

 Athenar bermani, a new species of dicraeosaurid sauropod from Dinosaur National Monument, Utah, U.S.A.
Athenar bermani, a new species of dicraeosaurid sauropod from Dinosaur National Monument, Utah, U.S.A.
Article number: 28.3.a50
https://doi.org/10.26879/1550
Copyright Society of Vertebrate Paleontology, October 2025
Author biographies
Plain-language and multi-lingual abstracts
PDF version
Submission: 8 March 2025. Acceptance: 16 September 2025.
ABSTRACT
Carnegie Museum of Natural History houses some of the most important specimens for our understanding of diplodocoid sauropod cranial anatomy. One specimen (CM 26552), a braincase and partial skull roof, is often overlooked in favor of more complete skulls, but bears an outsized historical importance as this specimen formed the basis for the modern description of braincase anatomy for Diplodocus as a genus. In light of several decades of new discoveries since the original description by Berman and McIntosh in 1978, here we re-evaluate the anatomy and taxonomic relationships of this specimen. We find that CM 26552 is a distinct not only from Diplodocus but also from the larger grouping Diplodocidae entirely, and is instead best regarded as a new genus of dicraeosaurid sauropod.
John A. Whitlock. Department of Science and Mathematics, Mount Aloysius College, Cresson, Pennsylvania 16630, U.S.A. and Section of Vertebrate Paleontology, Carnegie Museum of Natural History, Pittsburgh, Pennsylvania 15213, U.S.A. jwhitlock@mtaloy.edu
Juan Pablo Garderes. CONICET Consejo Nacional de Investigaciones Científicas y Técnicas, Ciudad Autónoma de Buenos Aires, Argentina and Fundación de Historia Natural Félix de Azara, Centro de Ciencias Naturales, Ambientales y Antropológicas, Universidad Maimónides, Hidalgo, Argentina. garderes.juanpablo@maimonides.edu
Pablo Gallina. CONICET Consejo Nacional de Investigaciones Científicas y Técnicas, Ciudad Autónoma de Buenos Aires, Argentina and Fundación de Historia Natural Félix de Azara, Centro de Ciencias Naturales, Ambientales y Antropológicas, Universidad Maimónides, Hidalgo, Argentina. gallina.pablo@maimonides.edu
Matthew C. Lamanna. Section of Vertebrate Paleontology, Carnegie Museum of Natural History, Pittsburgh, Pennsylvania 15213, U.S.A.: lamannam@carnegiemnh.org
Keywords: new genus; new species; Morrison Formation; Sauropoda; Late Jurassic; Dicraeosauridae
Final citation: Whitlock, John A., Garderes, Juan Pablo, Gallina, Pablo, and Lamanna, Matthew C. 2025. Athenar bermani, a new species of dicraeosaurid sauropod from Dinosaur National Monument, Utah, U.S.A.. Palaeontologia Electronica, 28(3):a50.
https://doi.org/10.26879/1550
palaeo-electronica.org/content/2025/5682-athenar-bermani-new-dicraeosaurid-sauropod
Copyright: October 2025 Society of Vertebrate Paleontology.
This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
creativecommons.org/licenses/by/4.0
https://zoobank.org/C612A8A6-4369-46AF-AA94-6E6BC6393082
INTRODUCTION
Carnegie Museum (CM) specimen 26552 is a braincase and partial skull roof from the Late Jurassic (ca. 150-151 Ma; Maidment et al., 2017; Whitlock et al., 2018; Hunt-Foster, 2024) Carnegie Quarry at Dinosaur National Monument.
Originally collected in 1913 by Earl Douglass, the specimen sat undescribed and unfigured for 65 years until briefly being described in a treatise most famous for “recapitating” Apatosaurus (Berman and McIntosh, 1978).
In that work, Berman and McIntosh (1978; p. 13) formally assigned CM 26552 to the genus Diplodocus on the basis of, “the common possession of characters of CM 11161 and the unquestionable Diplodocus skull CM 3452 that are not seen in the probable Apatosaurus skull CM 11162”. This was an important assignation, as CM 26552 formed the basis “on which [their] description of the braincase of Diplodocus mainly rests” (Berman and McIntosh, 1978; p.13). Unfortunately, the general anatomical similarity between the crania of Diplodocus and Apatosaurus (and flagellicaudatan diplodocoid sauropods in general; see Upchurch, 1998; Wilson and Sereno, 1998; Whitlock, 2011; Mannion et al., 2012; Whitlock and Wilson, 2013; Tschopp et al., 2015; Tschopp and Mateus, 2017) meant that the majority of evidence at the time was based on proportional differences and not explicit character states, which, due to the obvious effects of scaling and ontogeny, are perhaps no longer sufficient to provide a diagnosis given the vast increase in anatomical knowledge acquired over the intervening decades. As a result, a reevaluation of CM 26552 in the light of the past 47 years of additional information is valuable not only in the context of the specimen itself but also in terms of evaluating a fundamental paper in sauropod paleontology.
In particular, there has been a tremendous increase in the known taxonomic diversity of diplodocoid sauropods recovered from the Morrison Formation (Woodruff, 2019). At the time of the original description of CM 26552, only five valid diplodocoid genera (Amphicoelias, Apatosaurus, Barosaurus, Diplodocus and Haplocanthosaurus; Dystrophaeus may also be considered here) were known from the Morrison Formation, and only a single genus (Diplodocus) had a correctly assigned skull. Since then, Supersaurus (Jensen, 1985), Dyslocosaurus (McIntosh et al., 1992), Suuwassea (Harris and Dodson, 2004), Kaatedocus (Tschopp and Mateus, 2013), Galeamopus (Tschopp and Mateus, 2017), Smitanosaurus (Whitlock and Wilson Mantilla, 2020), and Ardetosaurus (van der Linden et al., 2024) have been described, three of which have been interpreted as dicraeosaurids (Suuwassea, Kaatedocus [but see Tschopp et al., 2015], and Smitanosaurus), and four of which preserve comparable cranial material (Galeamopus, Kaatedocus, Smitanosaurus, and Suuwassea). Furthermore, the addition of pertinent diplodocoid cranial material from Argentina (Amargasaurus, Bajadasaurus, Campananeyen, Lavocatisaurus, Leinkupal, Limaysaurus, and Sidersaura; Salgado and Bonaparte, 1991; Calvo and Salgado, 1995; Canudo et al., 2018; Gallina et al., 2014, 2019; Garderes et al., 2021; Lerzo et al., 2024a; Lerzo et al., 2024b), China (Lingwulong; Xu et al., 2018), Niger (Nigersaurus; Sereno et al., 2007and Spain (Demandasaurus; Torcida Fernandez-Baldor et al., 2011) has provided workers with a wealth of comparative anatomical data from which to glean discrete morphological characters.
