

 The antiarch fish Asterolepis orcadensis from the Scottish Middle Devonian
The antiarch fish Asterolepis orcadensis from the Scottish Middle Devonian
Article number: 22.2.37
https://doi.org/10.26879/970
Copyright Palaeontological Association, June 2019
Author biographies
Plain-language and multi-lingual abstracts
PDF version
Submission: 21 February 2019. Acceptance: 16 May 2019
ABSTRACT
The Givetian (Middle Devonian) antiarch Asterolepis orcadensis was first described in 1932 based on a limited amount of material. Newly collected material has allowed the head to be described in more detail, especially the premedian plate which has an embayment at the centre of the anterior edge. Also, the anterior section of supra-orbital sensory line has been identified on the premedian plate. Other elements newly described include the pectoral appendages which are shown to be quite broad at their proximal ends. The anterior ventro-lateral plate has been described in detail for the first time, particularly structures associated with the attachment of pectoral appendages. The posterior median dorsal plate has been shown not to have had a more pointed posterior edge as depicted in the 1932 description, but a flatter posterior edge reminiscent of Asterolepis saevesoederberghi which is shown to be closely related to A. orcadensis. The shortness of the posterior median dorsal plate has emphasised the broadness of the whole armour, making A. orcadensis one of the broadest species of Asterolepis. Whilst A. orcadensis is endemic to the Orcadian Basin of northern Scotland, it fits into a biostratigraphical column at the same level as A. saevesoederberghi so strengthening the evidence of a close relationship between these two forms.
Michael Newman. Vine Lodge, Vine Road, Johnston, Haverfordwest, Pembrokeshire, SA62 3NZ, United Kingdom. ichthyman@btinternet.com
Jan den Blaauwen. University of Amsterdam, Science Park 904, 1098 XH, Amsterdam, Netherlands. J.L.denBlaauwen@uva.nl
David Leather. Woodlands, Panorama Drive, Ilkley, West Yorkshire LS29 9RA, UK davidleather@btinternet.com
Keywords: Orkney; Givetian; biostratigraphy; morphology; placoderm; Scotland
Newman, Michael, den Blaauwen, Jan, and Leather, David. 2019. The antiarch fish Asterolepis orcadensis from the Scottish Middle Devonian. Palaeontologia Electronica 22.2.37A 1-24. https://doi.org/10.26879/970
palaeo-electronica.org/content/2019/2587-asterolepis-orcadensis
Copyright: June 2019 Palaeontological Association.
This is an open access article distributed under the terms of Attribution-NonCommercial-ShareAlike 4.0 International (CC BY-NC-SA 4.0), which permits users to copy and redistribute the material in any medium or format, provided it is not used for commercial purposes and the original author and source are credited, with indications if any changes are made.
creativecommons.org/licenses/by-nc-sa/4.0/
INTRODUCTION
Sir John Flett collected in Deerness and South Ronaldsay in the Orkney Islands in the 1890s with a local collector Mr Spence of Deerness (Flett, 1898). They collected a single anterior median dorsal plate (NMS G.1898.1.1) at Lang Geo in Deerness. The specimen was sent to Dr Ramsay Traquair keeper of palaeontology at the Royal Scottish Museum who determined it was a new species of Asterolepis. Traquair also determined another specimen NMS G.1898.1.20 from Brough Head (Hesta Head) in South Ronaldsay as belonging to the same species which he gave the manuscript name Asterolepis orcadensis Watson ex Traquair MS, 1932. Traquair (1908) stated that Asterolepis remains had been discovered in Orkney but did not erect a species. During mapping of the Orkney Islands in the 1930s by the British Geological Survey, a further five plates of A. orcadensis were collected by survey officer W. E. Strachan. Four came from Coopalash Shore, Shapinsay and one from an old quarry near to Monquhanny farm house in Shapinsay. Watson (1932) used these specimens to erect and formally describe A. orcadensis (see below).
Mykura (1976) noted that Asterolepis orcadensis was a zone fossil for the whole of the Rousay Flagstone Formation. Michie et al. (2015) considered that A. orcadensis was probably confined to the upper part of this unit, but as will be shown in the discussion Mykura (1976) was correct.
This was the situation until the authors started collecting more specimens of Asterolepis orcadensis. This new material includes plates that Watson (1932) did not have available to study, such as a posterior median dorsal plate, anterior ventro-lateral plates and a pectoral appendage. Other new plates, such as an anterior dorso-lateral plate are also much better preserved than the material Watson (1932) had at his disposal. This allows the authors to provide a more accurate restoration of the species and better comparison with other Asterolepis species, including ones erected after Watson’s (1932) work.
All specimens studies are housed in the collections of the National Museums of Scotland, Edinburgh (NMS G), the British Geological Survey, Edinburgh (GSE), and the Palaeontological Museum, Oslo (PMO).
HISTORY OF RESEARCH ON ASTEROLEPIS
Due to the erection and description of many species of Asterolepis in difficult to obtain German and Russian journals and books, it is probably best here to give a brief history of the important works on this genus.
The type species Asterolepis ornata had been originally erected by Eichwald in 1840, but this was based on isolated plates, none of which he figured. Agassiz (1844-5) figured some incomplete plates which were not very diagnostic. Pander (1857) was the first author to describe the species in detail, figuring diagnostic plates and providing reconstructions. In fact, Eichwald must have been so impressed that he reproduced Pander’s (1857, plate 5, figure 11) lateral reconstruction, albeit flipping it laterally (Eichwald, 1860, plate 56, figure 1). Gross (1931) collected many new specimens that allowed him to describe the species in great detail, making it the most anatomically understood species of Asterolepis at that time.
The next species to be erected was Asterolepis maxima Agassiz (1844). Agassiz (1844-5) based this species on a single, dorsally preserved anterior median dorsal plate (AMD). He thought it represented a median ventral plate of a Coccosteus, but Miller in a communication to Egerton (1848, p. 310-311) correctly identified it as an AMD of an antiarch. However, it was Pander (1857) who correctly placed it in the genus Asterolepis. Traquair (1888) became interested in describing the species in more detail firstly by describing an AMD plate including the visceral side which showed the characteristic areas of overlap. Woodward (1891a) also figured the visceral side of an AMD plate in the Natural History Museum. Later, Traquair (1894) had more material at his disposal and provided a thorough description of the species including a reconstruction of the complete fish.
The final 19th century species to be erected was Asterolepis scabra Woodward (1891b). Woodward (1891b) had only part of a headshield and part of an anterior dorso-lateral plate available to describe. The species was not recognised as an Asterolepis species until Stensiö (1918) made this determination. Later, Stensiö (1931) described new material of A. scabra and Asterolepis ornata. In this work he went into great detail on the antiarch morphology including all known Asterolepis species.
The last species to be erected before Asterolepis orcadensis was Asterolepis radiata Rohan,1900. However, as pointed out by Stensiö (1931), Rohon’s (1900) description was “imperfect” and the figures’ rough outline drawings making determination difficult.
This was the situation when Watson (1932) erected Asterolepis orcadensis. He noted it was different to the other known species by its wedged shaped head and short trunk armour among other characters. In the same paper Watson erected another species from Scotland Asterolepis thule Watson, 1932 which he differentiated from all other species by the narrowness of the skull.
In 1933 Obruchev erected a new species Asterolepis cristata Obruchev, 1933 based on specimens from the Leningrad region of Russia. He considered the ornamentation and other minor characters specifically different from Asterolepis radiata to warrant a new species. However, there is still some controversy on this assertion (see below).
In 1938 Stensiö erected the new species Asterolepis saevesoederberghi Stensiö, 1938 from the Middle Devonian of East Greenland. Stensiö (1938) noted the similarities between Asterolepis orcadensis and A. saevesoederberghi compared to the other Asterolepis species but noted some specific character differences as well.
In 1940 Gross erected two new species, Asterolepis estonica Gross, 1940 and Asterolepis dellei Gross, 1940 from the Middle Devonian of the Baltic region. He had numerous specimens of A. estonica which allowed him to describe the species in detail, although he had no articulated specimens. Gross (1940) had only a lateral plate and a posterior median dorsal plate at his disposal to describe A. dellei which meant it was poorly known.
Nilsson (1941) provided a new detailed description of Asterolepis scabra based on newly collected material, some of it articulated. He was able to provide a reconstruction of the head and compare it closely with other known species. Like Stensiö (1938) Nilsson (1941) noted the similarities between Asterolepis orcadensis and Asterolepis saevesoederberghi. Nilsson (1941, figure 18, plate 15, figures 4-5) described the anterior half of an anterior median dorsal plate (PMO A24374) of Asterolepis cf. orcadensis. However, due to its incomplete nature it is difficult to determine accurately.
Karatajūtė-Talimaa (1963) in a large work redescribed all the known Baltic and Russian species in minute anatomical detail. She was able to provide a much more detailed description of Asterolepis radiata and Asterolepis dellei than had hitherto been made. Of particular use was the reconstructions of the head of A. dellei and head and body of A. radiata. This has allowed comparison with Asterolepis orcadensis, although Karatajūtė-Talimaa (1963) did not attempt this herself.
In 1964 Wells’ described a new species Asterolepis chadwicki Wells, 1964 from the Late Devonian of New York state, the first species of that genus discovered in the Americas. Wells (1964) specimens were disarticulated plates. However, amongst the specimens was a morphologically important anterior median dorsal plate.
