

 New remains of Liopleurodon (Reptilia, Plesiosauria) from the Middle Jurassic of western France and paedomorphosis within pliosaurids
New remains of Liopleurodon (Reptilia, Plesiosauria) from the Middle Jurassic of western France and paedomorphosis within pliosaurids
Article number: 27.2.a34
https://doi.org/10.26879/1280
Copyright Paleontological Society, July 2024
Author biographies
Plain-language and multi-lingual abstracts
PDF version
Appendix
Submission: 13 March 2023. Acceptance: 18 June 2024.
ABSTRACT
A new specimen of the pliosaurid Liopleurodon ferox from the Callovian strata (Middle Jurassic) of France is described. This new specimen represents one of the most-complete post-cranial skeletons known for this species. Remains of Liopleurodon ferox were previously reported from central and southern England, Germany, the North of France, and possibly from Poland and Russia. The specimen examined here therefore extends the biogeographical area of the species into central France and represents the southernmost occurrence of this taxon. Based on osteological evidence, the specimen is here identified as an “adult” individual, confirming that the lack of fusion between the cervical neural arches and associated centra is not a reliable indicator of a juvenile condition in pliosaurids. Instead, such a lack of fusion in cervical vertebrae is more likely to be paedomorphic in adult pliosaurids, as noted in previous works. However, this statement does not hold true for dorsal and caudal vertebrae, or cervical ribs, in which the pattern of suture closure may vary according to taxa. Overall, our results indicate that the open or closed conditions of the neurocentral sutures of all the vertebrae of the vertebral column, as well as the suture between ribs and centra, is a poor indicator of ontogenetic stages in pliosaurids.
Peggy Vincent. CR2P, CNRS-MNHN-Sorbonne Université, CP 38, 57 rue Cuvier, F-75005, Paris, France. peggy.vincent@mnhn.fr
Didier Poncet. Communauté de Communes du Thouarsais, Pôle Aménagement durable du territoire, 7 rue Anne Desrays, 79100 Thouars, France. didier.poncet@thouars-communaute.fr
Anthony Rard. Communauté de Communes du Thouarsais, Pôle Aménagement durable du territoire, 7 rue Anne Desrays, 79100 Thouars, France. anthony.rard@thouars-communaute.fr
Jean-Pierre Robin. 13, rue du Pas, 86120 Beuxes, France. jean-pierre.robin9@wanadoo.fr
Rémi Allemand. Department of Anthropology, University of Toronto Scarborough, Toronto, ON, Canada. remi.allemand@gmail.com
Keywords: Callovian; Jurassic; Plesiosauria; paedomorphic character; France
Final citation: Vincent, Peggy, Poncet, Didier, Rard, Anthony, Robin, Jean-Pierre, and Allemand, Rémi. 2024. New remains of Liopleurodon (Reptilia, Plesiosauria) from the Middle Jurassic of western France and paedomorphosis within pliosaurids. Palaeontologia Electronica, 27(2):a34.
https://doi.org/10.26879/1280
palaeo-electronica.org/content/2024/5257-jurassic-liopleurodon-of-france
Copyright: July 2024 Paleontological Society.
This is an open access article distributed under the terms of Attribution-NonCommercial-ShareAlike 4.0 International (CC BY-NC-SA 4.0), which permits users to copy and redistribute the material in any medium or format, provided it is not used for commercial purposes and the original author and source are credited, with indications if any changes are made.
creativecommons.org/licenses/by-nc-sa/4.0/
INTRODUCTION
Pliosaurids (Sauropterygia, Plesiosauria, Pliosauridae) were among the largest predators of Mesozoic aquatic ecosystems and ranged stratigraphically from the uppermost Triassic (Rhaetian) or lowermost Jurassic (Hettangian), to the early Late Cretaceous (e.g., Benson et al., 2011; Schumacher et al., 2013; Bardet et al., 2014; Benson and Druckenmiller, 2014; Wintrich et al., 2017). They colonized both marine and freshwater environments (e.g., Benson et al., 2013a; O’Gormann et al., 2018) and achieved a cosmopolitan distribution during the Late Jurassic (Barrientos-Lara et al., 2015). In Europe, pliosaurid remains have been mainly reported from England, France, Germany, Poland, Switzerland, and Norway (Bardet et al., 2014). They are particularly abundant in the Upper Jurassic (Callovian) strata of both the Oxford and Kimmeridge Clay formations exposed in England (e.g., Owen, 1869; Tarlo, 1960; Ketchum and Benson, 2011; Benson et al., 2013b; Martill et al., 2023), but are comparatively scarce elsewhere. In France, pliosaurid occurrences have been reported from the Lower, Middle, and Upper Jurassic strata of the north-east of the country: Normandy and Hauts-de-France (Beckary, 1992; Bardet, 1993; Bardet et al., 1993a, 1993b; Maréchal et al., 1999; Vincent et al., 2013), and from Lower Cretaceous strata of eastern France (Buffetaut et al., 2005). However, the preserved material from France consists mostly of isolated teeth and/or partial post-cranial remains, and only a few specimens preserve skull elements (e.g., Pliosaurus, Liopleurodon; Bardet, 1993). The main goal of this contribution is to describe a pliosaurid specimen from the Callovian of France preserving an almost complete post-cranial skeleton and to assess its systematic status as well as its phylogenetic affinities. In addition, the geographical occurrence and ontogenetic status of the specimen are discussed. The analysis of neurocentral sutures closure in the cervical vertebrae of pliosaurids, previously addressed by various authors (e.g., Ketchum and Benson, 2022; Araújo and Smith, 2023), has been expanded to encompass the entire vertebral column, as well as the sutures between cervical ribs and centra.
Repositories and Institutional Abbreviations
BEDFM, Bedford Museum, Bedford, UK; BHN, Musée d´Histoire Naturelle, Boulogne-sur-Mer, France; BRSMG, Bristol City Museum and Art Gallery, Bristol, UK; CAMSM, Sedgwick Museum of Earth Sciences, University of Cambridge, Cambridge, UK; CM, Carnegie Museum, Pittsburgh, Pennsylvania, USA; CMN, Canadian Museum of Nature, Ottawa, Ontario, Canada; ComCom Thouarsais, Community of Communes of Thouarsais, Thouars, France; FHSM, The Sternberg Museum of Natural History, Fort Hays, Kansas, USA; GPIT, Institut und Museum für Geologie und Paläontologie der Universität Tübingen, Tübingen, Germany; HAUF, UrweltMuseum Hauff, Holzmaden, Germany; IGM, Colección Nacional de Paleontología, Instituto de Geología of the Universidad Nacional Autónoma de México, Mexico city, Mexico; MAE, Musée de l’Agglomération d’Elbeuff, Elbeuf-sur-Seine, France; MANCH, The Manchester Museum, Manchester, UK; MCZ, Museum of Comparative Zoology, Harvard University, Cambridge, Massachusetts, USA; MJACM, Museo El Fósil, Vereda Monquirá, Municipio de Villa de Leyva, Departamento de Boyacá, Colombia; MNA V, Museum of Northern Arizona, Flagstaff, USA; MNHNCu, Museo Nacional de Historia Natural, Havana, Cuba; MP, VL, Paleontological collection of the Servicio Geológico Colombiano, Bogotá D.C., Colombia; MUHNCAL, Museo de Historia Natural y Cultural del Desierto de Atacama, Calama, Chile; NAMU, Naturkunde-Museum Bielefeld, Bielefeld, Germany; NHMUK, Natural History Museum, London, UK; OUMNH, Oxford University Museum of Natural History, Oxford, UK; PETMG, Peterborough City Museum and Art Gallery, Peterborough, UK; PMO, University of Oslo Natural History Museum, Oslo, Norway; QM, Queensland Museum, Brisbane, Ausralia; SDSM, South Dakota School of Mines and Technology, Rapid City, South Dakota, USA; UANL-FCT, Universidad Autonóma de Nuevo León, Linares N.L., Mexico; YKM: Ulyanovskii Oblastnoi Kraevedcheskii Musei I.A. Goncharova, Ulyanovsk, Ulyanovsk Region, Russia.
GEOGRAPHICAL AND STRATIGRAPHICAL SETTING
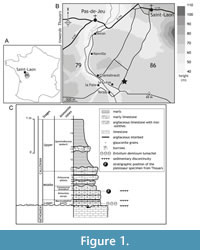 The specimen described herein was discovered on the municipality of Saint-Laon (Vienne, France), located in the Dive du Nord valley on the southwest margin of the Paris Basin (Dhoste et al., 1987). The deposits correspond to the Jurassic series extending from Poitiers to the region of Thouars (Figure 1A-B). The fossil remains come from a limestone quarry (the Grève Quarry) that was exploited for construction materials. In the 1970s and 1980s, the Grève Quarry was well known among fossil collectors (Saint-Léger, 1987), and provided abundant and diverse ammonites. In the past 30 years, this quarry has been abandoned and inaccessible, preventing the direct observation of the formerly exploited levels.
The specimen described herein was discovered on the municipality of Saint-Laon (Vienne, France), located in the Dive du Nord valley on the southwest margin of the Paris Basin (Dhoste et al., 1987). The deposits correspond to the Jurassic series extending from Poitiers to the region of Thouars (Figure 1A-B). The fossil remains come from a limestone quarry (the Grève Quarry) that was exploited for construction materials. In the 1970s and 1980s, the Grève Quarry was well known among fossil collectors (Saint-Léger, 1987), and provided abundant and diverse ammonites. In the past 30 years, this quarry has been abandoned and inaccessible, preventing the direct observation of the formerly exploited levels.
The sedimentary succession of the Grève Quarry was described by Legendre (1984, p. 98) and Cariou (in Gabilly et al., 1985, p. 45-47). The whole succession was about 4.50 m thick and divided into two formations corresponding to the Bathonian and the Callovian stages, which are separated by a hard and perforated surface, coated with iron oxides (Figure 1C). The Bathonian succession (2 m) corresponds to whitish, nodular, chalky arenaceous limestones and contain sponges, bivalves, and scleractinian corals of the genus Montlivaltia (Lamouroux, 1821). The Callovian succession (2.5 m) includes argillaceous limestones with ferruginous ooids and marls, locally known as “calcaires en platins”. The Callovian succession can be subdivided into three units: (i) a lower unit (beds 1 to 4) consisting of brown to red limestone with abundant and small ferruginous ooids, and characterized by a distinctive lumachel (i.e., shell beds containing partial to complete bivalve fossils) of Entolium demissum (Phillips, 1829); (ii) a middle unit (beds 5 to 11) corresponding to blue-grey limestones with glauconitic grains, containing less numerous and much larger ferruginous ooids; (iii) an upper unit (beds 12 to 14) corresponding to grey marls that present a discontinuous decimetric level of bioturbated hard limestone ("miches") and contain limited fossil remains of sponges and belemnites.
