooves laying parallel to branchial groove.
Etymology. The name joecollinsi is dedicated to Joseph (Joe) S.H. Collins (1927-2019), a specialist of fossil crustaceans, especially decapods.
Comparisons. The carapaces of L. laeve are less convex. Laeviprosopon joecollinsi n. sp. also differs from L. laeve in the absence of the symmetrical tubercles on the mesogastric region and in having a narrower posterior part of this region. The mesogastric region of L. joecollinsi n. sp. is not divided in the middle as in L. punctatum, L. laeve and L. musialiki. It has a distinct narrow anterior and broad posterior part more or less the same length unlike in L. lazarae, L. sublaeve, L. grandicentrum, L. laculatum, L. edoi, L. hispanicum, L. planum and L. crassum. The species has round epigastric regions, not short and wide as in L. sublaeve, L. fraasi, L. edoi, L. planum and L. hispanicum. The species does not have a transverse groove from the cervical to the branchio-cardiac groove as in L. sublaeve, L. fraasi, L. musialiki and L. icaunensis. The species does not have any large tubercles on the carapace as in L. musialiki, L. lanceatum n. sp. and L. lazarae. The epigastric, protogastric and hepatic regions are separated by transverse grooves as in L. laeve, L. lazarae, L. sublaeve, L. fraasi, L. ewakrzeminskae n. sp. and L. lanceatum n. sp.
Material Examined
Type material. Holotype: I-F/MP/6264/1599/12; type locality: Niegowonice; age: middle - late Oxfordian (upper Elizabethae Subzone - upper Wartae Subzone).
Paratypes. I-F/MP/5061/1543/09; type locality: Niegowonice; age: middle - late Oxfordian; I-F/MP/741/1502/08; type locality: Ogrodzieniec; age: early-middle Oxfordian; UF272115, UF272119; type locality: Szklary; age: middle Oxfordian.
Additional material. Niegowonice: I-F/MP/3037/1532/08; I-F/MP/3038/1532/08; I-F/MP/3873/1533/08; I-F/MP/4623/1534/08.
Dimensions. The state of preservation of the material is not good enough to take most of the measurements. The length of the holotype from the epigastric region to the posterior margin is 6.7 mm, 9.5 mm for paratype UF272115. The maximum width of paratype 5061 in the epibranchial region is 5.3 mm. The length of the mesogastric region of paratypes 5061 and 741 is 3.4 mm and 2.7 mm, respectively.
Description. The carapace is longer than wide, widest in the branchial region, strongly vaulted transversally and moderately vaulted longitudinally (Figure 9A, E, G-H).
The rostrum is not preserved on any specimen. The orbital margin is rounded, with a flattened suborbital spine (Figure 9A, marked with a black arrow). The anterior groove is distinct.
The epigastric regions are rounded and lay in front of the mesogastric region, which is distinctly bordered by grooves. The length of the anterior (narrow) part of this region is about 0.6x the posterior (wide) part, and the posterior part is the highest point of the carapace. There is no incision in the posterior border of this region. It is covered with many granules on the internal mold and the cuticle; gastric spines are absent. Epigastric, protogastric and hepatic regions are separated by transverse grooves (Figure 9C, E). A pair of small, elongated gastric pits is present in the cervical groove (Figure 9B).
The urogastric region is convex and divided into two parts by an incision, which connects to the postcervical groove (Figure 9A, D-E, H). Incisions laying on the epibranchial region, with the postcervical and branchial groove, bound surfaces directed to the cardiac region. The cardiac region is not distinctly bordered, widening anteriorly.
The grooves are very deep and wide. The cervical groove is the deepest. The branchiocardiac groove reaches the posterior margin of the carapace.
The posterior border of the carapace is rimmed and concave in the middle (Figure 9A, C). The whole surface of the carapace is covered with granules, which occur both on the internal mold and on the cuticle. In the anterior part, they are sparsely present, but they are denser in the posterior part. On the branchial region, they are positioned in lines parallel to the branchial groove (Figure 9C-D).
Genus Navarrahomola Artal, Van Bakel, Fraaije, Jagt, Klompmaker, 2012
Type species. Navarrahomola hispanica Artal, Van Bakel, Fraaije, Jagt, Klompmaker, 2012
Diagnosis. As in Artal et al. (2012: p. 405).
Navarrahomola hispanica Artal, Van Bakel, Fraaije, Jagt, Klompmaker, 2012
2012 Navarrahomola hispanica Artal et al., p. 405, figure 3.1, 3.3 (non 3.2).
2013 Navarrahomola hispanica Artal et al. - Klompmaker, p. 171, figure 12.
Diagnosis. As for the genus.
Remarks. Navarrahomola hispanica was described from the upper Albian reefal limestones of Spain (Monte Orobe and Koskobilo quarries). One of the paratypes (MGSB28141) is not conspecific to the holotype and the other paratype because the lateral borders of the mesogastric region are more continuously curved and the anterior carapace contains fewer large tubercles compared to N. hispanica (Artal et al., 2012: pl. 3.1, 3.3; Klompmaker, 2013: figure 12). Instead, MGSB28141 likely represents the largest known specimen of Laeviprosopon hispanicum, a species also reported from Koskobilo (Klompmaker, 2013). This specimen compares best to the specimen figured in Klompmaker (2013: figure 6H), which also shows evenly spaced granules, unlike on smaller specimens in which no clear granules are visible. Thus, we ascribe MGSB28141 to Laeviprosopon hispanicum for now. Apparently, granules become only visible on the internal mold in larger specimens for this species.
