

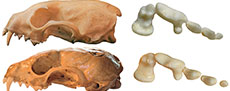
One or two species? Revision of fossil martens from the late Early Pleistocene sites Deutsch Altenburg 2 and 4 (Austria)
Article number: 27.2.a32
https://doi.org/10.26879/1397
Copyright Society of Vertebrate Paleontology, June 2024
Author biographies
Plain-language and multi-lingual abstracts
PDF version
Submission: 12 April 2024. Acceptance: 24 May 2024.
ABSTRACT
Marten fossils from the Austrian sites Deutsch Altenburg 2 and 4 have been revisited. Despite the previous view on the possible occurrence of two marten species, Martes cf. zibellina and Martes vetus, only the presence of the latter has been confirmed. Previous identifications of marten remains as M. zibellina are considered to be related to sexual dimorphism characteristic for M. vetus. The well-preserved viscerocranium and mandibles found at Deutsch Altenburg 2 and 4 are important materials in the European fossil record of this species. The revised material of M. vetus from Deutsch Altenburg 2 and 4 was compared with fossils from other European Early and early Middle Pleistocene sites, which showed that the species has intermediate features shared with M. martes, M. zibellina, and M. foina.
Adrian Marciszak. Department of Paleozoology, University of Wrocław, Sienkiewicza 21, 50-335 Wrocław, Poland. Corresponding author. adrian.marciszak@uwr.edu.pl
Doris Nagel. Institute of Paleontology, University Vienna, Josef-Holaubek-Platz 2, 1090 Wien, Austria, Vienna, Austria. doris.nagel@univie.ac.at
Keywords: Martes; morphotypes; biostratigraphy; sexual dimorphism
Final citation: Marciszak, Adrian and Nagel, Doris. 2024. One or two species? Revision of fossil martens from the late Early Pleistocene sites Deutsch Altenburg 2 and 4 (Austria). Palaeontologia Electronica, 27(2):a32.
https://doi.org/10.26879/1397
palaeo-electronica.org/content/2024/5240-fossil-martens-from-austria
Copyright: June 2024 Society of Vertebrate Paleontology.
This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
creativecommons.org/licenses/by/4.0
INTRODUCTION
The origin of the genus Martes Pinel, 1792, has been uncertain for a long time, although new findings have substantially improved our understanding of the evolution of martens (Flynn et al., 2005; Koepfli et al., 2008; Samuels et al., 2018). The earliest species that has been referred to the Guloninae is Martes laevidens Dehm, 1950, from the German site Wintershof West, dated to the early Miocene (MN 3, 20-17 Mya; Dehm, 1950, 1953). However, based on basicranial anatomy, mainly on the incompletely ossified supramental fossa, this assignment was questioned (Wolsan, 1993; Sato et al., 2003), and the feature was regarded as a plesiomorphic trait shared by different mustelids (Hughes, 2012). Among gulonines, the genus Martes has been consistently found to be the most closely related to the genus Gulo Pallas, 1780, and a sister group to Pekania pennanti (Erxleben, 1777) (Koepfli et al., 2008; Wolsan and Sato, 2010; Sato et al., 2012; Li et al, 2014; Malyarchuk et al., 2015; Zhu et al., 2016; Samuels et al., 2018). Koepfli et al. (2008) estimated the time when the Guloninae diverged from the mustelid branch at c. 11.0 Mya (12.5-9.4 Mya), while Sato et al. (2012) at c. 12.7 Mya (14.7-10.8 Mya). The study of mitochondrial genomes of gulonines showed that the Martes-Gulo clade diverged from Pekania around 7.6 Mya (8.9-7.1 Mya) (Li et al., 2014). The split between the genus Martes and the genus Gulo was assessed to have occurred 6.4-6.3 Mya (7.6-5.3 Mya) (Li et al., 2014), while Malyarchuk et al. (2015) suggested a time of 5.6 Mya (6.3-4.9 mya). These molecular estimates corroborate well with the earliest records of definite true gulonines from the late Miocene, i.e., Pekania occulta Samuels and Cavin, 2013, from North America and Martes palaeosinensis Zdansky, 1924, from Asia (Wang et al., 2012; Samuels and Cavin, 2013). According to them, true martens from the genus Martes appeared even later, c. 4.2-4.0 Mya (Stach, 1959; Wolsan, 1989b; Anderson, 1994; Sato et al., 2003; Montoya et al., 2011).
The Middle-Late Miocene history of possible ancestors of the genus Martes is full of marten-like species and forms, often of debatable and unclear taxonomical position (Anderson, 1994; Baskin, 1998; Ginsburg, 1999; Hughes, 2012). Some of these forms show morphological features closely related to those in the Guloninae, but many others represent stem groups outside of the crown clade Guloninae (Anderson, 1994; Sato et al., 2003; Wang et al., 2012; Li et al., 2014). In addition, some of these Martes -like mustelids show some similarity to the ischyrictines (Ginsburg and Morales, 1992; Montoya et al., 2011) or even have been referred to the ischyrictine genera Hoplictis Ginsburg, 1961, Plionictis Matthew, 1924, and Sthenictis Peterson, 1910 (Anderson, 1994; Baskin, 1998; Hughes, 2012; Samuels and Cavin, 2013; Samuels et al., 2018). Morphological similarities of these Martes -like mustelids with members of the Guloninae are likely the result of their ecomorphological convergence or retention of plesiomorphic traits (Samuels et al., 2018). Considering the high level of polymorphism observed in these early taxa, comparable with the highly polymorphic dentition of extant taxa and the scarcity of fossil materials, a detailed revision is needed to fully understand their taxonomic position (Samuels et al., 2018).
It is debatable whether the early Late Miocene (MN 9-10) Martes melibulla Petter, 1963, or Late Pliocene (MN 15) Martes wenzensis Stach, 1959, is the first true Martes (Valenciano, 2017; Valenciano et al., 2020, 2021). Contrary to the Miocene, the Eurasian Pliocene history of the genus is considerably poorly documented. Since Stach’s (1959) description, M. wenzensis has been regarded as the only European Pliocene marten (Anderson, 1970; Marciszak et al., 2024). The Early to the mid-Middle Pleistocene Eurasian martens are represented by M. vetus Kretzoi, 1942 (2.2-0.6 Mya; Heller, 1930, 1933, 1936; Brunner, 1933; Kretzoi, 1942, 1945; Dehm, 1962; Kurtén, 1968; Anderson, 1970; Wiszniowska, 1989; Ambros et al., 2005; Ambros, 2006; Marciszak, 2012; Marciszak et al., 2021, 2024) and M. crassidens Jiangzuo, Gimranov, Liu, Liu, Jin and Liu, 2021 (2.2-1.7 Mya; Jiangzuo et al., 2021). Although most authors tentatively interpret M. wenzensis as the most likely ancestor of the Martes lineage. The main goal of this paper is to revise old and to describe new materials of M. vetus from the famous paleontological site Deutsch Altenburg (Austria), the only Early Pleistocene site with marten fossils in Europe. The finds from this locality shed new light on the late Early Pleistocene evolution of the genus Martes and the relation of M. vetus with M. martes (Linnaeus, 1758) and M. foina (Erxleben, 1777). The article also tests the assumption about the hypothetical presence of a second European marten species, M. zibellina.
MATERIALS AND METHODS
All studied materials of Martes vetus are housed in the Department of Paleontology, University of Vienna, Austria. Measurements were taken with an electronic calliper to the nearest 0.01 mm, according to the schemes presented in Appendix 1 (Figure S1, Figure S2, Figure S3). Each value given here is the mean of three measurements. Measurements were also taken using an Olympus set for image analysis (Olympus stereo microscope ZSX 12, Olympus DP 71 camera, and Cell D software). This set coupled with a Canon EOS 5D camera was also used to take photographs. The dental terminology and tooth morphotypes follow Anderson (1970) and Gimranov and Kosintsev (2015). For comparison, materials from the following sites were used (unless otherwise noted, measurements were made personally by A.M.): Kamyk (1.9-1.8 Mya), Schernfeld (1.9-1.6 Mya), Żabia Cave (1.7-1.5 Mya), Kielniki 1 (1.3-1.1 Mya), Zalesiaki 1A (1.3-1.1 Mya), Somssich Hill 2 (1.1-1.0 Mya; Gasparik and Pazonyi, 2018), Stránská Skála (0.9-0.8 Mya), Kozi Grzbiet (0.8-0.7 Mya), and Południowa Cave (0.8-0.7 Mya).
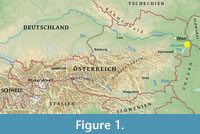 The site complex Deutsch Altenburg is located in the Hollitzer quarry (Pfaffenberg, the western foothill of the Hainburger Berge), where Mesozoic dolomites have been mined (Figure 1; Frank and Rabeder, 1997a, 1997b). It is near the Austrian-Slovak border (Bruck an der Leitha district, Lower Austria) (Figure 1). The first site discovered in 1900 was the Middle Pleistocene locality Deutsch Altenburg 1 (Toula, 1902, 1906; Freudenberg, 1914; Nagel and Rabeder, 1997). In 1908-1984, numerous crevices and caves were exposed during blasting (Pfeiffer-Deml, 2016). As a result, 52 localities were excavated in the quarry, each being referred to in chronological order of their discovery (Frank and Rabeder, 1997a, 1997b).
The site complex Deutsch Altenburg is located in the Hollitzer quarry (Pfaffenberg, the western foothill of the Hainburger Berge), where Mesozoic dolomites have been mined (Figure 1; Frank and Rabeder, 1997a, 1997b). It is near the Austrian-Slovak border (Bruck an der Leitha district, Lower Austria) (Figure 1). The first site discovered in 1900 was the Middle Pleistocene locality Deutsch Altenburg 1 (Toula, 1902, 1906; Freudenberg, 1914; Nagel and Rabeder, 1997). In 1908-1984, numerous crevices and caves were exposed during blasting (Pfeiffer-Deml, 2016). As a result, 52 localities were excavated in the quarry, each being referred to in chronological order of their discovery (Frank and Rabeder, 1997a, 1997b).
The most well-known site, Deutsch Altenburg 2 (DA2; 48°9’10” N, 16°55’2” E, c. 255 m a.s.l.), was discovered in 1971. The uppermost layers contain mostly microvertebrate remains (layer DA2A). Located below are brown coarse sands (layer DA2C). Layer DA2C1 consists of light brown grit interstratified with dolomite rubble and sandstone concretions. It is covered by sandstone slabs. The sands are of fluviatile origin and were probably sedimented by a precursor of the Danube. The karst cavity of the DA2C1 was connected to a wide shaft, which once passed the connection to the former surface of the Pfaffenberg. The shaft was filled mainly with rubble and blocks. Between them, partly hardened clay lenses were intercalated, which contained abundant molluscs and vertebrates. It consists also of fluviatile sand, which probably was delivered by the precursor of the Danube. The shaft was named Deutsch Altenburg 4 (280 m a.s.l.) and the clay deposits in the rubble were named layer DA4B. The Martes vetus remains analyzed in the paper were found in two horizons, DA2C1 and DA4B, which probably belonged to the same cave system. Previously it was connected to the surface of Pfaffenberg by a shaft of about 40 m wide (Rabeder et al., 2010).