Here, we revisit the anatomy of CM 26552, evaluate it in light of those subsequent discoveries, and place the specimen within an appropriate phylogenetic context. A full phylogenetic analysis including this specimen will be included as a separate paper within this volume.
Institutional abbreviations
ANS, Academy of Natural Sciences of Drexel University, Philadelphia; CM, Carnegie Museum of Natural History, Pittsburgh; CMC, Cincinnati Museum Center, Cincinnati; HMNS, Houston Museum of Natural Science, Houston; MACN, Museo Argentino de Ciencias Naturales, Buenos Aires; MB, Museum für Naturkunde, Berlin; MMCh, Museo Municipal de Villa el Chocón-Ernesto Bachmann, Neuquén; MUCP, Museo de Geología y Paleontología, Villa el Chocón; NMZ, Natural History Museum, University of Zurich, Zurich; SMA, Sauriermuseum Aathal, Aathal; SMM, Science Museum of Minnesota, St. Paul; TATE, Tate Geological Museum, Casper; USNM, National Museum of Natural History, Washington; YPM, Yale Peabody Museum, New Haven.
SYSTEMATIC PALEONTOLOGY
SAURISCHIA Seeley, 1887
SAUROPODA Marsh, 1878
DIPLODOCOIDEA Marsh, 1884 (Upchurch, 1995)
FLAGELLICAUDATA Harris and Dodson, 2004
DICRAEOSAURIDAE Janensch, 1914
Genus ATHENAR gen. nov.
Athenar bermani gen. et sp. nov.
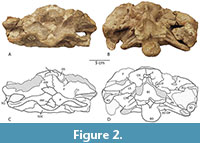
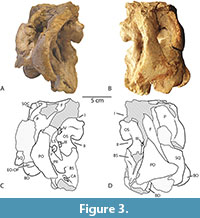
Figure 1, Figure 2, Figure 3
zoobank.org/6315E694-4BA7-44A4-B192-CB43F9D88B58
Type Species. Athenar bermani gen. et sp. nov.
zoobank.org/2F58D0D4
Holotype. CM 26552, a braincase and partial skull roof.
Locality and Horizon. Carnegie Quarry, Dinosaur National Monument, Utah, U.S.A. Uppermost Jurassic (Tithonian; Maidment et al., 2017; Whitlock et al., 2018) stratum of the Morrison Formation.
Etymology. The generic epithet Athenar is in honor of Cleveland musician and notable Pittsburgh Steelers fan Athenar, whose music was the soundtrack to the majority of the work done on this specimen and for whom no better paleontological comparison exists than a broken skull. The specific epithet bermani is in honor of David S. Berman, who, in collaboration with John S (“Jack”) McIntosh, did so much of the fundamental modern work on diplodocoid skulls at Carnegie Museum of Natural History and was responsible for the initial description of the specimen.
Diagnosis. Dicraeosaurid sauropod based upon: 1) the presence of postparietal and frontoparietal fenestrae, 2) the exclusion of the basioccipital from the dorsal margin of the occipital condyle by the exoccipitals, 3) the presence of a distinct prong on the squamosal, 4) contribution of the frontal to the margin of the supratemporal fenestra, 5) an expanded crista prootica, 6) free dorsal margin of the antotic process, 7) the presence of a "shelf” overhanging the foramen for the trigeminal (V) nerve, and 8) the flat distal margin of the paroccipital process (Whitlock and Wilson Mantilla, 2020; n.b. the specimen referred to Apatosaurus by Balanoff et al. [2010] also appears to display postparietal and frontoparietal fenestrae, although ongoing work by JAW as well as preliminary work by Moore and Mannion [2024] suggest dicraeosaurid affinities for that specimen as well). Distinct from Suuwassea emilieae based upon: 1) the presence of a single foramen for passage of the abducens (VI) nerve, 2) a narrow, midline ventral expansion of the foramen for the optic nerve and 3) greater size at maturity (see below). Additionally, there appears to be a notable gap in geologic age between the Tithonian A. bermani and the Kimmeridgian S. emilieae (but see also Maidment and Muxworthy, 2019 and Maidment, 2023 for dissenting opinion on relative ages). Distinct from Kaatedocus siberi based upon: 1) the presence of a frontoparietal fenestra and 2) the contribution of the frontal to the supratemporal fenestra in CM 26552. Distinct from Smitanosaurus agilis based upon: 1) the presence of a broad supraoccipital crest and 2) the contribution of the frontal to the supratemporal fenestra in CM 26552. Can be distinguished from all other dicraeosaurids by a single autapomorphy: the presence of a ‘tooth’ in the parietal/opisthotic suture.
DESCRIPTION OF CM 26552
Anatomical and orientational terminology uses traditional or ‘Romerian’ descriptors, following Wilson (2006). Orientational descriptors of cranial elements refer to a cranial position wherein the occiput is oriented vertically.
Preservation
CM 26552 is a braincase and partial skull roof. The elements have been slightly taphonomically distorted, resulting in a slight shearing in the transverse plane, such that the right half of the skull is shifted dorsally relative to the left, primarily affecting the dermal elements of the skull roof. An additional fragment containing the distal portion of the right paroccipital process and the proximal end of the right squamosal was found with the specimen and later attached with plaster, although this has since fractured and there do not appear to be any conforming surfaces to properly reunite the elements.
Ontogenetic Stage
Although quite large (width from center of occipital condyle to lateral margin of paroccipital process 89.9 mm, compared to the ‘subadult’ Diplodocus CM 3452 [81.0 mm] and the presumptively skeletally mature Diplodocus CM 11161 [85.5 mm]), the presence of patent, weakly interdigitated sutures between many of the braincase elements (e.g., between the right and left orbitosphenoids, and between the orbitosphenoids and the basisphenoid) suggests that CM 26552 died while still in a ‘subadult’ developmental stage.