DISCUSSION
Reassignment of Laeviprosopon to Homolidae
The overall shape of the carapace, regions and grooves show similarities between Laeviprosopon and Homolidae. For example, the Cretaceous species of Laeviprosopon from Spain, namely L. edoi, is similar to Homolopsis glabra Wright and Collins, 1972, in the shape of the mesogastric, hepatic and cardiac regions, the shape of grooves and the overall shape of the carapace. This species of Laeviprosopon has a relatively smooth carapace, while this Homolopsis is covered with small tubercles. This difference applies to other taxa as well. Representatives of Laeviprosopon have relatively smooth carapaces with some species exhibiting small and sparse tubercles, mostly in the posterior part, whereas most of the homolid genera and species have numerous large tubercles on their carapaces, which might be an argument against transferring Laeviprosopon to Homolidae. However, the ornamentation of the carapace is a variable character and might differ between genera of one family or even within species of one genus (Prosopon abbreviatum Schweitzer and Feldmann, 2009, and Prosopon mammillatum Woodward, 1868, Prosopon and Protuberosa in Schweitzer and Feldmann, 2009; Planoprosopon and Dioratiopus in Schweitzer and Feldmann, 2011). Among Homolidae, most of the genera have strong tubercles, but some species of Homolopsis have a smooth carapace (Bishop and Brannen, 1992, figure 1; Collins, 1997, figure 3). Moreover, many other characters match as mentioned above. Based on these data, herein we transfer the genus Laeviprosopon to the superfamily Homoloidea and family Homolidae. As specimens of Laeviprosopon show the oldest unequivocal molting lines on most of the dorsal carapace, we speculate that they may have given rise to other homoloids with molting lines entirely visible on the dorsal carapace in the Late Jurassic or Early Cretaceous.
Molting Lines in Jurassic and Cretaceous Homolodromioidea and Homoloidea - Homology or Homoplasy?
Homolodromioidea. The molting process in Homolodromioidea is insufficiently known. In the Jurassic and in modern homolodromioids, no molting lines are present on the dorsal carapace, along which it could break. There are only large, poorly calcified areas on the latero-ventral sides in modern representatives - the branchiostegites (Ng et al., 2008). The molting lines in these regions are clearly visible on the flanks or in ventral view in fossils ascribed to Homolodromioidea (e.g., Schweitzer and Feldmann, 2008; Klompmaker et al., 2011, 2012, 2016, 2020; Starzyk, 2013, 2015a), but they are rarely illustrated.
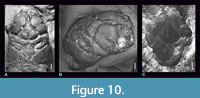 Regarding Laeviprosopon, insufficient preservation is often an issue when trying to determine the position of the molting line. As Franţescu (2011) wrote, the posterolateral sides of the carapace in Laeviprosopon lazarae are not preserved most likely due to their weak calcification or post-mortem damages, therefore it is not possible to ascertain the presence or absence of molting lines in that species. The carapace of Laeviprosopon punctatum is preserved with its lateral sides and shows no clear molting lines (Figure 10A-B, Schweitzer and Feldmann, 2008, figure 1.7; Klompmaker et al., 2020, figure 13B). Laeviprosopon musialiki has only small parts of the lateral flanks damaged, and there are also no remnants of molting lines visible (Starzyk, 2020, figure 3).
Regarding Laeviprosopon, insufficient preservation is often an issue when trying to determine the position of the molting line. As Franţescu (2011) wrote, the posterolateral sides of the carapace in Laeviprosopon lazarae are not preserved most likely due to their weak calcification or post-mortem damages, therefore it is not possible to ascertain the presence or absence of molting lines in that species. The carapace of Laeviprosopon punctatum is preserved with its lateral sides and shows no clear molting lines (Figure 10A-B, Schweitzer and Feldmann, 2008, figure 1.7; Klompmaker et al., 2020, figure 13B). Laeviprosopon musialiki has only small parts of the lateral flanks damaged, and there are also no remnants of molting lines visible (Starzyk, 2020, figure 3).
 However, the molting line on the dorsal carapace of L. laculatum is present; specimen NHMW 1990/0041/4221 is a molt. The margin of this molt is partially destroyed on the left side, but the right side is similar to the one in L. laeve (Figure 10C; Schweitzer and Feldmann, 2008, figure 1.9). Most of the specimens of L. fraasi have the posterolateral borders preserved. The molting line is preserved on the specimen NHMW 1990/0041/4051 (Figure 6B, E) and is also visible in the reconstruction of Wehner (1988, figure 32A), but there are no signs of such a line on the holotype. Patrulius (1966), Wright and Collins (1972) and Collins and Wierzbowski (1985) also mentioned the presence of linea homolica on specimens of L. fraasi. Prosopidae, to which Laeviprosopon was assigned repeatedly, do not bear molting lines on their dorsal carapaces, just as with other representatives of Homolodromioidea. Molting lines are probably laying on the ventral side of the carapace as in Tanidromites montreuilensis Crônier and Boursicot, 2009 (Figure 11A) and in other homolodromioids (see Karasawa et al., 2006, figures 1, 2; Klompmaker, 2013, figure 5; Klompmaker et al., 2011, figures 3, 5, 6; Klompmaker et al., 2020, figures 10, 11, 14, 15). Unfortunately, only the dorsal aspects are available or illustrated for many other homolodromioid taxa.