The stratigraphic level from which the pliosaur specimen came was not accurately located during its extraction. However, the identification of ammonites, i.e., Hecticoceras (Rossiensiceras) savoiense Zeiss, 1956, Rehmannia (Loczyceras) discrepans (Bourquin, 1968), Reineckeia (Reineckeia) anceps (Reinecke, 1818) forme corroyi, and Oecoptychus refractus (Reinecke, 1818), during the preparation of the material, indicates a possible provenance from bed 4 of the Callovian succession. Such a hypothesis about the stratigraphic position of the specimen is reinforced by the superficial erosion observed on some of the vertebrae, which is like the erosional facies that were visible on the top of the bed 4. The specimen was thus possibly partly embedded in the argillaceous limestones with ferruginous ooids of the Erymnoceras coronatum Zone, more precisely at the boundary between the Leuthardi sub-zone and the Rota sub-zone (Cariou, pers. com. 2006). The abundant pelagic fauna of these Callovian deposits indicates the strata were deposited over an area of shallow waters open to the oceanic domain (Gabilly and Cariou, 1974; Cariou, 1980) in a sublittoral environment (Faugeras, 1985; Legendre et al., 1989).
MATERIAL AND METHODS
The ComCom Thouarsais_Geol.0121 specimen examined here, informally called the ‘Thouarsais specimen’, is a partial post-cranial skeleton discovered in April 1979 by the fossil enthusiasts Camille Robin and Jean-Pierre Robin. They used a pickaxe and a crowbar to clear the overburden including a Quaternary cover (about 0.8 m thick) over an area of 25 to 30 m². The discoverers were concerned about the conservation of the specimen and donated it to the Community of Communes of the Thouarsais (Communauté de Communes du Thouarsais, Thouars, Deux-Sèvres, France) in 2001. In return, the latter provided the means to extract the skeleton from its limestone matrix and to exhibit it to the public. The preparation was performed out in Thouars in 2006. The removal of the calcareous matrix was first achieved by using a pneumatic micro-hammer and followed by treatment with 10% methanoic acid to precisely finalise the preparation. This was done after embedding the exposed bones with plexigum®, a resin utilized for consolidating, repairing, and reassembling fractured elements such as ribs and vertebrae. The specimen was presented to the public during a temporary exhibition for the “European Heritage Days” in Thouars, and then for the “Science Festival” in 2006. The Thouarsais specimen is currently kept in the paleontological collections of the Community of Communes of the Thouarsais.
To establish the phylogenetic relationships of the specimen within Plesiosauria, a cladistic analysis was performed using the data matrix of Sachs et al. (2023), one of the most recent cladistic data matrix of Plesiosauria including the greatest number of pliosaurid taxa. The data (see Appendix 1) for all plesiosaur taxa were retained except for Attenborosaurus conybeari (Sollas, 1881), as we consider that its coding requires revision, given that it is one of the few genera from Toarcian strata whose anatomy has not yet been revised. Data for the Thouarsais specimen were added to the data matrix. The character 172 (the shape of neurocentral suture in anterior-middle cervical vertebrae in lateral view) was not included in the analysis. Indeed, although an open neurocentral suture is considered as a peadomorphic feature in pliosaurid taxa (refer to the Ontogenetic stage paragraph), associated changes in the shape of the suture could potentially be linked to their ontogenetic status, as previously reported in some plesiosauroid taxa (Vincent et al., 2017). The data set, including 129 taxa and scored for 269 characters, was analyzed using TNT 1.6 (Goloboff et al., 2008; Goloboff and Morales, 2023). A Traditional Search (1000 replicates) using tree bisection-reconnection (TBR) branch-swapping with all characters unordered and unweighted was performed. Bremer support values were calculated by using the bremer.run script in TNT (Goloboff et al., 2008).
SYSTEMATIC PALAEONTOLOGY
Superorder SAUROPTERYGIA Owen, 1860
Order PLESIOSAURIA de Blainville, 1835
Family PLIOSAURIDAE Seeley, 1874
Clade THALASSOPHONEA Benson and Druckenmiller, 2014
Genus LIOPLEURODON Sauvage, 1873
Type species. Liopleurodon ferox Sauvage, 1873.
Liopleurodon ferox Sauvage, 1873
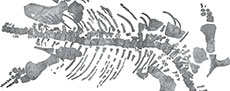
Figure 2, Figure 3, Figure 4, Figure 5, Figure 6, Figure 7, Figure 8
Type material. BHN 3R 197, a single tooth now kept in the Musée d’histoire naturelle de Lille (Lille, France).
Referred material. See Noè, 2001; ComCom Thouarsais_Geol.0121 (described herein).
Type locality and horizon. Le Wast, near Boulogne-sur-Mer (northern France), Quenstedtoceras lamberti Zone, Oxford Clay Formation, Upper Callovian (Sauvage, 1873; Noè, 2001).
Other localities. Bathonian-Callovian boundary to Lower Oxfordian of North and Central France, England, Switzerland and Germany. Possibly from the Lower Bathonian of France, Callovian of Poland, Callovian and Oxfordian of Russia, and Lower Tithonian of Germany (see Noè (2001) for details; Zverkov et al., 2024).
Diagnosis. Andrews, 1913; Tarlo, 1960 (in part); for the cranial diagnostic characters see Noè (2001). Pliosaurid possessing short cervical centra (length less than half width or height); cervical centra with a ventral lip and no ventral keel; two cervical rib facets.
Description
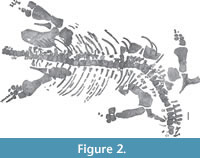 Preservation. The Thouarsais specimen measures around 320 cm in length and consists of a sub-complete post-cranial skeleton of a pliosaurid plesiosaur (Figure 2), including 45 vertebrae (10 cervicals, 4 pectorals, 22 dorsals, 4 sacrals, and 4 caudals), remains of ribs and gastralia, partial pectoral and pelvic girdles, as well as partial front- and hind-limbs. For convenience, the vertebrae will be described by using a letter (i.e., C: cervical, P: pectoral, D: dorsal, S: sacral and CA: caudal) followed by an Arabic number (e.g., C1 for the first cervical vertebra). This system does not correspond to the anatomical positions of the vertebrae.
Preservation. The Thouarsais specimen measures around 320 cm in length and consists of a sub-complete post-cranial skeleton of a pliosaurid plesiosaur (Figure 2), including 45 vertebrae (10 cervicals, 4 pectorals, 22 dorsals, 4 sacrals, and 4 caudals), remains of ribs and gastralia, partial pectoral and pelvic girdles, as well as partial front- and hind-limbs. For convenience, the vertebrae will be described by using a letter (i.e., C: cervical, P: pectoral, D: dorsal, S: sacral and CA: caudal) followed by an Arabic number (e.g., C1 for the first cervical vertebra). This system does not correspond to the anatomical positions of the vertebrae.
Cervical vertebrae. Ten cervical centra represent middle to posterior elements of the neck (Figure 3). They exhibit gently concave articular surfaces with a central depressed notochordal pit, as in Peloneustes and NHMUK R. 3536, a large specimen of Liopleurodon (Andrews, 1913; Tarlo, 1960; Ketchum, 2007). In anterior view, the centra are subcircular in outline, slightly heart-shaped with a slight ventral lip in the cervicals C1 to C8 (Figure 3A, C, H). The presence of a ventral lip is reported in Marmornectes, Liopleurodon, Peloneustes, ‘Pliosaurus’ andrewsi, Eardasaurus, and Simolestes (Andrews, 1913; Tarlo, 1960; Ketchum and Benson, 2011, 2022) and may be a synapomorphy of a derived clade within Pliosauridae (Ketchum and Benson, 2011). The cervical centra are wider than they are high and long (W>H>L), as in other pliosaurids (e.g., Andrews, 1913; Tarlo, 1960; O’Keefe, 2001; Ketchum and Benson, 2011). Centrum length represents approximately 1/2 to 3/5 of both the width and height of the vertebrae; similar proportions (length being less than half the vertical diameter of the articular faces) are given by Andrews (1913) for Liopleurodon ferox. The ventral surfaces of the centra are pierced by two small subcentral foramina (Figure 3I). Similar to the pliosaurids Marmornectes, Pliosaurus westburyensis, Pliosaurus carpenter, and Liopleurodon ferox, these foramina are not separated from one another by a ridge and differ from the condition reported in Peloneustes, Pliosaurus brachydeirus, and the pliosaurid OUMNH J.02247 (Andrews, 1913; Tarlo, 1960 ; Ketchum and Benson, 2011; Benson et al., 2013b). The cervical rib facets are large and located on the ventral to ventrolateral surfaces of the centra (Figure 3B-E). The centra of cervicals C1 to C8 bear two rib facets (diapophysis and parapophysis) subequal in size (Figure 3B, E, F). The diapophysis is triangular with the apex pointing dorsally whereas the parapophysis exhibits a bowl shape with a rounded ventral margin. The two facets, taken together, are pear-shaped like those described by Andrews (1913) for Liopleurodon ferox. The cervical centra C9 and C10 bear a single oval, dorsoventrally elongated rib facet (Figure 3G), similar to Liopleurodon ferox (Andrews, 1913). Four cervical neural arches are sub-complete and still attached to their centra showing a V-shaped neurocentral suture (C3-C6; Figure 3D-E). In anterior view, the neural canals exhibit an ovoid keyhole-shape due to a bony bulge located at mid-height on the medial surface of the neural arches, as in Marmornectes (Ketchum and Benson, 2011) and Liopleurodon ferox (Andrews, 1913). The neural canals represent approximately 50% of the centra height (Figure 3C). The preserved neural spines are straight and sub-rectangular. The neural spines are longer antero-posteriorly than they are wide, as in Liopleurodon but unlike the condition observed in Marmornectes (Andrews, 1913; Ketchum and Benson, 2011), and measure approximately half of the centra length. The tips of the neural arches widen as in Liopleurodon ferox (Andrews, 1913), showing a teardrop to oval outline in dorsal view and appearing sub-rectangular in lateral view. The zygapophyses are narrower than the centra. They are angled at approximately 45 degrees from the horizontal and are separated from one another, the right and left pre- and postzygapophyses being not fused medially. The sutures between cervical centra and associated neural arches, as well as the sutures between cervical centra and the corresponding ribs, are visible on the surface of the bones.
; Ketchum and Benson, 2011; Benson et al., 2013b). The cervical rib facets are large and located on the ventral to ventrolateral surfaces of the centra (Figure 3B-E). The centra of cervicals C1 to C8 bear two rib facets (diapophysis and parapophysis) subequal in size (Figure 3B, E, F). The diapophysis is triangular with the apex pointing dorsally whereas the parapophysis exhibits a bowl shape with a rounded ventral margin. The two facets, taken together, are pear-shaped like those described by Andrews (1913) for Liopleurodon ferox. The cervical centra C9 and C10 bear a single oval, dorsoventrally elongated rib facet (Figure 3G), similar to Liopleurodon ferox (Andrews, 1913). Four cervical neural arches are sub-complete and still attached to their centra showing a V-shaped neurocentral suture (C3-C6; Figure 3D-E). In anterior view, the neural canals exhibit an ovoid keyhole-shape due to a bony bulge located at mid-height on the medial surface of the neural arches, as in Marmornectes (Ketchum and Benson, 2011) and Liopleurodon ferox (Andrews, 1913). The neural canals represent approximately 50% of the centra height (Figure 3C). The preserved neural spines are straight and sub-rectangular. The neural spines are longer antero-posteriorly than they are wide, as in Liopleurodon but unlike the condition observed in Marmornectes (Andrews, 1913; Ketchum and Benson, 2011), and measure approximately half of the centra length. The tips of the neural arches widen as in Liopleurodon ferox (Andrews, 1913), showing a teardrop to oval outline in dorsal view and appearing sub-rectangular in lateral view. The zygapophyses are narrower than the centra. They are angled at approximately 45 degrees from the horizontal and are separated from one another, the right and left pre- and postzygapophyses being not fused medially. The sutures between cervical centra and associated neural arches, as well as the sutures between cervical centra and the corresponding ribs, are visible on the surface of the bones.