Dermal Skull
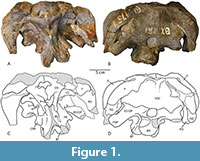 Frontal. Portions of both the right and left frontal bones are present in CM 26552, although both elements are poorly preserved (Figure 1, Figure 2). In dorsal view, the posterior portion of their midline contact and the medial portion of their contact with the parietals posteriorly can be seen, enclosing the anterior margin of a large foramen separating the frontals and parietals along the midline, the frontoparietal fenestra (Figure 1A, C). There is a patent, interdigitated suture separating the right and left frontals from each other, with a small boss present along the suture near the frontoparietal fenestra. The contact with the parietals is patent and interdigitated as well, bowing slightly posteriorly as it travels laterally from the frontoparietal fenestra toward the supratemporal fenestra. The frontal contributes to the entirety of the anterior margin of the supratemporal fenestra, as it does in the dicraeosaurids Suuwassea (Harris, 2006) and Bajadasaurus (Garderes et al., 2023), but unlike Amargasaurus or all other Morrison Formation sauropods (Wilson, 2002), including the dicraeosaurid Smitanosaurus (= `Morosaurus’ agilis; Whitlock and Wilson Mantilla, 2020).
Frontal. Portions of both the right and left frontal bones are present in CM 26552, although both elements are poorly preserved (Figure 1, Figure 2). In dorsal view, the posterior portion of their midline contact and the medial portion of their contact with the parietals posteriorly can be seen, enclosing the anterior margin of a large foramen separating the frontals and parietals along the midline, the frontoparietal fenestra (Figure 1A, C). There is a patent, interdigitated suture separating the right and left frontals from each other, with a small boss present along the suture near the frontoparietal fenestra. The contact with the parietals is patent and interdigitated as well, bowing slightly posteriorly as it travels laterally from the frontoparietal fenestra toward the supratemporal fenestra. The frontal contributes to the entirety of the anterior margin of the supratemporal fenestra, as it does in the dicraeosaurids Suuwassea (Harris, 2006) and Bajadasaurus (Garderes et al., 2023), but unlike Amargasaurus or all other Morrison Formation sauropods (Wilson, 2002), including the dicraeosaurid Smitanosaurus (= `Morosaurus’ agilis; Whitlock and Wilson Mantilla, 2020).
 The frontal contacts the orbitosphenoid along a massive pedicle approximately 25 mm wide. The frontals meet above a large foramen for the passage of the olfactory nerve; where the frontals contact there is a prominent ventral expansion that partially bifurcates this foramen dorsally, representing a midline division for the olfactory tracts not previously reported in other dicraeosaurids. The contact with the laterosphenoid is best preserved on the right side, where it appears to be gently sinuous.
The frontal contacts the orbitosphenoid along a massive pedicle approximately 25 mm wide. The frontals meet above a large foramen for the passage of the olfactory nerve; where the frontals contact there is a prominent ventral expansion that partially bifurcates this foramen dorsally, representing a midline division for the olfactory tracts not previously reported in other dicraeosaurids. The contact with the laterosphenoid is best preserved on the right side, where it appears to be gently sinuous.
In lateral view (Figure 3A), a portion of the posterior margin of the right frontal appears to preserve the rugose texture typical of diplodocoids. The lateral contact with the postorbital is a scarf joint angled subvertically.
 Parietal. Both parietals are preserved in CM 26552; the right is essentially complete, whereas the left is incomplete laterally. In dorsal view, the parietal is a strap-like element, forming only the posterior margin of the supratemporal fenestra (Figure 1A, C). The supratemporal fenestrae have an elliptical outline, with a long axis noticeably longer than the short axis (left 25 x 34 mm; right 22 x 35 mm [est.]). Although the skull roof is damaged near the contact between the right and left parietal, there appears to have been a brief, patent suture separating the elements anterior to the postparietal foramen near the contact with the supraoccipital. On either side of the midline, the supraoccipital is strongly concave posteriorly, forming the dorsolateral portion of the prominent occipital fossae.
Parietal. Both parietals are preserved in CM 26552; the right is essentially complete, whereas the left is incomplete laterally. In dorsal view, the parietal is a strap-like element, forming only the posterior margin of the supratemporal fenestra (Figure 1A, C). The supratemporal fenestrae have an elliptical outline, with a long axis noticeably longer than the short axis (left 25 x 34 mm; right 22 x 35 mm [est.]). Although the skull roof is damaged near the contact between the right and left parietal, there appears to have been a brief, patent suture separating the elements anterior to the postparietal foramen near the contact with the supraoccipital. On either side of the midline, the supraoccipital is strongly concave posteriorly, forming the dorsolateral portion of the prominent occipital fossae.
In posterior view, the parietal descends ventrolaterally along its contact with the supraoccipital (Figure 1B, D). Approximately 35 mm lateral to the supraoccipital crest, this suture features a prominent ‘tooth’, where a triangular expansion of the parietal inserts into the exoccipital-opisthotic below it. A similar, albeit less prominent sinuosity occurs in the probable Apatosaurus skulls CM 11162 and CMC VP7180 as well as in the ‘subadult’ Diplodocus CM 3452, but not in the mature Diplodocus CM 11161 or in the ‘juvenile’ Diplodocus SMM P84.15.3.
The sinuosity mostly resembles the condition in the dicraeosaurid Smitanosaurus (Whitlock and Wilson Mantilla, 2020). Above this contact, the parietal forms a prominent ‘wing’ that partially obscures the supratemporal fenestra. The parietal appears to be excluded from contributing to the posttemporal fenestra by the squamosal, as is typical of flagellicaudatans (Whitlock, 2011; but see Bajadasaurus; Gallina et al., 2019).
Postorbital. The postorbitals are triradiate elements with anterodorsal, posterior, and anteroventral processes. The postorbitals of CM 26552 preserve much of the anatomy of the anterodorsal and posterior processes, although much of the anteroventral processes are missing.
In lateral view, the scarf-like contact between the anterodorsal process and the frontal is visible (Figure 3). The anterodorsal process also contributes to a part of the anterior margin of the supratemporal fenestra and can be seen to contact the parietal medially within that opening. The ornamentation on the lateral margin of the frontal continues down onto the margin of the anterodorsal process of the postorbital near its contact with the frontal. A short, subtriangular posterior process inserts into a pocket in the squamosal, forming the separation between the supratemporal and lateral temporal fenestrae.
Squamosal. Portions of both right and left squamosals are preserved in CM 26552, between them representing all but the anteroventral process that forms much of the posteroventral margin of the lateral temporal fenestra in diplodocoids. In lateral view, the squamosal can be seen to contribute to a short (ca. 15 mm) section of the ventral margin of the supratemporal fenestra, separating the postorbital and the parietal laterally (Figure 3). The right squamosal preserves a large (ca. 17 mm), ventrally directed ‘prong’ just posterior to the fossa for the insertion of the quadrate, similar to that seen in dicraeosaurids like Bajadasaurus, Dicraeosaurus, Smitanosaurus and Suuwassea (Whitlock, 2011; Whitlock and Wilson Mantilla, 2020; Garderes et al., 2023).