However, the molting line on the dorsal carapace of L. laculatum is present; specimen NHMW 1990/0041/4221 is a molt. The margin of this molt is partially destroyed on the left side, but the right side is similar to the one in L. laeve (Figure 10C; Schweitzer and Feldmann, 2008, figure 1.9). Most of the specimens of L. fraasi have the posterolateral borders preserved. The molting line is preserved on the specimen NHMW 1990/0041/4051 (Figure 6B, E) and is also visible in the reconstruction of Wehner (1988, figure 32A), but there are no signs of such a line on the holotype. Patrulius (1966), Wright and Collins (1972) and Collins and Wierzbowski (1985) also mentioned the presence of linea homolica on specimens of L. fraasi. Prosopidae, to which Laeviprosopon was assigned repeatedly, do not bear molting lines on their dorsal carapaces, just as with other representatives of Homolodromioidea. Molting lines are probably laying on the ventral side of the carapace as in Tanidromites montreuilensis Crônier and Boursicot, 2009 (Figure 11A) and in other homolodromioids (see Karasawa et al., 2006, figures 1, 2; Klompmaker, 2013, figure 5; Klompmaker et al., 2011, figures 3, 5, 6; Klompmaker et al., 2020, figures 10, 11, 14, 15). Unfortunately, only the dorsal aspects are available or illustrated for many other homolodromioid taxa.
Homoloidea. As mentioned before, Laeviprosopon laeve has been transferred several times between the families Prosopidae (Homolodromioidea) and Homolidae (Homoloidea), giving rise to doubts about the presence or absence of the linea homolica. New specimens reported herein showing undoubted molting lines indicate that Laeviprosopon laeve is hitherto the oldest known species with a linea homolica. Specimens of L. fraasi from Tithonian also show linea homolica. For these reasons, Laeviprosopon is confirmed as a member of Homoloidea.
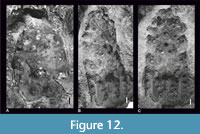 Until this paper, there was only one member of the family Homolidae known from the Jurassic, Doerflesia ornata Feldmann and Schweitzer, 2009, (Figure 12A) and four species of the family Tithonohomolidae (Figure 12B-C) from Štramberk and Ernstbrunn. They all have a confirmed linea homolica (Feldmann and Schweitzer, 2009, figures 2, 4, 5). The molting line in these species extends parallel to the anterolateral and posterolateral margins of the carapace, reaching the anterior margin (Figure 11).
Until this paper, there was only one member of the family Homolidae known from the Jurassic, Doerflesia ornata Feldmann and Schweitzer, 2009, (Figure 12A) and four species of the family Tithonohomolidae (Figure 12B-C) from Štramberk and Ernstbrunn. They all have a confirmed linea homolica (Feldmann and Schweitzer, 2009, figures 2, 4, 5). The molting line in these species extends parallel to the anterolateral and posterolateral margins of the carapace, reaching the anterior margin (Figure 11).
On the carapaces of the post-Jurassic representatives of Homolidae, the linea homolica extends in a different way than the representatives of Laeviprosopon (compare Figure 11B-C): it is visible on the dorsal side of the carapace along its entire length from the orbits to the posterior margin, for example in the genera Homolopsis (Figure 11C; Collins, 1997, figure 3.1, 3.3), Ariecornibus (Franţescu, 2014, figure 7), Peedeehomola (Garassino et al., 2015, figure 5), Zygastrocarcinus (Collins, 1997, figure 6.1), Latheticocarcinus (Feldmann and Schweitzer, 2015, figure 2.1; Schweitzer et al., 2004, figure 2), Cretalamoha (Nyborg and Garassino, 2017, figures 1, 2), Paromola (Nyborg and Garassino, 2017, figure 3, 4) Nogarhomola (De Angeli and Alberti, 2012, figure 2, pl. 1) and Paromolopsis (Schweitzer et al., 2004, figure 4). Linea homolicae extends this way also in recent Homolidae such as Homola (Guinot and Richer de Forges, 1995, figure 8B-D, 14B, 14D, 14F), Paromola (Guinot and Richer de Forges, 1995, figure 21, 23), Latreillopsis (Guinot and Richer de Forges, 1995, figure 29A, B, F, G), Hypsophrys (Guinot and Richer de Forges, 1995, figure 59B, D, F) and Homolomannia (Guinot and Richer de Forges, 1995, figure 63B-D, F).
Molting Process Conclusions. Laeviprosopon laeve, L. laculatum and L. fraasi are the only Jurassic species of Laeviprosopon in which it is possible to ascertain the presence of a molting line (Figure 6B, E; Figure 10C). This line extends partially through the dorsal side of the carapace. There are no carapaces of L. laeve with lateral parts preserved in the collection of the ISEA PAS or the NHMW collection. There is only one specimen (SMNS 70527/11), which is a complete carapace, that does not have any remains of molting line (Figure 5H).
In the reconstruction of L. fraasi presented by Wehner (1988, figure 32A), there is a molting line extending in a similar way as in L. laeve, but there are no traces of such a line on the holotype. In contrast, the molting line is visible on specimen NHMW 1990/0041/4051 (Figure 6B, E). It is located on the right side, where the posterolateral border is preserved.
