

 New species and evolution of the foraminiferal family Janischewskinidae in the middle–upper Mississippian of South China
New species and evolution of the foraminiferal family Janischewskinidae in the middle–upper Mississippian of South China
Article number: 26.1.a2
https://doi.org/10.26879/1238
Copyright Paleontological Society, February 2023
Author biographies
Plain-language and multi-lingual abstracts
PDF version
Submission: 16 August 2022. Acceptance: 2 December 2022.
ABSTRACT
Rich foraminiferal assemblages from the Bama Platform in South China allow clarifying the systematics, composition, and biostratigraphy of the family Janischewskinidae. The family is comprised of four genera, Janischewskina, Cribrospira, Bibradya, and Parajanischewskina. The genera Rhodesinella and Groessensella are considered to be synonyms of Cribrospira and Bibradya, respectively, and some specimens of the genus Ugurus have been included under the genus Bibradya. Seven new species are described in this study: Cribrospira evoluta, C. paradenticulata, Bibradya primitiva, B. densicamerata, B. maxima, B. subita, and Parajanischewskina nautiliformis. The biostratigraphy of the Janischewskinidae in South China resembles that in the Russian Platform, i.e., primitive Cribrospira are first recorded from the Tulian, primitive Bibradya from the Aleksinian, evolved Cribrospira (e.g., C. panderi) and large Janischewskina species from the Mikhailovian, Parajanischewskina and Bibradya maxima from the Venevian, and some species from the base of the Serpukhovian (Tarusian), such as Janischewskina delicata, J. gibshmanae, Bibradya subita, and Parajanischewskina nautiliformis. On the other hand, Janischewskina adtarusia is only documented from the late Serpukhovian strata (Zapaltyubian).
Chao Liu. Institute of Resources and Environment, Henan Polytechnic University, Shiji Road 2001, 454003 Jiaozuo, China. liuchao661030@126.com
Daniel Vachard. 1 rue des Tilleuls, 59152 Gruson, France. daniel.vachard@free.fr
Pedro Cózar. Instituto de Geociencias (CSIC-UCM), c/ Severo Ochoa 7, 28040 Madrid, Spain. p.cozar@igeo.ucm-csic.es
Ismael Coronado. Facultad de Ciencias Biológicas y Ambientales, Universidad de León, Campus de Vegazana s/n, 24071-León, Spain. icorv@unileon.es
Keywords: foraminifers; new species; late Viséan; Serpukhovian; biostratigraphy
Final citation: Liu, Chao, Vachard, Daniel, Cózar, Pedro, and Coronado, Ismael. 2023. New species and evolution of the foraminiferal family Janischewskinidae in the middle–upper Mississippian of South China. Palaeontologia Electronica, 26(1):a2. https://doi.org/10.26879/1238
palaeo-electronica.org/content/2023/3737-janischewskinidae-of-china
Copyright: February 2023 Paleontological Society.
This is an open access article distributed under the terms of Attribution-NonCommercial-ShareAlike 4.0 International (CC BY-NC-SA 4.0), which permits users to copy and redistribute the material in any medium or format, provided it is not used for commercial purposes and the original author and source are credited, with indications if any changes are made.
creativecommons.org/licenses/by-nc-sa/4.0/
https://zoobank.org/
INTRODUCTION
The Janischewskinidae is one of the most important families for the subdivision of the Viséan and Serpukhovian (Middle and Late Mississippian), and some genera and species can be used to construct precise biostratigraphy of some regional stages in the Palaeotethys. Since the pioneer studies in the Russian Platform (e.g., Rauser-Chernousova, 1948a; Reitlinger, 1950; Ganelina, 1956), the biostratigraphic and systematic knowledge of the group has been notably improved (Lipina and Reitlinger, 1971; Makhlina et al., 1993; Vdovenko, 2001; Gibshman, 2003; Cózar and Somerville, 2006; Gibshman and Baranova, 2007; Gibshman et al., 2020; Vachard and Le Coze, 2022). In the past, some of the genera discussed in the current study were not defined formally, and the others were included within the family Bradyinidae Reilinger, 1950, with a much wider concept for the family that it is currently understood. In more recent classifications, the family Janischewskinidae has been considered as independent of the family Bradyinidae (e.g., Rauser-Chernousova et al., 1996; Vachard and Le Coze, 2022). The family Janischewskinidae is rather diversified in the Moscow Basin, where in the southern part, there are better biostratigraphic and lithostratigraphic controls, but it is possibly more abundant in the western part, and there, some species exhibit longer stratigraphic ranges, such as the large Janischewskina (compare Makhlina et al., 1993 and Kabanov et al., 2016 with Savitsky et al., 2015 and Gibshman et al., 2020). In Western Europe, although species diversity is lower than that in the Russian Platform (e.g., Cózar and Somerville, 2006, 2021; Vachard et al., 2016; Cózar et al., 2022), the family is common (e.g., Conil et al., 1980), with some well-known species being absent and/or their first occurrences being much later than those in the Moscow Basin in some cases (e.g., Cózar et al., 2019; Cózar and Somerville, 2021). Species diversity in the Urals and Ukraine seems to be also lower than that in the Moscow Basin (see Aizenverg et al., 1968; Vdovenko, 2001; Stepanova and Kucheva, 2009; Kulagina et al., 2019; Kulagina, 2022).
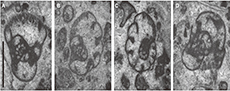
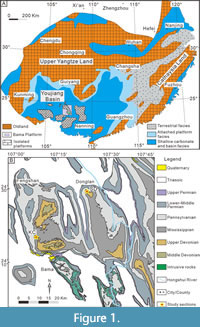 The Janischewskinidae has an important environmental dependence, and as an overall rule, they are more abundant in shallow-water facies (Gallagher, 1998; Somerville, 2008). Their abundance decreases obviously in the outer platform to slope far below the storm wave base, where the foraminiferal assemblages are alternatively dominated by other families better adapted to quiet environments, such as Lasiodiscidae (e.g., Cózar et al., 2018, 2019). This effect is also observed in the Mississippian deep-water settings in the Pengchong area, South China (Shen and Wang, 2016). In the Youjiang Basin (Figure 1), the development of isolated platforms allowed the formation of shallow-water carbonates surrounded by deep-water siliciclastic facies (Liu et al., 2014, 2015). The Bama Platform in the northern basin provides a recognizable trend from shallow- to relatively deeper-water environments, and from agitated to quiet environments, represented by the Kacai section (the shallowest and more agitated) and Shuidong section to the Gongchuan section (the deepest and quietest), and thus, the abundance of the Janischewskinidae decreases with the same trend. As a result, the family weakens its biostratigraphic importance for the Gongchuan section. The bio- and lithostratigraphy of the Bama Platform is still a work in progress, and this study is exclusively focused on the family Janischewskinidae.
The Janischewskinidae has an important environmental dependence, and as an overall rule, they are more abundant in shallow-water facies (Gallagher, 1998; Somerville, 2008). Their abundance decreases obviously in the outer platform to slope far below the storm wave base, where the foraminiferal assemblages are alternatively dominated by other families better adapted to quiet environments, such as Lasiodiscidae (e.g., Cózar et al., 2018, 2019). This effect is also observed in the Mississippian deep-water settings in the Pengchong area, South China (Shen and Wang, 2016). In the Youjiang Basin (Figure 1), the development of isolated platforms allowed the formation of shallow-water carbonates surrounded by deep-water siliciclastic facies (Liu et al., 2014, 2015). The Bama Platform in the northern basin provides a recognizable trend from shallow- to relatively deeper-water environments, and from agitated to quiet environments, represented by the Kacai section (the shallowest and more agitated) and Shuidong section to the Gongchuan section (the deepest and quietest), and thus, the abundance of the Janischewskinidae decreases with the same trend. As a result, the family weakens its biostratigraphic importance for the Gongchuan section. The bio- and lithostratigraphy of the Bama Platform is still a work in progress, and this study is exclusively focused on the family Janischewskinidae.
 Our investigation on foraminifers from the Bama Platform suggests that the foraminiferal assemblages from the middle Viséan to the lowermost upper Viséan are more similar to the Western European foraminifers than to those from the Russian Platform (Liu et al., in press). In contrast, for most of the upper Viséan and Serpukhovian, assemblages are comparable to those of the Russian Platform, a fact which justifies the use of the Russian lithostratigraphic and foraminiferal zonal schemes for comparison with the younger part of the succession (Liu et al., in press; Figure 2‒Figure 3). In the Bama Platform, representatives of the genera Cribrospira, Janischewskina, Bibradya, and Parajanischewskina have been recorded. These genera promise a biostratigraphic subdivision of the upper Viséan‒Serpukhovian in the platform, and the recognition of some regional substages described in Europe. Furthermore, these foraminifers allow a better understanding of the migration routes from basins in Europe to this sector in the eastern Palaeotethys, and the connections between basins.
Our investigation on foraminifers from the Bama Platform suggests that the foraminiferal assemblages from the middle Viséan to the lowermost upper Viséan are more similar to the Western European foraminifers than to those from the Russian Platform (Liu et al., in press). In contrast, for most of the upper Viséan and Serpukhovian, assemblages are comparable to those of the Russian Platform, a fact which justifies the use of the Russian lithostratigraphic and foraminiferal zonal schemes for comparison with the younger part of the succession (Liu et al., in press; Figure 2‒Figure 3). In the Bama Platform, representatives of the genera Cribrospira, Janischewskina, Bibradya, and Parajanischewskina have been recorded. These genera promise a biostratigraphic subdivision of the upper Viséan‒Serpukhovian in the platform, and the recognition of some regional substages described in Europe. Furthermore, these foraminifers allow a better understanding of the migration routes from basins in Europe to this sector in the eastern Palaeotethys, and the connections between basins.