Murphy et al (1976) described some Asterolepis material from the Late Devonian of Red Hill, Nevada. Some of the Murphy et al. (1976) material was articulated, including a complete head and a specimen with a tail (Schultze, 2010, figure 4). It is, therefore, surprising that Murphy et al. (1976) chose not to give it a formal name, as the description and material was adequate to do so.
In 1981 Lyarskaya described all the Baltic and Russian species in detail. She had at her disposal fully articulated (including the tail) newly discovered specimens of Asterolepis ornata found at Lode Quarry in Latvia. She also erected two new species, Asterolepis essica Lyarskaya, 1981 and Asterolepis syasiensis Lyarskaya, 1981. Lyarskaya (1981) had a lot of material available of A. essica including a partially articulated headshield. This allowed her to provide a reconstruction of the armour of the fish. Lyarskaya (1981) only had a limited number of disarticulated plates of A. syasiensis, although this did include the morphologically important anterior median dorsal plate. Lyarskaya (1981) also erected a possible new species Asterolepis ? amulensis Lyarskaya, 1981. This was based on a solitary anterior ventro-lateral plate which is strictly speaking not diagnostic for the genus.
Moloshnikov (2008) described new material from Russia of Asterolepis radiata. He also described some remains he referred to Asterolepis cf. A. syasiensis.
Following on from Lyarskaya’s (1981) work on Asterolepis ornata from Lode Quarry, Upeniece (2001; 2011) was able to describe the ontogenetic development of the species from very small, probable hatchlings to larger young adult specimens.
Finally, Newman and den Blaauwen (2018) provided a new description of Asterolepis thule including the description of a more complete headshield than Watson (1932) had at his disposal. This allowed Newman and den Blaauwen (2018) to furnish a new reconstruction of the head.
SYSTEMATIC PALAEONTOLOGY
Suborder ASTEROLEPIDOIDEI Miles, 1968
Family ASTEROLEPIDIDAE Traquair, 1888
Genus ASTEROLEPIS Eichwald, 1840
Type species. Asterolepis ornata Eichwald, 1840
Diagnosis. See Lyarskaya 1981.
Included species. Asterolepis chadwicki Wells, 1964, Asterolepis dellei Gross, 1940, Asterolepis essica Lyarskaya, 1981, Asterolepis estonica Gross, 1940, Asterolepis maxima (Agassiz, 1844), Asterolepis orcadensis Watson, 1932, Asterolepis radiata Rohan, 1900, Asterolepis saevesoederberghi Stensiö, 1938, Asterolepis scabra (Woodward, 1891), Asterolepis syasiensis Lyarskaya, 1981, Asterolepis thule Watson, 1932 and Asterolepis ? amulensis Lyarskaya, 1981.
Remarks. Asterolepis cristata Obruchev, 1933 was considered a valid species until Karatajūtė-Talimaa (1963) made it a junior synonym of Asterolepis radiata. Later, Moloshnikov (2008) commented on A. cristata and recognised several characters he thought unique to the species. He proposed it was probably a valid species. However, Ivanov and Lebedev (2011) still considered A. cristata to be a junior synonym of A. radiata and Baltic and Russian researchers are still studying these forms to get a resolution to this problem (Lukševičs pers. comm.)
Asterolepis orcadensis Watson ex Traquair MS, 1932
Figure 1, Figure 2, Figure 3, Figure 4, Figure 5, Figure 6, Figure 7, Figure 8, Figure 9, Figure 10, Figure 11, Figure 12, Figure 13, Figure 14, Figure 15, Figure 16
V*1932 Asterolepis orcadensis [Traquair MS] sp. nov.; Watson, p. 161-163, fig. 3, pl. 4.
1935 Asterolepis orcadensis (Traquair) Watson; Watson, p. 160, fig. 25.
1937 Asterolepis orcadensis; Westoll, p. 523.
1938 A. orcadensis Watson; Stensiö, p. 8, 10-11, 13, 18
1940 Asterolepis orcadensis Watson; Gross, p. 26-27, fig. 6.
1941 A. orcadensis Watson (1932); Nilsson, p. 5, 16, 18, 23-24, 33, 36, 42-44, 50.
1976 Asterolepis orcadensis Watson; Mykura, p. 78, pl. 14, fig. 5.
1978 A. orcadensis Watson 1932; Denison, p. 113.
1981 Asterolepis orcadensis Watson 1932 ex Traquair MS; Paton, p. 45.
1981 Asterolepis orcadensis Watson; Lyarskaya, p. 22, 29, 142, fig. 63.9.
1999 Asterolepis orcadensis; Dineley, p. 174.
2015 Asterolepis orcadensis Watson, 1932; Michie, Newman and den Blaauwen, p. 149, 154, fig. 1
2015 Asterolepis orcadensis; De Vleeschouwer, Leather and Claeys, fig. 2.
2018 Asterolepis orcadensis Watson, 1932; Newman and den Blaauwen, p. 69, 73
Lectotype. Watson (1932) when he erected the species described, five specimens thereby making them syntypes. We choose GSE 12828 (originally referred to as V.1810B in Watson (1932)), an impression of a headshield in dorsal aspect, as the lectotype as it is the most diagnostically informative.
Material and localities. From Coopalash Shore, Shapinsay (British Grid reference HY 540203), GSE 12829 (originally referred to as V.1820B in Watson (1932)) the posterior end of a right anterior dorsal lateral plate, GSE 12830 (originally referred to as V.1821B in Watson (1932)), a right mixilateral plate missing the posterior end and ventral margin and NMS G.2019.1.1 an impression of a right anterior ventral lateral plate; Lang Geo, Deerness (British Grid reference HY 593090), NMS G.1898.1.1 a complete anterior median dorsal plate; Long Geo, Holm, East Mainland (British Grid reference HY 530007), NMS G.1897.57.1, and the posterior part of a right posterior ventral lateral plate; Hesta Head Quarry, South Ronaldsay (British Grid reference ND 464883), NMS G.1898.1.20 an incomplete, badly preserved mixilateral plate; Taft, Westray (British Grid reference HY 502412), NMS G.2018.8.16.1 complete head and NMS G.2018.8.16.2 an associated right pectoral appendage, NMS G.2018.8.22 Nuchal plate missing part of the left side and NMS G.2018.8.17.1 and NMS G.2018.8.17.2 left ADL plate; Stancro Shore, Westray (British Grid reference HY 50127.40548), NMS G.2018.8.6 a posterior median dorsal plate; Loth Quarry, Sanday (British Grid reference HY 602343), NMS G.2018.8.3 a right anterior ventral lateral plate and NMS G.2018.8.5 a left anterior ventral lateral plate.
Type locality. Lang Geo, Deerness, Orkney (British grid reference HY 593090).
Remarks. Monquhanny Farm quarry, Shapinsay, 130 yards E. 43 degrees S of the farm (British Grid reference HY 503177) would normally be chosen as the type locality as this is where the lectotype came from. However, as this quarry has been filled in and no longer exists, we chose the Coopalash Shore locality instead as it is the most accessible and some of the specimens described by Watson (1932) when erecting the species came from there.
Diagnosis. A medium sized Asterolepis with a very wide head, particularly at the posterior margin with a maximum breadth/length index of around 1.55. The premedian plate is very long and wide with a length/breadth ratio of around 1.37. The post pineal plate is moderately short and wide with a breadth/length index of around 1.39. The nuchal plate is relatively wide and short. The thorax is also very broad with the anterior median dorsal plate being very wide, even taking into account compaction and diagenesis. It has a breadth/length index of around 0.88. The posterior median dorsal plate is short and wide with a breadth/length index of around 1.16. The posterior edge of the posterior median dorsal plate has a very shallow convex profile which turns into a shallow concave profile at the centre point.
 Description. Head. General characters. The two articulated headshields, GSE 12828 (Figure 1.1-2) and NMS G.2018.8.16.1 (Figure 1.3-4) are broadly similar. GSE 12828 is preserved as an impression of the dorsal surface (so Figure 1.2 has been flipped horizontally to show the headshield in true dorsal aspect) which is damaged at the anterior end, whilst NMS G.2018.8.61 is preserved dorsally, but is damaged on the right-side margin. The headshield is of moderate length but moderately wide, particularly at the posterior end. The orbital socket is fairly small being quite short and not very wide. There is a universal coverage of quite large tubercles.
Description. Head. General characters. The two articulated headshields, GSE 12828 (Figure 1.1-2) and NMS G.2018.8.16.1 (Figure 1.3-4) are broadly similar. GSE 12828 is preserved as an impression of the dorsal surface (so Figure 1.2 has been flipped horizontally to show the headshield in true dorsal aspect) which is damaged at the anterior end, whilst NMS G.2018.8.61 is preserved dorsally, but is damaged on the right-side margin. The headshield is of moderate length but moderately wide, particularly at the posterior end. The orbital socket is fairly small being quite short and not very wide. There is a universal coverage of quite large tubercles.
Premedian plate (Prm). The anterior end of the premedian plate is damaged in GSE 12828, so this description is mostly based on NMS G.2018.8.16.1. The plate is sub-square being fairly long and narrow with a breath/length index of 1.37 at the centre lines of the plate. The anterior margin is slightly concave in profile with the corresponding posterior margin being slightly convex in profile. The infraorbital sensory lines enter the plate from the two lateral plates and runs to the centre of the plate where it meets the anterior section of the supra-orbital sensory line. These sensory lines run quite far forward anteriorly about a third from the anterior margin. The tubercles run in rows laterally with no obvious radiation point.
