

 Wear-dependent molar morphology in hypsodont rodents: The case of the spalacine Pliospalax
Wear-dependent molar morphology in hypsodont rodents: The case of the spalacine Pliospalax
Article number: 26.3.a47
https://doi.org/10.26879/1322
Copyright Society of Vertebrate Paleontology, November 2023
Author biographies
Plain-language and multi-lingual abstracts
PDF version
Submission: 14 July 2023. Acceptance: 1 November 2023.
ABSTRACT
Studying the dental pattern of micromammals has proven to be a valuable tool in classifying and identifying extinct species, but problems may arise when dental morphology changes with wear. This holds particularly true for the high-crowned rodents, such as the Spalacinae. X-ray microtomography provides new opportunities to document the changes in dental morphology of the species with wear. Combining this technology with visualization and analysis software enables the examination of virtual occlusal surfaces of the molars mimicking the various wear stages. In this paper, we apply this technology on Pliospalax macoveii from the locality of Çalta in Turkey, P. tourkobouniensis and P. sotirisi from the Greek localities of Tourkobounia-1 and Maritsa, and the recent species Spalax microphthalmus from Hungary and Romania. This method enables the effective comparison between the spalacine species. As a result, we consider P. sotirisi as a junior synonym of P. macoveii.
Panagiotis Skandalos. Naturalis Biodiversity Center, P.O. Box 9517, 2300RA Leiden, The Netherlands. panos.skandalos@naturalis.nl
Lars W. van den Hoek Ostende. Naturalis Biodiversity Center, P.O. Box 9517, 2300RA Leiden, The Netherlands. lars.vandenhoekostende@naturalis.nl
Key words: Variation; Rodentia; Microtomography; Wear stage; Pliocene
Final citation: Skandalos, Panagiotis and van den Hoek Ostende, Lars W. 2023. Wear-dependent molar morphology in hypsodont rodents: The case of the spalacine Pliospalax Palaeontologia Electronica, 26(3):a47.
https://doi.org/10.26879/1322
palaeo-electronica.org/content/2023/4009-wear-dependent-pliospalax
Copyright: November 2023 Society of Vertebrate Paleontology.
This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
creativecommons.org/licenses/by/4.0
INTRODUCTION
Dental morphology plays an important role in mammal paleontology and particularly in the study of micromammals, based on which fossil species are mostly defined. The morphology of molar is very specific and in addition, molars have, in contrast to postcranial elements which may change over ontogeny, a relatively fixed morphology from the time they appear in juveniles. Thus, the combination of morphological and metrical data has been at the basis of the description for thousands of species of rodents, insectivores, lagomorphs and bats throughout the Cenozoic.
Of course, the morphology of a molar will change with wear. Generally, this is not a problem, as the wear pattern retains the original position of a cusp or crest. Moreover, species descriptions are often based on large assemblages that contain both old and younger individuals, so that features that disappear with wear can easily be identified. However, there are some groups in which the molar morphology changes dramatically with wear. Examples of this type are found in the high-crowned (hypsodont) molars of Castoridae (beavers), Hystricidae (porcupines), Rhizomyidae (bamboo rats) and Spalacinae (mole rats). Not all hypsodont molars necessarily show major changes with wear. The occlusal surfaces of voles, gerbils or lagomorphs, for instance, are relatively constant except for the very early stages of wear. But in groups that have a changing morphology depending on the individual age, taxonomical problems may arise (e.g., von Koenigswald, 1993; Lopatin, 2003; Van Weers, 2005).
This holds particularly true for the high crowned Spalacinae, which are the subject of this study. The fossil record of this subfamily is long and fairly continuous, dating back to the Late Eocene of southeastern Europe (De Bruijn et al., 2013; De Bruijn et al., 2015, De Bruijn et al., 2023). It comprises six genera: Vetusspalax De Bruijn et al., 2013, Debruijnia Ünay, 1996, Heramys Klein Hofmeijer and De Bruijn, 1985, Sinapospalax Sarıca and Şen, 2003, Pliospalax Kormos, 1932 and Spalax Güldenstädt, 1770. Although the subfamily appears in many rodent assemblages, it is usually represented by a few elements per locality only. Presumably, the "scarcity of fossils" is linked to the fossorial lifestyle, which makes spalacines a difficult target for birds of prey, one of the main agents for the accumulation of fossil remains (Andrews, 1990). Notably, the fossil range of the subfamily very much mirrors its current distribution (Ünay, 1999; De Bruijn et al., 2015; He et al., 2019).