 The high diversity of the family in South China allows the description of seven new species, and to clarify the systematics of the genera Cribrospira and Bibradya, as well as to confirm the validity of the genus Parajanischewskina.
The high diversity of the family in South China allows the description of seven new species, and to clarify the systematics of the genera Cribrospira and Bibradya, as well as to confirm the validity of the genus Parajanischewskina.
GEOLOGICAL AND STRATIGRAPHICAL SETTING
During the Mississippian, South China was located in the equatorial eastern Palaeotethys. The southwestern portion of the block consisted of the Upper Yangtze Land, the attached platforms along the oldland, and the Youjiang Basin (also known as the Nanpanjiang Basin) (Liu and Xu, 1994). These palaeogeographical elements brought about a striking sedimentary differentiation (Figure 1A).
Volcanic rocks and cherts suggest that during the Devonian, the Youjiang Basin was a rift basin, with eastward expansion generating two directional groups of faults (NW‒SE and NE‒SW), passing into a passive continental margin for the Carboniferous (Liu et al., 2015). The open platform facies were ascribed to the Du’an Formation (about 434 m-thick), ranging from the late Tournaisian up to the Serpukhovian (Yao and Aretz, 2020) or Viséan to Serpukhovian (Liu et al., 2015). This formation mostly contains thickly-bedded bioclastic pack- to grainstones, coral-microbial-bryozoan reefs, and intraclastic pack- to grainstones (Fang and Hou, 1989; Yao and Aretz, 2020). Persistent rifting and subsidence of the basin caused a number of tectonic blocks to tear away from the southern margin of the Yangtze Land, giving rise to a submarine landscape composed of numerous isolated shallow-water carbonate platforms surrounded by basinal facies during the Carboniferous (Liu et al., 2014) (Figure 1A).
The Bama Platform represented a typical isolated platform in the north, and its margins were associated with synsedimentary normal faults. Two shallow-water sections, the Shuidong (24º29’2’’N/107º22’58’’E; Figure 2) and Kacai (24º13’54’’N/107º9’34’’E; Figure 3) sections on the marginal position of the northern Bama Platform (Figure 1B), have been measured and sampled. These sections contain limestones of the middle Viséan to late Serpukhovian (work in progress), being laterally equivalent to the upper part of the Du’an Formation. The Kacai section (317.6 m thick) is mainly formed by shoal carbonates (i.e., gray thick-bedded to massive skeletal pack- to grainstones), and shoal and back-shoal subtidal to intertidal carbonates (i.e., gray or dark gray thick- to medium-bedded to massive skeletal wacke- to grainstones) for the Shuidong section (423 m thick) (Liu, 2017). No evident depositional hiatus was detected from the Kacai section, whereas in the Shuidong section, the lower and upper boundaries of the Plectostaffella Zone or the Zapaltyubian Substage are marked by karstic surfaces (Figure 2). Syn-rift normal-fault movements resulted in greater depositional thickness for these two sections at the platform margins than that in the platform interior Gongchuan section, which was deposited in lagoonal environments (Liu et al., 2015).
SYSTEMATIC PALAEONTOLOGY
(P. Cózar and D. Vachard)
Class FUSULINATA Maslakova, 1990 emend. Vachard et al., 2013
Subclass FUSULINANA Maslakova, 1990 emend. Vachard, 2016
Order ENDOTHYRIDA Brady, 1884 nom. translat Fursenko, 1958
Suborder ENDOTHYRINA Bogush, 1985
Superfamily BRADYINOIDEA Reitlinger, 1950 nom. translat. Rauser-Chernousova et al., 1996
Family JANISCHEWSKINIDAE Reitlinger in Rauser-Chernousova et al., 1996
Description. Test free, nautiloid to compressed laterally, with a juvenarium endothyroid passing to a planispiral final coiling. Coiling follows a progressive increase of the spire with a common rapid increase of the final whorl, in species trending to the uncoiling.
Secondary deposits absent. The most common septa are simple, curved backward, but also furrowed, blunt, swollen, and bifurcated. Cribrate aperture in the final whorl, rarely present in the penultimate chambers. Wall microgranular to granular with some agglutinated grains in the more ancestral forms, or with a porous tectum.
Composition. Bibradya Strank, 1983 (= Mirifica Shlykova, 1969, part; = Groessensella Strank in Somerville and Strank, 1984); Cribrospira Möller, 1878 (= Rhodesinella Conil and Longerstaey in Conil, 1980, part); Janischewskina Mikhailov, 1939; and Parajanischewskina Cózar and Somerville, 2006.
Remarks. The family was described in Rauser-Chernousova et al. (1996) as including Janischewskina, Bibradya (= Mirifica Shlykova, 1969, preoccupied, replaced name Ugurus Özdikmen, 2009), Cribrospira, and Groessensella. In contrast, Hance et al. (2011) and Vachard and Le Coze (2022) considered that the family only consists of Cribrospira, Janischewskina (= ? Parajanischewskina), and Rhodesinella.
The genus Ugurus (= Mirifica) shows a similar wall structure with Omphalotis Shlykova, 1969, i.e., thin dark outer tectum and thick grey inner layer, as well as a simple and basal aperture, although certainly, some furrowed and blunt septa are observed in the specimens included in the genus (e.g., Aizenverg et al., 1968, plate 8, fig. 3). The holotype of the type species Endothyra mirifica Rauser-Chernousova, 1948b is an axial section, which does not allow the recognition of the septa, nor the aperture, but it shows continuous low basal secondary deposits and a thin microgranular wall. However, the illustrated paratype is a nearly equatorial section, which shows strongly furrowed septa, incipient bifurcation of swollen septa of the sixth final chambers, and some small agglutinated grains in the wall. Although the inner whorls are a little obscure, the final whorl is free of secondary basal deposits. In addition, in the paratype, the aperture is not clearly visible, and the swollen last septum is joined to the apertural face, with a basal aperture and possibly another hole at the junction with the septum, which suggests that the last chamber is missing or has a rudimentary cribrate aperture. We interpret that in the type material of Rauser-Chernousova (1948b), there is a mixture of true Ugurus (holotype, fig. 17), and the paratype (fig. 18) belongs to Bibradya. Taking into consideration that the holotype of the type species of the genus is correct, there is no nomenclatural conflict, and this genus (skipping the features described from the paratype) should be included within the subfamily Omphalotininae Vdovenko in Rauser-Chernousova et al., 1996.
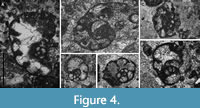 The genus Groessensella is not considered as a junior synonym of Bibradya (Figure 4A-4B, 4F), because the four specimens of the type material described by Strank (in Somerville and Strank, 1984, fig. 6h-k) are composed of rather oblique sections, which could be misinterpreted as finally uncoiled. However, a recent axial section recorded by Cózar et al. (2022, fig. 11G) shows this marked trend to the uncoiling in the final chambers (Figure 4B), but not a true uncoiling, and the genus Groessensella is thus thought to be redundant.
The genus Groessensella is not considered as a junior synonym of Bibradya (Figure 4A-4B, 4F), because the four specimens of the type material described by Strank (in Somerville and Strank, 1984, fig. 6h-k) are composed of rather oblique sections, which could be misinterpreted as finally uncoiled. However, a recent axial section recorded by Cózar et al. (2022, fig. 11G) shows this marked trend to the uncoiling in the final chambers (Figure 4B), but not a true uncoiling, and the genus Groessensella is thus thought to be redundant.
Vachard and Le Coze (2022) included Groessensella and Bibradya in the subfamily Endothyranopsinae Reitlinger, 1958, but these genera do not show planispiral coiling, nor nautiloid tests, nor thick agglutinated wall, nor quadratic chambers (except in the juvenarium), and nor a simple and basal aperture, missing mostly typical features of this subfamily, whereas, their initial skewed coiling, nautiloid tests, microgranular wall, convex chambers, and cribrate apertures are typically observed in the family Janischewskinidae.
The validity of the genus Rhodesinella is problematic based on its definition. The genus was described by Conil and Longerstaey in Conil et al. (1980) as Rhodesina (preoccupied) for the forms with granular to coarsely granular wall with agglutinated grains, using Cribrospira pansa Conil and Lys, 1965 as the type species. However, those authors also described a new species Rhodesina avonensis, which owns its particular chamber shape and coarsely granular to agglutinated wall, and was later selected as the type species of Holkeria Strank, 1982. Holkeria has been included in the subfamily Mstiniinae Lipina, 1989 by Rauser-Chernousova et al. (1996), or the family Mstiniidae Lipina, 1989 nom. translat and emend. Hance et al., 2011 (as also followed by Vachard and Le Coze, 2022), which belongs to the tournayellids/lituotubellids, not to the endothyrids. Apart from the completely different chambers and septa, the wall in Holkeria is more coarsely granular, and agglutinated grains are relatively common (Conil et al., 1980, plate XI, fig. 6, holotype; see also Figure 4C), whereas in Cribrospira pansa is microgranular, slightly granular, and with some sparse grains (agglutinated or recrystallization?) (Conil and Lys, 1965, plate III, fig. 24; see also Figure 4D‒E). These features are observed in all the primitive Cribrospira and Bibradya (e.g., Strank, 1983, plate 54, figs. 6‒8, plate 55, figs. 1‒2; Rauser-Chernousova, 1948c, plate 7, figs. 7‒8). If the small difference in the wall is admitted to be of generic value, then, the most primitive species of the genus should be considered certainly as Rhodesinella. Similarly, this progressive transition into purer microgranular walls is observed in the most evolved forms of Bibradya, which should be ascribed to a different new genus, to distinguish them from typical B. inflata Strank, 1983 and B. grandis Strank, 1983 (Figure 4A, 4F). This variation from those more granular to typical microgranular walls differentiated with tectum is considered as an intrageneric variation herein, and thus, the genus Rhodesinella is considered as a junior synonym of Cribrospira.