Braincase
As mentioned above, a number of patent sutures between the elements of the braincase suggest that this individual was still a subadult at the time of death. The braincase itself is largely undistorted, and preserves all but the distal portions of the parasphenoid rostrum and basipterygoid processes. The endocranial cavity is filled with matrix.
Basioccipital. The basioccipital is a midline element forming the majority of the occipital condyle and the basal tubera. The basioccipital is excluded from the foramen magnum by the exoccipital-opisthotics dorsally, which also form the `shoulders’ of the occipital condyle (Figure 1B, D).
In posterior view, the occipital condyle is vaguely ovate, although the dorsal margin is flat or slightly concave dorsally due to the presence of the prominent ‘shoulders’ created by the exoccipital-opisthotics. It is slightly wider than tall (45 x 31 mm). The right `shoulder’ is missing, and although a broken plaster surface provides evidence that the element was originally collected, the piece has been missing since at least 1978 (see figure 3b in Berman and McIntosh, 1978). Despite this absence, it is plain from the left ‘shoulder’ and the preserved contact on the basioccipital for the right `shoulder’ that the basioccipital was largely excluded from the foramen magnum and the dorsal surface of the occipital condyle by the exoccipital-opisthotics, as in Amargasaurus (MACN-N 15), Dicraeosaurus (Tschopp et al., 2015), Smitanosaurus (Whitlock and Wilson Mantilla, 2020), and Suuwassea (Harris, 2006). The articular surface of the condyle is strongly convex and rugose. The basioccipital forms the majority of the basal tubera, and their entire posterior face. The tubera are broader than the condyle (ca. 61 mm) and have gently concave posterior surfaces. The general character of the tubera more closely resembles the `pendulous' tubera characteristic of Diplodocus than the `globose' tubera of taxa like Apatosaurus (Whitlock et al., 2010).
In posterolateral view, the suture between the basioccipital and the basisphenoid can be seen in the parabasal fossae, rising toward the exoccipital and meeting that element approximately at the crista interfenestralis, which divides the fenestra ovalis anteriorly from the metotic foramen posteriorly (Figure 2, Figure 3). In lateral view, the occipital condyle appears to be angled posteroventrally at about 45 degrees relative to the skull roof, similar to the angle seen in other Morrison Formation diplodocoids (e.g., Diplodocus; Sereno et al., 2007).
Basisphenoid. The basisphenoid is largely complete, contacting the orbitosphenoid dorsally, the laterosphenoid and prootic posterodorsally, and the basioccipital posteriorly (Figure 2A, C). The parasphenoid rostrum is largely absent, preserved only as a low broken ridge on the anterior face of the element. The dorsal margin of the basisphenoid has a U-shaped invagination along the midline where it meets the opening for the optic nerves (II) -which conjoin medially as in several dicraeosaurids (e.g., Bajadasaurus, Lingwulong, and Kaatedocus; Garderes et al., 2023)-, effectively shortening the parasphenoid rostrum dorsoventrally. Ventrally, the basisphenoid makes up the majority of the body of the basal tubera. The bases of the basipterygoid processes are present, but the processes themselves are broken off very near their origin and so little can be inferred about their length, diameter, or angle of divergence.
Supraoccipital. The supraoccipital is roughly trapezoidal, with a wide ventral base and a narrow dorsal margin, and it contacts the parietal dorsolaterally and the exoccipital-opisthotic ventrolaterally (Figure 2, Figure 3). A substantial frontoparietal fenestra occurs at the midline between the parietals and the supraoccipital. Posterior to this fenestra, the posterodorsal corner of the supraoccipital is expanded into a massive tubercle that forms the dorsal origin of a broad, sagittally-oriented supraoccipital crest. Laterally, this tubercle grades into a pair of low crests that continue on to the parietals as the lateral occipital crests.
As is common for subadult sauropod braincases, the suture between the supraoccipital and the exoccipital-opisthotic is patent and traceable (Figure 1B, D). The supraoccipital can be seen to form part of the dorsal margin of the foramen magnum, as is the case in most neosauropods (Whitlock et al., 2010). The contact continues laterally towards the posttemporal fenestra, where it can be seen arching dorsally as a dorsolateral process of the exoccipital-opisthotic excludes the supraoccipital from participating in the margin of that fenestra.
Exoccipital and Opisthotic. As is typical of sauropods, the exoccipital and opisthotic are indistinguishably fused in this specimen and as such will be treated as a single element. The exoccipital-opisthotic complex contacts the supraoccipital dorsally along a highly sinuous suture that runs from the dorsolateral corner of the foramen magnum laterally to contact the parietal just dorsolateral to the posttemporal fenestra (Figure 1B, D). A prominent paroccipital process extends ventrolaterally to form the basal margin of the posttemporal fenestra and eventually contact and cover posteriorly the squamosal at the lateral margin of the skull. A prominent processus posttemporalis (sensu Harris, 2006) completes the contribution of the exoccipital-opisthotic to the posttemporal fenestra by forming the dorsal and dorsomedial margins and excluding the supraoccipital from participation. Tschopp et al. (2015; p. 44) characterized this condition, where the supraoccipital is excluded from the posttemporal fenestra, as being typical of all diplodocoids; however in diplodocids like Diplodocus (CM 3452; CM 11161; SMM P.84.15.3) and Apatosaurus (CM 11162; YPM 1860; probably also CMC VP7180 although the suture is difficult to distinguish), and in the dicraeosaurid Bajadasaurus (MMCh-Pv 75; Garderes et al., 2023), no spur exists on the exoccipital and the supraoccipital forms a significant part of the margin of the fenestra. In many of these cases (e.g., CM 11161), a similar ‘spur’ shape appears as the posttemporal fenestra invades the supraoccipital, but careful examination of the region reveals that there is no contribution to this spur by the exoccipital. However, a spur formed by the exoccipital (and excluding the supraoccipital from the margin of the fenestra) is also known in the North American dicraeosaurids Suuwassea (ANS 21122; Harris, 2006), Kaatedocus (Tschopp et al., 2015; figure 16) and Smitanosaurus (USNM 5384), as well as in other dicraeosaurids such as Amargasaurus (MACN-N 15; Salgado and Calvo, 1992) and Lingwulong (Xu et al., 2018; figure SI 2).