Biochronological analysis of micromammals, mostly rodents, allow the age of the DA2C1 horizon to be estimated as 1.3-1.2 Mya (Rabeder et al., 2010). The dominance of the genus Microtus (“Allophaiomys”- horizon) with the basal species Mimomys pliocaenicus Forsyth-Major, 1902, but also other arvicolids such as Lagurus arankae Kretzoi, 1954, Prolagurus pannonicus Kormos, 1930, Mimomys coelodus Kretzoi, 1954, Mimomys savini Hinton, 1910, Clethrionomys hintonianus Kretzoi, 1958, Pliomys episcopalis Méhely, 1914, and Ungaromys nanus Kormos, 1932, stratigraphically position DA2C very well into the Microtus pliocaenicus zone (Table S1 in Appendix 1; Rabeder, 1976, 1981; Rabeder et al., 2010). The layer DA4B is younger by one Microtus zone and is estimated at about 1.1-1.0 Mya (Table S1 in Appendix 1; Rabeder, 1981; Rabeder et al., 2010).
In DA2C, 98 vertebrate taxa were identified, including seven amphibians, 11 reptiles, four birds, and 76 mammals. The latter are represented by 14 insectivorans, 18 bats, 21 rodents, two lagomorphs, 15 carnivorans, one proboscidean, two perissodactyls, two artiodactyls, and one primate. From DA4B, remains of 98 vertebrate taxa were identified, including four amphibians, 12 reptiles, 15 birds, and 67 mammals. The latter are represented by 10 insectivorans, 13 bats, 25 rodents, two lagomorphs, 12 carnivorans, one proboscidean, two perissodactyls, one artiodactyl, and one primate (Table S1 in Appendix 1; Ehrenberg, 1929; Mais, 1971, 1973, 1978; Rabeder, 1972a, 1972b, 1972c, 1973a, 1973b, 1974a, 1974b, 1974c, 1976, 1978, 1981, 1982; Mais and Rabeder, 1977a, 1977b, 1979, 1984, 1997; Jánossy, 1981; Fladerer, 1984; Frank and Rabeder, 1997a, 1977b; Sapper, 1997; Rabeder and Withalm, 2006; Rabeder et al., 2010).
SYSTEMATIC PALEONTOLOGY
Class MAMMALIA Linnaeus, 1758
Order CARNIVORA Bowdich, 1821
Suborder CANIFORMIA Kretzoi, 1943
Infraorder ARCTOIDEA Flower, 1869
Parvorder MUSTELIDA Tedford, 1976
Superfamily MUSTELOIDEA Fischer, 1817
Family MUSTELIDAE Fischer, 1817
Genus MARTES Pinel, 1792
Martes vetus Kretzoi, 1942
Figure 2, Figure 3, Figure 4, Figure 5, Figure 6, Figure 7; Table 1, Table 2, Table 3
Material. Deutsch Altenburg 2C: a viscerocranium with a small part of the left neurocranium and preserved left I1, I3, C1, P2-P4, and right I1-M1; right P4; three M1 (2 left, 1 right); a left mandible without most of the ramus and present i3-m2; a right mandible without the ramus, with c1, p2-m3, and m1 (DA/2275/14/77); a right complete mandible with i3-c1 and p2-m2; a right c1; two trigonids, of the left and right m1. Deutsch Altenburg 4B: a left mandible with missing symphyseal part and with p2 and p4; a right mandibular body fragment with m1.
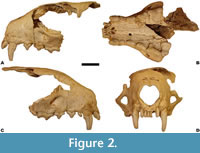 Emended diagnosis. Marten of about the size of Martes martes and M. foina; cranium with a short and broad viscerocranium; broad incisor row moderately extended forward; moderately convex frontal profile with gently marked concavity in the middle part; compressed, short, and broad temporal region with a broad and short postorbital bar situated almost exactly in the middle of the temporal region; large, inflated, and strongly convex tympanic bullae; short and broad palate, especially in its distal part; V-shaped and convex median indentation of palate; C1 with broad and short crown; large and oval P1; narrow P2; narrow P3 with weakly marked convex margin; P4 with short and low protocone, whose length is smaller than the distal breadth of P4; M1 with trigon length slightly greater than the talon length, highly complicated microrelief on the occlusal surface and moderately large metacone; mandible with a long and slim body, gently convex mandibular lower margin under m1 and vertical edges of the ramus; triangular and shallow masseteric fossa; moderately spaced mental foramens; large p1; m1 with a short trigonid and small metaconid; oval m2; postcranial bones morphologically resembling those of M. martes.
Emended diagnosis. Marten of about the size of Martes martes and M. foina; cranium with a short and broad viscerocranium; broad incisor row moderately extended forward; moderately convex frontal profile with gently marked concavity in the middle part; compressed, short, and broad temporal region with a broad and short postorbital bar situated almost exactly in the middle of the temporal region; large, inflated, and strongly convex tympanic bullae; short and broad palate, especially in its distal part; V-shaped and convex median indentation of palate; C1 with broad and short crown; large and oval P1; narrow P2; narrow P3 with weakly marked convex margin; P4 with short and low protocone, whose length is smaller than the distal breadth of P4; M1 with trigon length slightly greater than the talon length, highly complicated microrelief on the occlusal surface and moderately large metacone; mandible with a long and slim body, gently convex mandibular lower margin under m1 and vertical edges of the ramus; triangular and shallow masseteric fossa; moderately spaced mental foramens; large p1; m1 with a short trigonid and small metaconid; oval m2; postcranial bones morphologically resembling those of M. martes.
Description. The viscerocranium is short and wide, with a well-marked broadening at the level of canines and P4. The frontal profile in lateral view is moderately convex, with a well-marked concavity in the middle of the frontal part. The frontal line runs somewhat mesially to the nasal bones and depresses slightly on the boundary between the cerebral and facial areas. The muzzle area is broadened at the canines, narrowed behind them, and then gradually widened Table 1). The rostral area is proportionally short and massive. The nasal aperture is large and rounded, and the nasal bones are broad, short, and W-shaped. The oval and large orbits have minute lacrimal processes. The infraorbital foramina are large and oval. The upper tooth row is curved distally. The teeth are loosely arranged with short diastemas between I3 and C1, P1 and P2, and P2 and P3; the longest one is between I3 and C1. The broad and curved incisor row is moderately extended forward (Figure 2).
The simple build I1 is relatively short mesio-distally and moderately developed bucco-lingually. The crown is asymmetric, and its main axis runs obliquely from the upper-lingual to the lower-buccal side. The apex of the crown is oriented mesio-buccally and forms an asymmetric triangle. The mesial edge is larger than the distal one. The mesial valley between them is V-shaped, shallow, and both edges are connected to each other. The I2 has a massive and compact crown with a tip directed distally and slightly buccally. The buccal part of the crown is concave and separated from the convex lingual side by a thin and V-shaped cingulum, almost evenly developed mesially and distally. The large, canine-like I3 has a crown placed at an angle of 40o to the root axis. The crown, ovoid or oval in cross-section, is also flattened on both sides, especially on the lingual side. The top of the crown is twisted disto-lingually. Two edges extend from it, mesial and distal, which at the base of the crown connect with the moderately developed lingual cingulum, forming a thin ridge around the crown. The C1 are flattened laterally and relatively short mesio-distally; weak mesial and distal crests run from their apex to the base. The oval-shaped P1 is a small, monocuspid, single-rooted tooth. Its buccal and lingual margins are convex, and the lingual one is developed more strongly.
The P2 is an elongated tooth, with a stronger distal cingulum. The buccal margin is straight, and the lingual margin is slightly convex in its middle part. The mesial and distal margins are blunt. The protoconid is situated more mesially. Two thin edges run from the apex of the protoconid, the mesial one in a mesio-lingual direction. The distal edge ends exactly medially and is connected with the distal cingulum. The P3 is elongated and narrow. The crown bears an elongated distal cingular projection. The small mesio-buccal prominence of the cingulum forms a faint mesial crest to the apex of the protoconid. This main cusp is located medially and slightly mesially. The two edges running from the apex of the protoconid are thick and sharp. The distal one is not connected with the distal cingulum, and its end forms a small, swelling-like bulge. The cingulum is weakly developed on the lingual side. The mesial and distal margins are blunt or rounded, while the buccal margin bears a moderately developed, median concavity. There is a moderate convexity at the same level, but on the lingual margin.
The long and narrow P4 has straight buccal and lingual margins of the talon. The mesial and distal margins are blunt or rounded. The moderately high paracone bears a thin crest across the mesial border from the apex to the base of the crown. It is separated from the moderately long and low protocone, whose mesial margin is aligned with that of the paracone. Its length is, on average, smaller than the distal breadth of the crown (Table 2). The metacone is separated from the paracone by a deep valley. The cingulum is more strongly developed on the lingual margin of the metacone. The M1 is large, its breadth is smaller than the length of P4 (Table 2). The trigon is moderately wide and short, with a moderate and abrupt concavity of the buccal margin. The paracone and the metacone are elongated and well developed; the paracone is larger. They are well separated by a deep, narrow, V-shaped valley. Apexes of both main cusps are connected by a thin, long crest. The talon is shorter than the trigon and they are separated by a deep and broad depression running through the middle part of the crown. The protocone is low and long. It is divided by a shallow and wide valley into two parts of similar length and height. The reduced metaconule is not connected with any other cusp or crest, and is a low, elongated cuspid situated in the middle part of the crown. A long and thin crest corresponding to the buccal margin of the cingulum runs in parallel along the whole talon length. The well-developed lingual cingulum forms a thick crest.
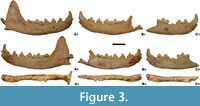 The mandibular body is long and moderately high (Figure 3). Its height measured behind the m1 is comparable to the m1 length (Table 3). Two rounded mental foramens are moderately spaced and similar in size. The mesial one is situated under the p2, while the distal mental foramen is located under the distal root of p3, slightly lower than the mesial one (Figure 3). The masseteric fossa is moderately deep and its rounded, mesial edge reaches the m1 and m2 boundary. The mesial part narrows dorso-ventrally and its ventral margin only slightly exceeds the midline of the mandibular body in dorso-ventral direction. The lover mandibular body margin forms a gently curved arch, uniformly domed mesially and distally, with the strongest curvature under m1. The symphysial part is moderately massive and elongated. The row of cheek teeth is almost straight and only the distal parts of the p2-p3 crowns moderately arch lingually. In the tooth row, the premolars are located more buccaly in relation to the molars. As a result, the lingual margin of the p4 is displaced more buccally relative to the lingual border of the m1 (Figure 3).
The mandibular body is long and moderately high (Figure 3). Its height measured behind the m1 is comparable to the m1 length (Table 3). Two rounded mental foramens are moderately spaced and similar in size. The mesial one is situated under the p2, while the distal mental foramen is located under the distal root of p3, slightly lower than the mesial one (Figure 3). The masseteric fossa is moderately deep and its rounded, mesial edge reaches the m1 and m2 boundary. The mesial part narrows dorso-ventrally and its ventral margin only slightly exceeds the midline of the mandibular body in dorso-ventral direction. The lover mandibular body margin forms a gently curved arch, uniformly domed mesially and distally, with the strongest curvature under m1. The symphysial part is moderately massive and elongated. The row of cheek teeth is almost straight and only the distal parts of the p2-p3 crowns moderately arch lingually. In the tooth row, the premolars are located more buccaly in relation to the molars. As a result, the lingual margin of the p4 is displaced more buccally relative to the lingual border of the m1 (Figure 3).