The specimens of L. leave, L. fraasi and L. laculatum that show a molting line raise the question whether specimens of the same species developed molting lines only in some periods of their lifetime because not all specimens show a clear molting line. The mentioned specimen of L. fraasi might show a premolting stage, when the carapace was about to break at the molting line. In that case, there is a possibility that other Jurassic specimens of Laeviprosopon, with the posterolateral carapaces preserved and no signs of molting lines, molted in the same way. Their carapaces could break through lines similar to L. laeve, where molting lines are laying on the dorsal surface of the branchial and epibranchial regions and crossing on the latero-ventral side of the hepatic region, not reaching the orbits (Figure 5, Figure 11B).
Other Jurassic and Cretaceous species of Laeviprosopon do not show any dorsal molting lines when complete carapaces are preserved (Figure 7, Figure 8, Figure 9, Figure 10; Wehner, 1988, pl. 8, figures 3, 4; Schweitzer and Feldmann, 2008, figure 1). In other cases, the lateral sides are not shown or studied. They might have molted in a way similar to L. laeve, but our state of knowledge is insufficient to confirm this. However, other morphological characters in the Jurassic and Cretaceous Laeviprosopon species provide convincing arguments for a close relationship between L. laeve and other species of Laeviprosopon that have not revealed their molting lines so far.
Among the Jurassic Homolidae and Tithonohomolidae, there are only molds broken along the linea homolica; therefore, it is uncertain whether the linea was developed on the carapace between molts (Figure 12; Feldmann and Schweitzer, 2009, figures 2, 4, 5).
Specimens of Homolidae from the Aptian, Albian and geologically younger stages described and documented in many papers (Figure 12C; Wright and Collins, 1972, pls. 6, 7, 8, figure 1; Bishop, 1986, figures 2.2, 3.1, 4.1, 4.2, 4.4; Bishop and Brannen, 1992, figure 1; Collins, 1997, figures 3.1, 3.2, 3.3, 9; Crawford, 2008, figures 2, 3; Feldmann et al., 2013, figure 7) show clear linea homolicae also when the carapace is not broken along it. It might mean it was visible during the time interval between molts for at least some Cretaceous species.
Contrary, the linea homolica does not seem to be visible on some of the carapaces of the Cretaceous homoloid species: Ariecornibus schweitzerae Franţescu, 2014 (Franţescu, 2014, figure 7C) and Zygastrocarcinus richardsoni (Woodward, 1896) (Bishop, 1986). Both of these specimens have lateral sides of the carapace preserved at least partially. There are also appendages visible on the specimen of A. schweitzerae, which suggests they are carcasses rather than molts. Similar to L. fraasi, molting lines in these specimens may have become visible only before molting. This observation in other Mesozoic homoloids supports placement of Laeviprosopon in Homoloidea. In contrast, molting lines are consistently visible during the time interval between molts in recent homoloids (Guinot and Richer de Forges, 1995, figures 8, 14, 29, 59, 63; Richer de Forges and Ng, 2007, figures 2, 3, 6, 7).
Because of the preservation of Jurassic homoloid crabs, it is hard to evaluate whether their molting pattern is homologous to that of the Cretaceous and younger ones. Other characters such as the thoracic sternum are usually not preserved in Jurassic crabs either; therefore, it is not possible to rely on them (Guinot, 2019). However, our research sheds some light on this process in the oldest brachyurans. In most Jurassic Laeviprosopon species (except L. fraasi), there are no confirmed molting lines developed between molting episodes (opposite to most Homoloidea, see paragraph above). This might be a matter of different preservational conditions in different localities and geological ages or, more likely, a genuine pattern fairly common in Jurassic homoloids, less common in Cretaceous specimens, and absent in modern homoloids.
The genus Laeviprosopon may be a possible phyletic link between Homolodromioidea and Homoloidea. Based on the current state of knowledge, it seems to be the first group with molting lines on the dorsal surface of the carapace, at least partially (Figure 11B). A possible scenario of molting line evolution is that the posterior part of the molting line shifted from the ventral to the dorsal surface of the carapace faster than the anterior part. In Laeviprosopon, its posterior part lies on the dorsal surface, but it is still present on the ventral side when reaching the cervical groove. This type of molting line lasted at least to the Albian and is present on the carapaces of L. crassum, L. edoi, L. hispanicum and L. planum. Subsequently, the molting line of homoloids was present solely on the dorsal carapace.
ACKNOWLEDGEMENTS
NS thanks Mike Reich for a warm welcome and help during her visit of the Bayerische Staatssammlung für Paläontologie und Geologie in Munich. Horst Kuschel kindly donated specimens for this study. AAK thanks the University of California Museum of Paleontology for access to photography equipment for the UF paratypes of L. joecollinsi and the Florida Museum of Natural History (invertebrate paleontology) for equipment to prepare those specimens. Roger Portell (Florida Museum of Natural History) is thanked for the loan of those paratypes. We would also like to thank the reviewers who provided useful input that improved this paper.
REFERENCES
Alcock, A. 1900. Materials for a carcinological fauna of India, No. 5. The Brachyura Primigenia or Dromiacea. Journal of the Asiatic Society of Bengal, 68:123–169.