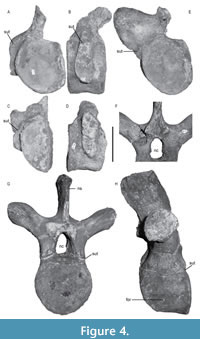 Pectoral vertebrae. Four pectoral vertebrae were identified based on rib facets that are present on both the centrum and the neural arch (Figure 4A-D). The pectoral centra are higher than long, but longer than the cervical centra. On the two first pectorals (P1 and P2), the rib facets show an open neurocentral suture (Figure 4B, D) and appear elongated dorsoventrally (Figure 4B, D). This differs from the last pectoral (P4), in which the rib facet is more rounded (Figure 4E). The subcentral foramen on the right side of the P4 centrum is situated more laterally than in other pectorals, and a larger supplementary foramen is present ventrally.
Pectoral vertebrae. Four pectoral vertebrae were identified based on rib facets that are present on both the centrum and the neural arch (Figure 4A-D). The pectoral centra are higher than long, but longer than the cervical centra. On the two first pectorals (P1 and P2), the rib facets show an open neurocentral suture (Figure 4B, D) and appear elongated dorsoventrally (Figure 4B, D). This differs from the last pectoral (P4), in which the rib facet is more rounded (Figure 4E). The subcentral foramen on the right side of the P4 centrum is situated more laterally than in other pectorals, and a larger supplementary foramen is present ventrally.
Dorsal vertebrae. Centra of the 22 dorsal vertebrae are as high as wide, shorter than high, and bear a central notochordal pit (Figure 4F-H). Both the lateral and ventral surfaces of the centra are concave. The articular surfaces of the centra are sub-oval and weakly concave. The centrum width is approximately half of the transverse process width. The rib facets are situated on short and robust transverse processes. Only one complete neural arch and neural spine is preserved in association with the centrum (Figure 4G, H). The zygapophyses are relatively short and proportionally smaller than those on the cervical neural arches. The neural spines extend antero-posteriorly, almost as long as the centrum length (around 70%), and higher than the associated centrum. The thin neural spines are inclined postero-dorsally and present a slightly curved anterior margin in lateral view (Figure 4H). The apices of the neural spines are convex in their centra and rectangular in cross-section. The posterior margin of the neural spine is vertically grooved, and the postzygapophyses are thus separated by deep medial fossae (observable on D6 and D19; Figure 4F).
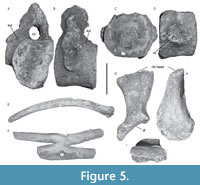 Sacral vertebrae. The centra of the four sacral vertebrae (Figure 5A-B) show an open neurocentral suture and bear a dorso-ventrally elongated rib facet, which is formed by both the neural arch and the centrum (Figure 5B). The neural spines are inclined postero-dorsally, the apices being strongly expanded and oval in dorsal view.
Sacral vertebrae. The centra of the four sacral vertebrae (Figure 5A-B) show an open neurocentral suture and bear a dorso-ventrally elongated rib facet, which is formed by both the neural arch and the centrum (Figure 5B). The neural spines are inclined postero-dorsally, the apices being strongly expanded and oval in dorsal view.
Caudal vertebrae. Five caudal vertebral centra are preserved. The articular surfaces are nearly circular in outline and are slightly concave with a pronounced median pit (Figure 5C-D). The caudal rib facets are simple. The chevron facets are located ventrally on the posterior edges of the centra. No neural arches have been preserved.
Ribs. The double-headed cervical ribs are short with a convex dorsal surface and separated by a wide aperture (Figure 3E). Proximally, the width of the aperture (Figure 3C) is much greater than that reported for Marmornectes (Ketchum and Benson, 2011), but similar to that reported for CAMSM J.46912 (holotype of Liopleurodon pachydeirus Seeley, 1869, considered con-specific with Liopleurodon ferox by Noè, 2001). The distal end of the complete ribs preserved on cervical vertebrae C3 and C6 (Figure 3E) shows an undeveloped anterior process, the short posterior process being rounded and flattened as in most pliosauroids (e.g., Peloneustes, Liopleurodon, Simolestes; Ketchum and Benson, 2011). Numerous fragments of elongated and curved dorsal ribs are preserved (Figure 5E). The ribs have slightly concave articulatory surfaces. Differentiating gastralia from dorsal ribs among the various broken isolated elements has proved difficult. One element is double-forked (Figure 5F) and is identified as a gastralium, a morphology already observed for gastralia in the Triassic sauropterygians Corosaurus (Storrs, 1991) and Nothosaurus (Koken, 1893), the plesiosauroids Plesiosaurus and Brancasaurus (Storrs, 1997; Sachs et al., 2016), both juvenile and adult specimens of the elasmosaurids Mauisaurus, Terminonatator, and Fluvionectes (Hector, 1874; Martin et al., 2007; Campbell et al., 2021), and in a single specimen of Peloneustes philarchus, BMNH R3318 (Ketchum, 2007). This character was not observed in the mounted specimen of Liopleurodon ferox GPIT-PV-30093 displayed in Tübingen (syn. PV 17998, GPIT/RE/3184, GPIT 1754/2; see Goller (2021) and Krahl et al. (2022)). This character is variously interpreted as an anomalous ossification (Sato, 2003) or a congenital deformity (Storrs, 1991), and considered rare (Martin et al., 2007), but it may be more widespread than initially thought.
Two sacral ribs are preserved (Figure 5G-I). Similar to other pliosaurids (e.g., Liopleurodon and Marmornectes), the distal end of the rib is dorsoventrally flattened and flared (Andrews, 1913; Ketchum and Benson, 2011). The proximal end has an elongated oval outline (Figure 5I). The rib shaft is twisted as in Marmornectes (Ketchum and Benson, 2011), so that the long axis of the proximal rib facet and the long axis of the distal ilium facet are strongly rotated relative to each other (~90°). A ridge extends dorsoventrally from the rib head to the flared distal end as in Peloneustes (Ketchum, 2007).
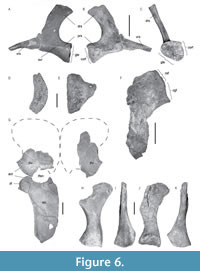 Pectoral girdle. The pectoral girdle is partially preserved, with only remains of the scapulae and coracoids present, whereas the clavicular arch is missing. Other small and fragmentary elements, probably belonging to the pectoral girdle are preserved; however, they could not be identified and are omitted in this description. Only the posterior ramus of the right scapula is preserved, whereas the almost complete left scapula is triradiate, with the ventral, dorsal, and posterior rami visible (Figure 6A-C). The medial surface of the scapula between the posterior and ventral rami, at the level of the pectoral fenestra, is concave. The ventral ramus is incomplete and directed antero-medially, whereas the complete dorsal process extends postero-laterally. The latter shows an approximately subequal antero-posterior width along its entire dorso-ventral height except for the apex that is slightly enlarged. The dorsal process presents a convex and acute anterior edge, as well as a roundly concave posterior margin. The anterior edge is slightly sinusoidal, as in Marmornectes and Peloneustes (Ketchum and Benson, 2011) and Liopleurodon ferox GPIT-PV-30093 mounted specimen (Linder, 1913; pers. obs.). The apex of the dorsal ramus is a rugose concave, elongated, teardrop-shaped facet, indicating that it was probably covered by cartilage in the living individual, and undulates, as in Marmornectes (Ketchum and Benson, 2011). The posterior ramus is robust and bears facets for articulation with the coracoid and the humerus. A ridge extends from the tip of the glenoid facet of the scapula along its lateral surface, enhancing the angle between the ventral and lateral surfaces of the scapula.
Pectoral girdle. The pectoral girdle is partially preserved, with only remains of the scapulae and coracoids present, whereas the clavicular arch is missing. Other small and fragmentary elements, probably belonging to the pectoral girdle are preserved; however, they could not be identified and are omitted in this description. Only the posterior ramus of the right scapula is preserved, whereas the almost complete left scapula is triradiate, with the ventral, dorsal, and posterior rami visible (Figure 6A-C). The medial surface of the scapula between the posterior and ventral rami, at the level of the pectoral fenestra, is concave. The ventral ramus is incomplete and directed antero-medially, whereas the complete dorsal process extends postero-laterally. The latter shows an approximately subequal antero-posterior width along its entire dorso-ventral height except for the apex that is slightly enlarged. The dorsal process presents a convex and acute anterior edge, as well as a roundly concave posterior margin. The anterior edge is slightly sinusoidal, as in Marmornectes and Peloneustes (Ketchum and Benson, 2011) and Liopleurodon ferox GPIT-PV-30093 mounted specimen (Linder, 1913; pers. obs.). The apex of the dorsal ramus is a rugose concave, elongated, teardrop-shaped facet, indicating that it was probably covered by cartilage in the living individual, and undulates, as in Marmornectes (Ketchum and Benson, 2011). The posterior ramus is robust and bears facets for articulation with the coracoid and the humerus. A ridge extends from the tip of the glenoid facet of the scapula along its lateral surface, enhancing the angle between the ventral and lateral surfaces of the scapula.
A crushed portion of the right coracoid is preserved (Figure 6F). It presents two adjacent facets: the anterior one, triangular, corresponds to the scapular facet of the coracoid, while the posterior facet is concave and oval, corresponding to the glenoid facet of the coracoid. An element preserved separately (Figure 6D-E), showing an antero-posterior curvature and a thickening that culminates in the flattened medial margin, is identified as the anterior-most part of the robust right bar of the coracoid. The part of the symphysis where the coracoids meet is curved dorsally. The anterior process of the coracoid is very short with a subrectangular outline in dorsal view as in Liopleurodon ferox (Andrews, 1913).
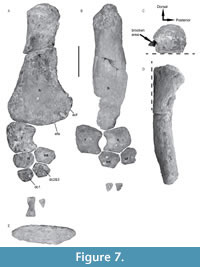 Forelimb. Both humeri are preserved, the left being almost complete (Figure 7A) whereas the right is severely damaged and lacks most of the proximal and distal parts (Figure 7B). The proximo-ventral part of the left humerus shows the tuberosity but is severely abraded, and the capitulum is not preserved (Figure 7C). The humeral shaft curves dorso-distally as in other pliosaurids (Figure 7D; Ketchum and Benson, 2011). Distally, the pre- and postaxial margins of the shaft are asymmetrical, similar to Liopleurodon ferox, Pliosaurus almanzaensis, and Peloneustes philarchus (Linder, 1913; Ketchum, 2007; O’Gorman et al., 2018). A prominent postaxial expansion is visible at the distal end and is much wider than in Pliosaurus funkei, Stenorhynchosaurus, and Marmornectes (Andrews, 1913; Ketchum and Benson, 2011; Knutsen et al., 2012; Páramo-Fonseca et al., 2016). The distal surface of the humerus is suboval in outline (Figure 7E). The radial and ulnar facets are distinct as in Liopleurodon ferox (Linder, 1913), contrasting with Marmornectes (although the propodials are abraded distally in the type and sole specimen of this taxon; Ketchum and Benson, 2011), and Stenorhynchosaurus, which both lack prominent articular surfaces for the epipodial elements (Páramo-Fonseca et al., 2016). Posterior to the ulna facet, a third facet is presumed to articulate with a postaxial accessory ossicle. This contrasts with Stenorhynchosaurus and Peloneustes in which the lack of postaxial ossicles in the epipodial row of the forelimb has been reported (Ketchum and Benson, 2011; Páramo-Fonseca et al., 2016).