The premolars are loosely arranged with diastemas between c1 and p1, p2 and p3, and p3 and p4. All teeth are situated at a similar level. The triangular crown of the i3 is double cusped, with the main protoconid occupying a larger surface. It is slightly asymmetrical, being more developed on the lingual side. Its vertical top is massive and rather blunt. The distoconid, located laterally to it on the buccal side, has the top of the crown directed slightly disto-buccally. It is a well-developed cuspid, although its shape and size are subject to considerable variability. It is separated from the protoconid by a thin and deep medial notch running almost to the base of the crown, which, in fact, divides the crown into two parts. The c1 is long and robust, with a proportionally short and hook-shaped crown (Figure 3; Table 3).
Two longitudinal grooves run on the buccal and lingual sides of the c1 crown. The relatively large and weakly reduced p1 is an oval, small, and one-rooted tooth. It is tightly squeezed between c1 and p2. Contrary to other premolars, the axis of its crown runs in an arrangement from mesio-lingual to bucco-distal. The two-rooted p2 is low-crowned, with the protoconid strongly displaced mesially. Its occlusal outline is almost rectangular, with an elongated distal part. Two thin crests run in the mesial and distal direction from the top of the protoconid. The distal cingulum forms a thin ridge, collaring the smooth area in the distal part. The mesial part of the crown is longer than the distal one. It bears an elongated distal cingular projection. The mesial and distal margins are blunt, while the buccal and lingual margins are almost straight. The larger p3 has a similar outline in occlusal view, with straight buccal and lingual margins. The mesial and distal margins are blunt. The protoconid is also displaced mesio-medially, although to a lesser degree than in p2. The distal part of p3 is shorter than the mesial one. An elongated distal, cingular projection is oriented slightly disto-buccally. Two thin edges run from the apex of the protoconid. On the distal edge, a small tubercular convexity is present just behind the top. The mesial and distal cingulum are relatively strongly developed. The two-rooted p4 is relatively high-crowned and has the protoconid placed almost exactly centrally and pushed slightly mesially. A relatively large hypoconid is present behind the protoconid. It is associated with the distal crest, running distally from the apex of the protoconid. The mesial ridge is thinner than the distal one. The mesial and distal halves of the tooth are equal in length. The mesial part is narrower than the distal one. The crown is slightly broadened distally. There is a gentle lingual convexity in the middle part of the crown. The lingual margin is straight. The mesial margin is blunt, while the distal one is rounded. The mesial and distal cingulum are relatively strongly developed. The distal, cingular projection is less elongated compared to the rest of the crown. Collared by a thick cingulum, the inner surface of this projection is crescent-shaped and shallow.
The elongated and large m1 has a proportionally short and massive trigonid with a low paraconid and high protoconid. The long and low talonid is similar in breath to the trigonid. The medium-sized, but well-recognized metaconid is connected with the protoconid. A thin, longitudinal ridge surrounds the talonid field, ends on the metaconid base, and runs from the elongated and low hypoconid. The edge of the paraconid is weakly developed. The mesial margin is rounded, while the distal one is blunt. The buccal margin is almost straight, with a gentle concavity at the transition between the trigonid and talonid. The lingual margin of the paraconid and half of the protoconid is straight, while there is a moderate convexity more distally. The cingulum is moderately developed. The m2 is a moderately reduced, one-rooted tooth. The crown has a slightly irregular, rounded occlusal outline, with a length slightly exceeding the breadth. On the trigonid are located the large and low paraconid and protoconid. The larger paraconid is located mesio-buccaly, while the smaller, but not lower protoconid is situated medially and lingually. The talonid is narrower, with a conical and low hypoconid. The moderately developed cingulum is stronger only on the distal margin.
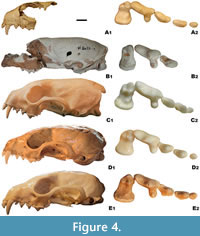 Comparison. Being the same metrically, the DA2 marten morphologically differs more from Martes martes than from M. foina. The viscerocranium is short and broad as in M. foina, with a broad and less extended forward incisor row. In lateral view, the frontal profile is weakly convex, with a slightly marked crossing between the maxillae and frontal region (Figure 4). In M. martes, the rostrum is proportionally longer and narrower, with a similarly less marked widening of the facial part at the level of canines.
Comparison. Being the same metrically, the DA2 marten morphologically differs more from Martes martes than from M. foina. The viscerocranium is short and broad as in M. foina, with a broad and less extended forward incisor row. In lateral view, the frontal profile is weakly convex, with a slightly marked crossing between the maxillae and frontal region (Figure 4). In M. martes, the rostrum is proportionally longer and narrower, with a similarly less marked widening of the facial part at the level of canines.
No particular differences were found in the morphology of the upper incisors and canines. The C1 of all three marten species are comparable dimensionally, where males are larger than females. On average, the C1 of Martes vetus is slightly more robust, but the ranges of variation strongly overlap. Morphologically, the C1 of M. vetus is characterised by a more curved, shorter and wider crown, which resembles more the C1 of M. foina than the more elongated and narrower C1 of M. martes. The right P1 of the DA2C marten measured 2.74 mm, which falls well into the range of variability of M. vetus (2.69 mm, 2.46-2.94 mm, n = 13) and M. martes (2.59 mm, 2.29-3.21 mm, n = 129), but distinctly exceed values obtained for M. foina (1.97 mm, 1.78-2.49 mm, n = 114). The P1 of M. vetus is not only larger, but also more robust, and the B/L index for this species is 68.5 (62.4-72.8, n = 13) compared to 69.7 in the DA2 marten. The P1 of M. martes is even more massive (71.9, 66.4-77.8, n = 129), while that of M. foina is notably narrower and more reduced (55.6, 52.9-64.3, n = 114). The P3 of M. vetus is similar to that of M. foina in having a narrow crown, with a weakly-marked convex margin (Figure 4). The P3 B/L mean ratio for M. vetus (48.7) is similar to that of M. foina (47.7). Morphotype A1, which is characteristic for M. foina (97%), without the lingual bulge, occurs also in the DA2 marten. This morphotype rarely occurs in M. martes (19%), for which the morphotype A2 is the most typical, with a small to moderate lingual bulge (73%). This species possesses much broader P3 (B/L = 62.4), which is consistent with the strongly developed lingual convexity.
The P4 of the DA2 marten is moderately long and low, and it is slightly shorter than the distal breadth of the crown (Figure 4). The protocone length to the distal breadth (L pr/Bp) index in the DA2 specimens (97.1, 96.2-98.6, n = 3) corresponds well with the data obtained for Martes vetus (94.5). Martes foina has distinctly shorter protocone (80.8), while its length in M. martes exceeds the distal breadth of P4 (L pr/Bp = 105.7). The P4 of the DA2 marten represents morphotype A2, with a notch between the protocone and parastyle, non-protruding protocone, and a concave buccal outline. This morphotype often occurs in M. foina (33%) and M. martes (24%). The P4 from the Sackdilling holotype belong to morphotype A1, with a notch between the protocone and parastyle, the protocone being shifted mesialy to the parastyle, and a concave buccal outline (Marciszak et al., 2021). This is also the most common morphotype in extant martens (58% in both). Other morphotypes are much rarer (Gimranov and Kosintsev, 2015).
The M1 of the DA2 marten has a relatively wide trigon, visibly broader than that in the Sackdilling holotype. The index of the trigon length to the M1 breadth (L ta/L tr) in DA2 is higher (78.7, 75.9-81.4, n = 4) than that obtained for the Sackdilling marten (69.0, 65.5-72.5, n = 2) (Marciszak et al. 2021). Data from both localities coincide with values of this index for Martes vetus (66.8, 59.2-76.1, n = 58). In this matter, M. vetus shows intermediate values between M. foina (62.6, 58.0-67.5, n = 114) and M. martes (75.4, 68.3-85.3, n = 129). The outline is irregular, and the occlusal surface is more complicated than in most specimens of M. foina and M. martes. The single M1 of the DA2 marten was assigned to morphotype A2, with straight lingual and rounded buccal outline and the preprotocrista bearing two small cusps. This morphotype (55%), together with A1 (32%), is the most common for M. martes, while others occur only rarely (Gimranov and Kosintsev, 2015). In M. foina, only group B morphotypes occur, with a deep groove between the paracone and metacone (Gimranov and Kosintsev, 2015). This is a reliable specific feature for M. foina, which was already pointed out by Wolsan et al. (1985) and Wolsan (1988, 1989a). The sole M1 from Sackdilling represents morphotype D2, with a straight lingual and rounded buccal outline, where the preprotocrista possesses two small cuspids and the hypocone and metaconule are present.
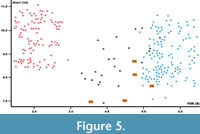 Previous authors, e.g., Heller (1930, 1933, 1936), Brunner (1933), or Dehm (1962), who described and studied remains of Martes vetus, practically did not deal with the mandible morphology. Only Anderson (1970) noted that the mandibular body is relatively shallow, the masseteric fossa reaches its maximum at the m1/m2 border, and the mental foramens are situated relatively far apart. She noted that the distance between them in M. vetus is 4.96 mm (3.70-6.10 mm, n = 13), while the mental foramens spacing ranged between 3.70-7.40 mm in M. martes and 2.00-3.40 mm in M. foina. A detailed revision showed that this feature is the most useful for taxonomic classification of European martens. The obtained results are close to the data presented by Anderson (1970), where the spacing of the mental foramens in the DA2 marten is 4.98 mm (4.02-5.74 mm, n = 5), and well corroborates with that of M. vetus (4.77 mm, 3.78-5.94 mm, n = 28). This measurement shows intermediate values between that of M. foina (2.53 mm, 1.97-3.41 mm, n = 114) and M. martes (6.13 mm, 5.36-7.97 mm, n = 129) (Figure 5).
Previous authors, e.g., Heller (1930, 1933, 1936), Brunner (1933), or Dehm (1962), who described and studied remains of Martes vetus, practically did not deal with the mandible morphology. Only Anderson (1970) noted that the mandibular body is relatively shallow, the masseteric fossa reaches its maximum at the m1/m2 border, and the mental foramens are situated relatively far apart. She noted that the distance between them in M. vetus is 4.96 mm (3.70-6.10 mm, n = 13), while the mental foramens spacing ranged between 3.70-7.40 mm in M. martes and 2.00-3.40 mm in M. foina. A detailed revision showed that this feature is the most useful for taxonomic classification of European martens. The obtained results are close to the data presented by Anderson (1970), where the spacing of the mental foramens in the DA2 marten is 4.98 mm (4.02-5.74 mm, n = 5), and well corroborates with that of M. vetus (4.77 mm, 3.78-5.94 mm, n = 28). This measurement shows intermediate values between that of M. foina (2.53 mm, 1.97-3.41 mm, n = 114) and M. martes (6.13 mm, 5.36-7.97 mm, n = 129) (Figure 5).
Considering other features as well, the mandible of Mares vetus is morphologically more similar to that of M. martes in having an elongated and moderately high mandibular body, the condyloid processes located slightly below or at the level of p4/m1, and a similar shape of the mandibular ramus. In M. vetus and M. martes, the mesial and distal edges of the coronoid process are approximately of the same length and form a not fully isosceles triangle. In M. foina, the mesial edge of this process is more inclined than the distal one, which is almost vertical and therefore considerably shorter. That is why, in relation to the mandibular body, the ramus is proportionally shorter and more vertical.