Artal, P., Van Bakel, B.W.M., Fraaije, R.H.B., Jagt, J.W.M., and Klompmaker, A.A. 2012. New Albian-Cenomanian crabs (Crustacea, Decapoda, Podotremata) from Monte Orobe, Navarra, northern Spain. Revista Mexicana de Ciencias Geológicas, 29:398—410.
Beurlen, K. 1928. Die fossilen Dromiaceen und ihre Stammesgeschichte. Paläontologische Zeitschrift, 10:144—183. https://doi.org/10.1007/BF03041570
Bishop, G. 1986. A new crab, Zygastrocarcinus cardsmithi (Crustacea, Decapoda), from the Lower Pierre Shale, Southern Montana. Journal of Paleontology, 60:1097—1102. https://doi.org/10.1017/S0022336000022629
Bishop, G. and Brannen, N.A. 1992. Homolopsis pikeae, new species (Decapoda), a crab from the Cretaceous of Texas. Journal of Crustacean Biology, 12:317—323. https://doi.org/10.2307/1549084
Collins, J.S.H. 1997. Fossil Homolidae (Crustacea; Decapoda). Bulletin of the Mizunami Fossil Museum, 24:51—71.
Collins, J.S.H. and Wierzbowski, A. 1985. Crabs from the Oxfordian sponge megafacies of Poland. Acta Geologica Polonica, 35:73—88.
Crônier, C. and Boursicot, P.-Y. 2009. A new decapod crustacean faunule from the Middle Jurassic of North-West France. Palaeontology, 52(6):1275—1289. https://doi.org/10.1111/j.1475-4983.2009.00896.x
Crawford, R.S. 2008. A new species of fossil homolid crab (Decapoda: Brachyura) from the Rίo Foyel Formation (Paleogene), Rίo Negro Province, Argentina. Journal of Paleontology, 82:835—841. https://doi.org/10.1666/06-111.1
De Angeli, A. and Alberti, R. 2012. Nogarhomola aurorae n. gen., n. sp. (Decapoda, Brachyura, Homolidae) dell’Eocene medio dei Monti Lessini orientali (Vicenza, Italia settentrionale). Studi Trentini di scienze naturali, 92:63—70.
De Haan, W. 1833-1850. Crustacea. In von Siebold, P.F. (ed.), Fauna Japonica sive Descriptio Animalium, quae in Itinere per Japoniam, Jussu et Auspiciis Superiorum, qui summum in India Batava Imperium Tenent, Suscepto, Annis 1823-1830 Collegit, Notis, Observationibus et Adumbrationibus Illustravit: i—xvii, i—xxxi, ix—xvi, 1—243, pls. A—J, L-Q, 1—55, circ. tab. 2, J. Müller et Co., Lugduni Batavorum.
Feldmann, R.M. and Tshudy, D. 1987. Ultrastructure in cuticle from Hoploparia stokesi (Decapoda: Nephropidae) from the López de Bertodano Formation (Late Cretaceous-Paleocene) of Seymour Island, Antarctica. Journal of Paleontology, 61:1194—1203. https://doi.org/10.1017/S0022336000029565
Feldmann, R.M., Tshudy, D.M., and Thomson, M.R.A. 1993. Late Cretaceous and Paleocene decapod crustaceans from James Ross Basin, Antarctic Peninsula. Memoir of the Palaeontological Society, 28:1—41. https://doi.org/10.1017/S0022336000062077
Feldmann, R.M. and Gaździcki, A. 1998. Cuticular ultrastructure of fossil and living homolodromiid crabs (Decapoda: Brachyura). Acta Palaeontologica Polonica, 43:1—19.
Feldmann, R.M. and Schweitzer, C.E. 2009. Revision of Jurassic Homoloidea De Haan, 1839, from the Ernstbrunn and Štramberk limestones, Austria and the Czech Republic. Annalen des Naturhistorischen Museums in Wien, 111A:183—206.
Feldmann, R.M., Schweitzer, C.E., Baltzly, L.M., Bennett, O.A., Jones, A.M.R., Mathias, F.F., Weaver, K.L., and Yost, S.L. 2013. New and previously known decapod crustaceans from the Late Cretaceous of New Jersey and Delaware, USA. Bulletin of the Mizunami Fossil Museum, 39:7—37.
Feldmann, R.M. and Schweitzer, C.E. 2015. Latheticocarcinus punctatus (Rathbun, 1917) (Decapoda, Brachyura) from the Ranch 777 site, Custer County, South Dakota, USA. Bulletin of the Mizunami Fossil Museum, 41:1—5.
Förster, R. 1985. Frühe Anomuren und Brachyuren (Decapoda, Crustacea) aus dem mittleren Dogger. Mitteilungen der Bayerischen Staatssammlung für Paläntologie und Historische Geologie, 25:45—60.