Prootic. The prootic forms the lateral wall of the braincase posterior to the laterosphenoid and dorsal to the basisphenoid. In lateral view, the prootic is divided by the prominent crista prootica, which forms the anterior margin of the parabasal fossae (Figure 3). The dorsal parts of the parabasal fossae are subdivided into two subfossae by a low crest extending from the metotic foramen laterally and running approximately subparallel to the crista prootica. The prootic forms the dorsal margin of the foramen for the facial nerve (VII) and the very large metotic foramen posterior to it. The fenestra ovalis is entirely within the prootic, anterior to the metotic foramen. Anterior to the crista prootica, the foramen for the trigeminal nerve (V) exits between the prootic and the laterosphenoid anterior to it. On the right side, a prominent shelf overhanging the trigeminal foramen is present, as in Smitanosaurus and other dicraeosaurids (Whitlock and Wilson Mantilla, 2020), and a broken margin along the contact between the prootic and laterosphenoid at the dorsal margin of the foramen on the left side indicates this was symmetrical.
In anterior view, the prootic is largely defined by the broad crista prootica, which begins at the ventrolateral corner of the bone at its contact with the exoccipital and forms a sweeping curve ventrally as it approaches the basisphenoid, terminating in a substantial lateral expansion similar to the ‘tab’ seen in Amargasaurus and Dicraeosaurus (Salgado and Bonaparte, 1991) and probably also in Bajadasaurus (Gallina et al., 2019) and Suuwassea (Harris, 2006).
Laterosphenoid. The laterosphenoids form a small part of the lateral wall of the braincase, between the prootic posteriorly and the orbitosphenoids anteriorly. In lateral view, the laterosphenoid forms the anterior margin of the trigeminal foramen (V), and the posterior margins of the foramina for the oculomotor (III) and trochlear (IV) nerves. At its ventral terminus, the laterosphenoid appears to completely enclose the abducens foramen (VI).
In anterior view, the laterosphenoid is mostly defined by the large crista antotica, which largely mirrors the shape of the crista prootica as it travels laterally to contact the postorbital. The dorsal contact with the frontal is thick and widely separated, and it seems likely that the large, tapering antotic process directly abutted the postorbital in life, similar to the condition in Amargasaurus (MACN-N 15), Bajadasaurus (MMCh-Pv 75), Dicraeosaurus (MB.R. 2379.1), Kaatedocus (NMZ 1000004 [formerly SMA 004]), Limaysaurus (MUCPv-205), Nigersaurus (MNN GAD512), and Suuwassea (Harris, 2006), but not Apatosaurus (TATE-099; CMC VP 7180), Diplodocus (CM 3452; CM 11161; SMM P84.15.3) or Galeamopus (HMNS 175). The dorsomedial corner of the laterosphenoid is marked by a short, thick pedicle which extends out toward the orbitosphenoid and forms the posterior margin of the trochlear foramen (IV).
Orbitosphenoid. The orbitosphenoids form the anterior wall of the braincase, as well as part of the margins of the foramina for the first four cranial nerves (Figure 1). In lateral view, the contact with the laterosphenoid posteriorly is marked primarily by the presence of large foramina for the oculomotor (III) and trochlear (IV) nerves, separated from each other by a large, pillar-like posterior expansion of the orbitosphenoids, as in Amargasaurus (MACN-N 15) and probably Bajadasaurus (MMCh-Pv 75). The anterior margin is marked by a prominent crest dorsally and a large opening for the optic nerve (II) ventrally. The contact with the basisphenoid is patent and mildly interdigitated; it takes a ‘stepped’ appearance, with a ventral expansion of the orbitosphenoid posteriorly and a dorsal expansion of the basisphenoid anteriorly.
In anterior view, the dorsal margin is gently convex ventrally, owing primarily to the contribution to the foramen for the passage of the olfactory nerve (I). The suture between the right and left orbitosphenoids is patent and without interdigitation. At the dorsomedial corner of each orbitosphenoid, a prominent tubercle appears, a trait shared with Amargasaurus (MACN-N 15) and, probably, Dicraeosaurus (MB.R. 2379.1). Ventral to the opening for the olfactory nerve, a large opening for the optic nerves occurs, which exit the braincase conjoined as in all dicraeosaurids except Amargasaurus and Dicraeosaurus (Garderes et al., 2023). A prominent ventral expansion of the orbitosphenoids along their midline contact incompletely divides this foramen, as also seen in Bajadasaurus (MMCh-Pv 75).
DISCUSSION
Phylogenetic Affinities of CM 26552
CM 26552 can be assigned to Diplodocoidea over Macronaria based upon two well-established characters. First, the squamosal extends anterior to the posterior margin of the orbit (Whitlock, 2011). Second, the angle of the skull roof relative to the occipital condyle indicates a downturned face, characteristic of diplodocoids (Wilson and Sereno, 1998; Wilson, 2002; Upchurch et al., 2004; Sereno et al., 2007). Within Diplodocoidea, CM 26552 can be placed within Flagellicaudata based upon the exclusion of the parietal from the margin of the posttemporal fenestra (Wilson, 2002) and the distance between the supratemporal fenestrae exceeding twice the greatest dimension of the supratemporal fenestrae themselves (Salgado and Calvo, 1992).
As is typical for a number of ‘subadult’ diplodocoid skulls from the Morrison Formation (e.g., Suuwassea [Harris, 2006; Whitlock, 2011], Smitanosaurus [Whitlock and Wilson Mantilla, 2020], Kaatedocus [Tschopp and Mateus, 2013; Tschopp et al., 2015; Whitlock and Wilson Mantilla, 2020), CM 26552 displays characteristics of both Diplodocidae and Dicraeosauridae. The broad supraoccipital crest (Whitlock, 2011) and wide basal tubera (Wilson, 2002) suggest diplodocid affinities, but unlike the members of that group the supratemporal fenestrae of CM 26552 are not notably larger than the foramen magnum.
Characters indicating dicraeosaurid affinities are much more numerous, however. CM 26552 has both frontoparietal and postparietal fenestrae, features that diagnose various subclades of Dicraeosauridae (Whitlock, 2011; Whitlock and Wilson Mantilla, 2020). It has been suggested that these are possibly juvenile characteristics (Tschopp et al., 2015; Woodruff et al., 2017), but the absence of these fenestrae in very small diplodocids (i.e., SMM P84.15.3), and their presence in sexually mature dicraeosaurids (e.g., Windholz and Cerda, 2021) would seem to suggest the presence or absence of these fenestrae are of phylogenetic rather than ontogenetic significance. Other characteristics that seem to unite CM 26552 with most dicraeosaurids include the exclusion of the basioccipital from the dorsal margin of the occipital condyle by the exoccipitals (Tschopp et al., 2015), the presence of a distinct prong on the squamosal (Whitlock, 2011), the contribution of the frontal to the margin of the supratemporal fenestra (Wilson and Sereno, 1998), an expanded crista prootica (Wilson, 2002; Whitlock and Wilson Mantilla, 2020), and the flat distal margin of the paroccipital process (Whitlock and Wilson Mantilla, 2020). Additionally, the free dorsal margin of the antotic process (Harris and Dodson, 2004) is typical of dicraeosaurids; originally described as typical for Diplodocus by Berman and McIntosh (1978) based on CM 26552, it is now known only from dicraeosaurids. Finally, the presence of a shelf overhanging the foramen for the trigeminal nerve on the right side (and most probably also on the left) also aligns CM 26552 with dicraeosaurid affinities (Whitlock and Wilson Mantilla, 2020).