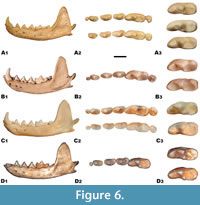 The morphology of the angular process is more variable; however, there are some differences (Figure 6). The morphology of the angular process in the DA2 marten strongly resembles that in Martes vetus and M. martes, which is longer and hooked, contrary to the short and straight angular process of M. foina. There are also some differences in the shape of the masseteric fossa, which, on average, is shallower with a more triangular mesial edge in M. vetus and M. martes. In M. foina, this structure is usually slightly deeper, with a more rounded mesial edge. However, the shape of the masseteric fossa is also highly variable. It is rather more related to age and sex (in males and older individuals the masseteric fossa is deeper), than to species-specific characteristics. Therefore, the diagnostic values of this feature is limited and cannot be taken into consideration during species discrimination. The mandible of M. vetus resembles that of M. foina and differs from that of M. martes simultaneously in having a strongly curved mandibular body, with a notably marked curvature of the lower margin under m1. In contrast, the mandibular body of M. martes is straighter, with a lower margin having no or only a gentle curvature. The variability of all the above features is relatively smaller than in most teeth, but still relevant and therefore diagnostic value of those characters should be considered with caution.
The morphology of the angular process is more variable; however, there are some differences (Figure 6). The morphology of the angular process in the DA2 marten strongly resembles that in Martes vetus and M. martes, which is longer and hooked, contrary to the short and straight angular process of M. foina. There are also some differences in the shape of the masseteric fossa, which, on average, is shallower with a more triangular mesial edge in M. vetus and M. martes. In M. foina, this structure is usually slightly deeper, with a more rounded mesial edge. However, the shape of the masseteric fossa is also highly variable. It is rather more related to age and sex (in males and older individuals the masseteric fossa is deeper), than to species-specific characteristics. Therefore, the diagnostic values of this feature is limited and cannot be taken into consideration during species discrimination. The mandible of M. vetus resembles that of M. foina and differs from that of M. martes simultaneously in having a strongly curved mandibular body, with a notably marked curvature of the lower margin under m1. In contrast, the mandibular body of M. martes is straighter, with a lower margin having no or only a gentle curvature. The variability of all the above features is relatively smaller than in most teeth, but still relevant and therefore diagnostic value of those characters should be considered with caution.
There are also some differences between the studied martens in the morphology of the lower dentition. Our studies confirmed all previously known differences and revealed a few others. The great size variability of the c1 of Martes vetus was already highlighted by Heller (1933). When comparing size values of different marten species, differences seemingly result from sexual dimorphism. The c1 of M. vetus is on average slightly narrower than c1 of M. martes and M. foina. However, the ranges of variation are almost the same, and c1 of all three martens also do not differ morphologically. The p1 of M. vetus tends to be larger and less reduced (2.67 mm, 2.32-3.14 mm, n = 33) than in M. foina (2.21 mm, 1.79-2.56 mm, n = 114), and shows great similarity to p1 of M. martes (2.54 mm, 2.14-3.19 mm, n = 129).
The p1 of Martes vetus is not only larger, but also more robust, with a B/L index of 73.8 (66.2-80.2, n = 33), while this ratio is 72.9 (64.9-84.6, n = 129) in M. martes and 54.1 (53.2-63.9, n = 114) in M. foina. The p1 of the DA2 marten is relatively small (L = 2.27 mm), but rather wide (B/L = 67.8), and the robustness index falls into the range of variability of M. vetus. Metrically, the p2, p3, and p4 of all three martens are similar, only M. foina tends to show slightly higher values. Morphologically, those lower premolars showed some tendency to variability, although its range is not so wide. The p2 and p4 of M. vetus are narrower than that of M. martes and M. foina. In M. vetus, the B/L index of p2 is 50.7 (46.8-56.8, n = 29) and 45.2 (41.6-48.6, n = 38) in p4. Values of this ratio are 54.8 (48.9-61.1) for p2 and 53.2 (48.2-57.6) for p4 in M. martes (n = 129). In M. foina (n = 114), it is 54.9 (48.7-64.3) for p2 and 54.5 (48.8-61.6) for p4. The robustness index of p3 is almost the same for all three martens. Of the three p3 from DA2, two were assigned to morphotype A2, where a small distal additional cuspid is present on the distal ridge of the protoconid. The third p3 represents morphotype A1, where this cuspid is absent, and which is the dominant morphotype in M. foina (97%) and M. zibellina (82%) (Gimranov and Kosintsev, 2015). In M. martes, the most numerous is morphotype A2 (73%), but A1 also occurs relatively often (17%). Two p4 from DA2 were assigned to morphotype A3, where a small distal additional cuspid is present on the distal ridge of the protoconid, at lower level. This morphotype occurs in all three extant martens, but in low proportion: M. foina (3%), M. zibellina (9%), and M. martes (10%). Much more numerous in those species are morphotypes A1 (58% in M. foina and M. martes, and 40% in M. zibellina), without the additional cuspid, and A2 (33% in M. foina, 24% in M. martes and M. zibellina), where this cuspid is located high in relation to the protoconid (Gimranov and Kosintsev, 2015).
In the morphology of the lower dentition, most authors focused on m1. Dehm (1962) and Anderson (1970) highlighted the larger value of the mean size of m1. Additionally, Anderson (1970) and Wiszniowska (1989) mentioned that the m1 of Martes vetus has a proportionally short trigonid as in M. martes, and a small metaconid as in M. foina. Rabeder (1976) noted the lack of the so-called “paraconid edge”, a strong, median concavity situated between the disto-lingual margin of the paraconid and the mesio-lingual margin of the metaconid. When dealing with the DA2 material, it shows to be small marten, dimensionally smaller from other populations and species. The mean of L m1 in DA2 is 8.38 mm (7.97-8.78 mm, n = 5), which is clearly smaller than in extant martens, where L m1 in M. martes is 10.71 mm (10.08-11.38 mm, n = 63) in males and 9.64 mm (9.16-10.27 mm, n = 75) in females. Similarly, in M. foina, L m1 has higher values in males (L m1 = 10.64 mm, 10.04-11.34 mm, n = 68) compared to females (9.35 mm, 8.76-9.88 mm, n = 44). When comparing the DA2 material with two relatively numerous populations from Schernfeld and Żabia Cave, differences seemingly result from sexual dimorphism. In Schernfeld, M. vetus is larger (L m1 = 10.76 mm, 10.17-11.89 mm, n = 23 in males, and 9.44 mm, 8.74-9.89 mm, n = 17 in females) than in the Żabia Cave (L m1 = 9.87 mm, 9.57-10.52 mm, n = 7 in males, and 8.78 mm, 7.98-9.47 mm, n = 15 in females). The DA2 marten is relatively small, but its dimensions fall into the range of size variability of M. vetus females, and indicates a predominance of females at this locality. Only a single, incomplete m1 documents the presence of a larger male.
The general morphology of m1 in Martes vetus is typical for the genus Martes, with a proportionally broad and trenchant talonid, comparable in width with the trigonid. This is well illustrated by the talonid (B ta) to trigonid breadth (B tr) index. The mean value of this ratio is 98.9 (94.1-103.1, n = 83) in M. vetus, 106.1 (103.1-116.9, n = 139) in M. martes, and 93.4 (87.5-103.1, n = 114) in M. foina. It has been also found that M. vetus has not only a moderately narrow talonid, but also a relatively short trigonid. The mean ratio of the talonid (L ta) to trigonid length (L tr) is 46.9 (36.2-58.1, n = 86) in M. vetus, while 56.8 (41.5-72.3, n = 139) in M. martes and 37.6 (26.7-47.5, n = 114) in M. foina. In both indexes (L ta/L tr and B ta/B tr), the m1 of M. vetus shows intermediate values between M. martes and M. foina. The same was obtained for the material from DA2, where the mean of L ta/L tr is 52.6 (49.2-55.4, n = 4) and the mean of B ta/B tr is 98.8 (98.1-99.7, n = 5). Two m1 of DA2 were assigned to morphotype A1, were the distinct and robust hypoconid, as a sole cusp, occupies approximately half of the talonid basin. Two other m1 represents morphotype A2, where the small, but well-recognizable hypoconulid is adjacent to the robust hypoconid. Finally, the last tooth belongs to morphotype B2, where the hypoconulid is adjacent to the relatively small-sized hypoconid. The frequency of occurrence of particular morphotypes in extant martens is highly variable. There is, however, a noticeable difference between M. foina, in which group A morphotypes predominate (55% A1, 40% A2), M. martes, for which group B morphotypes are typical (24% B1, 67% B2), and M. zibellina, in which morphotypes B2 (44%) and C2 (45%) are the most common (Gimranov and Kosintsev, 2015). Other morphotypes occur only rarely.
In this aspect, the DA2 marten resembles the most Martes foina, because group A morphotypes also predominate. But the limited number of specimens should also be taken into account, especially considering that other morphotypes in the much more numerous populations of Schernfeld and Żabia Cave occurred also relatively often. Anderson (1970) noted some differences in the morphology of m2, being long and narrow in M. vetus, slightly broader than longer in M. martes, and nearly round in M. foina. Both preserved m2 from DA2 were assigned to morphotype A2, where the closely spaced protoconid and metaconid form a crest. This is also the dominant morphotype in M. foina (83%) and M. zibellina (96%), but occurs rarely (9%) in M. martes. For this species, morphotype A1 is typical (91%), where the protoconid and metaconid are separated from each other (Gimranov and Kosintsev, 2015). When comparing the B/L index of m2, both teeth from DA2 (89.2, 87.9-90.5, n = 2) corroborate well with the data obtained for M. vetus (92.7, 84.5-99.5, n = 46). The m2 of this marten tends to be narrower than in extant martens, in which the mean B/L ratio is higher, approximately 107.4 (94.5-112.4, n = 138) in M. martes and 99.7 (91.6-102.5, n = 112) in M. foina (83%).
In sum, the analyzed marten material from the DA2 locality both metrically and morphologically corroborates well with the respective dentognathic material of Martes vetus, which allows assigning it to this species. The comparison with two extant martens, M. martes and M. foina, showed a similar number of differences and similarities between these three species. Moreover, many of those features and indexes of M. vetus showed intermediate values between M. martes and M. foina. A concluded list of differences and similarities between those three martens is given below.
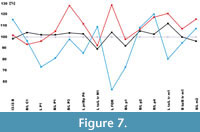 Comparison with Martes martes. Individuals of M. vetus from DA2 differ from M. martes in: (1) shorter and broader viscerocranium; (2) broader and less extended forward row of incisors; (3) strongly convex frontal profile in lateral view, with well-marked concavity in the middle part; (4) shorter, wider, and more strongly curved crown of C1; (5) narrower P2; (6) narrower P3 with weakly marked convex margin; (7) shorter and lower protocone and stronger lingual cingulum of P4; (8) less expanded trigon of M1 with more complicated microrelief on the chewing surface and less reduced metacone; (9) narrower spacing of mental foramens on the mandibular body; (10) slightly narrower c1; (11) narrower p2; (12) narrower p4 with an additional cuspid located low in relation to the protoconid; and (13) narrower m2 with closely spaced protoconid and metaconid that form a crest (Figure 7) (7) m1 with.
Comparison with Martes martes. Individuals of M. vetus from DA2 differ from M. martes in: (1) shorter and broader viscerocranium; (2) broader and less extended forward row of incisors; (3) strongly convex frontal profile in lateral view, with well-marked concavity in the middle part; (4) shorter, wider, and more strongly curved crown of C1; (5) narrower P2; (6) narrower P3 with weakly marked convex margin; (7) shorter and lower protocone and stronger lingual cingulum of P4; (8) less expanded trigon of M1 with more complicated microrelief on the chewing surface and less reduced metacone; (9) narrower spacing of mental foramens on the mandibular body; (10) slightly narrower c1; (11) narrower p2; (12) narrower p4 with an additional cuspid located low in relation to the protoconid; and (13) narrower m2 with closely spaced protoconid and metaconid that form a crest (Figure 7) (7) m1 with.