The validity of the genus Parajanischewskina has been questioned by some scholars (e.g., Pille et al., 2010; Vachard and Le Coze, 2022). And even, the establishment of the genus has been attributed to a possible misinterpretation of the wall nature due to oblique sections of the cribrate apertures present in the last three chambers of some Janischewskina (Gibshman et al., 2020). However, the keriotheca documented in the type species P. brigantiensis Cózar and Somerville, 2006 is more marked in the new species (P. nautiliformis) described from South China. The differentiation of the wall is used herein to justify the validity of the genus as independent of Janischewskina.
Genus Janischewskina Mikhailov, 1939
Type species. Janischewskina typica Mikhailov, 1939.
Diagnosis. Janischewskinidae with interseptal space in some cases with sutural apertures, and development of pre- and post-septal lamellae in the most advanced species.
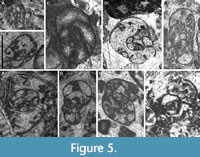 Description. Test free, nautiloid to compressed laterally, with a juvenarium endothyroid only in the first whorl, passing to planispiral. Coiling follows a progressive increase of the spire. Secondary deposits absent. The most common septa are simple, curved backward. Presence of pre- and post-septal lamellae in the most advanced species, and sutural apertures in the interseptal space. Cribrate aperture in the final whorl, rarely present in the penultimate chambers. Wall microgranular (Figure 5D).
Description. Test free, nautiloid to compressed laterally, with a juvenarium endothyroid only in the first whorl, passing to planispiral. Coiling follows a progressive increase of the spire. Secondary deposits absent. The most common septa are simple, curved backward. Presence of pre- and post-septal lamellae in the most advanced species, and sutural apertures in the interseptal space. Cribrate aperture in the final whorl, rarely present in the penultimate chambers. Wall microgranular (Figure 5D).
Composition. Janischewskina adtarusia Gibshman, Zaytseva, and Stepanova in Gibshman et al., 2020; ? J. banphitensis (Saurin, 1960); J. calceus (Ganelina, 1956); J. compressa Sosnina in Sosnina and Nikitina, 1976 (= ? J. delicata); J. delicata (Malakhova, 1956); J. gibshmanae Cózar et al. 2016; J. isotovae Lebedeva in Grozdilova et al., 1975; J. lusca (Saurin, 1960), J. minuscularia (Ganelina, 1956); J. perretae (Vachard and Cózar in Vachard et al., 2016); J. rovnensis (Ganelina, 1956); J. typica Mikhailov, 1939 (= Samarina operculata Rauser-Chernousova and Reitlinger in Rauser-Chernousova et al., 1940; = J. inflata Wang, 1982).
Remarks. Composition of the genus differs notably from that listed in Pille et al. (2010) and Gibshman et al. (2020). Some of the characters used by Gibshman et al. (2020) as distinctive for the genus Janischewskina, such as test shape, coiling symmetry, number of whorls, and wall thickness, are variable characters at species level, but in general, similar in all the genera included in the family Janischewskinidae. Only the interseptal spaces and the lamellae (in the most evolved species) are typical features of the genus, which are not observed in Samarina orbiculata Ganelina, 1956, nor in Janischewskina ladeinaensis Stephanova and Gibshman, 2017 (= Janischewskina compressa Grozdilova and Lebedeva in Grozdilova et al., 1978), and thus, they are considered herein as Cribrospira. It is noted that some specimens identified as Janischewskina orbiculata (Gibshman et al., 2020, pl. 1, fig. 7) are reinterpreted as oblique sections of J. typica.
Janischewskina compressa seems to be a junior synonym of J. delicata, although the type material shows very poor orientation as to confirm this synonymy (Gibshman et al., 2020). Janischewskina inflata shows a wide nautiloid test, with umbilical areas depressed, and secondary apertures in the interseptal space, and herein, it is considered as a junior synonym of J. typica. This overall shape was also observed in Janischewskina sp. in Groves et al. (2012), which is considered also to be this species.
The identification of J. minuscularia in older strata, for instance in the Aleksinian of the Moscow Basin, is a more robust identification than that in younger strata, because for those older rocks, there is no large Janischewskina, of which, its juveniles could be confused. However, in younger levels, the ontogenic evolution of the Janischewskina species has been never studied, and the juveniles of some species seem to be rather similar with those of J. minuscularia. Therefore, the identifications of this species in intervals co-existing with other large Janischewskina are questionable.
Occurrence. Late Viséan-Serpukhovian, northern Palaeotethyan (Vachard and Le Coze, 2022). The disappearance of the genus commonly occurred in the late Serpukhovian (e.g., Pazukhin et al., 2002; Mazuno and Ueno, 1997), which could allow the recognition of the biostratigraphy for the upper part of the Shuidong section, below the Severokeltmian top (Figure 2). However, the genus has been recorded in strata assigned to the Krasnopolyanian in the Saharan Platform, southern Morocco (Cózar et al., 2014a), and the Cantabrian Mountain, North Spain (Cózar et al., 2018), although the conditions in the former region are unusual, and a longer existence of some genera of benthic organisms was recognized (Cózar et al., 2014b).
Distribution in the Bama Platform. The first occurrence datum of the genus is equivalent to the Mikhailovian Substage, nearly from the base of the Kacai section (Figure 3). However, from the base, large species are recorded (as oblique sections), as well as J. rovnensis (Figure 5C) and J. aff. typica, whereas J. minuscularia (Figure 5A‒B) take place in intermediate positions within the Mikhailovian. The occurrence of large species of Janischewskina from the base of this interval suggests that it cannot discard the occurrence of J. minuscularia from older levels, as occurs in the Russian Platform. Only in the upper part of the Viséan, J. typica and J. isotovae are first recorded (Figure 5D‒E). The abundance of Janischewskina increases notably from the base of the Serpukhovian, and in addition to J. delicata, J. calceus and J. gibshmanae also first occur (Figure 5F‒H). Other species previously recorded in the Kacai section (Figure 3) commonly appear from the base of the Serpukhovian in the Shuidong section (Figure 2), where a late occurrence of some species is observed, i.e., J. gibshmanae only occurs from the early-late Serpukhovian transition, together with Bradyina cribrostomata Rauser-Chernousova and Reitlinger in Rauser-Chernousova and Fursenko, 1937 and Eostaffellina ex gr. paraprotvae (Rauser-Chernousova, 1948d). In the Shuidong section, J. adtarusia (Figure 5I) is first recorded from the Zapaltyubian, although the species is rare.
Genus Cribrospira Möller, 1878
Type Species. Cribrospira panderi Möller, 1878.
Diagnosis. Globose Janischewskinidae with a marked small skew-coiled juvenarium, low number of chambers, and short septa, curved backward, with swollen extremes or straight pointed. Wall granular in the primitive species, becoming microgranular to porous in more advanced forms. Cribrate aperture in the final chamber.
Composition. Cribrospira auriculata (Lin, 1981), C. baliamadeni Pille, Vachard and Argyriadis in Pille et al., 2010; C. denticulata Strank, 1983; C. evoluta sp. nov.; C. knetschi Omara and Conil, 1966; C. lebedevae Vachard and Cózar in Vachard et al., 2016 (= Janischewskina ladeinaensis Stepanova and Gibshman, 2017 = Janischewskina compressa Grozdilova and Lebedeva in Grozdilova et al., 1978), C. lianxianensis Lin, 1981; C. micula Vdovenko, 1982; C. mikhailovi Rauser-Chernousova, 1948c; C. mira Rauser-Chernousova, 1948c; C. orbiculata (Ganelina, 1956); C. panderi Möller, 1878; C. pansa Conil and Lys, 1965; C. paradenticulata sp. nov.; C. rara Rauser-Chernousova, 1948c.
Remarks. As mentioned previously, the most primitive species of the genus show a more granular wall with some agglutinated grains, and thus, they could be considered by some scholars as Rhodesinella, such as C. denticulata, C. knetschi, C. micula, C. pansa, C. paradenticulata, and C. rara. In contrast, more evolved species of the genus show a typical microgranular to porous wall (e.g., C. panderi, C. baliamadeni, and C. orbiculata).
Occurrence. Middle Viséan to late Serpukhovian in the Russian Platform, Urals, Belgium, Spain, England, Morocco, Ireland, Poland, Southern France and Ukraine. The genus has been also described in the same interval in Turkey, “Central Asia”, China, Laos, Viet-Nam, and Japan (Hance et al., 2011). Although as mentioned above, the First Occurrence Datum of the genus is usually in the middle Viséan or its equivalents, the First Appearance Datum (FAD) of the most primitive species of the genus, considered as Rhodesinella auct., is in the uppermost lower Viséan in Europe, upper Moliniacian in Belgium (Laloux, 1987), and upper Arundian in Britain (Figure 4D‒E; see also Fewtrell et al., 1981).
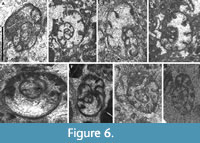 Distribution in the Bama Platform. The genus is rare below the Mikhailovian, and only Cribrospira paradenticulata has been recorded in the Bama Platform in strata equivalent to the Tulian, and Cribrospira mira (Figure 6B) from the upper part of the Aleksinian. The base of the Mikhailovian is marked by the first C. panderi (together with Eostaffella ikensis Vissarionova, 1948) (Figure 6C). These species are present in the entire Mikhailovian in the Shuidong section and only at the top of this substage C. orbiculata and C. mikhailovi first occur (Figure 6A, E). In the Kacai section, C. mikhailovi is also first recorded from the base of this substage.