Fraaije, R.H.B., Krzemiński, W., Van Bakel, B.W.M., Krzemińska, E., and Jagt, J.W.M. 2012a. The earliest record of pylochelid hermit crabs from the Late Jurassic of southern Poland, with notes on paguroid carapace terminology. Acta Palaeontologica Polonica, 57:647—654. https://doi.org/10.4202/app.2011.0002
Fraaije, R.H.B., Krzemiński, W., Van Bakel, B.W.M., Krzemińska, E., and Jagt, J.W.M. 2012b. The sixth abdominal tergites of paguroid anomurans - a newly recognized crustacean macrofossil type. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, 266:115—122. https://doi.org/10.1127/0077-7749/2012/0273
Fraaije, R.H.B., Krzemiński, W., Van Bakel, B.W.M., Krzemińska, E., and Jagt, J.W.M. 2014. New Late Jurassic symmetrical hermit crabs from the southern Polish Uplands and early paguroid diversification. Acta Palaeontologica Polonica, 59:681—688. https://doi.org/10.4202/app.2012.0022
Fraaije, R.H.B., Klompmaker, A.A., Jagt, J.W.M., Krobicki, M., and Van Bakel B.W.M. 2022. A new, highly diverse paguroid assemblage from the Oxfordian (Upper Jurassic) of southern Poland and its environmental distribution. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, 304:1–12. https://doi.org/10.1127/njgpa/2022/1054
Franţescu, O. 2011. Brachyuran decapods (including five new species and one new genus) from Jurassic (Oxfordian-Kimmeridgian) coral reefs limestones from Dobrogea, Romania. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, 259:271—297. https://doi.org/10.1127/0077-7749/2010/0110
Franţescu, O. 2014. Fossil decapods from the Cretaceous (late Albian) of Tarrant County, Texas. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, 273:221—239. https://doi.org/10.1127/0077-7749/2014/0427
Garassino, A. and Krobicki, M. 2002. Galicia marianae n. gen., n. sp. (Crustacea, Decapoda, Astacidea) from the Oxfordian (Upper Jurassic) of the Southern Polish Uplands. Bulletin of the Mizunami Fossil Museum, 29:51—59.
Garassino, A., Clements, D., and Vega, F.J. 2015. A new homolid crab, Peedeehomola deanbogani n. gen., n. sp., from the Peedee Formation (late Maastrichtian), Rocky Point Member, North Carolina. Paleontología Mexicana, 4:25—29.
Glaessner, M.F. 1929. Crustacea Decapoda, p. 1-464. In Pompeckj, J.F. (ed.), Fossilium Catalogus: Animalia, 41, Berlin.
Glaessner, M.F. 1933. Die Krabben der Juraformation. Centralblatt für Mineralogie, Geologie und Paläontologie, Abteilung B, 3:178—191.
Głowniak, E. 2012. The perisphinctid genus Prososphinctes Schindewolf (Ammonoidea, subfamily Prososphinctinae nov.): An indicator of palaeoecological changes in the Early Oxfordian Submediterranean sea of southern Poland. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, 264:117—179. https://doi.org/10.1127/0077-7749/2012/0235
Guinot, D. 2019. New hypotheses concerning the earliest brachyurans (Crustacea, Decapoda, Brachyura). Geodiversitas, 41:747—796. https://doi.org/10.5252/geodiversitas2019v41a22
Guinot, D. and Richer de Forges, B. 1995. Crustacea Decapoda Brachyura: Révision de la familie des Homolidae de Haan, 1839, p. 283—517. In Crosnier, A. (ed.), Résultats des campagnes MUSORSTOM, Volume 13. Mémoires du Muséum national d'histoire naturelle 163, Editions du Muséum, Paris.
Hoffmann, M., Kołodziej, B., and Skupien, P. 2017. Microencruster-microbial framework and synsedimentary cements in the Štramberk Limestone (Carpathians, Czech Republic): Insights into reef zonation. Annales Societatis Geologorum Poloniae, 87:325—347.
Karasawa, H., Kato, H., and Terabe, K. 2006. A new member of the family Prosopidae (Crustacea: Decapoda: Brachyura) from the Lower Cretaceous of Japan. Revista Mexicana de Ciencias Geológicas, 23:344—349.
Klompmaker, A.A. 2013. Extreme diversity of decapod crustaceans from the mid-Cretaceous (late Albian) of Spain: Implications for Cretaceous decapod paleoecology. Cretaceous Research, 41:150—185. https://doi.org/10.1016/j.cretres.2012.12.003
Klompmaker, A.A., Artal, P., and Gulisano, G. 2011. The Cretaceous crab Rathbunopon: Revision, a new species and new localities. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, 260:191—202. https://doi.org/10.1127/0077-7749/2011/0170
Klompmaker, A.A., Feldmann, R.M., and Schweitzer, C.E. 2012. A hotspot for Cretaceous goniodromitids (Decapoda: Brachyura) from reef associated strata in Spain. Journal of Crustacean Biology, 32:780—801. https://doi.org/10.1163/193724012X635340
Klompmaker, A.A., Flores-Ventura, J., and Vega, F.J. 2013. A new homolid crab, Zygastrocarcinus carolinasensis n. sp., from the Cretaceous (Campanian) of NE Mexico: Implications for paleobiogeography. Boletín de la Sociedad Geológica Mexicana, 65:265—271.
Klompmaker, A.A., Chistoserdov, A.Y., and Felder, D.L. 2016. Possible shell disease in 100 million-year-old crabs. Diseases of Aquatic Organisms, 119:91—99. http://doi.org/10.3354/dao02988
Klompmaker, A.A., Starzyk, N., Fraaije, R.H.B., and Schweigert, G. 2020. Systematics and convergent evolution of multiple reef-associated Jurassic and Cretaceous crabs (Decapoda, Brachyura). Palaeontologia Electronica, 23(2):a32. https://doi.org/10.26879/1045
Krzemińska, E., Krzemiński, W., Fraaije, R.H.B., Van Bakel, B.W.M., and Jagt, J.W.M. 2016. Allometric ontogenetic changes in two Late Jurassic gastrodorid hermit crabs (Crustacea, Decapoda, Anomura) from central Europe. Journal of Systematic Palaeontology, 14:139—148. http://doi.org/10.1080/14772019.2015.1018968
Latreille, P.A. 1802. Histoire naturelle, générale et particulière, des Crustacés et des Insectes, v. 3. F. Dufart, Paris.