Individuals of Martes vetus from DA2 resemble M. martes in: (1) larger, less reduced, and robust P1; (2) elongated and moderately high mandibular body; (3) condyloid processes located slightly below or at the level of p4/m1; (4) shape of the mandibular ramus, where the mesial and distal edges of the coronoid process are approximately of the same length and form a not fully isosceles triangle; (5) longer and hooked angular process; (6) larger, less reduced, and robust p1; and (7) m1 with short trigonid (Figure 7).
Comparison with Martes foina. Individuals of M. vetus from DA2 differ from M. foina in: (1) less massive crown of C1; (2) smaller, narrower, and less reduced P1; (3) M1 with more complicated microrelief on the chewing surface and less reduced metacone; (4) wider spacing of mental foramens on the mandibular body; (5) elongated and moderately high mandibular body; (6) condyloid processes located slightly below or at the level of p4/m1; (7) shape of the mandibular ramus, where the mesial and distal edges of the coronoid process are approximately of the same length and form a not fully isosceles triangle (in M. foina, the mesial edge is more inclined than the distal one, which is almost vertical and thus considerably shorter); (8) longer and hooked angular process (short and straight in M. foina); (9) slightly narrower c1; (10) larger, less reduced, and broader p1; (11) narrower p2; (12) narrower p4 with an additional cuspid located usually low in relation to the protoconid; (13) shorter trigonid of m1; and (14) narrower m2 (Figure 7).
Individuals of Martes vetus from DA2 resemble M. foina in: (1) shorter and broader viscerocranium; (2) broader and less extended forward row of incisors; (3) strongly convex frontal profile in lateral view, with well-marked concavity in the middle part; (4) shorter and broader crown of C1; (5) narrow P2; (6) narrow P3 with weakly marked convex margin; (7) P4 with short and low protocone, which length is smaller than the distal breadth; (8) M1 with less expanded trigon and irregular occlusal outline; (9) strongly curved mandibular body, with notably marked curvature of the lower margin under m1; (10) m1 with a small metaconid and a talonid slightly narrower than the trigonid; and (11) morphology of m2, where the closely spaced protoconid and metaconid are connected by a distinct crest (Figure 7).
Possible occurrence of Martes zibellina at DA2. Rabeder (1976) described a right mandible body with the ramus fr. and present c1, p2-p3, and m1 (collection no. UWPI 2275/14/77) from DA2 as Martes cf. zibellina. He classified this specimen to this species according to its small size, lower crowns of c1, p2, and p3, and the small metaconid and broad talonid of m1. Among other features of this individual, he also noted the low mandibular body, the two mental foramens being located under p2 and p3, the moderately elongated and curved c1, the p1 alveolus directed distally, the low m1 without median concavity situated between the disto-lingual margin of the paraconid and the mesio-lingual margin of the metaconid, the trigonid slightly narrower than the talonid, the low hypoconid located on the trenchant talonid, and the minute hypoconulid. All these features are also characteristic for the morphology of M. vetus.
Rabeder (1976) did not discuss the differences between mandibles classified as Martes cf. zibellina and those of M. vetus. Only when describing the trigonid of the isolated m1 as M. cf. vetus, he concluded that this tooth cannot be assigned to M. zibellina, which differs in having smaller size and more strongly reduced metaconid of m1. He compared both specimens with the mandible (L m1 = 9.28 mm) from Sackdilling Cave, which was studied before by Brunner (1933), Anderson (1970), and Marciszak et al. (2021). More recently, Jiangzuo et al. (2021) agreed with Rabeder’s (1976) original classification as M. cf. zibellina. However, they also pointed out some differences, such as the relatively wide m1 talonid and the presence of an additional small cuspid in the talonid basin or in the entoconid ridge, which is not present in the analyzed specimen.
Rabeder (1976) also highlighted his doubts about the correctness of the identification of this particular specimen. Considering that Martes zibellina is unknown from the European fossil record and that its extant populations show a remarkable geographic variability, this determination is even less probable. Of course, we cannot fully reject the possible presence of other marten species at DA2, but, based on our current state of knowledge, it is highly unlikely. There are no clear differences between M. zibellina and M. martes in the structure of the skull and teeth. As it was already pointed out above, the morphology of M. vetus showed intermediate values by many features and indexes between M. martes and M. foina, which makes unambiguous classification extremely difficult or even impossible. Additionally, the presence of small specimens, metrically and morphologically indistinguishable from the DA2 specimen, was also reported from the Schernfeld and Żabia Cave populations. All of them were, after detailed and careful revision, identified as females of M. vetus.
DISCUSSION
Contrary to a widespread view (Kurtén, 1968; Kaiser, 1999), there is no evidence of survival of Martes vetus beyond to the mid-Middle or even late Middle Pleistocene of Europe. All mentioned records, especially those post-Elsterian (MIS 12) and later, are highly uncertain and based on fossil materials of doubtful taxonomic value. Martes vetus is regarded as a typical Early and early Middle Pleistocene (2.2-0.6 Mya) component of the fauna (Kurtén, 1968; Anderson, 1970; Wiszniowska, 1989; Kot et al., 2022; Marciszak et al., 2021, 2024). Marten remains from the mid-Middle Pleistocene are most often described as displaying intermediate features and are classified differently. The main issue with those remains is that they are fragmentary, which does not allow for comparison with materials from other sites. Therefore, their taxonomic assignment remains unresolved. Many previous authors, including Miller (1912), Kurtén (1965, 1968), and Anderson (1970), tried to single out specific metric and morphological characters to distinguish the species M. martes, M. foina, and M. vetus. However, due to those attempts, there is a large overlap between their ranges of variation. As a result, it is difficult to make a specific attribution based on isolated teeth alone, and when species-specific characters are not observable in poorly preserved materials recovered from mid-Middle Pleistocene European sites, such as Cerè Cave (Ghezzo et al., 2014), it is preferred only to attribute such fragments to the genus Martes.
Among the European sites dated to the mid-Middle Pleistocene (MIS 16-12), there many examples. One of the most fascinating and long-lasting examples is the Spanish site Sima de los Huesos (MIS 13-12). García et al. (1997, 2023), based mostly on the proportions of P4 and M1, identified the marten fossils unearthed at this site as Martes sp. Later, however, after a detailed morphometric comparison, García (2003) classified those remains as Martes cf. martes, which was followed by later authors (García and Arsuaga, 2011; García et al., 2023). She highlighted some features, including the elongated protocone of P4, which length exceeds the distal breadth of P4, the large and broad M1, the distinct transition between the trigon and talon marked by two concavities, especially the distal one, the moderately developed metacone and protocone, the considerably expanded and wide trigon, and the m1 lacking the median concavity between the disto-lingual margin of the paraconid and the mesio-lingual margin of the metaconid. Simultaneously, all those authors (García, 2003; García and Arsuaga, 2011; García et al., 2023) highlighted the intermediate features and proof for the evolutionary lineage of M. martes → M. vetus. Part of a right mandibular body of a marten was illustrated and mentioned from the West Runton Freshwater Bed (MIS 19; England) (Stuart and Lister, 2010). Originally classified as Martes sylvatica (Newton, 1880), it was soon re-identified as M. martes (Newton, 1880, 1882, 1891; Reynolds, 1912; Kurtén, 1968; Stuart, 1974, 1975, 1981, 1982, 1988, 1992, 1996). It was never, however, described in detail, and even a provisional look at the relatively narrowly spaced mental foramens or the weak metaconid of m1 allows this determination to be seriously doubted. Similarly, Kaiser (1999) mentioned the presence of M. cf. vetus at the German locality Breitscheid-Erdbach 1 Cave, dated provisionally to the Middle Pleistocene. Jánossy (1969) briefly described a complete humerus (60 mm long) from the Hungarian site Tarkő (MIS 15-13) and a few postcranial remains from Uppony 5 (MIS 11). In both cases, it was impossible to exactly determine the marten species. He noted, however, the greatest similarity of the humerus from Tarkő to that of M. martes by its gracile structure. Ghezzo et al. (2014) described in detail a distal fragment of a left mandible from Ceré Cave (MIS 15-13) as Martes sp.
At least MIS 12 is the time of the first appearance of Martes martes. The earliest, undoubted occurrence of the species is known from Polish site Tunel Wielki Cave (Kot et al., 2022). A faunal turnover took place in the cold period of MIS 12. It was the time when tundra species expanded to southern and south-western Europe, entering the newly formed tundra-steppe ecosystem. At the same time, Asian steppe species dispersed to the north and north-west. The changes led to the formation of the earliest pan-Eurasian Mammoth fauna c. 460 kya, which dominated the late Middle Pleistocene and Late Pleistocene mammal assemblages (Kahlke, 2014; Kot et al., 2022). From the same horizon came also remains of two other, cold adapted mustelid species, Gulo gulo (Linnaeus, 1758) and Mustela nivalis Linnaeus, 1758. Martes martes appeared after the Mimomys-Arvicola transition (Koenigswald and Heinrich, 1999; Marciszak, 2012; Marciszak et al., 2021). The mid-Middle Pleistocene changes in mustelid paleoassemblages corroborate well with the evolution of rodents (Horáček and Ložek, 1988; Horáček et al., 2004; Horáček, 2008). It shows that the main rearrangement of rodent communities in Central Europe took place at the same time, between MIS 17 and 15. From this perspective, the evolution of these mustelids could be interpreted as a result of the selective pressure of the onset of environmental deterioration. In this model, the main selective pressure would be related to specialization in a different spectrum of prey and a life strategy associated with open grasslands (in case of Mustela palerminea (Petényi, 1864) → Mustela erminea Linnaeus, 1758, and Mustela praenivalis Kormos, 1934 → Mustela nivalis Linnaeus, 1758) and forests (in case of Martes vetus → Martes martes). As documented by Marciszak et al. (2021) and Kot et al. (2022), the earliest occurrence of M. martes in Europe is younger than MIS 16/15, and all of the earlier published records of this species are either a result of misidentification or based on material with no diagnostic characters.
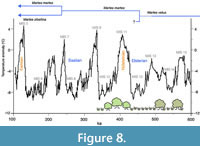 Similarly to Martes martes, M. zibellina also evolved from M. vetus during the Middle Pleistocene (0.6-0.55 Mya), probably in western Eurasia (Figure 8; Ishida et al. 2013). This hypothesis has been, however, poorly supported so far by fossil and mitochondrial data. Remains of M. zibellina older than the Late Pleistocene (~100 kya) are unknown. Additionally, the time of divergence from the most recent common ancestor was estimated for this species as c. 250 kya, with 95% confidence interval between 330-180 kya (Hope et al., 2013; Li et al., 2021). The extant M. zibellina is better adapted to cold environments than other Martes species (Davison et al. 2001). Accordingly, it was suggested that it could have first dispersed to eastern Eurasia during the glacials/stadials (Li et al., 2021). This is also supported by results obtained from the revision of the fossil material from the DA2 and DA4 localities, namely the absence of M. zibellina at both of these sites.