Distribution in the Bama Platform. The genus is rare below the Mikhailovian, and only Cribrospira paradenticulata has been recorded in the Bama Platform in strata equivalent to the Tulian, and Cribrospira mira (Figure 6B) from the upper part of the Aleksinian. The base of the Mikhailovian is marked by the first C. panderi (together with Eostaffella ikensis Vissarionova, 1948) (Figure 6C). These species are present in the entire Mikhailovian in the Shuidong section and only at the top of this substage C. orbiculata and C. mikhailovi first occur (Figure 6A, E). In the Kacai section, C. mikhailovi is also first recorded from the base of this substage.
More diversified species are recorded from the Venevian, including C. evoluta, C. baliamadeni, and C. lianxianensis (Figure 6D, F). However, this increase in the diversity does not coincide with an increase in its abundance, which is mostly concentrated in levels assigned to the Serpukhovian.
Cribrospira paradenticulata sp. nov.
Figure 6G‒H
zoobank.org/090413BF-19F5-4E87-B059-D1B7BA103028
Derivation of the Name. For its similarity with C. denticulata.
Material. Holotype (HPU-SDB114-2, Figure 6G) and four paratypes (Table 1).
Repository. School of Resources and Environment, Henan polytechnic University (HPU).
Type Locality and Horizon. Shuidong section, Mikhailovian (late Viséan).
Occurrence. Middle Viséan to early Serpukhovian in the Shuidong section, and early Serpukhovian in the Kacai section.
Diagnosis. Cribrospira of moderate-small size with tooth-like septa and moderate evolution rate between whorls.
Description. Test free, composed of 2‒2.5 whorls, of which, coiling initially irregular in a different plane, and the final or final 1.5 are planispiral, with a diameter (D) between 670 and 1200 μm. Wall granular, calcareous. Septa slightly directed backward, short, pointed to swollen. The evolution rate increases between whorls slowly and uniformly. Height (H) of the lumen in the final chamber is 130‒280 μm, with a low H/D ratio of 0.14‒0.29. Number of chambers is moderate, ranging in the last whorl between 9 and 11. Thickness of the wall in the last chamber is 20‒45 μm. Rudimentary cribrate aperture composed of a few holes concentrated in the lower part of the apertural face.
Remarks. The species differs from C. denticulata by a higher number of chambers for similar number of whorls, larger diameter, more rudimentary aperture, thicker wall, and the occurrence of some swollen septa. It differs from other species of the genus by the shape of the septa and the granular wall.
Cribrospira evoluta sp. nov.
Figure 7A‒E
zoobank.org/4E4E8AC1-3316-4FFE-9A0D-B60FFF00D9C3
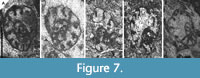 Derivation of the Name. From the evolute final whorl.
Derivation of the Name. From the evolute final whorl.
Material. Holotype (HPU-KC58-1, Figure 7A) and ten paratypes (Table 2).
Repository. School of Resources and Environment, Henan polytechnic University.
Type Hocality and Horizon. Kacai section, Tarusian (early Serpukhovian).
Occurrence. Upper part of the Mikhailovian (late Viséan) to basal Zapaltyubian (late Serpukhovian) in the Shuidong section and Venevian‒Tarusian in the Kacai section.
Diagnosis. Large Cribrospira with a high evolution rate in the final whorl, a mixture of septa (curved, furrowed, swollen), inner whorls irregularly coiled, and only the final whorl trends to be planispiral.
Description. Free large test with a diameter of 1200‒1600 μm for specimens of 2.5‒3 whorls. A juvenile specimen measures 940 μm in diameter for 1.5 whorls. Coiling irregular nearly up to the final whorl that becomes planispiral. The final whorl is located in a plane about 90º from the first whorl. Septa are usually blunt, swollen, and furrowed, but in the final chambers, they can be pointed, containing a relatively high number of chambers (9‒12 in the final whorl), with marked sutures in the final chambers and smooth inner ones. The evolution rate progresses uniformly in the inner whorls, and rapidly in the final whorl, which a height of the lumen in the final chamber between 450 and 600 μm, with high H/D ratios of 0.33‒0.40. Wall microgranular, comparatively thin, 20‒50 μm in the final chamber. Cribrate aperture in the entire apertural face.
Remarks. The species presents intermediate features between Cribrospira (nearly planispiral final coiling and curved septa) and Bibradya (more skew-coiled inner whorls and furrowed/swollen septa), allowing to distinguish from other species of Cribrospira and Bibradya.
Genus Bibradya Strank, 1983
Type Species. Bibradya inflata Strank, 1983.
Diagnosis. Janischewskinidae with irregular coiling up to the final whorl, septa bifurcated, swollen, and furrowed.
Composition. Bibradya densicamerata sp. nov.; B. grandis Strank, 1983 (= ? Mirifica mirifica part); B. inflata Strank, 1983; B. maxima sp. nov.; B. moldensis (Strank in Somerville and Strank, 1984); B. primitiva sp. nov.; B. subita sp. nov.; B. tenella (Ye et al., 1987).
Remarks. The composition of this genus has been usually restricted to the original species described by Strank (1984), although Cózar et al. (2022) also included B. moldensis, the type species of the monospecific genus Groessensella, and some ancestral forms (Bibradya ? sp. 1) from the base of the Asbian in Britain. This latter form is included herein in B. primitiva. In addition, B. tenella, originally described as a Cribrospira, is also included in the genus due to the more marked skew-coiled whorls.
Similar to the case of Cribrospira, it is possible to observe an intragenus variability of the wall structure in Bibradya, and thus, species such as B. primitiva, B. inflata, B. grandis, and B. moldensis show a more granular wall with some sparse agglutinated grains, whereas B. densicamerata, B. maxima, B. subita, and B. tenella have a typical microgranular wall. This variation seems to exist at intraspecies level, because comparing ‘Mirifica’ mirifica in Rauser-Chernousova (1948b, plate 5, fig. 18) and Aizenverg et al. (1968, plate 8, fig. 3), both specimens are homeomorphs, belonging to the same species, but the former specimen from Russia contains some agglutinated grains, whereas the latter specimens from Ukraine shows a typical microgranular wall.
 Occurrence. Late Asbian to Serpukhovian in Britain, Ireland, and China, Serpukhovian in the Sahara Platform, and in the Aleksinian from the Russian Platform (Cózar et al., 2014a; Cózar and Somerville, 2020).
Occurrence. Late Asbian to Serpukhovian in Britain, Ireland, and China, Serpukhovian in the Sahara Platform, and in the Aleksinian from the Russian Platform (Cózar et al., 2014a; Cózar and Somerville, 2020).
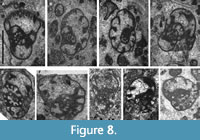
Distribution in the Bama Platform. The genus occurs from the Aleksinian (Figure 3). Surprisingly, the first recorded species is B. tenella (Figure 8G‒I), a fact which suggests that the stratigraphic record of Bibradya primitiva is not complete in the platform, a species which only occurs in strata assigned to the Mikhailovian upwards. In the upper part of the Mikhailovian, B. densicamerata and B. maxima are first recorded. The first occurrence of B. subita coincides in the two sections at the base of the Serpukhovian, and it could be considered as a regional marker.
Bibradya densicamerata sp. nov.
Figure 8A‒F
zoobank.org/CC448745-8DBE-4E77-923B-DBDF19EC8679
Derivation of the Name. For the numerous chambers in the juvenarium.
Material. Holotype (HPU-KC75-488, Figure 8B) and twenty-five paratypes (Table 3).
Repository. School of Resources and Environment, Henan polytechnic University.
Type Locality and Horizon. Kacai section, Tarusian (early Serpukhovian).
Occurrence. From the top of the Mikhailovian to the top of the Serpukhovian.
Diagnosis. Large Bibradya with a juvenarium densely packed composed of numerous quadratic chambers and a rapid expansion in the final 1.5 whorls composed of more rounded and irregular chambers.
Description. Large test composed of 3‒4 whorls for mature specimens, ranging from 1300‒1500 μm. Coiling is irregular throughout, changing progressively the plane, and increasing rapidly the evolution rate in the final 1.5 whorls. The height of the lumen in the final chamber reaches 320‒720 μm, with a H/D ratio of 0.2‒0.41. Chambers are numerous in the inner whorls, approximately 7‒8 in the second whorl, and 8‒12 in the fourth whorl. Septa are straight in the inner whorls and blunt with an incipient bifurcation in the final whorl. Wall microgranular. Cribrate aperture composed of multiple elements present in the entire apertural zone.
Remarks. Species differs from B. tenella (Figure 8G‒I) by the distinctively compressed juvenarium, more chambers, and higher number of whorls.
Bibradya primitiva sp. nov.
Figure 9A‒F
zoobank.org/81CE4253-CD52-455B-B1AC-494ABFA71371
2022 Bibradya ? sp. 1― Cózar et al., fig. 10M.
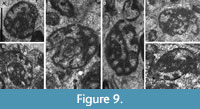 Derivation of the Name. From its primitive features.
Derivation of the Name. From its primitive features.
Material. Holotype (HPU-KC81-399, Figure 9A) and 11 paratypes (Table 4).
Repository. School of Resources and Environment, Henan polytechnic University.
Type Locality and Horizon. Kacai section, Steshevian (early Serpukhovian).
Occurrence. Mikhailovian (late Viséan) to Steshevian (early Serpukhovian) in the Kacai section and up to the Zapaltyubian in the Shuidong section.
Diagnosis. Small Bibradya with swollen, blunt septa, and incipient bifurcation of septa for a low evolution rate of the test.