Leinfelder, R.R., Krautter, M., Laternser, R., Nose, M., Schmid, D.U., Schweigert, G., Werner, W., Keupp, H., Brugger, H., Herrmann, R., Rehfeld-Kiefer, U., Schroeder, J.H., Reinhold, C., Koch, R., Zeiss, A., Schweizer, V., Christmann, H., Menges, G., and Luterbacher, H. 1994. The origin of Jurassic reefs. Current research developments and results. Facies, 31:1—56.
Leinfelder, R.R. and Keupp, H. 1995. Upper Jurassic mudmounds: Allochthonous sedimentation versus autochthonous carbonate production, p. 17—26. In Neuweiler, F. and Reitner, J. (ed.), Mudmounds: A polygenetic spectrum of fine-grained carbonate buildups. Facies 32,.
Leinfelder, R.R., Werner, W., Nose, M., Schmid, D.U., Krautter, M., Laternser, R., Takaćs, M., and Hartmann, D. 1996. Paleoecology, growthparameters and dynamics of coral, sponge and microbolite reefs from the Late Jurassic, p. 227—248. In Reitner, J., Neuweiler, F., and Gunkel, F. (eds.), Global and regional controls on biogenic sedimentation. I. Reef evolution. Research Reports. Göttinger Arbeiten zur Geologie und Paläontologie, Sonderband 2:.
Linnaeus, C. 1758. Systema Naturae per Regna tria Naturae, secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis (ed. 10), vol 1. Laurentius Salvius, Holmiae. https://doi.org/10.5962/bhl.title.542
Matyja, B.A. and Wierzbowski, A. 1996. Sea-bottom relief and bathymetry of Late Jurassic sponge megafacies in Central Poland, p. 333—340. In Riccardi, A.C. (ed.), Advances in Jurassic Research. GeoResearch Forum, 1—2.
Moericke, W. 1889. Die Crustaceen der Stramberger Schichten. Palaeontographica (Supplement-Bände), 2(6):43—72.
Moericke, W. 1897. Die Crustaceen der Stramberger Schichten. Palaeontographica, Supplement, 2(6):43—72.
Müller, P., Krobicki, M., and Wehner, G. 2000. Jurassic and Cretaceous primitive crabs of the family Prosopidae (Decapoda: Brachyura) - their taxonomy, ecology, and biogeography. Annales Societatis Geologorum Poloniae, 70:49—79.
Ng, P.K.L., Guinot, D., and Davie, P.J.F. 2008. Systema Brachyurorum: Part I. An annotated checklist of extant brachyuran crabs of the world. The Raffles Bulletin of Zoology, 17:1—286.
Nyborg, T. and Garassino, A. 2017. New occurrences of fossil Homolidae from the Eastern Pacific. Boletín de la Sociedad Geológica Mexicana, 69:135—148. https://www.redalyc.org/journal/943/94350664006/
Patrulius, D. 1966. Decapoda dolnego tytonu z Woźnik (Polskie Karpaty Zachodnie). Rocznik Polskiego Towarzystwa Geologicznego, 36:495—519.
Quenstedt, F.A. 1856—1857. Der Jura. Laupp, Tübingen.
Quenstedt, F.A. 1885. Handbuch der Petrefactenkund. Laupp, Tübingen.
Remeš, M. 1895. Beiträge zur Kenntnis der Crustaceen der Stramberger Schichten. Bulletin International de l’Académie des Sciences de Bohème, 2:200—204.
Remeš, M. 1905. Die Fauna der sogenannten exotischen Blöcke des Strambergerkalksteins in Rychalitz (Mähren). Bulletin International de l’Académie des Sciences de Bohème, 10:33—37.
Richer de Forges, B. and Ng, P.K.L. 2007. New records and new species of Homolidae De Haan, 1839, from the Philippines and French Polynesia (Crustacea: Decapoda: Brachyura). The Raffles Bulletin of Zoology, 16:29—45.
Schneider, S., Harzhauser, M., Kroh, A., Lukeneder, A., and Zuschin, M. 2013. Ernstbrunn Limestone and Klentnice beds (Kimmeridgian-Berriasian; Waschberg-Ždánice Unit; NE Austria and SE Czech Republic): State of the art and bibliography. Bulletin of Geosciences, 88:105—130.
https://doi.org/10.3140/bull.geosci.1360
Schweigert, G. 2019. The Upper Kimmeridgian limestones of Saal an der Donau (Bavaria, Germany) - the most diverse Late Jurassic crustacean fauna of a coral reef in Germany. 7th Symposium on Mesozoic and Cenozoic Decapod Crustaceans - Book of Abstracts, Ljubljana, p. 21—22.
Schweigert, G., Kuschel, H., and Scherzinger, A. 2016. Following the steps of Hermann v. Meyer: A newly collected brachyuran fauna from Late Jurassic sponge-microbial limestones near Geislingen an der Steige, SW Germany. - 6th Symposium on Mesozoic and Cenozoic Decapod Crustaceans, Villers-Sur-Mer, Normandy, France, p. 63—64.