Forelimb. Both humeri are preserved, the left being almost complete (Figure 7A) whereas the right is severely damaged and lacks most of the proximal and distal parts (Figure 7B). The proximo-ventral part of the left humerus shows the tuberosity but is severely abraded, and the capitulum is not preserved (Figure 7C). The humeral shaft curves dorso-distally as in other pliosaurids (Figure 7D; Ketchum and Benson, 2011). Distally, the pre- and postaxial margins of the shaft are asymmetrical, similar to Liopleurodon ferox, Pliosaurus almanzaensis, and Peloneustes philarchus (Linder, 1913; Ketchum, 2007; O’Gorman et al., 2018). A prominent postaxial expansion is visible at the distal end and is much wider than in Pliosaurus funkei, Stenorhynchosaurus, and Marmornectes (Andrews, 1913; Ketchum and Benson, 2011; Knutsen et al., 2012; Páramo-Fonseca et al., 2016). The distal surface of the humerus is suboval in outline (Figure 7E). The radial and ulnar facets are distinct as in Liopleurodon ferox (Linder, 1913), contrasting with Marmornectes (although the propodials are abraded distally in the type and sole specimen of this taxon; Ketchum and Benson, 2011), and Stenorhynchosaurus, which both lack prominent articular surfaces for the epipodial elements (Páramo-Fonseca et al., 2016). Posterior to the ulna facet, a third facet is presumed to articulate with a postaxial accessory ossicle. This contrasts with Stenorhynchosaurus and Peloneustes in which the lack of postaxial ossicles in the epipodial row of the forelimb has been reported (Ketchum and Benson, 2011; Páramo-Fonseca et al., 2016).
Both left and right radii (Figure 7A-B) form a compact and robust bone, longer (left radius: 92 mm) than wide (left radius: 87 mm), with concave postaxial borders and a slightly convex preaxial margin. The postaxial margin forms the preaxial margin of a large epipodial foramen that extends for 2/3 of the length of the epipodials. The humerus facet is extensive, being antero-posteriorly long and dorso-ventrally thick. Distally, the radius bears three facets. The middle one, being the largest, corresponds to the radial facet. Postaxially, a small but well-formed oblique facet contacts with the intermedium and appears relatively shorter than the one observed in Peloneustes philarchus (Ketchum, 2007). A third, very reduced and oblique facet, located preaxially to the radial facet, may have contacted a preaxial ossicle.
Only the right ulna is preserved (Figure 7B). It forms a pentagonal bone slightly wider (preaxially to postaxially) than long (proximally to distally), similar to the condition reported in other pliosauroids (Ketchum and Benson, 2011). The ulna (77 mm long; 95 mm wide) is shorter and wider than the radius, as in Simolestes vorax (Andrews, 1913). The ulna bears a straight proximal facet for the humerus, as observed in Simolestes vorax and Liopleurodon ferox (Andrews, 1913; Linder, 1913), but not for a postaxial accessory ossicle. The two distal facets articulate with the ulnare and the intermedium. The preaxial margin of the ulna forms the postaxial border of the epipodial foramen and is concave as in Liopleurodon ferox (Linder, 1913), but at a higher degree than in Marmornectes and Peloneustes (Andrews, 1913; Ketchum and Benson, 2011). The postaxial margin is almost straight and differs from the condition reported in Simolestes and Peloneustes (Andrews, 1913), but is similar to that observed in Liopleurodon ferox (Linder, 1913).
The forelimb intermedium (Figure 7A-B) is a large element (right intermedium: 67 mm wide; 47 mm long) and exhibits distinct facets for the radius, ulna, radiale, ulnare, distal carpals 2 and 3, and distal carpal 4. The intermedium participates in the epipodial foramen, but its involvement is minimal.
The left radiale is a dorsoventrally thick, sub-hexagonal element 62 mm wide and 46 mm long. It bears a long and thick proximal facet for union with the radius, and an anterior facet for the preaxial accessory ossicle (not preserved). Distally, the radiale contacts the distal carpal (dc) 1 through a long facet, and both the intermedium and dc2 and dc3 along its postaxial margin.
The right ulnare is the smallest of the three proximal carpals. It is rounded and contacts the ulna proximally as well as the intermedium medially. Its thickness reduces towards its anterior part to form a very thin edge marked by a profound notch on its extremity, as in Liopleurodon and Simolestes (Andrews, 1913; Linder, 1913).
The distal carpal (dc) 1 is rounded and articulates with the radiale and the dc2 and 3. The dc2 and 3 is sub-hexagonal and larger than dc1. Additional elements partially preserved may be identified as metacarpal and phalange remains. Only one of them is complete and appears proximodistally elongated and hourglass shaped.
Pelvic girdle. Both pubes are fragmentary (Figure 6G). The left pubis presents a gently concave acetabular facet that is longer than the ischial facet. Its posterior border, medial to the ischial facet, is concave, and forms the anterior border of the thyroid fenestra.
A large portion of the left ischium is preserved and forms a blade-like element, antero-posteriorly longer than wide (estimated length 560 mm), as in other pliosauroids (Tarlo, 1960; Ketchum and Benson, 2011) and polycotylids (Williston, 1903; O’Keefe and Carrano, 2005; Albright et al., 2007a), and which become thinner posteriorly. The dorsal surface of the bone is concave, and its anterior margin forms a broad and concave curve. On the anteromedial side, a forward projection comes to a broad point and forms the anterior end of the curved medial border of the element. Posteriorly, the shaft of the left ischium indicates that the two posterior parts of the ischia would probably diverge from one another, as in Simolestes but more pronounced than the divergence reported in Liopleurodon and Peloneustes (Andrews, 1913). The posterior-most part of the ischium is partially preserved. The thyroid fenestra seems to be more rounded than that of Peloneustes (Andrews, 1913; Ketchum, 2007).
Both ilia are partially preserved; the dorsal end of the left one as well as the ventral end of the right one are abraded. The ilia are elongate, left, and right measuring 258 mm and 256 mm, respectively. They are expanded at both extremities, the dorsal one (sacral end) more than the ventral (acetabular) one. The mid-shaft of the ilium is sub-oval in outline. The ventral end is approximately rotated about 90 degrees relative to the iliac blade and bears two facets, which are sub-equal in size: a posteromedial ischium facet and a more anterolateral facet contributing in the acetabulum margin. The ilium does not bear any ridge contra that reported for Peloneustes (Andrews, 1913; Ketchum, 2007).
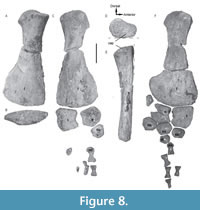 Hindlimb. Both femora are preserved (Figure 8) and appear relatively elongate with concave preaxial and postaxial margins. They are slightly longer (right femur: 480 mm long; left femur: 470 mm long) than the humeri, similar to Pliosaurus rossicus (Halstead, 1971) and Liopleurodon ferox (Linder, 1913), and as robust as the femora observed in other Callovian pliosauroids except Marmornectes (Ketchum and Benson, 2011). Both femora are fractured and abraded ventrally (Figure 8D-E). The proximal ends are poorly preserved, preventing the observation, if present, of the postaxial process and the rugosities. The posterodistal end of the right femur is only partially preserved (Figure 8A-B), whereas the distal portion of the left femur is well-preserved (Figure 8C). The latter is expanded and dorsoventrally flattened (Figure 8F) and bears two well-developed, but poorly distinctive, facets for contact with the epipodials. The facet for the tibia is slightly longer than the one for the fibula.
Hindlimb. Both femora are preserved (Figure 8) and appear relatively elongate with concave preaxial and postaxial margins. They are slightly longer (right femur: 480 mm long; left femur: 470 mm long) than the humeri, similar to Pliosaurus rossicus (Halstead, 1971) and Liopleurodon ferox (Linder, 1913), and as robust as the femora observed in other Callovian pliosauroids except Marmornectes (Ketchum and Benson, 2011). Both femora are fractured and abraded ventrally (Figure 8D-E). The proximal ends are poorly preserved, preventing the observation, if present, of the postaxial process and the rugosities. The posterodistal end of the right femur is only partially preserved (Figure 8A-B), whereas the distal portion of the left femur is well-preserved (Figure 8C). The latter is expanded and dorsoventrally flattened (Figure 8F) and bears two well-developed, but poorly distinctive, facets for contact with the epipodials. The facet for the tibia is slightly longer than the one for the fibula.
The tibia and fibula are morphologically similar to the radius and ulna, respectively, but longer (left tibia: 120 mm long; right tibia: 117 mm long; left fibula: 93 mm long; right fibula: 96 mm long) as is typical in pliosaurids (e.g., Andrews, 1913; Tarlo, 1960). The overall morphology of the tibia is similar to the one reported in Liopleurodon, Peloneustes, and Marmornectes (Andrews, 1913; Tarlo, 1960; Ketchum and Benson, 2011). The tibia (Figure 8B-C) is longer than wide, and its proximal facet for the femur is straight as in Peloneustes philarchus and Liopleurodon ferox (Andrews, 1913). The preaxial border is slightly convex as in Peloneustes and Liopleurodon ferox (Andrews, 1913; Ketchum, 2007), and the postaxial border is concave, forming the anterior margin of the epipodial foramen. The epipodial foramen is closed distally by contact between the tibia and fibula and is less extensive than that observed in the forelimb. The tibia shows a proximal facet for contact with the fibula, and a short and well-developed distal facet for the intermedium as in Peloneustes, Liopleurodon, and other pliosauroids (Andrews, 1913; Tarlo, 1960), but unlike Marmornectes (Ketchum and Benson, 2011). A long and concave distal facet for the tibiale is also visible on the tibia, as well as a short and oblique facet, located preaxially, for a preaxial accessory ossicle which is not preserved.
The fibula (Figure 8C, F) has a lunate outline with a concave preaxial margin that forms the posterior border of the epipodial foramen as in Liopleurodon ferox (Adrews, 1913; Linder, 1913). A posteriorly oblique proximal facet contacts a postaxial accessory ossicle preserved on the left paddle. The tibiale, intermedium, and ulnare are similar in shape to the elements of the forelimb. Only one distal tarsal (dt) is preserved and identified as the dt2 and 3. One metatarsal (mt) shows a triangular-shaped proximal end and is thus interpreted as the mt2. Only few phalanges are preserved preventing the determination of the phalangeal formula.
Cladistic Analysis
 The parsimony analysis of the data matrix yielded 2000 most parsimonious trees of 1937 steps. We do not detail the results within the Rhomaleosauridae, nor the results within the Plesiosauroidea. The topology of the strict consensus (Figure 9) positions the Thouarsais specimen as the sister taxon of Liopleurodon ferox, and this clade is supported by four synapomorphies: the presence of transversely concave/convex cervical zygapophyseal facets (character 169, state 1), cervical centrum mediolateral width subequal to height or less (character 171, sate 0), caudal ribs facet located at midheight of centrum or lower in proximal-middle caudal vertebrae (character 188, state 2), and the chevron facets of caudal vertebrae that are located mainly on the posterior edge of the centra (character 192, sate 1). Thalassiodracon hawkinsii is sister taxon of the clade grouping Rhomaleosauridae and Pliosauridae. The clade that includes the three species of Hauffiosaurus is the sister taxon to all other members of Pliosauridae. The overall relationships among taxa within this latter clade remain largely unresolved, similar to the relationships identified by Sachs et al. (2023) in their analysis using unweighted parsimony.