Description. Test free, small to moderate size, 490‒890 μm for specimens of 2‒3 whorls. The evolution rate is low and in some of the final chambers is even lower, showing a lower height of the lumen than that in the previous chambers. Height in the last chamber 70‒170 μm, H/D ratio very low, 0.10‒0.17 (0.26). Coiling is irregular, changing progressively the plane, and does not present any whorl completely in the same plane. Septa are blunt, swollen, and with incipient bifurcation, separating numerous chambers, usually 9-11 in the last whorl, but 8 chambers in specimens of only 2 whorls. Wall microgranular to granular, comparatively thick for the size of the specimens (20‒30 μm). Aperture cribrate.
Remarks. Differs from B. inflata (Figure 4F) by a lower evolution rate, higher number of chambers, slightly higher number of whorls, less pronounced bifurcated septa, and more rudimentary cribrate apertures. The primitive characters of the species allow to distinguish it from the rest of more evolved Bibradya.
Bibradya maxima sp. nov.
Figure 10A‒E
zoobank.org/BB49C3FF-9867-4CFA-A3DF-E4C0E1959116
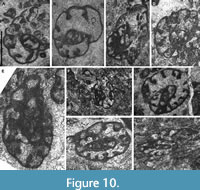 Derivation of the Name. For its large size.
Derivation of the Name. For its large size.
Material. Holotype (HPU-KC87-4, Figure 10D) and eighteen paratypes (Table 5).
Repository. School of Resources and Environment, Henan polytechnic University.
Type Locality and Horizon. Kacai section, Steshevian (early Serpukhovian).
Occurrence. From the top of the Mikhailovian to the top of the Serpukhovian.
Diagnosis. Giant Bibradya with a progressive evolution rate and furrowed and bifurcated septa in the final chambers.
Description. Large test composed of 2‒2.5 whorls, with a diameter ranging from 1600‒3000 μm. Coiling plane changes progressively and the final whorl is nearly planispiral, but not completely. Height of the lumen in the final chamber is 500‒1000 μm, with high H/D ratio of 0.3‒0.44, due to a progressive and rapid evolution rate in the successive whorls. The number of chambers is high, 10‒13.5 in the final whorl, whereas about half (5‒6) in the previous whorls. Septa in the juvenarium and early chambers of the final whorl are curved, whereas the final 5‒6 chambers are separated by swollen and furrowed septa, which tend to be bifurcated in the final 2‒3 chambers. Complex cribrate apertures occupy the entire apertural face.
Remarks. The most similar species is B. tenella, which presents similar number of chambers, whorls, septa, and coiling, although B. maxima is twice or three times larger.
B. maxima differs from B. densicamerata for the larger size, smaller number of whorls, and more expanded juvenarium.
Bibradya subita sp. nov.
Figure 10F‒I
zoobank.org/3BB52629-ECA6-44DC-87BD-6CA7C75CD8BC
Derivation of the Name. For the sudden change in the coiling plane of the final whorl.
Material. Holotype (HPU-SDB184-301, Figure 10G) and 17 paratypes (Table 6).
Repository. School of Resources and Environment, Henan polytechnic University.
Type Locality and Horizon. Shuidong section, Protvian (late Serpukhovian).
Occurrence. Tarusian to Protvian (Serpukhovian) in both sections.
Diagnosis. Large Bibradya with the inner whorls oscillating and the final whorl changing suddenly 90º the coiling plane.
Description. Large rounded to elongated test, ranging from 900 μm for specimens of 2‒2.5 whorls, up to 2000 μm for 3.5‒4 whorls. The inner whorls are nearly planispiral, with slightly oscillations, and the final whorl turns approximately 90º rapidly. This sudden change in the coiling axis is also observed in the smaller specimens (e.g., Figure 10F, I). The rate of evolution of the juvenarium is slow, and high rates for the final whorl. Height of the lumen in the final chamber is 210‒580 μm, with a moderate to high H/D ratio of 0.24‒0.48. Septa furrowed, swollen, and bifurcated in the final whorl, curved in the juvenarium. Chambers are numerous, 11‒13 in the final whorl, with marked final sutures. Wall microgranular, 30‒60 μm thick. Cribrate apertures occupy the entire apertural face.
Remarks. The arrangement of the final whorl with the sudden turn in the coiling plane distinguishes this species from other Bibradya.
Genus Parajanischewskina
Cózar and Somerville, 2006
Type Species. Parajanischewskina brigantiensis Cózar and Somerville, 2006.
Diagnosis. Janischewskinidae with slightly endothyroidal inner whorls and planispiral final whorls. Cribrate aperture in the final chambers and secondary sutural apertures with large pores. Thin, dark, and microgranular wall in most whorls, possibly formerly perforate, with an outer, darker layer. Keriotheca composed of an outer dark microgranular layer and an inner alveolar layer is present in the final septa and in the wall of the final chambers.
Composition. Parajanischewskina brigantiensis Cózar and Somerville, 2016; P. nautiliformis sp. nov.
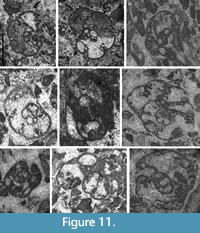
Remarks. The genus Parajanischewskina was interpreted by Cózar and Somerville (2006) as an intermediate form between Bradyina and Janischewskina, mostly due to the stratigraphic record of these three genera in Britain. However, the biostratigraphy of these genera in other basins does not support the phylogenetic lineage. Parajanischewskina is restricted in Britain to the uppermost Asbian to the lowermost Brigantian (Cózar and Somerville, 2004; Waters et al., 2017; Cózar et al., 2022), whereas the first Janischewskina (J. typica) occurred in the late Brigantian (= early Serpukhovian) (Cózar and Somerville, 2021), as a result, there is a significant gap without any representatives of both genera. Richer Janischewskinidae assemblages from the Russian Platform show small species of Janischewskina from the Aleksinian (FAD), at coeval levels with Bradyina, whereas the large species of Janischewskina occur from the Mikhailovian (Kabanov et al., 2016; Gibshman et al., 2020). Parajanischewskina has been never reported from the Russian Platform, and as mentioned previously, it was questioned if the keriotheca might be an artifact of oblique sections over double or triple cribrate apertures. However, the case can be the contrary, and the keriotheca wall in the final chambers might be confused with oblique sections of cribrate apertures (e.g., Gibshman et al., 2020, pl. 1, figs. 11‒12). Furthermore, although unrecognized by previous authors, the genus has been recorded by us in the Venevian in the Western Moscow Basin of the Russian Platform (Figure 11A‒B).
 Occurrence. Latest Asbian to late Serpukhovian in Britain, questionable in the Brigantian of SW Spain, Serpukhovian of Ireland and NW Spain, latest Viséan to late Serpukhovian in France and Sahara Platform, late Serpukhovian in Ukraine (Cózar and Somerville, 2006; Cózar et al., 2014a, 2018; Vachard et al., 2016). Venevian in the western Moscow Basin, questionable in the Serpukhovian (Khudolazian) in the Urals. The FAD of the genus Parajanischewskina might be in the late Viséan, equivalent to the Aleksinian or Mikhailovian in the Donets Basin, Ukraine (C1vf2 horizon) (Cózar and Somerville, 2006). It must be noted that the FAD of Janischewskina is located in the Aleksinian (e.g., Gibshman et al., 2020), and thus, it is not clear yet which genera, Parajanischewskina or Janischewskina, occurred first.
Occurrence. Latest Asbian to late Serpukhovian in Britain, questionable in the Brigantian of SW Spain, Serpukhovian of Ireland and NW Spain, latest Viséan to late Serpukhovian in France and Sahara Platform, late Serpukhovian in Ukraine (Cózar and Somerville, 2006; Cózar et al., 2014a, 2018; Vachard et al., 2016). Venevian in the western Moscow Basin, questionable in the Serpukhovian (Khudolazian) in the Urals. The FAD of the genus Parajanischewskina might be in the late Viséan, equivalent to the Aleksinian or Mikhailovian in the Donets Basin, Ukraine (C1vf2 horizon) (Cózar and Somerville, 2006). It must be noted that the FAD of Janischewskina is located in the Aleksinian (e.g., Gibshman et al., 2020), and thus, it is not clear yet which genera, Parajanischewskina or Janischewskina, occurred first.
Distribution in the Bama Platform. Parajanischewskina brigantiensis (Figure 11C-D) has been recorded from the upper part of the latest Viséan, whereas P. nautiliformis only occurs at the base of the Serpukhovian in both sections. The former species disappears at the base of the lower Serpukhovian, whereas the latter extends up to the top of the lower Serpukhovian.
Parajanischewskina nautiliformis sp. nov.
Figure 11E‒I
zoobank.org/A5097972-22E2-4A92-AA53-5A69AC993459
2006 Parajanischewskina ? sp. 1.― Cózar and Somerville, pl. 1, fig. 2.
Derivation of the Name. For the nautiloid form of the test.
Material. Holotype (HPU-KC75-391, Figure 11F) and six paratypes (Table 7).
Repository. School of Resources and Environment, Henan polytechnic University.
Type Locality and Horizon. Kacai section, Tarusian (early Serpukhovian).
Occurrence. Early Serpukhovian.
Diagnosis. Parajanischewskina with keriothecal wall presenting in the 4 final septa and chambers.
Description. Large nautiloid test with umbilical areas poorly depressed (e.g., Figure 11E), ranging from 1150 μm in diameter for specimens of 2 whorls up to 2150 μm for 3‒3.5 whorls. Width of the test in the umbilical region in well-oriented specimens 1200 μm. Coiling is planispiral throughout, only the first whorl slightly endothyroid. Progressive evolution rate, composed of a low number of chambers per whorl, 6‒6.5 in the final whorl. Septa are thin in the inner whorls, curved backward, passing to thick septa with keriotheca in the final 4‒5 chambers, up to 110 μm thick. Normally microgranular wall in the rest of the test, comparatively thin, 30‒50 μm thick. Cribrate apertures in the last 3‒4 chambers.