Lateral plate (L). The left and right lateral plates are symmetrical. The plates are long and relatively narrow, being broader at the posterior end. A prominent process on the anterior postmarginal angle is present. The middle of the inner surface forms the lateral edges of the orbital socket. The infraorbital sensory line runs the whole length of the plate from the anterior section of the supra-orbital sensory line at the suture of the premedian plate to the paranuchal plate. The tubercles form rows that follow the length of the plate, particularly the orbital margin and there is no obvious radiation centre of the plate.
Post pineal plate (Pp). In the post pineal plate of GSE 12828, the anterior edge is still covered in matrix and so the description of this plate is mostly based on NMS G.2018.8.16.1. This plate is sub-square in shape being slightly wider than long with a breadth/length index of 1.39 at the centre lines of the plate. The anterior end forms the posterior margin of the orbital socket which is mostly horizontal except for a slightly convex profile at the centre. The lateral edges are sub-vertical with the posterior margin forming a shallow V-shape. There is no obvious radiation point of the plate and the tubercles are densely pack in a random fashion. There are no sensory lines visible on the plate.
 Nuchal plate (Nu). The nuchal plate can be morphologically variable but is of the usual Asterolepis form. The plate is short and wide with a breadth/length ratio of 1.63 at the widest point, at the lateral corners in NMS G.2018.8.16.1 which is the most complete specimen. The lateral corners are placed about half the length of the lateral edges. The Nu is better preserved in NMS G.2018.8.16.1 than GSE 12828. NMS G.2018.8.22 (Figure 2) is a third isolated specimen preserved of this plate, although most of the left-hand side is missing. This specimen is reasonably well-preserved along the posterior margin which shows a thin, smooth mesial division forming the posterior border of the headshield. In NMS G.218.8.16.1 the middle pit-line groove (mp) enters the plate from the paranuchal plates and runs across the whole width of the plate. However, the plate is somewhat broken in the centre of this specimen and it is difficult to discern between cracks and pit lines. The middle pit-line grooves of GSE 12828 are faint and appear to come from the paranuchal plate as in NMS G.2018.8.16.1, but do not meet in the centre. In GSE 12828 there is a pair of short posterior pit-line grooves (pp) that aim diagonally down to the lateral edges. There are no clear external openings of the canal for the endolymphatic duct. However, none of the specimens are well-preserved in this area. GSE 12828 the tubercles form ridges running in rows radiating from the centre point of the plate.
Nuchal plate (Nu). The nuchal plate can be morphologically variable but is of the usual Asterolepis form. The plate is short and wide with a breadth/length ratio of 1.63 at the widest point, at the lateral corners in NMS G.2018.8.16.1 which is the most complete specimen. The lateral corners are placed about half the length of the lateral edges. The Nu is better preserved in NMS G.2018.8.16.1 than GSE 12828. NMS G.2018.8.22 (Figure 2) is a third isolated specimen preserved of this plate, although most of the left-hand side is missing. This specimen is reasonably well-preserved along the posterior margin which shows a thin, smooth mesial division forming the posterior border of the headshield. In NMS G.218.8.16.1 the middle pit-line groove (mp) enters the plate from the paranuchal plates and runs across the whole width of the plate. However, the plate is somewhat broken in the centre of this specimen and it is difficult to discern between cracks and pit lines. The middle pit-line grooves of GSE 12828 are faint and appear to come from the paranuchal plate as in NMS G.2018.8.16.1, but do not meet in the centre. In GSE 12828 there is a pair of short posterior pit-line grooves (pp) that aim diagonally down to the lateral edges. There are no clear external openings of the canal for the endolymphatic duct. However, none of the specimens are well-preserved in this area. GSE 12828 the tubercles form ridges running in rows radiating from the centre point of the plate.
Paranuchal plate (Pnu). The paranuchal plates are sub-square in shape and somewhat variable. The infraorbital sensory lines enter the paranuchal plates from the lateral plates. The sensory line then splits with the cephalic division of main lateral line running at right angles to the lateral edge and the middle pit-line groove running at right angles into the nuchal plate. The plates are not well enough preserved to infer any distribution of the tubercles although they appear heavily tuberculated.
Postmarginal plate (Pmg). A left postmarginal plate is known from GSE 12828. It is sub-ovoid in shape and sutures to the right to the paranuchal plate and anteriorly onto the lateral plate. It is quite broad with a breadth/length index of 1.00 at its widest point. The lateral margin is slightly convex, with the lateral posterior corner being quite pronounced. The posterior edge is horizontally straight. The plate is heavily ornamented with tubercles in no obvious pattern. There are no sensory lines on this plate.
 Thorax. Anterior median dorsal plate (AMD). Only one specimen is known of this plate being NMS G.1898.1.1 (Figure 3.1) a plate preserved viscerally. It is flattened and therefore quite broken, but all the overlaps areas are clear. A natural impression of this plate is also preserved (Figure 3.2). As with other species of Asterolepis there is no ventral pit in this plate. The plate is broad with a breadth/length index of 0.88 at its widest point. However, great caution is required in using this figure due to the flattening of the plate and the index would have been considerably smaller. The median ridge seen as a hollow in the visceral surface is shallow but may have been considerably deeper before compaction. The areas overlapping the anterior dorsal lateral plates (cf.ADL) are narrow and occupy the anterior half of the lateral edges. The areas overlapping the mixilateral plates (cf.MxL) are much broader and occupy the posterior half of the plate. There is a semi-circular impression of the area overlapped by the posterior median dorsal plate (i.oa.PMD) on the posterior margin. Near the lateral edge of the right-hand side, a small patch of the plate is broken away revealing an impression of the dorsal surface. This shows closely packed, quite small tubercles. No pattern can be discerned from this small patch. The visceral surface does display growth lines emanating from the central part of the plate.
Thorax. Anterior median dorsal plate (AMD). Only one specimen is known of this plate being NMS G.1898.1.1 (Figure 3.1) a plate preserved viscerally. It is flattened and therefore quite broken, but all the overlaps areas are clear. A natural impression of this plate is also preserved (Figure 3.2). As with other species of Asterolepis there is no ventral pit in this plate. The plate is broad with a breadth/length index of 0.88 at its widest point. However, great caution is required in using this figure due to the flattening of the plate and the index would have been considerably smaller. The median ridge seen as a hollow in the visceral surface is shallow but may have been considerably deeper before compaction. The areas overlapping the anterior dorsal lateral plates (cf.ADL) are narrow and occupy the anterior half of the lateral edges. The areas overlapping the mixilateral plates (cf.MxL) are much broader and occupy the posterior half of the plate. There is a semi-circular impression of the area overlapped by the posterior median dorsal plate (i.oa.PMD) on the posterior margin. Near the lateral edge of the right-hand side, a small patch of the plate is broken away revealing an impression of the dorsal surface. This shows closely packed, quite small tubercles. No pattern can be discerned from this small patch. The visceral surface does display growth lines emanating from the central part of the plate.
 Posterior median dorsal plate (PMD). NMS G.2018.8.6 is the only known PMD plate and is preserved in three dimensions (Figure 4.1). A natural cast of the dorsal surface (Figure 4.2) is also preserved. The specimen is flattened and broken in places, but the cast gives a good overall dimension of the plate. The plate is very broad for Asterolepis with a breadth/length index of 1.16. The dorsal median ridge (dmr) is fairly pronounced along the whole length of the plate. The posterior end of the plate is very flat with no posterior corner evident. The lateral corners (l) are rounded as is the anterior margin. The dorsal surface has tubercles radiating near the posterior central part of the plate. Growth lines reflect this, showing the plate grew from this central area. On the visceral side (Figure 4.3-4) the two contact faces for the mixilateral plates (cf.MxL) are clear to see on the lateral edges, although they are broken and incomplete. The contact face for the anterior median dorsal plate at the anterior end has been broken off and is not preserved. The posterior transversal internal crest (cr.tp) is low and has only slight posterior curvature reflecting the rather flat posterior end of the plate. The posterior marginal area (pma) whilst narrow, is consistent and does not seem to narrow at the lateral margins. The ventral pit (pt) is broad and with the middle slightly curved to the anterior as seen in other Asterolepis species. Other characters are not discernible due to the broken nature of the plate.
Posterior median dorsal plate (PMD). NMS G.2018.8.6 is the only known PMD plate and is preserved in three dimensions (Figure 4.1). A natural cast of the dorsal surface (Figure 4.2) is also preserved. The specimen is flattened and broken in places, but the cast gives a good overall dimension of the plate. The plate is very broad for Asterolepis with a breadth/length index of 1.16. The dorsal median ridge (dmr) is fairly pronounced along the whole length of the plate. The posterior end of the plate is very flat with no posterior corner evident. The lateral corners (l) are rounded as is the anterior margin. The dorsal surface has tubercles radiating near the posterior central part of the plate. Growth lines reflect this, showing the plate grew from this central area. On the visceral side (Figure 4.3-4) the two contact faces for the mixilateral plates (cf.MxL) are clear to see on the lateral edges, although they are broken and incomplete. The contact face for the anterior median dorsal plate at the anterior end has been broken off and is not preserved. The posterior transversal internal crest (cr.tp) is low and has only slight posterior curvature reflecting the rather flat posterior end of the plate. The posterior marginal area (pma) whilst narrow, is consistent and does not seem to narrow at the lateral margins. The ventral pit (pt) is broad and with the middle slightly curved to the anterior as seen in other Asterolepis species. Other characters are not discernible due to the broken nature of the plate.