The parsimony analysis of the data matrix yielded 2000 most parsimonious trees of 1937 steps. We do not detail the results within the Rhomaleosauridae, nor the results within the Plesiosauroidea. The topology of the strict consensus (Figure 9) positions the Thouarsais specimen as the sister taxon of Liopleurodon ferox, and this clade is supported by four synapomorphies: the presence of transversely concave/convex cervical zygapophyseal facets (character 169, state 1), cervical centrum mediolateral width subequal to height or less (character 171, sate 0), caudal ribs facet located at midheight of centrum or lower in proximal-middle caudal vertebrae (character 188, state 2), and the chevron facets of caudal vertebrae that are located mainly on the posterior edge of the centra (character 192, sate 1). Thalassiodracon hawkinsii is sister taxon of the clade grouping Rhomaleosauridae and Pliosauridae. The clade that includes the three species of Hauffiosaurus is the sister taxon to all other members of Pliosauridae. The overall relationships among taxa within this latter clade remain largely unresolved, similar to the relationships identified by Sachs et al. (2023) in their analysis using unweighted parsimony.
INTERPRETATION AND DISCUSSION
Comparison and Identification
The Thouarsais specimen displays several typical pliosaurid traits including wide cervical centra related to their length and height, a slightly convex ventral surface on cervical centra, a dorsal tip of the ilium flaring asymmetrically dorsally, and a convex preaxial margin on the tibia (e.g., Andrews, 1913; Tarlo, 1960; O’Keefe, 2001; Ketchum and Benson, 2010, 2011). The specimen also possesses a large antero-posterior expansion of the dorsal part of the ilium, the only unambiguous synapomorphy characterizing the clade Thalassophonea (Benson and Druckenmiller, 2014).
Information regarding the postcranial morphology in pliosaurids are limited, thus comparisons with Pliosaurus patagonicus Gasparini and O’Gorman, 2014, Gallardosaurus iturraldei Gasparini, 2009, Pliosaurus westburyensis Benson et al., 2013b, and Pliosaurus kevani Benson et al., 2013b, are not possible as these taxa are only or mostly known through skull remains. However, comparisons with pliosaurids in which postcranial skeletons are known show that (1) the absence of ventral ridge on cervical vertebrae of the Thouarsais specimen, as well as (2) the short size of its cervical centra, (3) the presence of a ventral lip on the latter, and (4) the presence of double-headed cervical rib facets, differ from (1) Peloneustes, P. brachydeirus, and Eardasaurus powelli Ketchum and Benson, 2022, (2) Makhaira, (3) Stenorhynchosaurus, BRSMG Cc332 (Pliosaurus sp., previously Pliosaurus brachyspondylus), as well as Pliosaurus rossicus, and (4) Stenorhynchosaurus, Plesiopleurodon, Brachauchenius, and Sachicasaurus, respectively (Tarlo, 1959; Halstead, 1971; Taylor and Cruickshank, 1993; Carpenter, 1996; Albright et al., 2007a, b; Ketchum and Benson, 2011; Benson et al., 2013b; Fischer et al., 2015; Páramo-Fonseca et al., 2016, 2018). In addition, the presence of ventral foramina on cervical centra in the Thouarsais specimen differs from most brachauchenine pliosaurids, except for Sachicasaurus and Luskhan (Benson and Druckenmiller, 2014; Fischer et al., 2017; Páramo-Fonseca et al., 2018). The cervical rib facets in the Thouarsais specimen are located ventro-laterally, as is typical in Jurassic pliosaurids (Fischer et al., 2017), and differ from the Cretaceous taxa Kronosaurus, Luskhan, and Brachauchenius in which the facets are located at mid-height of cervical centra (Fischer et al., 2017). The specimen ‘Pliosaurus’ andrewsi (NHMUK R3891) is distinguishable from the Thouarsais specimen by showing cervical neural spines that are transversely expanded dorsally, being wider medio-laterally than long antero-posteriorly (Andrews, 1913).
In the pectoral girdle, the anterior edge of the scapular dorsal process is sinusoidal in the Thouarsais specimen and differs from the one described in Simolestes, ‘Pliosaurus’ andrewsi (NHMUK R3891), Pliosaurus carpenteri (BRSMG Cd6172), and Pliosaurus rossicus (Tarlo, 1960; Ketchum and Benson, 2011).
Comparisons between propodial elements show that the Thouarsais specimen differs from Pliosaurus funkei in which the distal pre- and postaxial margins of the humeral shaft are symmetrical with no prominent postaxial expansion at the distal end (Knutsen et al., 2012). The femur in the Thouarsais specimen presents an expanded distal end that contrasts with the slender femur and the slightly expanded distal end observed in the pliosaurids Luskhan and Stenorhynchosaurus, as well as in the specimen MUHNCAL.20188 (Páramo-Fonseca et al., 2016; Fischer et al., 2017; Otero et al., 2020).
The epipodials in the Thouarsais specimen, longer proximo-distally than wide antero-posteriorly, differ from those of the pliosaurid specimen CAMSM J.35990 (Tarlo, 1960; Knutsen et al., 2012), previously identified as Stretosaurus macromerus Phillips, 1871, but provisionally referred to Pliosaurus cf. kevani by Benson et al. (2013b), ‘Pliosaurus’ andrewsi (NHMUK R3891), and Pliosaurus brachydeirus (Tarlo, 1960), as well as Stenorhynchosaurus, Pliosaurus almanzaensis, and Pliosaurus funkei (Knutsen et al., 2012; Páramo-Fonseca et al., 2016; O’Gorman et al., 2018). In addition, the presence of an epipodial foramen in the Thouarsais specimen allows to distinguish it from Luskhan (Fischer et al., 2017). The straight proximal surfaces of both the radius and ulna in the Thouarsais specimen contrast with the highly convex surfaces observed in Pliosaurus carpenteri (BRSMG Cd61172; Benson et al., 2013b).
Although the osteological features observed in the Thouarsais specimen contrast with most of pliosaurids, it shares some of the diagnostic characters that were reported from the post-cranial skeleton of Liopleurodon ferox (Andrews, 1913; Tarlo, 1960). Indeed, both the Thouarsais specimen and Liopleurodon ferox exhibit a ventral lip developed on the anterior surface of the cervical centrum without any ventral keel (Tarlo, 1960), and double-headed rib facets on the cervical vertebrae (Andrews, 1913). In addition, the Thouarsais specimen presents a combination of characters that can be relevant for its assignation to the species Liopleurodon ferox following Andrews (1913): short cervical centra (length equal or less than half width or height); cervical centra with a ventral lip and no ventral keel; two cervical rib facets on anterior and middle cervical centra; transversely compressed cervical neural spines longer antero-posteriorly than wide transversely with a teardrop- to oval-shaped dorsal tip; zygapophyses transversely narrower than the cervical centra; dorsal centra as high as wide and shorter than high; thin dorsal neural spine inclined postero-dorsally; sacral rib shaft twisted with a ridge extending dorsoventrally from the rib head to the flared distal end; dorsal process of the scapula with a convex, acute, and slightly sinusoidal anterior edge; humeral shaft that curves dorsodistally; humerus with pre- and postaxial margins of the shaft asymmetrical, distally, and with distinct radial and ulnar facets; radius longer than wide with concave postaxial borders forming the preaxial margin of a large epipodial foramen; ulna with a concave preaxial margin forming the postaxial border of the epipodial foramen, and a nearly straight postaxial margin; a femur longer than the humerus; epipodial foramen enclosed by tibia and fibula; tibia longer than wide.
Differences are also observed between the Thouarsais specimen and specimens previously assigned to Liopleurodon ferox. Thus, the length of the cervical centra, represents approximately 2/3 of the width and height of the vertebra in the Thouarsais specimen but less than half of the width and height in Liopleurodon ferox specimen NHMUK R.3536 (Tarlo, 1960). In addition, the neurocentral suture between the cervical neural arches and associated centra displays a V-shaped configuration in the Thouarsais specimen differing from the rounded suture observed in mounted specimen of Liopleuodon ferox (GPIT-PV-30093; and see character 172 in Sachs et al., 2023). Overall, these comparisons indicate possible intraspecific (including ontogenetic) variation in Liopleurodon ferox, and further studies should consider such a variability.
Based on comparisons performed here, the Thouarsais specimen is assigned to Liopleurodon ferox. Recently, Madzia et al. (2022) questioned the taxonomic status of the species L. ferox and recommended a taxonomic revision of the genus Liopleurodon. Pending a possible revision and according to Noè (2001), the genus Liopleurodon is monospecific, Liopleurodon pachydeirus being con-specific with Liopleurodon ferox, and ‘Liopleurodon’ macromerus as well as ‘Liopleurodon’ rossicus belonging to the genus Pliosaurus (Noè et al., 2004; Knutsen, 2012; Benson et al., 2013b). Among the 14 specimens referred to Liopleurodon ferox (Noè, 2001), excluding the isolated teeth present in numerous museum collections (Noè, 2001), only four include post-cranial remains: GPIT 2 (Huene, 1934), GPIT-PV-30093 mounted specimen (Linder, 1913), NHMUK R3536 and NHMUK R2446 (Andrews, 1913; Tarlo, 1960). Both specimens GPIT 2 and NHMUK R2446 being less complete, the Thouarsais specimen represents the most complete specimen from France and corresponds to the second or third most-complete post-cranial skeleton of a Liopleurodon ferox described so far, depending to what degree the specimen GPIT-PV-30093 is a composite skeleton (Noè, 2001; Martill et al., 2023).
Ontogenetic Stage
Various characters can be used to determine the ontogenetic status of plesiosaur specimens (Araújo and Smith, 2023). Among these characters, the fusion of the vertebral centra with the neural arches is one of the most widely used to determine whether the specimen is mature or not (Brown, 1981; Araújo et al., 2015). Indeed, the neurocentral junction in juvenile vertebrates is an important zone of growth for both the neural arch and the centrum (Vital et al., 1989), which connects these two separately ossified elements by cartilage (Ikejiri, 2012). The neurocentral junction fuses by the ossification of cartilage at or before the time of maturity (Romer, 1956). In plesiosaurs, open neurocentral sutures are present in juvenile and sub-adult plesiosaur specimens and are probably one of the last juvenile characters to disappear during ontogeny (Vincent, 2010). Here, we follow Brochu (1992, 1996) and Irmis (2007) who have specified certain definitions on this point: “an open suture is one that is visible on the surface of the bone, regardless of whether the two elements are firmly attached to each other. A closed suture has no trace of the suture on the surface of the bone. Whether or not two bony elements have actually ‘fused’ in the traditional sense cannot be determined without histologically sectioning across the boundary (Brochu, 1996; Cole et al., 2003) or using noninvasive high-resolution imaging such as computed tomography” (Irmis, 2007).