Remarks. Differs from P. brigantiensis for the occurrence of the keriotheca in more numerous septa and parts of the wall, as well as lower number of chambers per whorl.
BIOSTRATIGRAPHIC AND PALAEOBIOGEOGRAPHIC IMPLICATIONS
The occurrence of Bibradya seems to be incomplete in the studied sections, because the first B. tenella (more evolved Bibradya) from the top of the Aleksinian occurred earlier than other species apparently more primitive from the Mikhailovian. This does not follow a logical phylogenetic lineage. Thus, it is inferred that B. primitiva should first occur also from the Aleksinian. Bibradya densicamerata and B. maxima can be used for the recognition of the upper part of the Mikhailovian and on the other hand, B. subita has been documented consistently from the basal Serpukhovian. The disappearance of the genus is at the base of the Zapaltyubian or in the lower levels of this substage, which can be used as another important indicator. The last species recorded in the Zapaltyubian is B. densicamerata.
The genus Cribrospira shows rather variable first occurrences in the Bama Platform, similar to the Western Palaeotethys. The primitive forms can occur from the middle Viséan (the oldest strata have not been studied in detail in this region of China), whereas the large species were widespread in the late Viséan and Serpukhovian. It can be highlighted the first occurrence of C. panderi nearly coinciding with the first large Janischewskina, a few meters below the first Eostaffella ikensis, and that has been used for the recognition of the Mikhailovian in the basin. Rare occurrences of C. orbiculata, C. lianxianensis, C. baliamadeni, and C. evoluta are associated with the Venevian strata.
The occurrence of large species of Janischewskina is an index for the Mikhailovian. The first occurrences of J. delicata and J. gibshmanae can be used as Serpukhovian markers.
Although Parajanischewskina nautiliformis is restricted to the Serpukhovian, its rarity prevents considering this species as robust as would be desirable.
Aretz et al. (2014) proposed that foraminiferal assemblages from South China were influenced by the Western European basins and the Russian Platform during the Tournaisian and early‒middle Viséan, whereas for the late Viséan and early Serpukhovian, assemblages show more affinities with those from the Russian Platform. The parallel diversity in species of Cribrospira and Janischewskina, as well as their similar stratigraphic ranges confirm this close relationship with the Russian Platform.
However, the occurrence of Parajanischewskina and Bibradya in China is more problematic. In previous studies, the genus Parajanischewskina was restricted to Britain, Ireland, Spain, Morocco, France, and Ukraine (questionable in Sverdrup Basin, Canada), and thus, in the western Palaeotethys basins. The specimens reinterpreted from the Russian platform, as well as those recorded by us from this region, suggest that the Russian Platform might provide the source for the foraminiferal assemblages recorded in South China. As a result, it can be assumed that Parajanischewskina originated in Ukraine, but migrated to China through Russia, and the apparent absence in that region is only a matter of misidentifications with Janischewskina. The rarity of Bibradya, which is restricted to the western extreme of the Palaeotethys (Britain, Ireland, Sahara), complicates our understanding on the migration routes to China if specimens of ‘ Mirifica ’ mirifica in the Russian Platform and Ukraine are not considered to belong to this genus (see family ‘Remarks’).
CONCLUSIONS
Rich foraminiferal assemblages from the Bama Platform allow clarification of the systematic, composition, and biostratigraphy of the family Janischewskinidae.
The family is considered to be composed of four genera, Janischewskina, Cribrospira, Bibradya, and Parajanischewskina. The genera Rhodesinella and Groessensella are considered as synonyms of Cribrospira and Bibradya, respectively. Some specimens of the genus Ugurus are included under the genus Bibradya.
Seven new species have been described: Cribrospira evoluta, C. paradenticulata, Bibradya primitiva, B. densicamerata, B. maxima, B. subita, and Parajanischewskina nautiliformis.
The biostratigraphy of the Janischewskinidae in South China resembles that in the Russian Platform, i.e., primitive Cribrospira are first recorded from the Tulian, primitive Bibradya from the Aleksinian, evolved Cribrospira (e.g., C. panderi) and large Janischewskina from the Mikhailovian, Parajanischewskina and Bibradya maxima from the Venevian, and Janischewskina delicata, J. gibshmanae, Bibradya subita, and Parajanischewskina nautiliformis from the base of the Serpukhovian. On the other hand, Janischewskina adtarusia is only recorded from the upper Serpukhovian.
ACKNOWLEDGEMENTS
We would like to thank I.D. Somerville and two anonymous reviewers for their constructive comments. This contribution is financially supported by the National Natural Science Foundation of China (C. Liu, grant Numbers 41902102, 42172120, U1812402, and 41872117) and the Fundamental Research Funds for the Universities of Henan Province (C. Liu).
REFERENCES
Aizenverg, D.E., Brazhnikova, N.E., and Potievskaya, P.D. 1968. Biostratigraphic division of the Carboniferous deposits of the southern slope of the Voronezhsky Massif. Akademiya Nauk Ukrainskoi SSR, Institut Geologicheskii Nauk, “Naukova Dumka”, Kiev. (In Russian)
Aretz, M., Nardin, E., Vachard, D., Wang, Y., and Liu, M. 2014. Diversity patterns and palaeobiogeographical relationships of latest Devonian-Lower Carboniferous foraminifers from South China: What is global, what is local? Journal of Palaeogeography, 3:35-59. https://doi.org/10.3724/SP.J.1261.2014.00002
Bogush, O.I. 1985. Foraminifera and stratigraphy of the Early Carboniferous from the Western Siberia Plan. In Dubatolov, V.N. and Kanygin, A.V. (eds.), Biostratigraphy of the Paleozoic of western Siberia. Akademiya Nauk SSSR, Sibirskoe Otdelenie, Trudy Instituta Geologii i Geofiziki, 619:49-68, 195-199. (In Russian)
Brady, H.B. 1884. Report of the Foraminifera dredged by H.M.S. Challenger during the years 1873-1876. Reports on the Scientific Results of the Exploratory Voyage of the H.M.S. Challenger, Zoology, 9:1-814.
Conil, R. 1980. Note sur quelques foraminifères du Strunien et du Dinantien d’Europe occidentale. Annales de la Société géologique de Belgique, 103:43-53.
Conil, R. and Lys, M. 1965. Précisions complémentaires sur la micropaléontologie du Dinantien. Annales de la Société Géologique de Belgique, 88:B23-B44.
Conil, R., Longerstaey, P.J., and Ramsbottom, W.H.C. 1980. Matériaux pour l´étude micropaléontologique du Dinantien de Grande-Bretagne. Mémoires de l’Institut de Géologie de l’Université de Louvain, 30:1-187 (imprinted 1979).
Cózar, P. and Somerville, I.D. 2004. New algal and foraminiferal assemblages and evidence for recognition of the Asbian-Brigantian boundary in northern England. Proceedings of the Yorkshire Geological Society, 55:43-65. https://doi.org/10.1144/pygs.55.1.43
Cózar, P. and Somerville, I.D. 2006. Significance of the Bradyinidae and Parajanischewskina n. gen for biostratigraphic correlations of the late Visean (Mississippian) in Western Paleotethyan basins. Journal of Foraminiferal Research, 36:267-272. https://doi.org/10.2113/gsjfr.36.3.262
Cózar, P. and Somerville, I.D. 2020. Foraminifers in upper Viséan-lower Serpukhovian (Mississippian) limestones from South Wales: regional correlation and implications for the British foraminiferal zonal schemes. Proceedings of the Yorkshire Geological Society. 63:PGYS2020-009. https://doi.org/10.1144/pygs2020-009
Cózar, P. and Somerville, I.D. 2021. The Serpukhovian in Britain: use of foraminiferal assemblages for dating and correlating. Journal of the Geological Society, London, 178: jgs2020-170. https://doi.org/10.1144/jgs2020-170
Cózar, P., Medina-Varea, P., Somerville, I.D., Vachard, D., Rodríguez, S., and Said, I. 2014a. Foraminifers and conodonts from the late Viséan to early Bashkirian succession in the Saharan Tindouf Basin (southern Morocco): biostratigraphic refinements and implications for correlations in the western Palaeotethys. Geological Journal, 49:271-302. https://doi.org/10.1002/gj.2519
Cózar, P., Vachard, D., Somerville, I.D., Medina-Varea, P., Rodríguez, S., and Said, I. 2014b. The Tindouf Basin, a marine refuge during the Serpukhovian (Carboniferous) mass extinction in the northwestern Gondwana platform. Palaeogeography, Palaeoclimatology, Palaeoecology, 394:12-28. https://doi.org/10.1016/j.palaeo.2013.11.023
Cózar, P., Somerville, I.D., Sanz-López, J., and Blanco-Ferrera, S. 2016. Foraminiferal biostratigraphy across the Visean/Serpukhovian boundary in the Vegas de Sotres section (Cantabrian Mountains, Spain). Journal of Foraminiferal Research, 46:171-192. https://doi.org/10.2113/gsjfr.46.2.171
Cózar, P., Somerville, I.D., Blanco-Ferrera, S., and Sanz-López, J. 2018. Palaeobiogeographic context in the development of shallow-water late Viséan-early Bashkirian benthic foraminifers and calcareous algae in the Cantabrian Mountains (Spain). Palaeogeography, Palaeoclimatology, Palaeoecology, 511:620-638. https://doi.org/10.1016/j.palaeo.2018.09.031
Cózar, P., Vachard, D., Aretz, M., and Somerville, I.D. 2019. Foraminifers of the Viséan-Serpukhovian boundary interval in Western Palaeotethys: a review. Lethaia, 52:260-284. https://doi.org/10.1111/let.12311
Cózar, P., Somerville, I.D., Hounslow, M.W., and Kamenikova, T. 2022. A late Asbian (Mississippian) stratotype for England: Trowbarrow Quarry, S. Cumbria, UK. Papers in Palaeontology, 8:e1451. https://doi.org/10.1002/spp2.1451
Fang, S.X. and Hou, F.H. 1989. Bryozoan-coral patch reefs of the Carboniferous carbonate platform, Langping area, Tianlin County, Guangxi, China, p. 19-25. In Jin, Y. and Li, C. (eds.), XIe Congrès International de Stratigraphie et de Géologie du Carbonifère, Beijing 1987, Compte Runde 4.