Within Dicraeosauridae, three coeval North American taxa can be compared to CM 26552: Kaatedocus, Smitanosaurus, and Suuwassea. Kaatedocus has been variously recovered as a diplodocid (Tschopp and Mateus, 2013; Tschopp et al., 2015, 2017) or a dicraeosaurid (Whitlock and Wilson Mantilla, 2020). In either event, Kaatedocus and CM 26552 differ in two significant ways: Kaatedocus lacks the frontoparietal fenestra and contribution of the frontal to the supratemporal fenestra seen in CM 26552. CM 26552 differs from Smitanosaurus by the broad supraoccipital crest in the former and the exclusion of the frontal from the supratemporal fenestra margin in the latter. CM 26552 differs from Suuwassea by the broad supraoccipital crest and the single (not double) abducens foramen seen in CM 26552, as well as most likely a greater size at skeletal maturity. Additionally, CM 26552 is potentially slightly stratigraphically younger (Carnegie Quarry dated to the Tithonian ca. 151-150 Ma [Maidment et al., 2017; Whitlock et al., 2018] versus the older Kimmeridgian Rattlesnake Ridge site that yielded Suuwassea [Myers and Storrs, 2007]). However, CM 26552 and Suuwassea do share a potential synapomorphic trait: the expanded contribution of the frontal to the supratemporal fenestra. CM 26552 and Suuwassea also share the presence of a patent interfrontal suture, although the ‘subadult’ developmental age of CM 26552 makes it unclear whether this is an ephemeral, ‘juvenile’ trait.
Relative to Argentinean dicraeosaurid skulls (e.g., those of Amargasaurus and Bajadasaurus), CM 26552 differs in the shallow depression at the base of the basisphenoid, the basal tubera wider than the occipital condyle, the absence of a columellar recess in the parabasal fossae, and the extended contribution of the frontal to the supratemporal fenestra. Alternatively, CM 26552 shares with Bajadasaurus and Amargasaurus the crest that divides the parabasal fossae, although in the latter two taxa its extension is reduced inside the columellar recess. In particular, CM 26552 and Bajadasaurus are distinguished by the presence of the following traits in the former: the anterodorsal tubercles in the orbitosphenoids, the presence of a dorsal spur in the paroccipital process and the exclusion of the supraoccipital from the posttemporal fenestrae, and the wide dorsal exposure of the supratemporal fenestrae. However, CM 26552 and Bajadasaurus share the ventral expansion of the orbitosphenoids in the incomplete rostral division of CN II’s, and the subequal diameter between the frontoparietal and postparietal fenestrae. CM 26552 differs from Amargasaurus by the conjoined CN II, and the extended dorsal exposure of the supratemporal fenestrae. Nevertheless, CM 26552 shares with Amargasaurus the anterodorsalmost tubercles and the anterior crest in the orbitosphenoids. Both the presence of a crest in the parabasal fossa and the paired tubercles in the anterior border of the orbitosphenoid are remarkable traits. These could represent novel information to be considered in future phylogenetic analyses.
Overall, within non-North American Dicraeosauridae, CM 26552 shares many non-synapomorphic traits with several taxa. CM 26552 shares with Bajadasaurus and Lingwulong a conjoined passage for the optic nerve (also shared with Kaatedocus, Smitanosaurus, and Suuwassea); with Amargasaurus and Dicraeosaurus a prominent dorsomedial tubercle of the orbitosphenoid; and with Amargasaurus and Lingwulong a dorsomedial spur in the exoccipital-opisthotic excluding the supraoccipital from the posttemporal fenestra (also shared with Kaatedocus, Suuwassea, and Smitanosaurus). This last character is probably also shared with Dicraeosaurus, in which a massive bony lamina separates the sagittal crest from the posttemporal fenestra.
CONCLUSIONS
CM 26552 can be assigned to the clade Dicraeosauridae based upon the following traits: 1) the presence of postparietal and frontoparietal fenestrae, 2) the exclusion of the basioccipital from the dorsal margin of the occipital condyle by the exoccipitals, 3) the presence of a distinct prong on the squamosal, 4) contribution of the frontal to the margin of the supratemporal fenestra, 5) an expanded crista prootica, 6) free dorsal margin of the antotic process, 7) the presence of a ‘shelf’ overhanging the foramen for the trigeminal (V) nerve, and 8) the flat distal margin of the paroccipital process. Although it bears only a single autapomorphy (the presence of a ‘tooth’ in the parietal/opisthotic suture), CM 26552 exhibits a mosaic of traits that enable it to be confidently differentiated from the other known North American dicraeosaurid taxa. Based on this, we assign CM 26552 to the new genus Athenar with the specific epithet bermani. Athenar shares with Suuwassea a frontal that contributes to the supratemporal fenestra, which may indicate close phylogenetic affinity between the two taxa.
Considering CM 26552 as a new dicraeosaurid species enhances the knowledge of the diversity of this family in the Morrison Formation and in North America more generally, expanding the historically underestimated sauropod diversity of the unit.
ACKNOWLEDGMENTS
We thank A. Henrici and S. Davis (Carnegie Museum of Natural History) for their help, guidance, and patience with extended loans for this specimen. D.C. Woodruff and an anonymous reviewer provided helpful reviews that greatly improved this manuscript. Special thanks to I.L. for support.
REFERENCES
Balanoff, A., Bever, G.S., and Ikejiri. T. 2010. The braincase of Apatosaurus (Dinosauria: Sauropoda) based on computed tomography of a new specimen with comments on variation and evolution in sauropod neuroanatomy. American Museum Novitates, 2010:1-32.
https://doi.org/10.1206/591.1
Berman, D.S. and McIntosh, J.S. 1978. Skull and relationships of the Upper Jurassic sauropod Apatosaurus (Reptilia, Saurischia). Bulletin of Carnegie Museum of Natural History 8:1-35.
https://doi.org/10.5962/p.228587
Calvo, J.O. and Salgado, L. 1995. Rebbachisaurus tessonei sp. nov. a new Sauropoda from the Albian-Cenomanian of Argentina; new evidence on the origin of the Diplodocidae. Gaia 33:13-33.