In the vertebral column of the Thouarsais specimen, the neural arches or ribs are attached to their corresponding centra (cervical, pectoral, dorsal, and sacral vertebrae) and show a marked suture on their surface (Figure 3C-E), which is identified as an open suture following Brochu (1992, 1996) and Irmis (2007). The specimen should thus not to be considered as an osteologically mature specimen sensu Araújo et al. (2015), but as a ‘juvenile’ individual sensu Brown (1981). However, as previously noted by McHenry (2009), Knutsen et al. (2012), and more recently by Ketchum and Benson (2022) as well as Araújo and Smith (2023), the neurocentral sutures between the cervical vertebral centra and associated neural arches remain open (still visible) in all large pliosaurids (see Table 1 and references herein). This differs from the sutures between the neural arches and associated centra of dorsal vertebrae that remain open in all pliosaurids (Table 1) except Brachauchenius (adult specimen MNA V9433), in which the suture is open on the cervical vertebrae but closed on the dorsal vertebrae (Albright et al, 2007b). The data available for caudal vertebrae remain rare; however, when observed, the sutures between the neural arches and centra of caudal vertebrae remain open in all pliosaurids (Table 1) except in Peloneustes (CAMSM J.46913, holotype), in which closed neural sutures are observed on the posterior caudals (Ketchum, 2007). The sutures between cervical ribs and centra remain open in the Thouarsais specimen. Such a feature is also observed in all pliosaurids in which cervical rib and centra are preserved (Table 1), except for Gallardosaurus (Gasparini, 2009) and Peloneustes (CAMSM J.46913; Ketchum, 2007).
Such a variability makes interpretation of ontogenetic stage from neurocentral sutures difficult in pliosaurids and requires specific attention. Without the observation of pliosaurids exhibiting a closed suture between neural arches and their cervical centra, it is impossible to know whether the neurocentral junctions fused even in old age. Thus, the lack of fusion between cervical neural arches and their centra in pliosaurids does not appear as a reliable indicator of a juvenile condition and could be considered as a paedomorphic feature of adult pliosaurids as already proposed by McHenry (2009), Knutsen et al. (2012), Ketchum and Benson (2022), and Araújo and Smith (2023). However, the lack of fusion between cervical ribs and their centra, as well as dorsal or caudal neural arches and their centra, remains questionable as it is not present in all pliosaurids. The characterization of being an osteologically mature individual of Plesiosauria (sensu Araújo et al., 2015) when the neurocentral suture is closed, should thus not been applied to taxa belonging to pliosaurids. Based on other post-cranial characters, such as the presence of well-defined articular facets on limb bones, the Thouarsais specimen is here interpreted as an ‘adult’ individual (sensu Brown 1981), showing a paedomorphic character of its neurocentral suture closure.
In some extant archosaurs (e.g., Alligator mississippiensis), the neurocentral sutures commonly remain open in at least some parts of the vertebral column (e.g., the presacral vertebrae in A. mississippiensis) long after sexual maturity (Brochu, 1996; Ikejiri, 2010, 2012). Similarly, the neurocentral junctions remain unfused in sauropod dinosaurs even as the animals attained very large body sizes (Fronimos and Wilson, 2017). Such a feature, in archosaurs, is required to delay the closure and thus extends the period of rapid growth at the neurocentral junction that permit large body size (Vital et al., 1989; Ikejiri, 2012; Fronimos and Wilson, 2017). In this context, it is not surprising to observe open neurocentral sutures between neural arches and their centra in pliosaurids, as members of this clade usually achieve a large body size.
Paleogeographic Distribution of Liopleurodon ferox
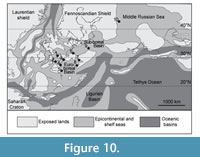 Most of the remains of the pliosaurid Liopleurodon ferox have been found in England, but have also been previously reported from France, Germany, Switzerland, Russia and possibly from Poland, although the available remains in this last locality are fragmentary and identified on the basis of teeth only (Lydekker, 1889; Persson, 1963; Bardet, 1993; Bardet et al., 1993b; Noé, 2001; Zverkov et al., 2017, 2024). The Thouarsais specimen extends the biogeographical area of the species Liopleurodon ferox into Central France (Figure 10) and represents the southernmost occurrence of this taxon. Liopleurodon ferox was thus present in three basins: the Sub-Boreal (i.e., Yorkshire, northern Germany and Poland) and Euro-Boreal basins (i.e., Paris, Central France, Wessex, Swabian basins), and a third one corresponding to the Moscow basin (Persson, 1963; Noé, 2001; Zverkov et al., 2017, 2024).
Most of the remains of the pliosaurid Liopleurodon ferox have been found in England, but have also been previously reported from France, Germany, Switzerland, Russia and possibly from Poland, although the available remains in this last locality are fragmentary and identified on the basis of teeth only (Lydekker, 1889; Persson, 1963; Bardet, 1993; Bardet et al., 1993b; Noé, 2001; Zverkov et al., 2017, 2024). The Thouarsais specimen extends the biogeographical area of the species Liopleurodon ferox into Central France (Figure 10) and represents the southernmost occurrence of this taxon. Liopleurodon ferox was thus present in three basins: the Sub-Boreal (i.e., Yorkshire, northern Germany and Poland) and Euro-Boreal basins (i.e., Paris, Central France, Wessex, Swabian basins), and a third one corresponding to the Moscow basin (Persson, 1963; Noé, 2001; Zverkov et al., 2017, 2024).
ACKNOWLEDGMENTS
We thank the team of the Centre d’Interprétation Géologique du Thouarsais for assistance and access to material in their care. We would like to thank I. Werneburg (Universität Tübingen, Tübingen) for the valuable information he provided on the palaeontological collections housed in Tübingen. We thank O. Bethoux (MNHN, Paris) for providing some photographs of the Liopleurodon mounted specimen kept in Tübingen. We thank the Willi Hennig Society for providing free access to the TNT program. This paper benefited from the highly valuable comments of L. Noè (Universidad de los Andes, Chile), D. Martill (University of Portsmouth, UK), D. Madzia (Polish Academy of Sciences, Poland), and an anonymous reviewer.
REFERENCES
Albright, L.B., Gillette, D.D., and Titus, A.L. 2007a. Plesiosaurs from the Upper Cretaceous (Cenomanian-Turonian) Tropic Shale of southern Utah, part 2: Polycotylidae. Journal of Vertebrate Paleontology, 27:41–58.
https://doi.org/10.1671/0272-4634(2007)27[41:PFTUCC]2.0.CO;2
Albright, L.B., Gillette, D.D., and Titus, A.L. 2007b. Plesiosaurs from the Upper Cretaceous (Cenomanian-Turonian) Tropic Shale of southern Utah, part 1: new records of the pliosaur Brachauchenius lucasi. Journal of Vertebrate Paleontology, 27:31–40.
https://doi.org/10.1671/0272-4634(2007)27[31:PFTUCC]2.0.CO;2
Andrews, C.W. 1913. A descriptive catalogue of the marine reptiles of the Oxford Clay, Part II. British Museum (Natural History), London.
Araújo, R. and Smith, A.S. 2023. Recognising and quantifying the evolution of skeletal paedomorphosis in Plesiosauria. Fossil Record, 26:85–101.
https://doi.org/10.3897/fr.26.97686
Araújo, R., Polcyn, M.J., Lindgren, J., Jacobs, L.L., Schulp, A.S., Mateus, O., Gonçalves, A.O., and Morais, M.-L. 2015. New aristonectine elasmosaurid plesiosaur specimens from the Early Maastrichtian of Angola and comments on paedomorphism in plesiosaurs. Netherlands Journal of Geosciences – Geologie en Mijnbouw, 94:93–108.
https://doi.org/10.1017/njg.2014.43
Bardet, N. 1993. Pliosaurs and plesiosaurs from the Middle Jurassic (Callovian) of Normandy. Revue de Paléobiologie, 7:1–7.
Bardet, N., Mazin, J.-M., and Martin, M. 1993a. A mandible of Pliosaurus brachyspondylus (Reptilia, Sauropterygia) from the Kimmeridgian of the Boulonnais (France). Neues Jahrbuch für Geologie und Paläontologie – Monatshefte, 7:385–392.
Bardet, N., Pennetier, G., Pennetier, E., and Queromain, J. 1993b. Présence du Pliosaure Liopleurodon ferox Sauvage dans le Jurassique moyen (Callovien) de Villers-sur-Mer, Normandie. Bulletin trimestriel de la Société géologique de Normandie et des amis du Muséum du Havre, 80:11–14.
Bardet, N., Falconnet, J., Fischer, V., Houssaye, A., Jouve, S., Pereda Suberbiola, X., Pérez-García, A., Rage, J.-C., and Vincent, P. 2014. Mesozoic marine reptile palaeobiogeography in response to drifting plates. Gondwana Research, 26:869–887.
https://doi.org/10.1016/j.gr.2014.05.005
Barrientos-Lara, J.I., Fernández, M.S., and Alvarado-Ortega, J. 2015. Kimmeridgian pliosaurids (Sauropterygia, Plesiosauria) from Tlaxiaco, Oaxaca, southern Mexic. Revista mexicana de ciencias geológicas, 32:293–304.
Beckary, S. 1992. Le pliosaure de Marquise. Annales de la Société géologique du Nord, 1:153–155.
Benson, R.B.J., Bates, K.T., Johnson, M.R., and Withers, P.J. 2011. Cranial anatomy of Thalassiodracon hawkinsii (Reptilia, Plesiosauria) from the Early Jurassic of Somerset, United Kingdom. Journal of Vertebrate Paleontology, 31:562–574.
https://doi.org/10.1080/02724634.2011.572937
Benson, R.B.J., Fitzgerald, E.M.G., Rich, T.H., and Vickers-Rich, P. 2013a. Large freshwater plesiosaurian from the Cretaceous (Aptian) of Australia. Alcheringa: An Australasian Journal of Palaeontology, 37:456–461.
https://doi.org/10.1080/03115518.2013.772825
Benson, R.B.J., Evans, M., Smith, A.S., Sassoon, J., Moore-Faye, S., Ketchum, H.F., and Forrest, R. 2013b. A giant pliosaurid skull from the Late Jurassic of England. PLoS ONE, 8 (5):e65989.
https://doi.org/10.1371/journal.pone.0065989
Benson, R.B.J. and Druckenmiller, P.S. 2014. Faunal turnover of marine tetrapods during the Jurassic-Cretaceous transition. Biological Reviews, 89:1–23.
https://doi.org/10.1111/brv.12038
Blainville de, H.D. 1835. Description de quelques espèces de reptiles de la Californie: précédée de l’analyse d’un système général d’erpétologie et d’amphibiologie. Nouvelles Annales du Muséum (national) d’Histoire Naturelle, Paris (serie 3), 4:233–296.
Bourquin, J. 1968. Les Reineckeidés. Annales Scientifiques de l’Université de Besançon, 3ème série, Géologie, 3:1–169.
Brochu, C.A. 1992. Ontogeny of the postcranium in crocodylomorph archosaurs. Unpublished M.A. thesis. The University of Texas at Austin, Austin, Texas.
Brochu, C.A. 1996. Closure of neurocentral sutures during crocodilian ontogeny: implications for maturity assessment in fossil archosaurs. Journal of Vertebrate Paleontology, 16:49–62.
https://doi.org/10.1080/02724634.1996.10011283
Brown, D.S. 1981. The English Upper Jurassic Plesiosauroidea (Reptilia) and a review of the phylogeny and classification of the Plesiosauria. Bulletin of the British Museum, (Natural History) Geology Series, 35:253–347.