Similarly to Martes martes, M. zibellina also evolved from M. vetus during the Middle Pleistocene (0.6-0.55 Mya), probably in western Eurasia (Figure 8; Ishida et al. 2013). This hypothesis has been, however, poorly supported so far by fossil and mitochondrial data. Remains of M. zibellina older than the Late Pleistocene (~100 kya) are unknown. Additionally, the time of divergence from the most recent common ancestor was estimated for this species as c. 250 kya, with 95% confidence interval between 330-180 kya (Hope et al., 2013; Li et al., 2021). The extant M. zibellina is better adapted to cold environments than other Martes species (Davison et al. 2001). Accordingly, it was suggested that it could have first dispersed to eastern Eurasia during the glacials/stadials (Li et al., 2021). This is also supported by results obtained from the revision of the fossil material from the DA2 and DA4 localities, namely the absence of M. zibellina at both of these sites.
CONCLUSIONS
The Early Pleistocene marten fossils from the Austrian sites Deutsch Altenburg 2 and 4 showed a strong similarity to Martes vetus. As a result of a detailed revision, all marten remains have been assigned to this species, despite previous statements about the possible occurrence of another marten species, M. cf. zibellina. The supposed earlier presence of Martes zibellina is regarded as a result of misidentification due to sexual dimorphism. Thanks to their good state of preservation, the viscerocranium and mandibles from Deutsch Altenburg 2 and 4 are important materials in the European fossil record of M. vetus. A comparison with marterns from other European Early and early Middle Pleistocene sites showed the studied materials have intermediate features shared with M. martes, M. zibellina, and M. foina.
ACKNOWLEDGEMENTS
We are indebted to M. Wolsan and one anonymous reviewer for their useful comments and suggestions, which made meaningful improvements to the original manuscript. The research was financed by subsidies for the activities of the University of Wrocław, subsidy 2024 - research activities (501), MPK 2599280000 ZP, discipline 73 - biological sciences. It was also financed by an internal grant no. BPIDUB.4610.6.2021.KP.A. ‘The Middle Pleistocene Revolution - How the modern theriofauna of Eurasia was developed’, from the Faculty of Biological Sciences, University of Wrocław.
REFERENCES
Ambros, D.C. 2006. Morphologische und metrische Untersuchungen an Phalangen und Metapodien quartärer Musteliden unter besonderer Berücksichtigung der Unterscheidung von Baum- und Steinmarder (Martes martes (Linné 1758) und Martes foina (Erxleben 1777). Der Andere Verlag, Tönning, Lübeck, Marburg.
Ambros, D., Hilpert, B., and Kaulich, B. 2005. Das Windloch bei Sackdilling. Lage, Forschungsgeschichte, Geologie, Paläontologie und Archäologie, pp. 365-382. In Ambros, D., Gropp, C., Hilpert, B., and Kaulich, B. (eds.), Neue Forschungen zum Höhlenbären in Europa. Abhandlungen der Naturhistorischen Gesellschaft Nürnberg, 45.
Anderson, E. 1970. Quaternary evolution of the genus Martes (Carnivora, Mustelidae). Acta Zoologica Fennica, 130:1-132.
Anderson, E. 1994. Evolution, prehistoric distribution and systematics of Martes, pp. 13-25. In Buskirk, S.W., Harestad, A.S., Raphael, M.G., and Powell, R.A. (eds.), Martens, Sables and Fishers, Biology and Conservation. Cornell University Press, Ithaca.
Baskin, J.A. 1998. Mustelidae, pp. 152-173. In Janis, C.M., Scott, K.M., and Jacobs, L.L. (eds.), Evolution of Tertiary Mammals of North America. Volume 1, Terrestrial Carnivores, Ungulates, and Ungulate-like Mammals. Cambridge University Press, Cambridge.
Bowdich, T.E. 1821. An Analysis of the Natural Classifications of Mammalia for the Use of Students and Travelers. J. Smith, Paris.
Brunner, G. 1933. Eine präglaciale Fauna aus dem Windloch bei Sackdilling (Oberpfalz). Neues Jahrbuch für Mineralogie, Geologie und Paläontologie, 71B:303-328.
Davison, A., Birks, J.D.S., Brookes, R.C., Messenger, J.E., and Griffiths, H.I. 2001. Mitochondrial phylogeography and population history of pine martens Martes martes compared with polecats Mustela putorius. Molecular Ecology, 10:2479-2488.
https://doi.org/10.1046/j.1365-294X.2001.01381.x
Dehm, R. 1950. Die Nagetiere aus dem Mittel-Miozän (Burdigalium) von Wintershof-West bei Eichstätt in Bayern. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen, 91B:321-428.
Dehm, R. 1953. Die Raubtiere aus dem Mittel-Miozän (Burdigalium) von Wintershof-West bei Eichstätt in Bayern. Abhandlungen Bayerische Akademie der Wissenschaften, Neue Folge, 58:1-141.
Dehm, R. 1962. Altpleistozäne Säuger von Schernfeld bei Eichstätt in Bayern. Mitteilungen der Bayerischen Staatssammlung für Paläontologie und Historische Geologie, 2:17-61.
Ehrenberg, K. 1929. Zur Frage der systematischen und phylogenetischen Stellung der Bärenreste von Hundsheim und Deutsch-Altenburg in Niederösterreich. Palaeobiologica, 2:213-222.
Erxleben, J.C.P. 1777. Systema regni animalis per classes, ordines, genera, species, varietates cum synonymia et historia animalium. Classis I. Mammalia. Weigand, Leipzig.
Fischer de Waldheim, G. 1817. Adversaria zoologica. Mémoires de la Société Impériale des Naturalistes de Moscou, 5:357-472.
Fladerer, F.A. 1984. Das Vordergliedmaßenskelett von Hypolagus beremendensis und von Lepus sp. (Lagomorpha, Mammalia) aus dem Altpleistozän von Deutsch-Altenburg (Niederösterreich). Beiträge zur Paläontologie, 11:71-148.
Flower, W.H. 1869. On the value of the characters of the base of the cranium in the classification of the order Carnivora, and on the systematic position of Bassaris and other disputed forms. Proceedings of the Zoological Society of London, 37(1):4-37.
Flynn, J.J., Finarelli, J.A., Zehr, S., Hsu, J., and Nedbal, M.A. 2005. Molecular phylogeny of the carnivora (Mammalia), assessing the impact of increased sampling on resolving enigmatic relationships. Systematic Biology, 54(2):317-337.
https://doi.org/10.1080/10635150590923326
Forsyth-Major, C.I. 1902. Some jaws and teeth of Pliocene voles (Mimomys gen. nov.), from the Norwich Crag at Thorpe, and from the upper Val d’Arno. Proceedings of the Zoological Society of London, 1:102-107.
Frank, C. and Rabeder, G. 1997a. Deutsch-Altenburg, pp. 238-240. In Döppes, D. and Rabeder, G. (eds.), Pliozäne und pleistozäne Faunen Österreichs. Ein Katalog der wichtigsten Fossilfundstellen und ihrer Faunen. Mitteilungen der Kommission für Quartärforschung Österreichische Akademie der Wissenschaften, 10.
Frank, C. and Rabeder, G. 1997b. Deutsch-Altenburg 2-4-16-30, pp. 241-250. In Döppes, D. and Rabeder, G. (eds.), Pliozäne und pleistozäne Faunen Österreichs. Ein Katalog der wichtigsten Fossilfundstellen und ihrer Faunen. Mitteilungen der Kommission für Quartärforschung Österreichische Akademie der Wissenschaften, 10.
Freudenberg, W. 1914. Die Säugetiere des älteren Quartärs von Mitteleuropa mit besonderer Berücksichtigung der Fauna von Hundsheim und Deutschaltenburg in Niederösterreich nebst Bemerkungen über verwandte Formen anderer Fundorte. Geologische und Paläontologische Abhandlungen, Neue Folge, 12(4-5):455-671.
García, N.G. 2003. Osos y otros carnivoros de la Sierra de Atapuerca. Fundación Oso de Asturias, Oviedo.
García, N. and Arsuaga, J.L. 2011. The Sima de los Huesos (Burgos, northern Spain): palaeoenvironment and habitats of Homo heidelbergensis during the Middle Pleistocene. Quaternary Science Reviews, 30(11-12):1413-1419.
https://doi.org/10.1016/j.quascirev.2010.11.008
García, N., Arsuaga, J.L., and Torres, T. 1997. The carnivore remains from the Sima de los Huesos, Middle Pleistocene site (Sierra de Atapuerca, Spain). Journal of Human Evolution, 33(2-3):155-174.
https://doi.org/10.1006/jhev.1997.0154
García, N., Jimenez, I.J., Blázquez-Orta, R., and Arsuaga, J.L. 2023. Updates to the carnivore fauna from the Sima de los Huesos. The Anatomical Record.
https://doi.org/10.1002/ar.25199
Gasparik, M. and Pazonyi, P. 2018. The macromammal remains and revised faunal list of the Somssich Hill 2 locality (late Early Pleistocene, Hungary) and the Epivillafranchian faunal change. Fragmenta Palaeontologica Hungarica, 35:153-178.
https://doi.org/10.17111/FragmPalHung.2018.35.153
Ghezzo, E., Berté, D.F., and Sala, B. 2014. The revaluation of Galerian Canidae, Felidae and Mustelidae of the Cerè Cave (Verona, Northeastern Italy). Quaternary International, 339-340:76-89.
https://doi.org/10.1016/j.quaint.2012.12.031
Gimranov, D. and Kosintsev, P. 2015. Differentiation of three Martes species (M. martes, M. zibellina, M. foina) by tooth morphotypes. Comptes Rendus Palevol, 14(8):647-656.
https://doi.org/10.1016/j.crpv.2015.06.007
Ginsburg, L. 1961. La faune des carnivores miocènes de Sansan. Mémoires du Muséum national d'Histoire naturelle, Série C, 9:1-190.
Ginsburg, L. 1999. Order Carnivora, pp. 109-148. In Rössner, G.E. and Heissig, K. (eds.), Land Mammals of Europe. Pfeil, München.
Ginsburg, L. and Morales, J. 1992. Contribution à la connaissance des Mustélidés (Carnivora, Mammalia) du Miocène d’Europe Trochictis et Ischyrictis, genres affines et genres nouveaux. Comptes Rendus de l’Académie des Sciences de Paris, 315:111-116.
Heller, F. 1930. Eine Forest-Bed-Fauna aus der Sackdillinger Höhle (Oberpfalz). Neues Jahrbuch für Geologie und Paläontologie, 63B:247-298.
Heller, F. 1933. Ein Nachtrag zur Forest Bed Fauna aus der Sackdillinger Höhle (Oberpfalz). Centralblatt für Mineralogie, Geologie und Paläontologie, Abteilung B, 1933:60-68.
Heller, F. 1936. Eine Forest Bed Fauna aus der Schwäbischen Alb. Sitzungsberichte der Heidelberger Akademie der Wissenschaften, Mathematisch Naturwissenschaftliche Klasse, 2: 1-29.
Hinton, M.A.C. 1910. A preliminary account of the British fossil voles and lemmings; with some remarks on the Pleistocene climate and geography. Proceedings of the Geologists Association, 21:489-507.
Hope, A.G., Takebayashi, N., Galbreath, K.E., Talbot, S.L., and Cook, J.A. 2013. Temporal, spatial and ecological dynamics of speciation among amphi-Beringian small mammals. Journal of Biogeography, 40(3):415-429.
https://doi.org/10.1111/jbi.12056
Horáček, I. 2008. Jak se zviřátka rozkmotřila a co z toho pošlo, pp. 63-76. In Pokorný, P. and Bárta, M. (eds.), Něco překrásného se končí - Kolapsy v přírodě a společnosti. Dokořán, Praha.