 Anterior dorso-lateral plate (ADL). Only one right ADL plate GSE 12829 (Figure 5) was used in this study. The other specimen mentioned by Watson (1932) in the GSE collection (V.1819B) is unfortunately lost and cannot be considered further here. GSE 12829 is preserved mostly as a natural mould of the outer surface, although part of the bone is preserved viscerally on the lateral side of the plate. The mesial margin of the dorsal lamina (dlm) is missing, so the area overlapping the anterior median dorsal plate is also missing. This also means we cannot give the relative dimensions of GSE 12829. The anterior part of the lateral edge of the lateral lamina (llm) is also broken away with the viscerally preserved bone covering the rest of the lateral edge. This, therefore, obscures the area that overlaps the anterior ventral lateral plate. The anterior margin of the plate is mostly broken and reveals little information. Large portions of the plate are preserved either side of the dorso-lateral ridge (dlr). The main lateral line groove (lcg) is visible as an impression at the top third of the preserved plate. It runs subparallel to the length of the plate before it is obscured by the preserved bone of the visceral part of the plate. The ornamentation consists of closely packed tubercles running in rows subparallel to the dlr. The visceral part of the bone that is preserved shows growth lines that indicate the bone grew outward from the centre of the plate.
Anterior dorso-lateral plate (ADL). Only one right ADL plate GSE 12829 (Figure 5) was used in this study. The other specimen mentioned by Watson (1932) in the GSE collection (V.1819B) is unfortunately lost and cannot be considered further here. GSE 12829 is preserved mostly as a natural mould of the outer surface, although part of the bone is preserved viscerally on the lateral side of the plate. The mesial margin of the dorsal lamina (dlm) is missing, so the area overlapping the anterior median dorsal plate is also missing. This also means we cannot give the relative dimensions of GSE 12829. The anterior part of the lateral edge of the lateral lamina (llm) is also broken away with the viscerally preserved bone covering the rest of the lateral edge. This, therefore, obscures the area that overlaps the anterior ventral lateral plate. The anterior margin of the plate is mostly broken and reveals little information. Large portions of the plate are preserved either side of the dorso-lateral ridge (dlr). The main lateral line groove (lcg) is visible as an impression at the top third of the preserved plate. It runs subparallel to the length of the plate before it is obscured by the preserved bone of the visceral part of the plate. The ornamentation consists of closely packed tubercles running in rows subparallel to the dlr. The visceral part of the bone that is preserved shows growth lines that indicate the bone grew outward from the centre of the plate.
 NMS G.2018.8.17.2 (Figure 6) is the only left ADL plate that was available for study. It is preserved as a natural mould of the outer surface of the whole plate. NMS G.2018.8.1 (Figure 7) is the anterior part of the bone preserved from this mould. It is preserved in three dimensions so allowing description of both the outer and visceral surface. NMS G2018.8.17.1 gives the overall shape of the plate which is sub-ovoid with a flat anterior end. The dorsal lamina (dlm) is moderately broad with a breadth/length ratio of 0.33. The lateral lamina (llm) is narrower with a breadth/length ration of 0.24, thus giving the whole plate a breadth/length ratio of 0.62. The dorso-lateral ridge (dlr) is not so clear on NMS G.2018.8.17.2, but the closely packed tubercles running subparallel to it are. Also clear is the area overlapping the anterior median dorsal plate (oa.AMD) and the area overlapping the anterior ventro-lateral plate (oa.AVL). The oa.AMD runs nearly two thirds the length of the plate from the anterior end of the plate. It is shaped like a long, thin oval. The oa.AVL is shorter at a little under half the length of the plate from the anterior end. It is similarly shaped to the oa.AMD.
NMS G.2018.8.17.2 (Figure 6) is the only left ADL plate that was available for study. It is preserved as a natural mould of the outer surface of the whole plate. NMS G.2018.8.1 (Figure 7) is the anterior part of the bone preserved from this mould. It is preserved in three dimensions so allowing description of both the outer and visceral surface. NMS G2018.8.17.1 gives the overall shape of the plate which is sub-ovoid with a flat anterior end. The dorsal lamina (dlm) is moderately broad with a breadth/length ratio of 0.33. The lateral lamina (llm) is narrower with a breadth/length ration of 0.24, thus giving the whole plate a breadth/length ratio of 0.62. The dorso-lateral ridge (dlr) is not so clear on NMS G.2018.8.17.2, but the closely packed tubercles running subparallel to it are. Also clear is the area overlapping the anterior median dorsal plate (oa.AMD) and the area overlapping the anterior ventro-lateral plate (oa.AVL). The oa.AMD runs nearly two thirds the length of the plate from the anterior end of the plate. It is shaped like a long, thin oval. The oa.AVL is shorter at a little under half the length of the plate from the anterior end. It is similarly shaped to the oa.AMD.
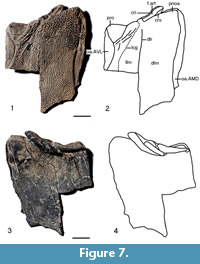 The outer surface of NMS G2018.8.17.1 (Figure 7.1-2) mirror images NMS G.2018.8.17.2, although large parts of the posterior part of the plate are missing. Also, care must be taken as the specimen is crushed flat and there is a bit of distortion and breakage. On NMS G.2018.8.17.1 the area overlapping the anterior median dorsal plate (oa.AMD) and the area overlapping the anterior ventro-lateral plate (oa.ADL) are clear. They are also now in the correct orientation of the right and left side of the plate retrospectively. The dorso-lateral ridge (dlr) and the subparallel rows of tubercles running alongside are also more prominent than in NMS G.2018.8.17.1. Of more importance is that the anterior end of the plate is much better preserved than NMS G.2018.8.17.1. The postnuchal ornamented corner (pnoa) is prominent. The processus obstans (pro) is quite a large, relatively deep, smooth triangular area. There are a lot of large ridges running parallel to the right side of the pro, but they may be a preservational feature. The main lateral line groove (lcg) runs to the right of these ridges in the usual position for Asterolepis. The articular fossa for the exoskeletal cervical joint (f.art) is clearly demarcated by the supra-articular cristae (crs) and the intra-articular cristae (cri).
The outer surface of NMS G2018.8.17.1 (Figure 7.1-2) mirror images NMS G.2018.8.17.2, although large parts of the posterior part of the plate are missing. Also, care must be taken as the specimen is crushed flat and there is a bit of distortion and breakage. On NMS G.2018.8.17.1 the area overlapping the anterior median dorsal plate (oa.AMD) and the area overlapping the anterior ventro-lateral plate (oa.ADL) are clear. They are also now in the correct orientation of the right and left side of the plate retrospectively. The dorso-lateral ridge (dlr) and the subparallel rows of tubercles running alongside are also more prominent than in NMS G.2018.8.17.1. Of more importance is that the anterior end of the plate is much better preserved than NMS G.2018.8.17.1. The postnuchal ornamented corner (pnoa) is prominent. The processus obstans (pro) is quite a large, relatively deep, smooth triangular area. There are a lot of large ridges running parallel to the right side of the pro, but they may be a preservational feature. The main lateral line groove (lcg) runs to the right of these ridges in the usual position for Asterolepis. The articular fossa for the exoskeletal cervical joint (f.art) is clearly demarcated by the supra-articular cristae (crs) and the intra-articular cristae (cri).
The visceral side (Figure 7.3-4) of the articular fossa for the exoskeletal cervical joint (f.art) is thick and long, running subparallel to the anterior edge. It has been stretched and broken up with compaction and would have been a single structure. Whilst it does not reach as far as to be underneath the postnuchal ornamented corner (pnoa), it does run under the processus obstans (pro). Growth ridges are visible that indicate growth started from the anterior middle part of the plate.
 Mixilateral plate (MxL). GSE 12830 (Figure 8) is a left MxL with the ventral and anterior ends missing. The specimen is mostly preserved as an impression, although on the right anterior part of the plate the bone is preserved viscerally. Due to the incomplete nature of the plate it is not possible to give relative dimensions of it. The area overlapped by the posterior median dorsal plate (oa.PMD) is preserved on the lower left edge of the specimen being a half, thin semicircle. It is posterior of the prominent dorsal corner (d). The oa.PMD is preserved posteriorly and anteriorly as in indent caused by a break in the plate on the visceral bone. The area overlapped by the anterior median dorsal plate is covered by the visceral surface and so not visible. No other areas of overlap are visible. The dorso-lateral ridge (dlr) is preserved as a shallow impression running the length of the preserved plate. The main lateral line groove (lcg) is less distinct but is preserved as a shallow ridge to the right of the dlr. The tubercles are tightly packed and often form ridges that run from the dlr. These ridges can be seen as impressions on the visceral bone. The visceral bone also has growth ridges that indicate growth from what would have been the centre of the plate.