Canudo, J.I., Carballido, J.L., Garrido, A., and Salgado, L. 2018. A new rebbachisaurid sauropod from the Aptian-Albian, Lower Cretaceous Rayoso Formation, Neuquén, Argentina. Acta Palaeontologica Polonica 63: 679-691.
https://doi.org/10.4202/app.00524.2018
Gallina, P.A., Apesteguía, S., Canale, J.I., and Haluza, A. 2019. A new long-spined dinosaur from Patagonia sheds light on sauropod defense system. Scientific Reports 9:1392.
https://doi.org/10.1038/s41598-018-37943-3
Gallina, P.A., Apesteguía, S., Haluza, A., and Canale, J.I. 2014. A diplodocid sauropod survivor from the Early Cretaceous of South America. PLoS ONE 9:e97128-e97128.
https://doi.org/10.1371/journal.pone.0097128
Garderes, J.P., Gallina, P.A., Whitlock, J.A., and Toledo, N. 2021. Neuroanatomy of a diplodocid sauropod dinosaur from the Lower Cretaceous of Patagonia, Argentina. Cretaceous Research 129: 105024.
https://doi.org/10.1016/j.cretres.2021.105024
Garderes, J.P., Gallina, P.A., Whitlock, J.A., Toledo, N., 2023. Cranial osteology of Bajadasaurus pronuspinax (Sauropoda, Dicraeosauridae). Historical Biology 36, 1343-1367.
https://doi.org/10.1080/08912963.2023.2212389
Harris, J.D. 2006. Cranial osteology of Suuwassea emilieae (Sauropoda: Diplodocoidea: Flagellicaudata) from the Upper Jurassic Morrison Formation of Montana, USA. Journal of Vertebrate Paleontology 26:88-102.
https://doi.org/10.1671/0272-4634(2006)26[88:cooses]2.0.co;2
Harris, J.D. and Dodson, P. 2004. A new diplodocoid sauropod dinosaur from the Upper Jurassic Morrison Formation of Montana, USA. Acta Palaeontologica Polonica 49:197-210.
Hunt-Foster, R. 2024. Discovery, preservation, and protection of notable paleontological resources from Dinosaur National Monument, Utah and Colorado. Parks Stewardship Forum 40:243-256.
https://doi.org/10.5070/p540162933
Janensch, W. 1914. Übersicht über die Wirbeltierfauna der Tendaguru-Schichten nebst einer kurzen Charakterisierung der neu aufgeführten Arten von Sauropoden. Archiv für Biontologie, 3:81-110.
Jensen, J.A. 1985. Three new sauropod dinosaurs from the Upper Jurassic of Colorado. Great Basin Naturalist 45:697-709.
https://doi.org/10.5962/bhl.part.4439
Lerzo, L.N., Gallina, P.A., Canale, J.I., Otero, A., Carballido, J.L., Apesteguía, S., and Makovicky, P.J. 2024a. The last of the oldies: a basal rebbachisaurid (Sauropoda, Diplodocoidea) from the early Late Cretaceous (Cenomanian-Turonian) of Patagonia, Argentina. Historical Biology 37:208-233.
https://doi.org/10.1080/08912963.2023.2297914
Lerzo, L.N., Torcida Fernández-Baldor, F., Canale, J.I., Whitlock, J.A., Otero, A., and Gallina, P.A., 2024b. They all floated in the cretaceous: new rebbachisaurid (Sauropoda, Diplodocoidea) with a highly pneumatized skeleton from the Upper Cretaceous (lower Cenomanian) of Patagonia, Argentina. Historical Biology 37:1795–1808.
https://doi.org/10.1080/08912963.2024.2383708
Mannion, P.D., Upchurch, P., Mateus, O., Barnes, R.N., and Jones, M.E.H. 2012. New information on the anatomy and systematic position of Dinheirosaurus lourinhanensis (Sauropoda: Diplodocoidea) from the Late Jurassic of Portugal, with a review of European diplodocoids. Journal of Systematic Palaeontology 10:521-551.
https://doi.org/10.1080/14772019.2011.595432
Maidment, S. C. 2023. Diversity through time and space in the Upper Jurassic Morrison Formation, western USA. Journal of Vertebrate Paleontology, 43(5), e2326027.
https://doi.org/10.1080/02724634.2024.2326027
Maidment, S. C., and Muxworthy, A. 2019. A chronostratigraphic framework for the Upper Jurassic Morrison Formation, western USA. Journal of Sedimentary Research, 89(10): 1017-1038.
https://doi.org/10.2110/jsr.2019.54
Maidment, S.C.R., Balikova, D., and Muxworthy, A.R. 2017. Magnetostratigraphy of the Upper Jurassic Morrison Formation at Dinosaur National Monument, Utah, and prospects for using magnetostratigraphy as a correlative tool in the Morrison Formation. p. 279-302. In Ziegler, K.E. and Parker, W. (eds), Terrestrial Depositional Systems. Elsevier.
https://doi.org/10.1016/b978-0-12-803243-5.00007-8
Marsh, O.C. 1878. Principal characters of American Jurassic dinosaurs, Part I. American Journal of Science, 16: 411-416.
https://doi.org/10.2475/ajs.s3-16.95.411
Marsh, O.C. 1884. Principal characters of American Jurassic dinosaurs, Part VII, Diplodocidae, a new family of Sauropoda. American Journal of Science, 27:161-167.
https://doi.org/10.2475/ajs.s3-27.158.161
McIntosh, J.S., Coombs, Jr., W.P., and Russell, D.A. 1992. A new diplodocid sauropod (Dinosauria) from Wyoming, U.S.A. Journal of Vertebrate Paleontology 12:158-167.
https://doi.org/10.1080/02724634.1992.10011446
Moore A., and Mannion, P.D. 2024. Critical reappraisal of a putative dicraeosaurid sauropod dinosaur from the Middle Jurassic of Gondwana and a revised view of diplodocoid evolutionary relationships. Journal of Vertebrate Paleontology, Program and Abstracts, 2024:392-393.
Myers, T.S. and Storrs, G.W. 2007. Taphonomy of the Mother’s Day Quarry, Upper Jurassic Morrison Formation, south-central Montana, USA. Palaios 22:651-666.
https://doi.org/10.2110/palo.2005.p05-123r
Salgado, L. and Bonaparte, J.F. 1991. Un nuevo sauropod Dicraeosauridae, Amargasaurus cazaui gen. et sp. nov., de la Formación La Amarga, Neocomiano de la Provincia del Neuquén, Argentina. Ameghiniana 28:333-346.