Horáček, I. and Ložek, V. 1988. Palaeozoology and the mid-European Quaternary past: scope of the approach and selected results. Rozpravy Ceskoslovenkse Akademe Ved, Rada Matematickych a Prirodnich Ved, 98:1-102.
Horáček, I., Ložek, V., and Fejfar, O. 2004. The Middle Pleistocene revolution and the Central European vertebrate and molluscs fossil record. Terra Nostra, Schriften der Alfred-Wegener-Stiftung, 2004:119-121.
Hughes, S.S. 2012. Synthesis of Martes evolutionary history, pp. 3-22. In Aubry, K.B., Zieliński, W.J., Raphael, M.G., Proulx, G., and Buskirk, S.W. (eds.), Biology and Conservation of Martens, Sables, and Fishers. Cornell University Press, Ithaca.
Ishida, K., Sato, J.J., Kinoshita, G., Hosoda, T., Kryukov, A.P., and Suzuki, H. 2013. Evolutionary history of the sable (Martes zibellina brachyura) on Hokkaido inferred from mitochondrial Cytb and nuclear Mc1r and Tcf25 gene sequences. Acta Theriologica, 58:13-24.
https://doi.org/10.1007/s13364-012-0103-z
Jánossy, D. 1969. Stratigraphische Auswertung der europäischen mittelpleistozänen Wirbeltierfauna. Teil II. Berichte der deutschen Geselschaft für geologische Wissenschaft, Abteilung A, Geologie und Paläontologie, 5:573-643.
Jánossy, D. 1981. Die altpleistozänen Vogel-Faunen von Deutsch-Altenburg 2 und 4 (Niederösterreieh). Beiträge zur Paläontologie und Geologie Österreich, 8:375-391.
Jiangzuo, Q., Gimranov, D., Liu, J., Liu, S., Jin, Ch., and Liu, J. 2021. A new fossil marten from Jinyuan Cave, north-eastern China reveals the origin of the Holarctic marten group. Quaternary International, 531:47-58.
https://doi.org/10.1016/j.quaint.2020.10.064
Kahlke, R.D. 2014. The origin of Eurasian mammoth faunas (Mammuthus-Coelodonta Faunal Complex). Quaternary Science Reviews, 96:32-49.
https://doi.org/10.1016/j.quascirev.2013.01.012
Kaiser, T.M. 1999. New Pleistocene vertebrate assemblages in the Breitscheid-Erdbach Cave system (Iberg limestone, Dill Basin, Germany). Journal of Cave and Karst Studies, 61(3):145-149.
Koenigswald, W.V. and Heinrich, W.-D. 1999. Mittelpleistozäne Säugetierfaunen aus Mitteleuropa - der Versuch einer biostratigraphischen Zuordnung. Kaupia, 9:53-112.
Koepfli, K.-P., Deere, K.A., Slater, G.J., Begg, C., Begg, K., Grassman, L., Lucherini, M., Veron, G., and Wayne, R.K. 2008. Multigene phylogeny of the Mustelidae, resolving relationships, tempo and biogeographic history of a mammalian adaptive radiation. BMC Biology, 6(1):10.
https://doi.org/10.1186/1741-7007-6-10
Kormos, T. 1930. Diagnosen neuer Säugetiere aus der oberpliozänen Fauna des Somlyóberges bei Püspökfürdő. Annales Musei Historico-Naturalis Hungarici, 27:237-246.
Kormos, T. 1932. Neue Wühlmause aus dem Oberpliozän von Püspökfürdo. Neues Jahrbuch für Mineralogie, Geologie und Paläontologie, 69:323-346.
Kormos, T. 1934. Neue und wenig bekannte Musteliden aus dem ungarischen Oberpliozän. Folia Zoologica Hydrobiologica, 5(2):129-158.
Kot, M., Berto, C., Krajcarz, M.T., Moskal-del Hoyo, M., Gryczewska, N., Szymanek, M., Marciszak, A., Stefaniak, K., Zarzecka-Szubińska, K., Lipecki, G., Wertz, K., and Madeyska, T. 2022. Frontiers of the Lower Palaeolithic expansion in Europe: Tunel Wielki Cave (Poland). Scientific Reports, 12:16355.
https://doi.org/10.1038/s41598-022-20582-0
Kretzoi, M. 1942. Präokkupierte und durch Ältere zu ersetzende Säugetiernamen. Földtani Közlöny, 72:345-349.
Kretzoi, M. 1943. Kochitis centenii n.g. n.sp., ein alterümlicher Creodonte aus dem Oberoligozän Siebenbürgens. Földtani Közlöny, 73:268-271.
Kretzoi, M. 1945. Bemerkungen über das Raubtiersystem. Annales Historico-Naturales Musei Nationalis Hungarici, 38:59-83.
Kretzoi, M. 1954. Bericht über die Calabrische (Villafranchische) Fauna von Kisláng, Kom. Feyér. Jahresbericht der Ungarisches Geologischen Anstalt, 1953:212-264.
Kretzoi, M. 1958. New names for arvicolid homonyms. Annales Musei historico-naturalis Hungarici, 50:55-58.
Kurtén, B. 1965. The Carnivora of the Palestine caves. Acta Zoologica Fennica, 107:1-74.
Kurtén, B. 1968. Pleistocene Mammals of Europe. Weidenfeld and Nicolson, London.
Linnaeus, C. 1758. Systema naturae per regna tria naturae: secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis. Holmiae, Impensis Direct. Laurentii Salvii.
Li, B., Wolsan, M., Wu, D., Zhang, W., Xu, Y., and Zeng, Z. 2014. Mitochondrial genomes reveal the pattern and timing of marten (Martes), wolverine (Gulo), and fisher (Pekania) diversification. Molecular Phylogenetics and Evolution, 80:156-164.
https://doi.org/10.1016/j.ympev.2014.08.002
Li, B., Lu, J., Monakhov, V. Kang, H., Xu, Y., An, B., Ghani, M.U., Li, M., Peng, W., and Ma, X. 2021. Phylogeography of subspecies of the sable (Martes zibellina L.) based on mitochondrial genomes: implications for evolutionary history. Mammalian Biology, 101:105-120.
https://doi.org/10.1007/s42991-020-00092-0
Mais, K. 1971. Entdeckung einer Knochenspalte im Pfaffenberg bei Bad Deutsch-Altenburg. Höhlenkundliche Mitteilungen, 1971:43-44.
Mais, K. 1973. Das Karstgebiet Pfaffenberg bei Bad Deutsch-Altenburg (Niederösterreich) - ein vorläufiger Überblick. Die Höhle, 24(1):1-8.
Mais, K. 1978. Neue Ergebnisse speläologischer Untersuchungen in den Hainburger Bergen (NÖ.) und ihre wissenschaftliche Bedeutung. International Journal of Speleology, 10(2):167-178.
Mais, K. and Rabeder, G. 1977a. Eine pliozäne Höhlenfüllung im Pfaffenberg bei Bad Deutsch-Altenburg (Niederösterreich). Die Höhle, 28(1):1-7.
Mais, K. and Rabeder, G. 1977b. Eine weitere pliozäne Höhlenfauna aus dem Steinbruch Hollitzer bei Bad Deutsch-Altenburg (Niederösterreich). Die Höhle, 28(3):84-86.
Mais, K. and Rabeder, G. 1979. Das Karstgebiet der Hainburger Berge. Veröffentlichungen aus dem Naturhistorischen Museum Wien, Neue Funde, 17:51-63.
Mais, K. and Rabeder, G. 1984. Das große Höhlensystem im Pfaffenberg bei Bad Deutsch-Altenburg (NÖ) und seine fossilen Faunen. Die Höhle, 35(3-4):213-230.
Mais, K. and Rabeder, G. 1997. Eine pliozäne Höhlenfüllung im Pfaffenberg bei Deutsch-Altenburg (Niederösterreich). Die Höhle, 28(1):1-7.
Malyarchuk, B.A., Derenko, M.V., and Denisova, G.A. 2015. Mitochondrial genome variability in the wolverine (Gulo gulo). Russian Journal of Genetics, 51(11):1113-1118.
https://doi.org/10.1134/S1022795415090069
Marciszak, A. 2012. Ssaki łasicowate (Mustelidae, Carnivora, Mammalia) z plejstocenu Polski. PhD Thesis. University of Wrocław, Wrocław.
Marciszak, A., Ambros, D., and Hilpert, B. 2021. Mustelids from Sackdilling Cave (Bavaria, Germany) and their biostratigraphic significance. Geobios, 68:83-107.
https://doi.org/10.1016/j.geobios.2021.04.004
Marciszak, A., Lipecki, G., and Spassov, N. 2024. Martes wenzensis Stach, 1959 within the early history of the genus Martes Pinel, 1792. Acta Geologica Polonica, 74(1).
https://doi.org/10.24425/agp.2023.148027
Matthew, W.D. 1924. Third contribution to the Snake Creek Fauna. Bulletin of the American Museum of Natural History, 50:59-210.
Méhely, L. 1914. Fibrinae Hungariae: Der tertiären andquartären wurzelzänigen Wühlmäuse Ungarns. Annales Musei historico-naturalis Hungarici, 12:155-243.
Miller, G.S. 1912. Catalogue of the Mammals of Western Europe (Europe Exclusive of Russia) in the Collection of the British Museum. BMNH, London.
Montoya, P., Morales, J., and Abella, J. 2011. Musteloidea (Carnivora, Mammalia) from the Late Miocene of Venta del Moro (Valencia, Spain). Estudios Geológicos, 67(2):193-206.
https://doi.org/10.3989/egeol.40576.183
Nagel, D. and Rabeder, G. 1997. Revision der mittelpleistozänen Großsäugerfauna von Deutsch-Altenburg 1. Wissenschaftliche Mitteilungen Niederösterreichisches Landesmuseum, 10:231-249.
Newton, E.T. 1880. Notes on the vertebrata of the pre-glacial Forest Bed series of the East of England. Part II - Carnivora. On the occurrence of the glutton, Gulo luscus, Linn., in the Forest Bed of Norfolk. Geological Magazine, 9(7):1-4.
Newton, E.T. 1882. The vertebrata of the Forest Bed Series of Norfolk and Suffolk. Memoirs of the Geological Survey of Great Britain: England and Wales, H.M. Stationery Office, London.
Newton, E.T. 1891. The vertebrata of the Pliocene deposits of Britain. Memoirs of the Geological Survey of Great Britain: England and Wales, H.M. Stationery Office, London.
Pallas, P.S. 1780. Spicilegia zoologica: quibus novae imprimis et obscurae animalium species iconibus, descriptionibus atque commentariis illustrantur, fasciule 14. Berolini, Prostant apud Gottl. August. Lange.
Petényi, S.J. 1864. Hátrahagyott Munkái. Magyar Tudományos Akadémia jegyzőkönyvei, 130:44-50.
Peterson, O.A. 1910. Description of new carnivores from the Miocene of western Nebraska. Memoirs of the Carnegie Museum, 4(5):205-278.
Petter, G. 1963. Contribution a l’étude des mustélidés des Bassins Néogènes de Vallès-Pénédès et de Calatayud-Teruel (Espagne orientale). Mémoires de la Société Géologique de France, nouvelle série, 42(2):1-44.
Pfeiffer-Deml, T. 2016. Deer from the Pliocene site of Bad Deutsch-Altenburg 26 (Lower Austria, Leithagebirge): Conclusions based on skeletal morphology. Annalen des Naturhistorischen Museums in Wien, Serie A, 118:133-173.