Mixilateral plate (MxL). GSE 12830 (Figure 8) is a left MxL with the ventral and anterior ends missing. The specimen is mostly preserved as an impression, although on the right anterior part of the plate the bone is preserved viscerally. Due to the incomplete nature of the plate it is not possible to give relative dimensions of it. The area overlapped by the posterior median dorsal plate (oa.PMD) is preserved on the lower left edge of the specimen being a half, thin semicircle. It is posterior of the prominent dorsal corner (d). The oa.PMD is preserved posteriorly and anteriorly as in indent caused by a break in the plate on the visceral bone. The area overlapped by the anterior median dorsal plate is covered by the visceral surface and so not visible. No other areas of overlap are visible. The dorso-lateral ridge (dlr) is preserved as a shallow impression running the length of the preserved plate. The main lateral line groove (lcg) is less distinct but is preserved as a shallow ridge to the right of the dlr. The tubercles are tightly packed and often form ridges that run from the dlr. These ridges can be seen as impressions on the visceral bone. The visceral bone also has growth ridges that indicate growth from what would have been the centre of the plate.
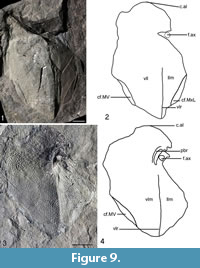 Anterior ventro-lateral (AVL). Two left AVL plates are known with NMS G.2018.8.5 (Figure 9.1-2) being an impression of the visceral side. NMS G.2018.8.5 is not very well preserved with the axillary foramen (f.ax) missing. The edges of the plate are missing either due to them not being preserved or slightly covered by matrix. The area overlapping the posterior ventro-lateral plate is not preserved well enough to observe. The overall shape of the plate is sub-oval with a breadth/length index of about 0.63. The ventral lamina (vlm) is about three times as large as the lateral lamina (llm) and about twice as broad. The anterio-lateral corner of subcephalic division (c.al) at the anterior edge of the plate is rounded as in other Asterolepis species. The impression of the ventro-lateral ridge (vlr) is seen running the length of the plate, except where there is damage. The area overlapping the mixilateral plate (cf.MxL) forms a thin sub-oval on the posterior lateral edge. The area overlapping the median ventral plate (cf.MV) is partly preserved on the posterior ventral edge of the plate. No other overlap or areas overlapped can be identified with confidence.
Anterior ventro-lateral (AVL). Two left AVL plates are known with NMS G.2018.8.5 (Figure 9.1-2) being an impression of the visceral side. NMS G.2018.8.5 is not very well preserved with the axillary foramen (f.ax) missing. The edges of the plate are missing either due to them not being preserved or slightly covered by matrix. The area overlapping the posterior ventro-lateral plate is not preserved well enough to observe. The overall shape of the plate is sub-oval with a breadth/length index of about 0.63. The ventral lamina (vlm) is about three times as large as the lateral lamina (llm) and about twice as broad. The anterio-lateral corner of subcephalic division (c.al) at the anterior edge of the plate is rounded as in other Asterolepis species. The impression of the ventro-lateral ridge (vlr) is seen running the length of the plate, except where there is damage. The area overlapping the mixilateral plate (cf.MxL) forms a thin sub-oval on the posterior lateral edge. The area overlapping the median ventral plate (cf.MV) is partly preserved on the posterior ventral edge of the plate. No other overlap or areas overlapped can be identified with confidence.
 The other left anterior ventro-lateral plate (AVL) NMS G.2019.1.1 (Figure 9.3-4) is preserved as an impression of the ventral and lateral side. The plate is missing much of the anterior ventral edge as the area overlapped by the right AVL plate is not visible. A small, thin semi-circle impression on the posterior ventral edge may be the area overlapping the median ventral plate cf.MV superimposed from the visceral side of the plate. Like NMS G.2018.8.5 the ventro-lateral ridge (vlr) runs the length of the plate to the axillary foramen (f.ax) which is situated about three quarters the length of the plate from the posterior edge on the lateral edge. This makes the subcephalic division about 25 % of the length of the ventral lamina (vlm). Preservation around the f.ax is not particularly good, although the impression of the process brachialis (pbr) is placed anteriorly medially of the f.ax. The f.ax is quite small and sub-rounded. The anterio-lateral corner of subcephalic division (c.al) is rounded like NMS G.2018.8.3. The tubercles run in ridge sub-parallelly to the vlr but spread out in a fan shape away from it. They appear to emanate from the region of the f.ax.
The other left anterior ventro-lateral plate (AVL) NMS G.2019.1.1 (Figure 9.3-4) is preserved as an impression of the ventral and lateral side. The plate is missing much of the anterior ventral edge as the area overlapped by the right AVL plate is not visible. A small, thin semi-circle impression on the posterior ventral edge may be the area overlapping the median ventral plate cf.MV superimposed from the visceral side of the plate. Like NMS G.2018.8.5 the ventro-lateral ridge (vlr) runs the length of the plate to the axillary foramen (f.ax) which is situated about three quarters the length of the plate from the posterior edge on the lateral edge. This makes the subcephalic division about 25 % of the length of the ventral lamina (vlm). Preservation around the f.ax is not particularly good, although the impression of the process brachialis (pbr) is placed anteriorly medially of the f.ax. The f.ax is quite small and sub-rounded. The anterio-lateral corner of subcephalic division (c.al) is rounded like NMS G.2018.8.3. The tubercles run in ridge sub-parallelly to the vlr but spread out in a fan shape away from it. They appear to emanate from the region of the f.ax.
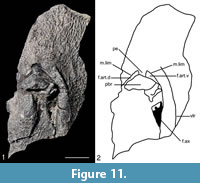 NMS G.2018.8.3 is a right anterior ventro-lateral plate (AVL) plate mostly preserved as an impression of the ventral and lateral side (Figure 10). However, a small piece of bone (Figure 11) is also preserved in three dimensions from NMS G.2018.8.3 which includes the axillary foramen (f.ax). The impression of NMS G.2018.8.3 is poorly preserved and less complete than NMS G.2019.1.1 and is, therefore, less useful for descriptive purposes. The lateral side of the small piece of bone, however, is useful anatomically. The visceral side of this piece of bone is badly damaged and not of much use anatomically. The lateral side is also broken in places but is generally well preserved. The f.ax is quite long although it is difficult to be precise about its dimensions as there is broken bone at the anterior end. The processus brachialis (pbr) is fairly broad and to the anterior left of the f.ax. The posterior right of the pbr is quite broken, so it is difficult to determine the nature of the funnel pit or its surroundings. The margo limitans (m.lim) is anterior of the f.ax and pbr and is moderately long forming a semi-circular shape broken by the pars pedalis of the processus brachialis (pe) which is fairly broad. The pe emanates from the pbr which divides the fossa articularis pectoralis into dorsal and ventral branches (f.art.d and f.art.v respectfully). Neither the f.art.d and f.art.v are seen to pass posteriorly of the pbr halfway point, but there is damage and distortion in this area, so this may be a preservational feature. Finally, the ventro-lateral ridge is quite pronounced. It runs as a shallow curve to the right of the f.ax and m. lim. It continues a short way anterior of the pbr before shallowing out.
NMS G.2018.8.3 is a right anterior ventro-lateral plate (AVL) plate mostly preserved as an impression of the ventral and lateral side (Figure 10). However, a small piece of bone (Figure 11) is also preserved in three dimensions from NMS G.2018.8.3 which includes the axillary foramen (f.ax). The impression of NMS G.2018.8.3 is poorly preserved and less complete than NMS G.2019.1.1 and is, therefore, less useful for descriptive purposes. The lateral side of the small piece of bone, however, is useful anatomically. The visceral side of this piece of bone is badly damaged and not of much use anatomically. The lateral side is also broken in places but is generally well preserved. The f.ax is quite long although it is difficult to be precise about its dimensions as there is broken bone at the anterior end. The processus brachialis (pbr) is fairly broad and to the anterior left of the f.ax. The posterior right of the pbr is quite broken, so it is difficult to determine the nature of the funnel pit or its surroundings. The margo limitans (m.lim) is anterior of the f.ax and pbr and is moderately long forming a semi-circular shape broken by the pars pedalis of the processus brachialis (pe) which is fairly broad. The pe emanates from the pbr which divides the fossa articularis pectoralis into dorsal and ventral branches (f.art.d and f.art.v respectfully). Neither the f.art.d and f.art.v are seen to pass posteriorly of the pbr halfway point, but there is damage and distortion in this area, so this may be a preservational feature. Finally, the ventro-lateral ridge is quite pronounced. It runs as a shallow curve to the right of the f.ax and m. lim. It continues a short way anterior of the pbr before shallowing out.
 Posterior ventro-lateral (PVL). Only one specimen is known being NMS G.1897.57.1 (Figure 12). It is preserved for the most part viscerally, although part of the bone is missing revealing the impression of the tubercles which are tightly packed with no obvious rows. Growth lines are visible on the visceral surface which indicate the plate grew from the centre. However, the plate is very incomplete and only the posterior part is preserved. As there is no overlap area visible on the visceral surface for a right PVL plate we can say this NMS G.1897.57.1 is a right PVL plate. Other than that, the plate shows few specific characters other than being curved at the posterior end, typical of this plate in other Asterolepis species.
Posterior ventro-lateral (PVL). Only one specimen is known being NMS G.1897.57.1 (Figure 12). It is preserved for the most part viscerally, although part of the bone is missing revealing the impression of the tubercles which are tightly packed with no obvious rows. Growth lines are visible on the visceral surface which indicate the plate grew from the centre. However, the plate is very incomplete and only the posterior part is preserved. As there is no overlap area visible on the visceral surface for a right PVL plate we can say this NMS G.1897.57.1 is a right PVL plate. Other than that, the plate shows few specific characters other than being curved at the posterior end, typical of this plate in other Asterolepis species.