Salgado L. and Calvo, J.O. 1992. Cranial osteology of Amargasaurus cazaui Salgado and Bonaparte (Sauropoda, Dicraeosauridae) from the Neocomian of Argentina. Ameghiniana 29: 337-346.
Seeley, H.G. 1887. On the Classification of the Fossil Animals Commonly Named Dinosauria. Proceedings of the Royal Society of London, 43, 165-171.
https://doi.org/10.1098/rspl.1887.0117
Sereno, P.C., Wilson, J.A., Witmer, L.M., Whitlock, J.A., Maga, A., Ide, O.A., and Rowe, T.A. 2007. Structural extremes in a Cretaceous dinosaur. PLoS ONE 2:e1230.
https://doi.org/10.1371/journal.pone.0001230
Torcida Fernández-Baldor, F., Canudo, J.I., Huerta, P., Montero, D., Pereda Suberbiola, X., and Salgado, L. 2011. Demandasaurus darwini, a new rebbachisaurid sauropod from the Early Cretaceous of the Iberian Peninsula. Acta Palaeontologica Polonica 56:535-552.
https://doi.org/10.4202/app.2010.0003
Tschopp, E. and Mateus, O. 2013. The skull and neck of a new flagellicaudatan sauropod from the Morrison Formation and its implication for the evolution and ontogeny of diplodocid dinosaurs. Journal of Systematic Palaeontology 11:853-888.
https://doi.org/10.1080/14772019.2012.746589
Tschopp, E. and Mateus, O. 2017. Osteology of Galeamopus pabsti sp. nov. (Sauropoda: Diplodocidae), with implications for neurocentral closure timing, and the cervico-dorsal transition in diplodocids. PeerJ 5:e3179.
https://doi.org/10.7717/peerj.3179
Tschopp, E., Mateus, O., and Benson, R.B.J. 2015. A specimen-level phylogenetic analysis and taxonomic revision of Diplodocidae (Dinosauria, Sauropoda). PeerJ 3:e857.
https://doi.org/10.7717/peerj.857
Upchurch, P. 1995. The evolutionary history of sauropod dinosaurs. Philosophical Transactions of the Royal Society of London, Series B: Biological Sciences, 349, 365-390.
https://doi.org/10.1098/rstb.1995.0125
Upchurch, P. 1998. The phylogenetic relationships of sauropod dinosaurs. Zoological Journal of the Linnean Society 124:43-103.
https://doi.org/10.1006/zjls.1997.0138
Upchurch, P., Barrett, P.M., and Dodson, P. 2004. Sauropoda, p. 259- 322. In Weishampel, D.B., Dodson, P., and Osmólska, H. (eds), The Dinosauria, 2nd edn. University of California Press, Berkeley.
https://doi.org/10.1525/california/9780520242098.003.0015
van der Linden, T.T.P., Tschopp, E., Sookias, R.B., Wallaard, J.J.W., Holwerda, F.M., and Schulp, A.S. 2024. A new diplodocine sauropod from the Morrison Formation, Wyoming, USA. Palaeontologia Electronica, 27(3):a50.
https://doi.org/10.26879/1380
Whitlock, J.A. 2011. A phylogenetic analysis of Diplodocoidea (Saurischia: Sauropoda). Zoological Journal of the Linnean Society 161:872-915.
https://doi.org/10.1111/j.1096-3642.2010.00665.x
Whitlock, J.A. and Wilson, J.A. 2013. Character distribution maps: a visualization method for comparative cladistics. Acta Zoologica 94:490-499.
https://doi.org/10.1111/azo.12006
Whitlock, J. A. and Wilson Mantilla, J.A. 2020. The Late Jurassic sauropod dinosaur ‘Morosaurus’ agilis Marsh, 1889 reexamined and reinterpreted as a dicraeosaurid. Journal of Vertebrate Paleontology 40:e1780600.
https://doi.org/10.1080/02724634.2020.1780600
Whitlock, J.A., Wilson, J.A., and Lamanna, M.C. 2010. Description of a nearly complete juvenile skull of Diplodocus (Sauropoda: Diplodocoidea) from the Late Jurassic of North America. Journal of Vertebrate Paleontology 30:442-457.
https://doi.org/10.1080/02724631003617647
Whitlock, J.A., Trujillo, K.C., and Hanik, G.M. 2018. Assemblage-level structure in Morrison Formation dinosaurs, Western Interior, USA. Geology of the Intermountain West 5:9-22.
https://doi.org/10.31711/giw.v5.pp9-22
Windholz, G.J. and Cerda, I.A. 2021. Paleohistology of two dicraeosaurid dinosaurs (Sauropoda; Diplodocoidea) from La Amarga Formation (Barremian-Aptian, Lower Cretaceous), Neuquén Basin, Argentina: paleobiological implications. Cretaceous Research, 128: 104965.
https://doi.org/10.1016/j.cretres.2021.104965
Wilson, J.A. 2002. Sauropod dinosaur phylogeny: critique and cladistic analysis. Zoological Journal of the Linnean Society 136:215-275.
https://doi.org/10.1046/j.1096-3642.2002.00029.x
Wilson, J.A. 2006. Anatomical nomenclature of fossil vertebrates: standardized terms or “lingua franca”? Journal of Vertebrate Paleontology 26:511-518.
https://doi.org/10.1671/0272-4634(2006)26[511:anofvs]2.0.co;2
Wilson, J.A. and Sereno, P.C. 1998. Early evolution and higher-level phylogeny of sauropod dinosaurs. Society of Vertebrate Paleontology Memoir 5:1-68.
https://doi.org/10.2307/3889325
Woodruff, D.C. 2019. What factors influence our reconstructions of Morrison Formation sauropod diversity? Geology of the Intermountain West, 6: 93-112.
https://doi.org/10.31711/giw.v6.pp93-112
Woodruff, D. C., Fowler, D.W., and Horner, J.R. 2017. A new multi-faceted framework for deciphering diplodocid ontogeny. Palaeontologia Electronica 20.3.43A: 1-53.
https://doi.org/10.26879/674
Xu, X., Upchurch, P., Mannion, P.D., Barrett, P.M., Regalado-Fernandez, O.R., Mo, J., Ma, J., and Liu, H. 2018. A new Middle Jurassic diplodocoid suggests an earlier dispersal and diversification of sauropod dinosaurs. Nature Communications 9:2700.
https://doi.org/10.1038/s41467-018-05128-1