Buchy, M.C., Frey, E., Stinnesbeck, W., and López-Oliva, J.G. 2003. First occurrence of a gigantic pliosaurid plesiosaur in the Late Jurassic (Kimmeridgian) of Mexico. Bulletin de la Société Géologique de France, 174:271–278.
https://doi.org/10.2113/174.3.271
Buffetaut, E., Colleté, C., Dubus, B., and Petit, J.L. 2005. The ‘sauropod’ from the Albian of Mesnil-Saint-Père (Aube, France): a pliosaur, not a dinosaur. Carnets de Géologie, Letter 2005/01.
Campbell, J.A., Mitchell, M.T., Ryan, M.J., and Anderson, J.S. 2021. A new elasmosaurid (Sauropterygia: Plesiosauria) from the non-marine to paralic Dinosaur Park Formation of southern Alberta, Canada. PeerJ, 9:e10720.
https://doi.org/10.7717/peerj.10720
Cariou, E. 1980. L’étage Callovien dans le Centre-Ouest de la France. Première partie: stratigraphie et paléogéographie; deuxième partie: les Reineckeiidae (Ammonitina), systématique, dimorphisme et évolution. Thèse d’état, Université de Poitiers.
Carpenter, K. 1996. A review of short-necked plesiosaurs from the Cretaceous of the Western Interior, North America. Neues Jahrbuch für Geologie und Paläontologie – Abhandlungen, 201:259–287.
https://doi.org/10.1127/njgpa/201/1996/259
Cole, A., Fedak, T., Hall, B., Olson, W., and Vickaryous, M. 2003. Sutures joining ontogeny and fossils. The Palaeontological Association Newsletter, 52:29–32.
Cruickshank, A.R.I., Martill, D.M., and Noè, L.F. 1996. A pliosaur (Reptilia, Sauropterygia) exhibiting pachyostosis from the Middle Jurassic of England. Journal of the Geological Society, 153:873–879.
https://doi.org/10.1144/gsjgs.153.6.08
Dhoste, M., Legendre, L., and Coubès, L. 1987. Carte géologique à 1/50 000, feuille Thouars (587), 1ère édition. Orléans, BRGM.
Faugeras, P. 1985. Stratigraphie, sédimentologie et paléoécologie comparées du Callovien du seuil du Poitou. Unpublished M.A. thesis, Poitiers University.
Fischer, V., Arkhangelsky, M.S., Stenshin, I.M., Uspensky, G.N., Zverkov, N.G., and Benson, R.B.J. 2015. Peculiar macrophagous adaptations in a new Cretaceous pliosaurid. Royal Society Open Science, 2:150552.
https://doi.org/10.1098/rsos.150552
Fischer, V., Benson, R.B.J., Zverkov, N.G., Soul, L.C., Arkhangelsky, M.S., Lambert, O., Stenshin, I.M., Uspensky, G.N., and Druckenmiller, P.S. 2017. Plasticity and Convergence in the Evolution of Short-Necked Plesiosaurs. Current Biology, 27:1667–1676.
https://doi.org/10.1016/j.cub.2017.04.052
Fronimos, J.A. and Wilson, J.A. 2017. Neurocentral suture complexity and stress distribution in the vertebral column of a sauropod dinosaur. Ameghiniana, 54:36.
https://doi.org/10.5710/AMGH.05.09.2016.3009
Gabilly, J. and Cariou, E. 1974. Journées d'étude et excursion en Poitou. Laboratoire de géologie sédimentaire et paléobiologie, Poitiers.
Gabilly, J., Cariou, E., and Hantzpergue, P. 1985. Géodynamique des seuils et des hauts-fonds. Poitiers, Laboratoire de géologie stratigraphique et structurale, Poitiers.
Gasparini, Z. 2009. A new Oxfordian pliosaurid (Plesiosauria, Pliosauridae) in the Caribbean Seaway. Palaeontology, 52:661–669.
https://doi.org/10.1111/j.1475-4983.2009.00871.x
Gasparini, Z. and O’Gorman, J.P. 2014. A New Species of Pliosaurus (Sauropterygia, Plesiosauria) from the Upper Jurassic of Northwestern Patagonia, Argentina. Ameghiniana, 51:269–283.
https://doi.org/10.5710/AMGH.03.04.2014.2225
Goller, T. 2021. Die Ära Ernst Koken und Friedrich von Huene 1903, 1904, 1906 und 1922: Plesiosaurier - die Unterwasserflieger, p. 320-335. In Seidl, E., Bierende, E., and Werneburg, I. (eds.), Aus der Tiefenzeit. Die Paläontologische Sammlung der Universität Tübingen. Tübingen (Schriften des Museums der Universität Tübingen MUT, Band 20).
Goloboff, P.A. and Morales, M.E. 2023. TNT version 1.6, with a graphical interface for MacOS and Linux, including new routines in parallel. Cladistics, 39:144–153.
https://doi.org/10.1111/cla.12524
Goloboff, P.A., Farris, J.S., and Nixon, K.C. 2008. TNT, a free program for phylogenetic analysis. Cladistics, 24:774–786.
https://doi.org/10.1111/j.1096-0031.2008.00217.x
Halstead, L.B. 1971. Liopleurodon rossicus (Novozhilov): a pliosaur from the lower Volgian of the Moscow Basin. Palaeontology, 14:566–570.
Hampe, O. 2005. Considerations on a Brachauchenius skeleton (Pliosauroidea) from the lower Paja Formation (late Barremian) of Villa de Leyva area (Colombia). Fossil Record, 8:37–51.
https://doi.org/10.1002/mmng.200410003
Hector, J. 1874. On the Fossil Reptilia of New Zealand. Transactions and Proceedings of the New Zealand Institute, 6:333–358.
Ikejiri, T. 2010. Morphology of the neurocentral junction during postnatal growth of Alligator (Reptilia, Crocodylia). Unpublished Ph.D. Thesis, University of Michigan, Ann Arbor, USA.
http://deepblue.lib.umich.edu/handle/2027.42/78850
Ikejiri, T. 2012. Histology-Based Morphology of the Neurocentral Synchondrosis in Alligator mississippiensis (Archosauria, Crocodylia). The Anatomical Record: Advances in Integrative Anatomy and Evolutionary Biology, 295:18–31.
https://doi.org/10.1002/ar.21495
Irmis, R.B. 2007. Axial skeleton ontogeny in the Parasuchia (Archosauria: Pseudosuchia) and its implications for ontogenetic determination in archosaurs. Journal of Vertebrate Paleontology, 27:350–361.
https://doi.org/10.1671/0272-4634(2007)27[350:ASOITP]2.0.CO;2
Ketchum, H.F. 2007. The anatomy, taxonomy and systematics of three British Middle Jurassic pliosaurs (Sauropterygia: Plesiosauria), and the phylogeny of Plesiosauria. Unpublished Ph.D. Thesis, University of Cambridge, UK.
Ketchum, H.F. and Benson, R.B.J. 2010. Global interrelationships of Plesiosauria (Reptilia, Sauropterygia) and the pivotal role of taxon sampling in determining the outcome of phylogenetic analyses. Biological Reviews, 85:361–392.
https://doi.org/10.1111/j.1469-185X.2009.00107.x
Ketchum, H.F. and Benson, R.B.J. 2011. A new pliosaurid (Sauropterygia, Plesiosauria) from the Oxford Clay Formation (Middle Jurassic, Callovian) of England: evidence for a gracile, longirostrine grade of Early-Middle Jurassic pliosaurids. Special Papers in Palaeontology, 86:109–129.
Ketchum, H.F. and Benson, R.B.J. 2022. A new pliosaurid from the Oxford Clay Formation of Oxfordshire, UK. Acta Palaeontologica Polonica, 67:297–315.
https://doi.org/10.4202/app.00887.2021
Knutsen, E.M., Druckenmiller, P.S., and Hurum, J.H. 2012. A new species of Pliosaurus (Sauropterygia: Plesiosauria) from the Middle Volgian of central Spitsbergen, Norway. Norwegian Journal of Geology, 92:235–258.
Koken, E. 1893. Beitrage zur Kenntniss der Gattung Nothosaurus. Zeitschrift der Deutschen Geologischen Gesellschaft, 45:337–377.
Krahl, A., Smith, A.S., and Werneburg, I. 2022. Historically transposed flipper pairs in a mounted plesiosaurian skeleton. Paläontologische Zeitschrift, 96:805–813.
https://doi.org/10.1007/s12542-022-00616-8
Lamouroux, J.V.F. 1821. Exposition méthodique des genres de l’ordre des polypiers: avec leur description et celle des principales espèces, figurées dans 84 planches, les 63 premières appartenant à l’Histoire naturelle des zoophytes d’Ellis et Solander. Veuve Agasse, Paris.
https://doi.org/10.5962/bhl.title.11328
Legendre, L. 1984. Les transgressions mésozoïques sur le promontoire oriental du Massif vendéen. Unpublished Ph.D. Thesis, University of Poitiers, France.
Legendre, L., Dhoste, M., and Coubès, L. 1989. Notice explicative de la carte géologique à 1/50 000, feuille Thouars (539). Orléans, BRGM.
Linder, H. 1913. Beiträge zur Kenntnis der Plesiosauria-Gattungen Peloneustes und Pliosaurus. Geologische und Palaeontologische Abhandlungen, 11:339–409.
Longman, A.H. 1924. A new gigantic marine reptile from the Queensland Cretaceous, Kronosaurus queenslandicus new genus and species. Memoirs of the Queensland Museum, 8:26–28.
Lydekker, R. 1889. Catalogue of the fossil Reptilia and Amphibia in the British Museum (Natural history), Part II. British Museum, London, UK.
Madzia, D., Sachs, S., and Klug, C. 2022. Historical significance and taxonomic status of Ischyrodon meriani (Pliosauridae) from the Middle Jurassic of Switzerland. PeerJ, 10:e13244.
https://doi.org/10.7717/peerj.13244
Maréchal, M., Debris, J.-P., Lepage, G., Cousin, R., and Watte, J.-P. 1999. Nouvelle découverte de restes de Sauroptérygiens dans le Kimméridgien du Cap de la Hève. Annales du Museum du Havre, 61:1–4.
Martill, D.M., Jacobs, M.L., and Smith, R.E. 2023. A truly gigantic pliosaur (Reptilia, Sauropterygia) from the Kimmeridge Clay Formation (Upper Jurassic, Kimmeridgian) of England. Proceedings of the Geologists’ Association, 134:361–373.
https://doi.org/10.1016/j.pgeola.2023.04.005
Martin, J.E., Sawyer, J.F., Reguero, M., and Case, J.A. 2007. Occurrence of a young elasmosaurid plesiosaur skeleton from the Late Cretaceous (Maastrichtian) of Antarctica. Online proceedings of the 10th International Symposium on Antarctic Earth Sciences. Antarctica: a keystone in a changing world. USGS Open-File Report, 1047.
McHenry, C.R. 2009. Devourer of gods: the palaeoecology of the Cretaceous pliosaur Kronosaurus queenslandicus. Unpublished Ph.D. Thesis, University of Newcastle, Australia.
Noè, L.F. 2001. A taxonomic and functional study of the Callovian (Middle Jurassic) Pliosauroidea (Reptilia, Sauropterygia). Unpublished Ph.D. Thesis, University of Derby, UK.