Pectoral appendages. Only one pectoral appendage is known being NMS G.2018.8.16.2 (Figure 13) a right pectoral appendage missing most of the distal end. Even considering compaction with fossilisation the proximal segment is short at around twice as long as wide. Care must be taken in describing NMS G.2018.8.16.2 as it is broken in places and the entirety of the individual plates is not always preserved. Plus, sometimes the sutures between the plates can be difficult to ascertain. The arrangement of the individual bones is the same as other Asterolepis species. The ventral central plate 1 (Cv1) is about a third as long as broad at the broadest point at the distal margin. The external articular area (ar3d) is of the normal form. The plate sutures mesially onto the mesial marginal plate 1 (Mm1) and the mesial lateral plate 2 (Ml2). The Cv1 sutures posteriorly on to the ventral central 2 plate (Cv2). The Mm1 plate is unusual as it has a distinct process (pMm1) on the upper right margin. The Cv2 is quite long and thin being about twice as long as broad at the broadest point at the distal margin. The Cv2 sutures mesially to the mesial marginal plate 2 (Mm2) and the Ml2.  The Cv2, Ml2 and Mm2 plates articulate at their distal ends onto the ventral central plate 3 (Cv3), the lateral marginal plate 3 (Ml3) and the mesial marginal plate 3 (Mm3). The contacts are, however, not well preserved and the Cd3, Ml3 and Mm3 plates are not complete at their distal ends. The dorsal central plate 1 (Cd1) is a about 2½ times as long as broad at its broadest point on the distal margin. It sutures the Ml2 plate, Mm2 plate and dorsal central plate 2 (Cd2). The Cd2 plate is about twice as long as broad. The distil end of this plate articulates with the dorsal central plate 3 (Cd3), Mm3 plate and the Ml3 plate. These plates are of course incomplete at their distal ends. Ventrally, Ml2 forms nearly the whole lateral margin of the proximal segment. The ornamentation consists of rows of tubercles running subparallel along the lengths of the individual plates. Short, blunt spines are present along the lateral and mesial edges of the plates. The broken nature of the plates makes it impossible to discern any sensory lines.
The Cv2, Ml2 and Mm2 plates articulate at their distal ends onto the ventral central plate 3 (Cv3), the lateral marginal plate 3 (Ml3) and the mesial marginal plate 3 (Mm3). The contacts are, however, not well preserved and the Cd3, Ml3 and Mm3 plates are not complete at their distal ends. The dorsal central plate 1 (Cd1) is a about 2½ times as long as broad at its broadest point on the distal margin. It sutures the Ml2 plate, Mm2 plate and dorsal central plate 2 (Cd2). The Cd2 plate is about twice as long as broad. The distil end of this plate articulates with the dorsal central plate 3 (Cd3), Mm3 plate and the Ml3 plate. These plates are of course incomplete at their distal ends. Ventrally, Ml2 forms nearly the whole lateral margin of the proximal segment. The ornamentation consists of rows of tubercles running subparallel along the lengths of the individual plates. Short, blunt spines are present along the lateral and mesial edges of the plates. The broken nature of the plates makes it impossible to discern any sensory lines.
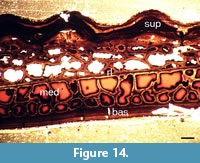 Histology. A thin section of an incomplete, unidentifiable plate NMS G.2018.8.11.4 from Stancro Shore, Westray was made (Figure 14). It can be seen that the histology of Asterolepis orcadensis follows very closely the pattern of Asterolepis ornata described by Gross (1931). Comparison of Figure 14 with Gross’s 1931 plate 1 are practically identical. However, this seems to be a general pattern in antiarchs, which consists of a three-layered structure, with a basal superficial lamellar layer (bas), a cancellous spongiosa middle layer (med) and an upper compact basal lamellar layer (sup). This structure was described in detail by Giles et al. (2013, e.g., figure 5). As in Giles et al. (2013) description a resting line (rl) is also present. It is, therefore, unnecessary to describe in detail the histology of NMS G.2018.8.11.4 as it is identical to the description of antiarch histology in Giles et al. (2013).
Histology. A thin section of an incomplete, unidentifiable plate NMS G.2018.8.11.4 from Stancro Shore, Westray was made (Figure 14). It can be seen that the histology of Asterolepis orcadensis follows very closely the pattern of Asterolepis ornata described by Gross (1931). Comparison of Figure 14 with Gross’s 1931 plate 1 are practically identical. However, this seems to be a general pattern in antiarchs, which consists of a three-layered structure, with a basal superficial lamellar layer (bas), a cancellous spongiosa middle layer (med) and an upper compact basal lamellar layer (sup). This structure was described in detail by Giles et al. (2013, e.g., figure 5). As in Giles et al. (2013) description a resting line (rl) is also present. It is, therefore, unnecessary to describe in detail the histology of NMS G.2018.8.11.4 as it is identical to the description of antiarch histology in Giles et al. (2013).
COMPARISON OF ASTEROLEPIS ORCADENSIS WITH OTHER ASTEROLEPIS SPECIES
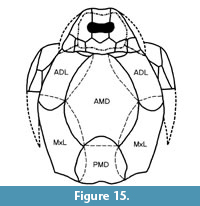 When Watson (1932, figure 3) reconstructed Asterolepis orcadensis he recognised it as a very broad fish. With the new material the authors provide a new reconstruction (Figure 15) which emphasises this character even more so, particularly with its shorter posterior median dorsal plate. As detailed below, A. orcadensis is a distinct species easily identified by key characters.
When Watson (1932, figure 3) reconstructed Asterolepis orcadensis he recognised it as a very broad fish. With the new material the authors provide a new reconstruction (Figure 15) which emphasises this character even more so, particularly with its shorter posterior median dorsal plate. As detailed below, A. orcadensis is a distinct species easily identified by key characters.
The simplified reconstruction of Asterolepis orcadensis (Figure 16.1) show that it has some distinctive characters (e.g., very broad anterior median dorsal plate) that are directly comparable with many of the other species of Asterolepis. It has a long and narrow premedian plate that clearly distinguish it from most other species, such as Asterolepis ornata (Figure 16.2), Asterolepis maxima (Figure 16.3), Asterolepis scabra (Figure 16.4), Asterolepis radiata (Figure 16.5), Asterolepis thule (Figure 16.6) and Asterolepis essica (Figure 16.7).
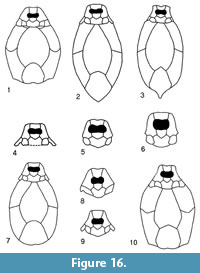 There are no premedian plates (Prm) of Asterolepis dellei known. However, Karatajūtė-Talimaa (1963, plate 4, figure 1) made a model reconstruction (see Figure 16.8 for a simplified version) of the known plates where the Prm plate is long and narrow like Asterolepis orcadensis. The limited amount of A. dellei plates does make further comparison difficult, but the nuchal plate is narrower than A. orcadensis and the ornamentation of all the plates is quite different.
There are no premedian plates (Prm) of Asterolepis dellei known. However, Karatajūtė-Talimaa (1963, plate 4, figure 1) made a model reconstruction (see Figure 16.8 for a simplified version) of the known plates where the Prm plate is long and narrow like Asterolepis orcadensis. The limited amount of A. dellei plates does make further comparison difficult, but the nuchal plate is narrower than A. orcadensis and the ornamentation of all the plates is quite different.
Another distinctive character of Asterolepis orcadensis is the short and wide anterior median dorsal plate. It also has a comparatively broad anterior end where it contacts the head. This immediately distinguishes it from Asterolepis chadwicki (Wells, 1964, plate 73, figures 1-2) and Asterolepis syasiensis (Lyarskaya, 1981, figure 100) which have very narrow anterior ends. This is a useful diagnostic character as otherwise the two species are poorly known.
Asterolepis ? amulensis is based on an anterior ventro-lateral plate. Lyarskaya (1981, figure 104) considered it characterised by a very narrow lateral lamina as well as a very small axillary foramen. The lateral lamina is much wider in Asterolepis orcadensis thereby showing the two forms to be distinct.
Asterolepis estonica has a very variable anterior median dorsal plate (Karatajute-Talimaa, 1963, figure 14) making comparison with Asterolepis orcadensis difficult in regard to this plate. However, the nuchal plate in A. estonica is much narrower, with a very narrow posterior end (Gross, 1940, figure 8), quite unlike A. orcadensis.
There is also an un-named species described by Murphy et al (1976). Not only is there an articulated headshield of this material, there are also complete specimens including the tail (Schultze, 2010, figure 4). Reed (1979) described the material in an unpublished PhD thesis but did not give it a formal name. Lyarskaya (1981, figure 63.8) figured one of Reed’s (1979) reconstructions of the headshield and Figure 16.9 is a simplified drawing of that figure. The headshield is similar to Asterolepis orcadensis but differs in its very long premedian plate and strong curvature of the lateral plates. In general terms the headshield of A. orcadensis is much shorter and wider.