Schweigert, G. and Kuschel, H. 2018. Schwäbische und Fränkische Alb - ein Paradies für Jura-Krabben. Fossilien, 35:14—23.
Schweitzer, C.E. and Feldmann, R.M. 2008. Revision of the genus Laeviprosopon Glaessner, 1933 (Decapoda: Brachyura: Homolodromioidea: Prosopidae) including two new species. Neues Jahrbuch für Geologie und Paläontologie, Abhandungen, 250:273—285. https://doi.org/10.1127/0077-7749/2008/0250-0273
Schweitzer, C.E. and Feldmann, R.M. 2009. Revision of the Prosopinae sensu Glaessner, 1969 (Crustacea: Decapoda: Brachyura) including four new families, four new genera, and five new species. Annalen des Naturhistorischen Museums in Wien, 110A:55—121.
Schweitzer, C.E. and Feldmann, R.M. 2011. Revision of some fossil podotrematous Brachyura (Homolodromiidae; Longodromitidae; Tyronommidae). Neues Jahrbuch für Geologie und Paläontologie, Abhandungen, 260:237—256. https://doi.org/10.1127/0077-7749/2011/0138
Schweitzer, C.E., Nyborg, T.G., Feldmann, R.M., and Ross, R.L.M. 2004. Homolidae De Haan, 1839 and Homolodromiidae Alcock, 1900 (Crustacea: Decapoda: Brachyura) from the Pacific Northwest of North America and a reassessment of their fossil records. Journal of Paleontology, 78:133—149. https://doi.org/10.1666/0022-3360(2004)078<0133:HDHAHA>2.0.CO;2
Starzyk, N. 2013. Jurassic crabs: New characters of carapace diagnostic to known tanidromitid species. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, 269:173—187. https://doi.org/10.1127/0077-7749/2013/0343
Starzyk, N. 2015a. Reappraisal of the primitive crab Eodromites, with description of three new species from European localities (Decapoda: Brachyura: Goniodromitidae). Palaeontologia Electronica, 18.3.50A:1—19. https://doi.org/10.26879/513
Starzyk, N. 2015b. New species of the genus Tanidromites (Decapoda: Brachyura: Tanidromitidae) from the Oxfordian of Poland. Acta Zoologica Cracoviensia, 58:129—136. https://doi.org/10.3409/azc.58_2.129
Starzyk, N. 2016. Three new species of the genus Tanidromites (Decapoda: Brachyura: Tanidromitidae) from the Late Jurassic (Oxfordian) of Poland. Palaeontologia Electronica, 19.3.45A:1—14. https://doi.org/10.26879/619
Starzyk, N. 2020. The oldest species of the genus Laeviprosopon (Decapoda: Brachyura: Prosopidae) from the Oxfordian of Poland. Acta Zoologica Cracoviensia, 63:9—13. https://doi.org/10.3409/azc.63.02
Starzyk, N., Krzemińska, E., and Krzemiński, W. 2011. Intraspecific variation in the Jurassic crab Bucculentum bucculentum (Decapoda: Homolodromioidea: Bucculentidae). Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, 260:203–210. https://doi.org/10.1127/0077-7749/2011/0169
Starzyk, N., Krzemińska, E., and Krzemiński, W. 2012. A new crab species from the Oxfordian of Poland (Decapoda: Brachyura: Goniodromitidae). Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, 263:143—153. https://doi.org/10.1127/0077-7749/2012/0219
Van Straelen, V. 1925. Contribution à l’étude des crustacés décapodes de la période jurassique. Mémoires d’Academie Royale de Belgique, Classe des Sciences, collected in number 4, series 2, 7:1—462.
Van Straelen, V. 1936. Crustacés décapodes nouveaux ou peu connus de l'époque crétacique. Bulletin du Musée royal d'Histoire naturelle de Belgique, 13:1—50.
Vašíček, Z., Skupien, P., and Jagt, J.W.M. 2018. Current knowledge of ammonite assemblages from the Štramberk Limestone (Tithonian-Lower Berriasian) at Kotouč Quarry, Outer Western Carpathians (Czech Republic). Cretaceous Research, 90:185—203. https://doi.org/10.1016/j.cretres.2018.04.016
von Meyer, H. 1857. Briefliche Mittheilungen, an Prof. Bronn gerichtet. Neues Jahrbuch für Mineralogie, Geognosie, Geologie und Petrefaktenkunde, 1857:556.
von Meyer, H. 1860. Die Prosoponiden oder die Familie der Maskenkrebse. Palaeontographica, 7:183—222.
Wehner, G. 1988. Über die Prosoponiden (Crustacea, Decapoda) des Jura. Dissertation zur Erlangung des Doktorgrades der Fakultät für Geowissenschaften der Ludwig-Maximilians-Universität zu München; PhD thesis, München.
Woodward, H. 1868. On a new brachyurous Crustacean (Prosopon mammillatum) from the Great Oolite, Stonesfield. Geological Magazine, 5:3—5, pl. 1. https://doi.org/10.1017/S0016756800207140
Wright, C.W. and Collins, J.S.H. 1972. British Cretaceous crabs. Palaeontographical Society Monographs, 126(533):1—113.