Noè, L.F., Smith, D.T.J., and Walton, D.I. 2004. A new species of Kimmeridgian pliosaur (Reptilia; Sauropterygia) and its bearing on the nomenclature of Liopleurodon macromerus. Proceedings of the Geologists’ Association, 115:13–24.
Noè, L.F. and Gómez-Pérez, M. 2022. Giant pliosaurids (Sauropterygia; Plesiosauria) from the Lower Cretaceous peri-Gondwanan seas of Colombia and Australia. Cretaceous Research, 132:105–122.
https://doi.org/10.1016/j.cretres.2021.105122
O’Gorman, J.P., Gasparini, Z., and Spalletti, L.A. 2018. A new Pliosaurus species (Sauropterygia, Plesiosauria) from the Upper Jurassic of Patagonia: new insights on the Tithonian morphological disparity of mandibular symphyseal morphology. Journal of Paleontology, 92:240–253.
https://doi.org/10.1017/jpa.2017.82
O’Keefe, F.R. 2001. A cladistic analysis and taxonomic revision of the Plesiosauria (Reptilia: Sauropterygia). Acta Zoologica Fennica, 213:1–63.
O’Keefe, F.R. and Carrano, M.T. 2005. Correlated Trends in the Evolution of the Plesiosaur Locomotor System. Paleobiology, 31:656–675.
https://doi.org/10.1666/0094-8373(2005)031[0656:CTITEO]2.0.CO;2
Otero, R.A., Soto-Acuña, S., Rojas, J., and Rojas, M.O. 2020. First pliosaur remains (Sauropterygia, Pliosauridae) from the Oxfordian of the Atacama Desert. Journal of South American Earth Sciences, 104:102811.
https://doi.org/10.1016/j.jsames.2020.102811
Owen, R. 1860. On the orders of fossil and recent Reptilia and their distribution in time. Report of the British Association for the Advancement of Science, 1859:153–166.
Owen, R. 1869. Monographs on the British Fossil Reptilia from the Kimmeridge Clay. No. III, Containing Pliosaurus grandis, Pl. trochanterius, and Pl. portlandicus. Monographs of the Palaeontographical Society, 22:1–12.
Páramo-Fonseca, M.E., Gómez-Pérez, M., Noé, L.F., and Etayo-Serna, F. 2016. Stenorhynchosaurus munozi, gen. et sp. nov. a new pliosaurid from the Upper Barremian (Lower Cretaceous) of Villa de Leiva, Colombia, South America. Revista de la Academia Colombiana de Ciencias Exactas, Físicas y Naturales, 40:84.
Páramo-Fonseca, M.E., Benavides-Cabra, C.D., and Gutiérrez, I.E. 2018. A new large Pliosaurid from the Barremian (Lower Cretaceous) of Sáchica, Boyacá, Colombia. Earth Sciences Research Journal, 22:223–238.
Páramo-Fonseca, M.E., Benavides-Cabra C.D., and Gutiérrez, I.E. 2019. A new specimen of Stenorhynchosaurus munozi Páramo-Fonseca et al., 2016 (Plesiosauria, Pliosauridae), from the Barremian of Colombia: new morphological features and ontogenetic implications. Journal of Vertebrate Paleontology, 39:e1663426.
https://doi.org/10.1080/02724634.2019.1663426
Persson, P.O. 1963. A revision of the classification of the Plesiosauria with a synopsis of the stratigraphical and geographical distribution of the group. Lunds Universitets Årsskrift, N.F. Avdelningen, 59:1–60.
Phillips, J. 1829. Illustrations of the geology of Yorkshire. York Wilson, York.
Phillips, J. 1871. Geology of Oxford and the valley of the Thames. Clarendon Press, Oxford.
Rauhut, O.W.M. and Carrano, M.T. 2016. The theropod dinosaur Elaphrosaurus bambergi, from the Late Jurassic of Tendaguru, Tanzania. Zoological Journal of the Linnean Society, 178:546–610.
https://doi.org/10.1111/zoj.12425
Reinecke, M. 1818. Maris protogaei Nautilos et Aronautas, vulgo Cornua Ammonis in Agro Coburgico et vicino reperiundos. L.C.A. Ahlii imp. Cobourg.
Romer, A.S. 1956. Osteology of the reptiles. University Chicago Press, Chicago, USA.
Romer, A.S. and Lewis, A.D. 1959. A mounted skeleton of the giant plesiosaur Kronosaurus. Breviora, 112:1–15.
Sachs, S., Rauhut, O.W.M., and Weigert, A. 1997. Mesozoische Reptilien aus Nordrhein-Westfalen. Terra Nostra, 1:22–27.
Sachs, S., Hornung, J.J., and Kear, B.P. 2016. Reappraisal of Europe’s most complete Early Cretaceous plesiosaurian: Brancasaurus brancai Wegner, 1914 from the “Wealden facies” of Germany. PeerJ, 4:e2813.
https://doi.org/10.7717/peerj.2813
Sachs, S. and Kear, B.P. 2018. A rare new Pliensbachian plesiosaurian from the Amaltheenton Formation of Bielefeld in northwestern Germany. Alcheringa, 42:487–500.
https://doi.org/10.1080/03115518.2017.1367419
Sachs, S., Madzia, D., Thuy, B., and Kear, B.P. 2023. The rise of macropredatory pliosaurids near the Early-Middle Jurassic transition. Scientific Reports, 13:17558.
https://doi.org/10.1038/s41598-023-43015-y
Saint-Leger, P. 1987. Excursion aux confins de la Touraine et du Poitou. Minéraux et fossiles, le guide du collectionneur, 144:38–41.
Sato, T. 2003. Terminonatator ponteixensis, a new elasmosaur (Reptilia: Sauropterygia) from the Upper Cretaceous of Saskatchewan. Journal of Vertebrate Paleontology, 23:89–103.
https://doi.org/10.1671/0272-4634(2003)23[89:TPANES]2.0.CO;2
Sauvage, H.E. 1873. Notes sur les reptiles fossiles. Bulletin de la Société Géologique de France, 1:365.
Schumacher, B.A., Carpenter, K., and Everhart, M.J. 2013. A new Cretaceous Pliosaurid (Reptilia, Plesiosauria) from the Carlile Shale (middle Turonian) of Russell County, Kansas. Journal of Vertebrate Paleontology, 33:613–628.
https://doi.org/10.1080/02724634.2013.722576
Seeley, H.G. 1869. Index to the Fossil Remains of Aves, Ornithosauria, and Reptilia, from the Secondary System of Strata, Arranged in the Woodwardian Museum of the University of Cambridge. Deighton, Bell, and Co., Cambridge, UK.
Seeley, H.G. 1874. Note on some of the generic modifications of the plesiosaurian pectoral arch. Quarterly Journal of the Geological Society, 30:436–449.
Sollas, W.J. 1881. On a new species of Plesiosaurus (P. conybeari) from the Lower Lias of Charmouth; with observation on P. macrocephalus, Stuchbury, and P. brachycephalus, Owen. Quarterly Journal of the Geological Society of London, 37:440–480.
Storrs, G.W. 1991. Anatomy and relationships of Corosaurus alcovensis (Diapsida: Sauropterygia) and the Triassic Alcova Limestone of Wyoming. Bulletin of the Peabody Museum of Natural History Yale University 44, 1–151.
Storrs, G.W. 1997. Morphological and taxonomic clarification of the genus Plesiosaurus, p. 145-190. In Callaway, J.M. and Nicholls, E.L. (eds.), Ancient Marine Reptiles. Academic Press, San Diego.
Tarlo, L.B. 1959. Pliosaurus brachyspondylus (Owen) from the Kimmeridge clay. Palaeontology, 1:283–291.
Tarlo, L.B. 1960. A review of the Upper Jurassic pliosaurs. Bulletin of the British Museum (Natural History), Geology, 14:145–189.
Taylor, M.A. and Cruickshank, A.R.I. 1993. A plesiosaur from the Linksfield erratic (Rhaetian, Upper Triassic near Elgin, Morayshire). Scottish Journal of Geology, 29:191–196.
Vincent, P. 2010. A juvenile plesiosaur specimen from the Lower Jurassic of Holzmaden, Germany. Palaeontographica Abteilung A, 291:45–61.
https://doi.org/10.1127/pala/291/2010/45
Vincent, P. 2011. A Re-Examination of Hauffiosaurus zanoni, a Pliosauroid from the Toarcian (Early Jurassic) of Germany. Journal of Vertebrate Paleontology, 31:340–351.
Vincent, P., Bardet, N., and Mattioli, E. 2013. A New Pliosaurid from the Pliensbachian, Early Jurassic of Normandy, Northern France. Acta Palaeontologica Polonica, 58:471–485.
http://doi.org/10.4202/app.2011.0113
Vincent, P., Allemand, R., Taylor, P.D., Suan, G., and Maxwell, E.E. 2017. New insights on the systematics, palaeoecology and palaeobiology of a plesiosaurian with soft tissue preservation from the Toarcian of Holzmaden, Germany. The Science of Nature, 104:51.
https://doi.org/10.1007/s00114-017-1472-6
Vital, J.M., Beguiristain, J.L., Algara, C., Villas, C., Lavignolle, B., Grenier, N., and Sénégas, J. 1989. The neurocentral vertebral cartilage: Anatomy, physiology and physiopathology. Surgical and Radiologic Anatomy, 11:323–328.
https://doi.org/10.1007/BF02098705
von Huene, F. 1934. Ein großer Pliosaurus aus dem schwäbischen Ornatenton. Jahreshefte des Vereins für vaterländische Naturkunde in Württemberg, 90:31–46.
White, T.E. 1940. The holotype of Plesiosaurus longirostris Blake and classification of the plesiosaurs. Journal of Paleontology, 14:451–467.
Wierzbowski, H. and Rogov, M. 2011. Reconstructing the palaeoenvironment of the Middle Russian Sea during the Middle-Late Jurassic transition using stable isotope ratios of cephalopod shells and variations in faunal assemblages. Palaeogeography, Palaeoclimatology, Palaeoecology, 299:250–264.
https://doi.org/10.1016/j.palaeo.2010.11.006
Williston, S.W. 1903. North American plesiosaurs. Part 1. Field Columbian Museum, Publication 73, Geology Series, 2:1–79.
Wintrich, T., Hayashi, S., Houssaye, A., Nakajima, Y., and Sander, P.M. 2017. A Triassic plesiosaurian skeleton and bone histology inform on evolution of a unique body plan. Science Advances, 3:e1701144.
https://doi.org/10.1126/sciadv.1701144
Zeiss, A. 1956. Hecticoceras und Reineckeia im Mittel- und Ober-Callovien von Blumberg (südbaden). Verlag der Bayerischen Akademie der Wissenschaften; in Kommission bei Beck. Mathematisch-Naturwissenschaftliche Klasse Abhandlungen, N.F., 80:1–101.
Zverkov, N.G., Shmakov, A.S., and Arkhangelsky, M.S. 2017. Jurassic marine reptiles of Moscow and surroundings, p. 230-263. In Rogov, M.A. and Zakharov, V.A. (eds.), Jurassic deposits of the southern part of the Moscow syneclyse and their fauna. Transaction of the Geological Institute, Moscow, Russia.
Zverkov, N.G., Arkhangelsky, M.S., Gulyaev, D., Ippolitov, A., and Shmakov, A. 2024. Callovian Marine Reptiles of European Russia. Diversity, 16:290.
https://doi.org/10.3390/d16050290