Finally, the species that has been noted by a number of authors (e.g., Stensiö, 1938; Nilsson, 1941) to be most similar to Asterolepis orcadensis is Asterolepis saevesoederberghi (Figure 16.10). They both share a similar dimensioned premedian plate. Stensiö (1938) noted that the posterior end of the posterior median dorsal plate (PMD) was quite different to Asterolepis orcadensis. However, this was based on Watson’s (1932) reconstruction which he seems to have based on the PMD of Asterolepis maxima. To all intend and purposes the PMD is identical between A. saevesoederberghi and A. orcadensis. Stensiö (1938) also thought the nuchal plate (Nu) was longer in A. saevesoederberghi but had some doubt on the variability of the length as he figured a much shorter Nu (Stensiö, 1938, figure 9) from the same block as the holotype. So, in many respects the heads of A. orcadensis and A. saevesoederberghi could be very difficult to tell apart if indeed the Nu plate is at times shorter than the restoration (Figure 16.10). Regarding the body trunk, the anterior median dorsal plate of A. saevesoederberghi is much longer and narrower than A. orcadensis, but apart from this the other post cranial plates are very similar. It seems these two species are very closely related and if more material were found, particularly the Nu and AMD plates, they may even be co-specific.
DISCUSSION
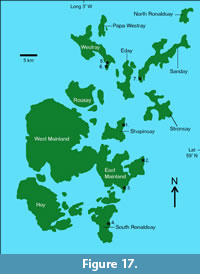 Asterolepis orcadensis had previously been found high up in the Rousay Flagstone Formation, not far below the Eday Group (Michie et al., 2015). However, in this study we describe NMS G.2018.8.6, a posterior median dorsal plate of A. orcadensis from Stancro Shore in Westray that was found associated with the tetrapodamorph fish Osteolepis panderi (Pander, 1860). Fish beds with O. panderi present mark the base of the Rousay Flagstone Formation (Michie et al., 2015). This confirms the view of Mykura (1976) that the species is found throughout the formation. However, it has a limited distribution only being found at a few localities (Figure 17). It has been noted by the authors that the species is generally found in shallow water deposits.
Asterolepis orcadensis had previously been found high up in the Rousay Flagstone Formation, not far below the Eday Group (Michie et al., 2015). However, in this study we describe NMS G.2018.8.6, a posterior median dorsal plate of A. orcadensis from Stancro Shore in Westray that was found associated with the tetrapodamorph fish Osteolepis panderi (Pander, 1860). Fish beds with O. panderi present mark the base of the Rousay Flagstone Formation (Michie et al., 2015). This confirms the view of Mykura (1976) that the species is found throughout the formation. However, it has a limited distribution only being found at a few localities (Figure 17). It has been noted by the authors that the species is generally found in shallow water deposits.
Marshall and Astin (1996) noted something similar in East Greenland with Asterolepis saevesoederberghi being found in certain facies, namely fluvial and shallow water deposits. Therefore, some caution must be taken when making up a biostratigraphical column based on the genus Asterolepis. Nonetheless, from Figure 18 we see that the strata where Asterolepis saevesoederberghi is confirmed as being present, the Genvejdalen Member of the Ankerbjergselv Formation of East Greenland, is at about the same horizon as the top of the Rousay Flagstone Formation (Blom et al., 2007). Whilst at present we consider the two Asterolepis species to be separate species they are undoubtedly closely related. This may indicate a close faunal contact as other co-specific genera are also present such as Homostius, Glyptolepis, Gyroptychius and Thursius (Blom et al., 2007). Indeed, Jarvik (1950; 1984) noted the similarity between the Scottish species Gyroptychius milleri Jarvik, 1948 and the East Greenland species Gyroptychius groenlandicus Jarvik, 1950. At the time of his descriptions (Jarvik, 1950; 1984), only disarticulated heads were known of G. milleri. However, now complete, articulated specimens of G. milleri have been collected. Perhaps, closer examination between these two species might show even closer similarities?
 In the Baltic deposits of Estonia, the equivalent stratum is the Tarvastu Member of the Arükula Regional Stage, where Asterolepis estonica is the Asterolepis species present. As the authors pointed out above, Asterolepis orcadensis and A. estonica can clearly be differentiated by the morphology of the nuchal plate. However, due to the limited material available of A. estonica, it is difficult to make a more detailed comparison between the two species to see how closely related they are to one another. They are probably closely related as Newman et al., 2015 showed that the Scottish species Millerosteus minor (Miller, 1858) was probably present in the Tarvastu Member (being described as Millerosteus cf. minor). In fact, a number of other recent papers have shown the close faunal content between the Baltic Devonian and the Scottish Orcadian Basin (Burrow et al., 2016; Newman et al., 2017; Newman and Trewin, 2008).
In the Baltic deposits of Estonia, the equivalent stratum is the Tarvastu Member of the Arükula Regional Stage, where Asterolepis estonica is the Asterolepis species present. As the authors pointed out above, Asterolepis orcadensis and A. estonica can clearly be differentiated by the morphology of the nuchal plate. However, due to the limited material available of A. estonica, it is difficult to make a more detailed comparison between the two species to see how closely related they are to one another. They are probably closely related as Newman et al., 2015 showed that the Scottish species Millerosteus minor (Miller, 1858) was probably present in the Tarvastu Member (being described as Millerosteus cf. minor). In fact, a number of other recent papers have shown the close faunal content between the Baltic Devonian and the Scottish Orcadian Basin (Burrow et al., 2016; Newman et al., 2017; Newman and Trewin, 2008).
As regards the Spitsbergen form Asterolepis scabra, it is found in the Fiskløfta Member of the Tordalen Formation in the Mimer Valley. There has been some debate on the age of these deposits (see Berry and Marshall, 2015; Piepjohn and Dallmann, 2014) but Newman et al. (2019) have shown without doubt that the Fiskeløfta Member is stratigraphically higher than the Rousay Flagstone Formation, notwithstanding the morphological differences between the two species pointed out above. The other species of Asterolepis can all be shown to be in higher strata than the Rousay Flagstone Formation with the American forms even being in the Late Devonian.
CONCLUSIONS
The species Asterolepis orcadensis is now one of the better-known species of Asterolepis. It is most closely related to Asterolepis saevesoederberghi, but limited material of both species makes comparison difficult. This is due to the lack of material of key plates, such as the nuchal and anterior median dorsal plate, which mean we do not know their species variability of these plates. Regarding regional biostratigraphy, the genus might not be as useful as once thought (certainly outside the Baltic region), as it is often ecologically controlled, albeit in a fairly narrow stratigraphical range. A. orcadensis appears endemic to the Orcadian Basin, which appears to be a trait of a number of species, such as Asterolepis thule. This further reduces the use Asterolepis in regional biostratigraphy.
ACKNOWLEDGEMENTS
We thank two anonymous reviewers and the editors for suggestions to improve the manuscript. We also thank G. Tulloch at the British Geological Survey in Edinburgh and S. Walsh at the National Museums of Scotland for access to specimens in their care. This is a contribution to IGCP596: Climate change and biodiversity patterns in the mid-Palaeozoic (Early Devonian to Late Carboniferous).
REFERENCES
Agassiz, J.L.R. 1844-45. Monographie des Poissons du Vieux Grès Rouge ou Systeme Devonien (Old Red Sandstone) des Isles Britanniques et de Russie. Jent and Gassmann, Neuchâtel. https://doi.org/10.5962/bhl.title.5752
Berry, C.M. and Marshall, J.E.A. 2015. Lycopsid forests in the early Late Devonian paleoequatorial zone of Svalbard. Geology, 43:1043-1046. https://doi.org/10.1130/g37000.1
Blom, H., Clack, J.A., Ahlberg, P.E., and Friedman, M. 2007. Devonian vertebrates from East Greenland: a review of faunal composition and distribution. Geodiversitas, 29:119-141.
Burrow, C.J., den Blaauwen, J.L., Newman, M.J., and Davidson, R.G. 2016. The diplacanthid fishes (Acanthodii, Diplacanthiformes, Diplacanthidae) from the Middle Devonian of Scotland. Palaeotologia Electronica, 19.1.10A:1-83. https://doi.org/10.26879/601
De Vleeschouwer, D., Leather, D., and Claeys, P. 2015. Ripple marks indicate Mid-Devonian paleo-wind directions in the Orcadian Basin (Orkney Isles, Scotland). Palaeogeography, Palaeoclimatology, Palaeoecology, 426:68-74. https://doi.org/10.1016/j.palaeo.2015.03.001
Denison, R.H. 1978. Placodermi, p. 1-128. In Schultze, H.-P. (ed.), Handbook of Paleoichthyology, Part 2. Gustav Fischer Verlag, Stuttgart and New York.
Dineley, D.L. 1999. Mid-Devonian fossil fish sites of Scotland, p. 167-222. In Dineley, D.L. and Metcalf, S.J. (eds.), Fossil Fishes of Great Britain. Joint Nature Conservation Committee, Peterborough, UK.
Egerton, P.G. 1848. Palichthyologic notes, supplemental to the works of Prof. Agassiz. Quarterly Journal of the Geological Society, 4:302-314. https://doi.org/10.1144/gsl.jgs.1848.004.01-02.43
Flett, J.F. 1898. The Old Red Sandstone of the Orkneys. Transactions of the Royal Society of Edinburgh, 39:383-424. https://doi.org/10.1017/s008045680003413x
Eichwald, E. 1840. Die Tier- und Pflanzenreste des alten roten Sandsteins und Bergkalks im Nowgorodschen Gouvernement. Saint-Petersburg Academy Imperial of Sciences, Bulletin of Sciences, 7:78-91.