Fewtrell, M.D., Ramsbottom, W.H.C., and Strank, A.R.E. 1981. Carboniferous, p. 13-69. In Jenkins, D.G. and Murray, J.W. (eds.), Stratigraphical Atlas of fossil Foraminifera. British Micropalaeontological Society Series, Ellis Horwood, Chichester.
Fursenko, A.V. 1958. Fundamental stages of development of foraminiferal fauna in the geological past. Trudy Instituta Geologicheskikh Nauk, Akademiya Nauk Beloruskoi SSR, 1:10-29. (In Russian)
Gallagher, S.J. 1998. Controls on the distribution of calcareous Foraminifera in the Lower Carboniferous of Ireland. Marine Micropaleontology, 34:187-211.
Ganelina, R.A. 1956. Viséan foraminifers from the northwestern areas of the Moscow Depression. Trudy Vsesoyuznogo Nauchno-Issledovatel’skogo Geologorazvedochnogo Instituta (VNIGRI) 98, Mikrofauna SSSR, 8:61-159. (In Russian)
Gibshman, N.B. 2003. Foraminifers from the Serpukhovian Stage stratotype, the Zabor’e Quarry site (Moscow region). Stratigraphy and Geological Correlation, 11:36-60.
Gibshman, N.B. and Baranova, D.V. 2007. The foraminifers Janischewskina and ‘ Millerella ’, their evolutionary patterns and biostratigraphical potential for the Visean-Serpukhovian boundary, p. 269-281. In Wong, Th.E. (ed.), Proceedings of the XVth International Congress on Carboniferous and Permian Stratigraphy. Utrecht, the Netherlands, 2003. Royal Netherlands Academy of Arts and Sciences, Utrecht.
Gibshman, N.B., Vevel, Y.A., Zaytseva, E.L., and Stepanova, T.I. 2020. Foraminifers of the Genus Janischewskina Mikhailov from the Upper Viséan-Serpukhovian (Mississippian) of Eurasia. Paleontological Journal, 54:91-110. https://doi.org/10.1134/S0031030120020045
Groves, J.R., Wang, Y., Qi, Y., Richards, B.C., Ueno, K., and Wang, X. 2012. Foraminiferal biostratigraphy of the Visean-Serpukhovian (Mississippian) boundary interval at slope and platform sections in southern Guizhou (South China). Journal of Paleontology, 86:753-774. https://doi.org/10.1666/11-111.1
Grozdilova, L.P. and Lebedeva, N.S. 1978. Paleontological Description. Foraminifers, p. 45-57. In Sultanaev A.A., Grozdilova L.P., Degtyarev D.D., Ermakova K.A., Kruchinina O.N., Lapina N.N., Lebedeva N.S., Muromtseva V.A., Rakshin P.P., Simakova M.A., Stanichnikova M.S., Tkacheva I.D., and Chudinova I.I. (eds.), The Reference Sections and Fauna of the Visean and Namurian Stages of the Middle and South Urals. Nedra, Leningrad. (In Russian)
Grozdilova, L.P., Lebedeva, N.S., Lipina, O.A., Malakhova, N.P., Mikhailova, Z.P., Chermnykh, V.A., Postoyalko, M.V., Simonova, Z.G., Sinitsyna, Z.A., Krylova, D.L., Grozdilova, A.K., Pozner, L.P., and Sultanaev, A.A. 1975. Foraminifera. In Stepanov D.L., Krylova, A.K., Grozdilova, L.P., Pozner, V.M., and Sultanaev, A.A. (eds.), Paleontological Atlas of the Carboniferous Deposits from the Urals. Trudy Vsesoyuznogo Nauchno-Issledovatel’skogo Geologorazvedochnogo Instituta (VNIGRI), 383:27-64. (In Russian)
Hance, L., Hou, H., and Vachard, D. 2011. Upper Famennian to Visean Foraminifers and some Carbonate Microproblematica from South China–Hunan, Guangxi and Guizhou. Geological Publishing House, Beijing.
Kabanov, P.B., Alekseev, A.S., Gibshman, N.B., Gabdullin, R.R., and Bershov, A.V. 2016. The upper Viséan-Serpukhovian in the type area for the Serpukhovian Stage (Moscow Basin, Russia): Part 1. Sequences, disconformities, and biostratigraphic summary. Geological Journal, 51:163-194. https://doi.org/10.1002/gj.2612
Kulagina, E.I. 2022. Foraminiferal sequence of the middle-upper Viséan (Mississippian) of the Sikasya River Valley in the South Urals and correlation with the Belgian standard. Paläontologische Zeitschrift, 96:495-516. https://doi.org/10.1007/s12542-021-00556-9
Kulagina, E.I., Gorozhanina, E.N., Gorozhanin, V.M., and Filimonova, T.V. 2019. Upper Viséan and Serpukhovian Biostratigraphy and Lithofacies of the Southeast of the East European Platform. Stratigraphy and Geological Correlation, 27:613-637. https://doi.org/10.1134/S0869593819060030
Laloux, M. 1987. Méthode de corrélations biostratigraphique et migrations; application raisonnée au Carbonifère d’Europe. Unpublished PhD Thesis, Thèse Doctorat ès Sciences Université Catholique de Louvain, Faculté des Sciences, Louvain, Belgium.
Lin, J. 1981. The Early Carboniferous Foraminifera in Guangdong and Hunan and their stratigraphical significance. Bulletin of the Yichang Institute of Geology and Mineral Resources, Special Issue, 1981:1-41.
Lipina, O.A. and Reitlinger, E.A. 1971. Stratigraphie zonale et paléozoogéographie du Carbonifère Inférieur d’après les foraminifères, p. 1101-1112. In Comptes Rendus, VI Congrès International du Carbonifère, Sheffied 1967, vol. 3, University of Sheffield, Sheffield.
Lipina, O.A. 1989. Some modifications in the systematics of tournayellids. Voprosy Mikropaleontologii, 30:30-47. (In Russian)
Liu, B. and Xu, X. 1994. Lithofacies Paleogeographic Atlas of South China: Sinian-Triassic. Science Press, Beijing, pp. 1118-1119. (In Chinese)
Liu, C. 2017. Sedimentary features and evolution of the late Paleozoic isolated Bama Platform in the Youjiang Basin: Responses to and implications for the deep-time climate change. PhD Thesis, School of Earth Sciences, China University of Geosciences, Wuhan, China.
Liu, C., Lu, G., Zhang, X., Zhou, F., Huang, X., and Du, Y. 2014. Sedimentary characteristics and evolutionary stages in late Paleozoic of Tian’e isolated platform in northwestern Guangxi. Geological Review, 60:55-70. (In Chinese with English abstract)
Liu, C., Jarochowska, E., Du, Y., Vachard, D., and Munnecke, A. 2015. Microfacies and carbon isotope records of Mississippian carbonates from the isolated Bama Platform of Youjiang Basin, South China: Possible responses to climate-driven upwelling. Palaeogeography, Palaeoclimatology, Palaeoecology, 438:96-112. https://doi.org/10.1016/j.palaeo.2015.07.048
Liu, C., Vachard, D., Cózar, P., and Coronado, I. in press. Middle to Late Mississippian and Early Pennsylvanian foraminiferal zonal scheme of South China ― a case study from the Youjiang Basin: biostratigraphic and palaeobiogeographic implications. Lethaia.
Makhlina, M.Kh., Vdovenko, M.V., Alekseev, A.S., Byvsheva, T.V., Donakova, L.M., Zhulitova, V.E., Kononova, L.I., Umnova, N.I., and Chik, E.M. 1993. Early Carboniferous of the Moscow Syneclise and Voronezh Anteclise. Rossiiskaya Akademiya Nauk, P Moskovskoe Obshchestvo Ispytatelei Pridory, Komitet po Geologii i Ispolzonalniyu Nedr Pri Pravitelstve Rossiikoi Federalii, Moskva “Nauka”. (In Russian)
Malakhova, N.P. 1956. Foraminifera of the limestones of the Shartym River in the southern Urals. Akademiya Nauk SSSR, Uralskii Filial, Trudy Gorno-Geologicheskogo Instituta, 24:26-71. (In Russian)
Maslakova, N.I. 1990. Criteria for establishing the higher taxa of Foraminifera, p. 22-27. In Menner, V.V. (ed.), Systematics and Phylogeny of Invertebrata: The Criteria for Establishing Higher Taxa. Nauka, Moscow. (In Russian)
Mazuno, Y. and Ueno, K. 1997. Conodont and foraminiferal faunal changes across the Mid-Carboniferous Boundary in the Hina Limestone Group, Southwest Japan, 1 p. 189-200. In Podemski, M., Dybova-Jachowicz, S., Jaworowski, K., Jureczka, J., and Wagner, R. (eds.), Proceedings of the XIII International Congress on the Carboniferous and Permian, Krakow 1995, vol. 3. Polish Geological Institute, Warszawa.
Mikhailov, A. 1939. On characteristic genera of Early Carboniferous foraminifers in territories of the USSR; the Lower Carboniferous deposits of the northwestern limb of Moscow basin. Sbornik Leningradskogo Geologicheskogo Upraleniya, 3:47-62. (In Russian)
Omara, S. and Kenawy, A. 1966. Upper Carboniferous microfossils from Wadi Araba, Eastern Desert, Egypt. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, 124:56-83.