The earliest genera of the Spalacinae already have high crowns, but hypsodonty increases over time towards the hyperhypsodont condition of the extant species (Sarıca and Şen 2003; Şen and Sarıca, 2011). Much of this increase in crown height occurs in Pliospalax. Ünay (1996, 1999) suggests separating Spalax from the older Pliospalax species using the different number of the reentrant folds in the lower first molars. Spalax has three while Pliospalax four to five. In addition, she cites De Bruijn and Van der Meulen (1975) who added the relatively shorter m1 and m3 and the posterior lophid of the latter not being isolated from the anterior part of the molar as typical for Spalax.
Sarıca and Şen (2003) separated the Middle and early Late Miocene species of Pliospalax into a genus of their own, Sinapospalax. Like Pliospalax, these species have four re-entrant folds and mostly differ from Pliospalax by their larger size and more complex occlusal surfaces. De Bruijn et al. (2023) did not recognize this genus, but here we follow the classification of Sarıca and Şen (2003) and Şen and Sarıca (2011). So, in our classification the oldest representatives of Pliospalax occurred at the end of the Late Miocene of eastern Europe and Anatolia (Şen and Sarıca, 2011). Pliospalax is present in Turkey with P. complicatus from Amasya (MN 13) (Şen and Sarıca, 2011) and P. macoveii from Çalta (MN 15) (Şen, 1977). It is also present in Ukraine with P. compositodontus in the MN 13 locality of Andreevka (Topachevski, 1969), in Romania with P. macoveii in the MN 15 locality of Mălușteni (Simionescu, 1930), and with the same species in Bulgaria in the MN 15 locality of Muselievo (Popov, 2004). In Greece, it is present in the MN 12 locality of Samos and the MN14 locality of Maritsa with P. sotirisi (De Bruijn et al., 1970; Vasileiadou and Sylvestrou, 2009), in the MN13/14 locality of Silata with P. macoveii (Vasileiadou et al., 2003), and in the MN 16 locality of Tourkobounia-1 with P. tourkobouniensis (De Bruijn and Van der Meulen, 1975).
Despite the frequent appearance of the genus in the fossil record, its relation with the extant Spalax is unclear (De Bruijn, 1984; Nevo et al., 1995; Sarıca and Şen, 2003; Şen and Sarıca, 2011) Here, we include in that genus Nannospalax Palmer, 1903 (Musser and Carleton, 2005), although recent molecular studies seem to favor keeping the two genera separate (e.g., Arslan et al., 2016; He et al., 2019). In literature, Microspalax [= Spalax ] is mentioned from the Pliocene of Ukraine (Topachevski, 1969) and the Early Pliocene (Ruscinian) of Greece with S. odessanus (Topachevski 1969) (De Bruijn, 1984). The localities that Topachevski (1969) mentions for this species, such as Kotlovina, Kryzhanivka, Kairy and Nogaisk, are currently considered to be of an Early Pleistocene age (Pushkina, personal. commun., May 2023). This is in line with the record of S. odessanus in the late Early Pleistocene Ukrainian localities of Tiligul Liman and Zhevakhova Gora 5 and 9 (Rekovets and Nadachowski, 1995). According to De Bruijn's (1984) illustrations, the Greek Early Pliocene occurrence of S. odessanus is mainly based on a senile specimen with heavily worn molars, which in our opinion even puts doubt on the generic classification.
Topachevski (1969, figure 60) and Şen (1977, plates 8 and 9) illustrated ontogenetic variations of molar pattern in Spalax odessanus from Odesa and Pliospalax macoveii from Çalta, respectively. Both studies were limited by the sample size and the available wear stages. However, X-ray microtomography allows us to study parts inside a molar that are hidden to the naked eye. Van Dam et al. (2011) used Micro computed tomography (Micro-CT) on micromammals in order to measure the enamel thickness of their molars and the hypsodonty based on volume indices. In this paper, we combine this technology with visualization and analysis softwares, in order to examine cross-cuts of molars as virtual occlusal surfaces, mimicking the various wear stages. In doing so, we aim to address taxonomical issues that appear due to wear and the shape of the molars, in order to review the distinction between the different Pliospalax species.