Eichwald, E. 1860. Lethaea Rossica ou paléontologie de la Russié, 1, seconde sectione de l’ancienne période, p. 681-1657. E. Schweizerbart, Stuttgart. https://doi.org/10.5962/bhl.title.52391
Giles, S., Rucklin, M., and Donoghue, P.C.J. 2013. Histology of “placoderm” dermal skeletons: implications for the nature of the ancestral gnathostome. Journal of Morphology, 274:627-644. https://doi.org/10.1002/jmor.20119
Gross, W. 1931. Asterolepis ornata Eichw. und das Antiarchi-Problem. Palaeontographica, 75:1-61.
Gross, W. 1940. Acanthodier und Placodermen aus den Heterostius-Schichten Estlands und Lettlands. Publications of the Geological Institution of the University of Tartu, 60:1-88.
Ivanov, A. and Lebedev, O. 2011. Devonian Vertebrate Localities in the Lug River Basin (Leningrad Region, Russia: Guidebook for the Field Trip. Department of Palaeontology, Saint Petersburg University, St. Petersburg.
Jarvik, E. 1948. On the morphology and taxonomy of the Middle Devonian osteolepid fishes of Scotland. Kungliga Svenska Vetenskapsakademiens Handlinger, 25:1-301.
Jarvik E. 1950. Middle Devonian vertebrates from Canning Land and Wegeners Halvo (East Greenland). Part II. Crossopterygii. Meddelelser om Grønland, 96:1-132.
Jarvik E. 1985. Devonian osteolepiform fishes from East Greenland. Meddelelser om Gronland, Geoscience, 13:1-52.
Karatajūtė-Talimaa, V. 1963. Genus Asterolepis from the Devonian of the Russian Platform, p. 65-169. In Grigelis, A. and Karatajūte-Talimaa, V. (eds.). The Data of Geology of the Lithuania. Academy of Sciences of the Lithuanian SSR, Vilnius. [In Russian, with English summary].
Lyarskaya L. 1981. Baltic Devonian Placodermi. Asterolepididae. Zinātne, Rīga. (In Russian with English summary)
Marshall, J.E.A. and Astin, T.R. 1996. An ecological control on the distribution of the Devonian fish Asterolepis. Newsletters on Stratigraphy, 33:133-144. https://doi.org/10.1127/nos/33/1995/133
Michie, U.M., Newman, M.J., and den Blaauwen, J.L. 2015. The vertebrate biostratigraphy of the Rousay sequence in the Middle Devonian of Orkney, Scotland. Scottish Journal of Geology, 51:149-156. https://doi.org/10.1144/sjg2014-011
Miller, H. 1858. Cruise of the Betsey. Thomas Constable & Co., Edinburgh. https://doi.org/10.5962/bhl.title.30934
Moloshnikov, S.V. 2008. Devonian Antiarchs (Pisces, Antiarchi) from Central and Southern European Russia. Paleontological Journal, 42:691-773. https://doi.org/10.1134/s0031030108070010
Murphy, M.A., Morgan, T.G., and Dineley, D.L. 1976. Asterolepis sp. from the Upper Devonian of Central Nevada, Journal of Paleontology, 50:467-471.
Mykura, W. 1976. British Regional Geology. Orkney and Shetland. HMSO, Edinburgh.
Newman, M.J., Burrow, C.J., and den Blaauwen, J.L. 2019. The Givetian vertebrate fauna from the Fiskekløfta Member (Mimerdalen Subgroup), Svalbard. Part I. Stratigraphic and faunal review. Part II. Acanthodii. Norwegian Journal of Geology, 99:1-16. https://doi.org/10.17850/njg99-1-01
Newman, M.J. and den Blaauwen, J.L. 2018. A redescription of the endemic antiarch placoderm Asterolepis thule from the Middle Devonian (Givetian) of Shetland and its biostratigraphical horizon. Scottish Journal of Geology, 54:69-75. https://doi.org/10.1144/sjg2018-005
Newman, M.J., den Blaauwen, J.L., and Tuuling, T. 2017. Middle Devonian coccosteid (Arthrodira, Placodermi) biostratigraphy of Scotland and Estonia. Scottish Journal of Geology, 53:63-69. https://doi.org/10.1144/sjg-2016-012
Newman, M.J., Mark-Kurik, E., den Blaauwen, J.L., and Zupiņš, I. 2015. Scottish Middle Devonian fishes in Estonia. Scottish Journal of Geology, 51:141-147. https://doi.org/10.1144/sjg2014-006
Newman, M.J. and Trewin, N.H. 2008. Discovery of the arthrodire genus Actinolepis (class Placodermi) in the Middle Devonian of Scotland. Scottish Journal of Geology, 44:83-88. https://doi.org/10.1144/sjg44010083
Nilsson, T. 1941. The Downtonian and Devonian vertebrates of Spitsbergen. VII. Order Antiarchi. Skrifter om Svalbard og Ishavet, 82:1-54.
Obruchev, D.V. 1933. Description of four new fish species of the Leningrad Devonian. Materials of TsNIGRI: Paleontology and Stratigraphy, 1:12-15. (In Russian)
Pander, C.H. 1857. Über die Placodermen des Devonischen Systems. Buchdruckerei der Kaiserlichen Akademie der Wissenschaften, St. Petersburg.
Pander, C.H. 1860. Über die Saurodipterinen, Dendrodipterinen, Glyptopiden und Cheirolepiden des Devonischen Systems. Buchdruckerei der Kaiserlichen Akademie der Wissenschaften, St. Petersburg.
Paton, R. L. 1981. A catalogue of fossil vertebrates on the Royal Scottish Museum, Edinburgh. Placodermi. Royal Scottish Museum Information Series, Geology, 6:1-70.
Piepjohn, K. and Dallmann, W.K. 2014. Stratigraphy of the uppermost Old Red Sandstone of Svalbard (Mimerdalen Subgroup): Polar Research, 33:1-22. https://doi.org/10.3402/polar.v33.19998
Reed, J. W. 1979. The Devonian Fish Fauna of Red Hill, Nevada. Unpublished Ph.D. Thesis, University of California, Berkeley, California.
Rohon, J.V. 1900. Die Devonischen Fische von Timan in Russland. Sitzungsberichte der Königliche Böhmischen Gesellschaft der Wissenschaften. Mathematisch naturwissenschaltliche Classe (1899), 8:3-77.
Schultze, H.-P. 2010. The late Middle Devonian fauna of Red Hill I, Nevada, and its paleobiogeographic implications. Fossil Record, 13:285-295. https://doi.org/10.5194/fr-13-285-2010
Stensiö, E. 1918. Zur Kenntnis des Devons und des Kulms an der Klaas Billenbay, Spitsbergen. Bulletin of the Geological Institution of the University of Uppsala, 16:65-80.
Stensiö, E.A. 1931. Upper Devonian vertebrates from East Greenland. Meddelelser om Grønland, 86:1-212.
Stensiö, E.A. 1938. Placodermi, p. 8-34. In Stensiö, E.A. and Säve-Söderbergh, G. Middle Devonian vertebrate from Canning Land and Wegener Peninsula (East Greenland). Meddelelser om Grønland, 96:1-38.
Traquair, R.H. 1888. On the structure and classification of the Asterolepididae. Annals and Magazine of Natural History Sixth Series, 2:485-504. https://doi.org/10.1080/00222938809487519
Traquair, R.H. 1894-1914. A monograph of the fossil fishes in the Old Red Sandstone of Britain, Part 2, The Asterolepidae. Palaeontographical Society [Monographs], 48(229):63-130. https://doi.org/10.1080/02693445.1894.12028032
Traquair, R.H. 1908. On fossil fish remains collected by J.S. Flett M.A., D. Sc., from the Old Red Sandstone of Shetland. Transactions of the Royal Society of Edinburgh, 46:321-329. https://doi.org/10.1017/s0080456800002787
Upeniece, I. 2001. The unique fossil assemblage from the Lode Quarry (Upper Devonian, Latvia). Mitteilungen aus dem Museum fur Naturkunde in Berlin. Geowissenschaftliche Reihe, 4:101-119. https://doi.org/10.5194/fr-4-101-2001
Upeniece, I. 2011. Palaeoecology and juvenile individuals of the Devonian placoderm and acanthodian fishes from Lode site, Latvia. Disertationes Geologicae Universitas Latviensis, 21:1-221.
Watson, D.M.S. 1932. On three new species of fish from the Old Red Sandstone of Orkney and Shetland. Summary of Progress of the Geological Survey, U.K. for 1931, 2:157-166.
Watson, D.M.S. 1935. Fossil fishes of the Orcadian Old Red Sandstone, p. 157-169. In Edwards, W., Knox, J., Jones, R.C.B., and Stephens, J.V. The Geology of the Orkneys. Memoirs of the Geological Survey, Scotland.
Wells, J.W. 1964. The antiarch Asterolepis in the Upper Devonian of New York. Journal of Paleontology, 38:492-495.
Westoll, T.S. 1937. On a specimen of Eusthenopteron from the Old Red Sandstone of Scotland. Geological Magazine, 74:507-524. https://doi.org/10.1017/s0016756800089986
Woodward, A.S. 1891a. Catalogue of the Fossil Fishes in the British Museum (Natural History). Part 2. British Museum (Natural History), London. https://doi.org/10.5962/bhl.title.61854
Woodward, A.S. 1891b. The Devonian Fish-Fauna of Spitzbergen. Annals and Magazine of Natural History Sixth Series, 8:1-15. https://doi.org/10.1080/00222939109460384