Özdikmen, H. 2009. Substitute names for some unicellular animal taxa (Protozoa). Munis Entomology and Zoology, 4:233-256.
Pazukhin, V.N., Kulagina, E.I., Nikolaeva, S.V., Kochetkova, N.N., and Konovalova, V.A. 2002. Zonal subdivision of the late Visean and Serpukhovian deposits in the Verkhnayaya Karadailovka section (eastern slope of southern Urals), p. 220-229. In Chuvashov, B.I. and Amon, E.O. (eds.), Carboniferous Stratigraphy and Paleogeography in Eurasia. Institut Geologii i Geokhimii UrO RAN, Ekaterinburg. (In Russian)
Pille, L., Vachard, D., Argyriadis, I., and Aretz, M. 2010. Revision of the late Visean-Serpukhovian (Mississippian) calcareous algae, foraminifers and microproblematica from Balia-Maden (NW Turkey). Geobios. 43:531-546. https://doi.org/10.1016/j.geobios.2010.03.005
Rauser-Chernousova, D.M. 1948a. Foraminifera and stratigraphy of Viséan and Namurian stages of central part of Russian Platform and Cis-Urals. Akademiya Nauk SSSR, Trudy Instituta Geologicheskikh Nauk 62, geologicheskaya seriya, 19:102-142. (In Russian)
Rauser-Chernousova, D.M. 1948b. On some endothyras of the group of Endothyra bradyi Mikhailov. Akademiya Nauk SSSR, Trudy Instituta Geologicheskikh Nauk 62, geologicheskaya seriya, 19:176-181. (In Russian)
Rauser-Chernousova, D.M. 1948c. Genus Cribrospira Moeller. Akademiya Nauk SSSR, Trudy Instituta Geologicheskikh Nauk 62, geologicheskaya seriya, 19:186-189. (In Russian)
Rauser-Chernousova, D.M. 1948d. Some new species of foraminifera of the Early Carboniferous deposits of the Submoscovite basin. Akademiya Nauk SSSR, Trudy Instituta Geologicheskikh Nauk 62, geologicheskaya seriya, 19:227-238. (In Russian)
Rauser-Chernousova, D.M., and Fursenko, A.V. 1937. Determination of Foraminifera from the petroliferous regions of the USSR. Glavnaya Redaktsya Gorno-Toplivnoi Literatury, Leningrad. (In Russian)
Rauser-Chernousova, D.M., Belyaev, G.M. and Reitlinger, E.A. 1940. On the Foraminifera of the Carboniferous deposits of the Samara Bend. Trudy VNIGRI, Trudy Neftyanogo Geologo-razvdochnogo Instituta, novaya seriya, 7:1-88. (In Russian with English summary)
Rauser-Chernousova, D.M., Bensh, F.R., Vdovenko, M.V., Gibshman, N.B., Leven, E.Ya., Lipina, O.A., and Reitlinger, E.A. 1996. Reference-Book on the Systematics of Paleozoic Foraminifers (Endothyroids, Fusulinoids). Nauka, Moscow. (In Russian).
Reitlinger, E.A. 1950. Foraminifera from Middle Carboniferous deposits of the central part of the Russian Platform (excepting the family Fusulinidae). Akademiya Nauk SSSR, Trudy Instituta Geologicheskikh Nauk, 126, seriya geologichevskaya, 47:1-126. (In Russian)
Reitlinger, E.A. 1958. On the systematics and phylogeny of the superfamily Endothyridea. Voprosy Mikropaleontologii, 2:53-73. (In Russian)
Saurin, E. 1960. Foraminifères viséens de Ban Phit (Laos). Annales de la Faculté des Sciences de Saïgon, 1960:345-376.
Savitsky, Y.V., Kossovaya, O.L., Vevel’ Y.A., and Evdokimova, I.O. 2015. Lower Carboniferous of the St. Petersburg region (Northwestern Russia), p. 1-77. In Savitsky, Y.V., Kossovaya, O.L., and Evdokimova, I.O. (eds.), Field Guidebook of XVIII International Congress on the Carboniferous and Permian. A.P. Karpinsky All-Russian Research Geological Institute, St-Petersburg State University, St. Peterburg.
Shen, Y. and Wang, X. 2016. Howchinia Cushman, 1927 (Foraminifera) from the Mississippian Bei'an Formation and its distribution in South China. Alcheringa: An Australasian Journal of Palaeontology, 41:169-180. https://doi.org/10.1080/03115518.2016.1196437
Shlykova, T.I. 1969. New early Carboniferous foraminiferal genus. Voprosy Mikropaleontologii, 12:47-50. (In Russian)
Somerville, I.D. 2008. Biostratigraphic zonation and correlation of Mississippian rocks in Western Europe: some case studies in the late Viséan/Serpukhovian. Geological Journal, 43:209-240. https://doi.org/10.1002/gj.1097
Somerville, I.D. and Strank, A.R.E. 1984. Discovery of Arundian and Holkerian faunas from a Dinantian platform succession in North Wales. Geological Journal, 19:85-104.
Sosnina, M.I. and Nikitina, A.P. 1976. Carboniferous foraminifers of the Primorski Krai. In. Stratigraphy and Paleontology of the Carboniferous. Proceedings of the All-Union Scientific Research Geological Institute (VSEGEI), 247:16-69. (In Russian)
Stepanova, T.I. and Gibshman, N.B. 2017. A new replacement name for Janischewskina compressa Grozdilova et Lebedeva, 1978 (Foraminifera). Paleontological Journal, 51:573-573. https://doi.org/10.1134/S0031030117050136
Stepanova, T.I. and Kucheva, N.A. 2009. Palaeontological substantiation of the horizons of the Serpukhovian Stage of the eastern-Uralian subregion in the stratotype section of the Khudolaz River, p. 97-128. In Puchtov, V.N., Kulagina, E.I., Nikolaeva, S.V., and Kochetova, N.N. (eds.), Carboniferous Type Sections in Russia and Potential Global Stratotypes. Design Polygraph Service Ltd., Ufa. (In Russian)
Strank, A.R.E. 1982. Holkeria gen. nov., a foraminifer characteristic of the Holkerian Stage of the British Dinantian. Proceedings of the Yorkshire Geological Society, 44:145-151.
Strank, A.R.E. 1983. New stratigraphically significant foraminifera from the Dinantian of Great Britain. Palaeontology, 26:435-442.
Vachard, D. 2016. Macroevolution and biostratigraphy of Paleozoic foraminifers, p. 257-323. In Montenari, M. (ed.), Stratigraphy & Timescales. Elsevier, Amsterdam. https://doi.org/10.1016/bs.sats.2016.10.005
Vachard, D. and Le Coze, F. 2022. Carboniferous smaller Foraminifera: convergences and divergences, p. 247-326. In Lucas, S.G., Schneider, J.W., Wang, X., and Nikolaeva, S. (eds.), The Carboniferous Timescale. Geological Society, London, Special Publications, 512. https://doi.org/10.1144/SP512-2020-42
Vachard, D., Krainer, K., and Lucas, S. 2013. Pennsylvanian (Late Carboniferous) calcareous microfossils from Cedro Peak (New Mexico, USA); Part 2: smaller foraminifers and fusulinids. Annales de Paléontologie, 99:1-42. https://doi.org/10.1016/j.annpal.2012.08.002
Vachard, D., Cózar, P., Aretz, M., and Izart, A. 2016. Late Viséan-early Serpukhovian foraminifers in the Montagne Noire (France): Biostratigraphic revision and correlation with the Russian substages. Geobios, 49:469-498. https://doi.org/10.1016/j.geobios.2016.09.002
Vdovenko, M.V. 1982. New species of Early-Carboniferous Foraminifera in the south-western margin of the east European Platform. Geologicheskii Zhurnal, 42(5):81-86. (In Russian)
Vdovenko, M. 2001. Atlas of Foraminifera from the upper Visean and lower Serpukhovian (Lower Carboniferous) of the Donets Basin (Ukraine). Abhandlungen und Berichte für Naturkunde, 23:93-178.
Vissarionova, A.Ya. 1948. Primitive fusulinids from the Lower Carboniferous of the European part of the Union. Akademiya Nauk SSSR, Trudy Instituta Geologicheskikh Nauk 62, geologicheskaya seriya, 19:216-226. (In Russian)
von Möller, V. 1878. Die spiral-gewundenen Foraminiferen des russischen Kohlenkalks. Mémoires de l’Académie Impériale des Sciences de St Pétersbourg, 7th series, 25(9):1-147.
Wang, K. 1982. Carboniferous and Permian Foraminifera of Xizang, p. 1-32. In Nanjing Institute of Geology and Paleontology, Palaeontology of Xizang, Book IV. Science Press, Beijing. (In Chinese)
Waters, C.N., Cózar, P., Somerville, I.D., Haslam, R.B., Millward, D., and Woods, M. 2017. Lithostratigraphy and biostratigraphy of the Lower Carboniferous (Mississippian) carbonates of the southern Askrigg Block, North Yorkshire, UK. Geological Magazine, 154:305-333. https://doi.org/10.1017/S0016756815000989
Yao, L. and Aretz, M. 2020. Upper Visean (Mississippian) metazoan-microbial reefs from Guangxi, South China: Insights regarding reef recovery after the end-Devonian extinction. Palaeogeography, Palaeoclimatology, Palaeoecology, 560:109994. https://doi.org/10.1016/j.palaeo.2020.109994
Ye, G., Lin, J., and Gu, D. 1987. Early Carboniferous Foraminifera from Longmenshan, Sichuan, China. Acta Micropalaeontologica Sinica, 4:281-292. (In Chinese)