Pinel, P. 1792. Recherches sur une nouvelle classification des quadrupèdes, fondée sur la structure méchanique des parties osseuses qui servent à l'articulation de la mâchoire inférieure. Actes de la Société d’Histoire Naturelle de Paris, 1:50-60.
Rabeder, G. 1972a. Die Insectivoren und Chiropteren (Mammalia) aus dem Altpleistozän von Hundsheim (Niederösterreich). Annalen des Naturhistorischen Museums in Wien, 76:375-474.
Rabeder, G. 1972b. Eine fossile Höhlenfauna aus dem Steinbruch Hollitzer bei Bad Deutsch-Altenburg (NÖ.). Die Höhle, 23:89-95.
Rabeder, G. 1972c. Ein neuer Soricide (Insectivora) aus dem Alt-Pleistozän von Deutsch-Altenburg 2 (Niederösterreich). Neues Jahrbuch für Geologie und Paläontologie Monatshefte, 1972:625-642.
Rabeder, G. 1973a. Weitere Grabungsergebnisse von der altpleistozänen Wirbeltierfundstelle Deutsch-Altenburg 2. Die Höhle, 24:8-15.
Rabeder, G. 1973b. Ein neuer Mustelide (Carnivora) aus dem Altpleistozän von Deutsch-Altenburg 2. Neues Jahrbuch für Geologie und Paläontologie Monatshefte, 1973:674-689.
Rabeder, G. 1974a. Eine fossile Höhlenfauna aus dem Steinbruch Hollitzer bei Bad Deutsch-Altenburg (NÖ). Die Höhle, 25:89-95.
Rabeder, G. 1974b. Plecotus und Barbastella (Chiroptera) aus dem Altpleistozän von Deutsch-Altenburg 2. Naturkunde-Jahrbuch Stadt Linz, 1973:159-184.
Rabeder, G. 1974c. Fossile Schlangenreste aus den Höhlenfüllungen des Pfaffenberges bei Bad Deutsch-Altenburg (NÖ). Die Höhle, 25(4):145-149.
Rabeder, G. 1976. Die Carnivoren (Mammalia) aus dem Altpleistozän von Deutsch-Altenburg 2. Mit Beiträgen zur Systematik einiger Musteliden und Caniden. Beiträge zur Paläontologie Österreich, 1:1-119.
Rabeder, G. 1978. Das fossilführende Pleistozän-Profil im Höhlensystem „Deutsch-Altenburg 2-4-16" im Pfaffenberg bei Bad Deutsch-Altenburg (NÖ.), pp. 449-458. In Nagl, H. (ed.), Festschrift z. 60. Geburtstag von Julius Fink. Beiträge zur Quartär- und Landschaftsforschung.
Rabeder, G. 1981. Die Arvicoliden (Rodentier, Mammalia) aus dem Pliozäen und dem ältesten Pleistozän von Niederösterreich. Beiträge zur Paläontologie Österreich, 8:1-373.
Rabeder, G. 1982. Die Gattung Dimylosorex (Insectivora, Mammalia) aus dem Altpleistozän von Deutsch-Altenburg (Niederösterreich). Beiträge zur Paläontologie Österreich, 9:233-251.
Rabeder, G. and Withalm, G. 2006. Brown bear remains (Ursidae, Mammalia) from Early Pleistocene cave fillings of Deutsch-Altenburg (Lower Austria). 12th Internat. Cave Bear Symposium, Aridea, Macedonia, Greece 2006, abstract book: 47, Aridea/Loutrá.
Rabeder, G., Pacher, M., and Withalm, G. 2010. Early Pleistocene bear remains from Deutsch-Altenburg (Lower Austria). Mitteilungen der Kommission für Quartärforschung Österreichische Akademie der Wissenschaften, 17:1-135.
Reynolds, S.H. 1912. A Monograph of the British Pleistocene Mammalia. Vol. 2, part 4. The Mustelidae. Palaeontographical Society, London.
Samuels, J.X. and Cavin, J. 2013. The earliest known fisher (Mustelidae), a new species from the Rattlesnake Formation of Oregon. Journal of Vertebrate Palaeontology, 33(2):448-454.
https://doi.org/10.1080/02724634.2013.722155
Samuels, J.X., Bredehoeft, K.E., and Wallace, S.C. 2018. A new species of Gulo from the Early Pliocene Gray fossil site (Eastern United States); rethinking the evolution of wolverines. PeerJ, 6:e4648.
https://doi.org/10.7717/peerj.4648
Sapper, N.R. 1997. Hufeisennasen (Rhinolophiden - Chiroptera, Mammalia) im pleistozänen Höhlensystem von Bad Deutsch-Altenburg. Wissenschaftliche Mitteilungen Niederösterreichisches Landesmuseum, 10:391-441.
Sato, J.J., Hosoda, T., Wolsan, M., Tsuchiya, K., Yamamoto, M., and Suzuki, H. 2003. Phylogenetic relationships and divergence times among mustelids (Mammalia, Carnivora) based on nucleotide sequences of the nuclear interphotoreceptor retinoid binding protein and mitochondrial cytochrome b genes. Zoological Science, 20(2):243-264.
https://doi.org/10.2108/zsj.20.243
Sato, J.J., Wolsan, M., Prevosti, F.J., D’Elía, G., Begg, C., Begg, K., Hosoda, T., Campbell, K.L., and Suzuki, H. 2012. Evolutionary and biogeographic history of weasel-like carnivorans (Musteloidea). Molecular Phylogenetics and Evolution, 63(3):745-757.
https://doi.org/10.1016/j.ympev.2012.02.025
Stach, J. 1959. On some Mustelinae from the Pliocene bone breccia of Węże. Acta Palaeontologica Polonica, 4(2):101-117.
Stuart, A.J. 1974. Pleistocene history of the British vertebrate fauna. Biological Reviews of the Cambridge Philosophical Society, 49(2):225-266.
https://doi.org/10.1111/j.1469-185X.1974.tb01574.x
Stuart, A.J. 1975. The vertebrate fauna of the type Cromerian. Boreas, 4:63-76.
Stuart, A.J. 1981. A comparison of the Middle Pleistocene mammalian faunas of Voigtstedt (Thuringia, German Democratic Republic) and West Runton (Norfolk, England). Quartärpaläontologie, 4:155-163.
Stuart, A.J. 1982. Pleistocene Vertebrates in the British Isles London. Longman Financial Service, London.
Stuart, A.J. 1988. The vertebrate faunas from West Runton (Crag and Freshwater Bed), pp. 152-157. In Gibbard, P.L. and Zalasiewicz, J.A. (eds.), Pliocene-Middle Pleistocene of East Anglia. Field Guide. Quaternary Research Association, Cambridge.
Stuart, A.J. 1992. The Pleistocene vertebrate faunas of West Runton, Norfolk, England. Cranium, 9(2):77-84.
Stuart, A.J. 1996. Vertebrate faunas from the early Middle Pleistocene of East Anglia, pp. 9-24. In Turner, C. (ed.), The Early Middle Pleistocene in Europe. A.A. Balkema, Rotterdam.
Stuart, A.J. and Lister, A.M. 2010. The West Runton Freshwater Bed and the West Runton Mammoth: Summary and conclusions. Quaternary International, 228:241-248.
https://doi.org/10.1016/j.quaint.2010.07.033
Tedford, R.H. 1976. Relationships of pinnipeds to other Carnivores (Mammalia). Systematic Zoology, 25:363-374.
Toula, F. 1902. Das Nashorn von Hundsheim: Rhinoceros (Ceratorhinus Osborn) hundsheimensis nov. form. mit Ausfuhrungen uber die Verhaltnisse von elf Schadeln von Rhinoceros (Ceratorhinus) sumatrensis. Abhandlungen der Kaiserischen Königlichen Geologischen Reichsanstalt, 19(1):1-92.
Toula, F. 1906. Das gebiss und Reste der Nasenbeine von Rhinoceros (Ceratorhinus Osborn) hundsheimensis. Abhandlungen der Kaiserischen Königlichen Geologischen Reichsanstalt, 20(2):1-38.
Valenciano, A. 2017. Mofetas, martas, tejones y rateles gigantes del Cerro de los Batallones, pp. 322-336. In Morales, J. (ed.), La Colina de los Tigres Dientes de Sable. Los Yacimientos Miocenos del Cerro de los Batallones (Torrejón de Velasco, Comunidad de Madrid). Museo Arqueológico Regional, Madrid.
Valenciano, A., Pérez-Ramos, A., Abella, J., and Morales, J. 2020. A new hypercarnivorous mustelid (Mammalia, Carnivora, Mustelidae) from Batallones, late Miocene (MN10), Torrejón de Velasco, Madrid, Spain. Geodiversitas, 42:103-121.
https://doi.org/10.5252/geodiversitas2020v42a8
Valenciano, A., Morales, J., Azanza, B., and Demiguel, D. 2021. Aragonictis araid, gen. et sp. nov., a small-sized hypercarnivore (Carnivora, Mustelidae) from the upper middle Miocene of the Iberian Peninsula (Spain). Journal of Vertebrate Palaeontology, 41(5):e2005615.
https://doi.org/10.1080/02724634.2021.2005615
Wang, X., Tseng, Z.J., and Takeuchi, G.T. 2012. Zoogeography, molecular divergence, and the fossil record - the case of an extinct fisher, Pekania palaeosinensis (Mustelidae, Mammalia), from the Late Miocene Baogeda Ula Formation, Inner Mongolia. Vertebrata PalAsiatica, 50(3):293-307.
Wiszniowska, T. 1989. Middle Pleistocene Carnivora (Mammalia) from Kozi Grzbiet in the Świętokrzyskie Mts, Poland. Acta Zoologica Cracoviensia, 32(14):589-630.
Wolsan, M. 1988. Morphological variations of the first upper molar in the genus Martes (Carnivora, Mustelidae). Mémoires du Muséum National d’Histoire Naturelle Série C, 53:241-254.
Wolsan M. 1989a. Dental polymorphism in the genus Martes (Carnivora: Mustelidae) and its evolutionary significance. Acta Theriologica, 34(40): 545-593.
Wolsan, M. 1989b. Drapieżne - Carnivora, pp. 177-197. In Kowalski, K. (ed.), Historia i ewolucja lądowej fauny Polski. Folia Quaternaria, 59-60.
Wolsan, M. 1993. Phylogeny and classification of early European Mustelida (Mammalia, Carnivora). Acta Theriologica, 38(4):345-384.
https://doi.org/10.4098/AT.arch.93-29
Wolsan, M. and Sato, J.J. 2010. Effects of data incompleteness on the relative performance of parsimony and Bayesian approaches in a supermatrix phylogenetic reconstruction of Mustelidae and Procyonidae (Carnivora). Cladistics, 26(2):168-194.
https://doi.org/10.1111/j.1096-0031.2009.00281.x
Wolsan M., Ruprecht A.L., and Buchalczyk, T. 1985. Variation and asymmetry in the dentition of the pine and stone martens (Martes martes and M. foina) from Poland. Acta Theriologica, 30(3):79-114.
Zdansky, O. 1924. Jungtertiäre carnivoren Chinas. Palaeontologica Sinica, 2:1-149.
Zhu, S., Gao, Y., Liu, H., Zhang, S., Bai, X., and Zhang, M. 2016. Phylogenetic relationship of wolverine Gulo gulo in Mustelidae revealed by complete mitochondrial genome. Mitochondrial DNA Part A, 27(4) 2937-2938.
https://doi.org/10.3109/19401736.2015.1060449