MATERIAL AND METHODS
The dental elements studied here include the lower first molars (m1) of Pliospalax macoveii (Simionescu, 1930) from Çalta in Turkey (Şen, 1977; ACA 858, ACA 859, ACA 881), stored in Muséum national d’ Histoire naturelle in France (MNHN), P. sotirisi (De Bruijn et al., 1970) from Maritsa (MR 1286) and P. tourkobouniensis De Bruijn and Van der Meulen, 1975 from Tourkobounia-1 in Greece (TB1 481, TB1-901), stored in the Department of Earth Sciences at Utrecht University and Spalax microphthalmus Güldenstaedt, 1770 from Hungary (REG 22729) and Romania (REG 22726), stored in Naturalis Biodiversity Center. The generic classification and the nomenclature of the molars follows Sarıca and Şen (2003, figure 6.1). In addition, the central transverse ridge oriented towards the labial border of the m1, is called the ectomesolophid following De Bruijn et al. (2023, figure 1).
The molars were scanned using a ZEISS Xradia 520 Versa equipment at Naturalis Biodiversity Center in The Netherlands and the AST-RX platform (Accès Scientifique à la Tomographie à Rayons X) of the MNHN. Subsequently, high resolution 3D models were made using the segmentation editor of the Avizo 2020.3.1 software. In order to explore the relationship between the morphological change of the occlusal surface on the Pliospalax species and wear, the ortho-slice module on the Z-axes of the 3D models was used to create cross-cuts of the molars. Since the wear surface of Spalacinae is flat, except for the very early stages of wear, these cross-cuts represent good representations of the wear surface at different heights. Thus, this method provides a complete reconstruction of the wear surface in subsequent slices, making it possible to observe the pattern changes in the enamel and dentine through the molar.
SYSTEMATIC PALEONTOLOGY
Order RODENTIA Bowdich, 1821
Family MURIDAE Illiger, 1811
Subfamily SPALACINAE Thomas, 1896
Genus PLIOSPALAX Kormos, 1932
Type Species. Pliospalax macoveii (Simionescu, 1930)
Sarıca and Şen (2003) were the last to revise the Spalacinae. According to them, the medium-sized genus Pliospalax is characterized by four reentrant folds; two in the labial and two in the lingual side of the m1. In addition, the molar has a rounded anterior part and a weak mesolophid. The protosinusids of the m1 and m2 are shallow and form enamel islands, even in younger specimens. The m2 mesolophid is weak or absent. The protocone of the M1 is entirely incorporated into the anteroloph and a weak mesoloph may be present. The M2 anterocone is also fused with the anteroloph and the mesoloph is completely fused with the paracone, surrounding the mesosinus as an enamel island. The species included in Pliospalax are P. macoveii (Simionescu, 1930); P. compositodontus Topachevski, 1969; P. sotirisi (De Bruijn et al., 1970), P. tourkobounensis De Bruijn and Van der Meulen, 1975, and Pliospalax complicatus Şen and Sarıca, 2011.
Pliospalax macoveii (Simionescu, 1930)
This species was described from the Romanian Pliocene locality of Mălușteni. Originally assigned to Prospalax by Simionescu (1930), it was selected as type species of Pliospalax by Kormos (1932). Topachevski (1969), in his research on the fauna from the USSR, considered Pliospalax synonymous to Microspalax, thus listing the species as M. macoveii. According to Topachevski (1969), the species lacks the mesoloph on the M1 and M2. The m1 of younger specimens have a free mesolophid that fuses with the metaconid in a later wear stage. In addition, the entoconid fuses with the posteroloph.
Pliospalax compositodontus (Topachevski, 1969)
The species was first described from the type locality of Andreevka (MN 13), in Ukraine (Topachevski, 1969). The Ukrainian spalacine was originally assigned to the genus Microspalax. Şen (1977) assigned the species to Pliospalax. However, later examination by the same author determined that some molars from Çalta had been mistakenly determined as P. compositodontus and they were in fact unworn m2 of P. macoveii. Finally, De Bruijn (1984), Sarıca and Şen (2003) and Şen and Sarıca (2011), assigned the species to Pliospalax.
Examining the occlusal surface of the molars, Topachevski (1969) distinguishes the species based on the well-developed mesoloph of the M1 which does not disappear throughout the different stages of wear. The mesolophid of the m1 is well developed, separated from the metaconid. The entoconid is not incorporated into the posterolophid (Topachevski, 1969).
Pliospalax sotirisi (De Bruijn et al., 1970)
The species was first described from the Greek locality of Maritsa (MN 14), on Rhodes island. De Bruijn et al. (1970) originally assigned the species to Spalax. However, five years after the first publication of the species, De Bruijn and Van der Meulen (1975) assigned the species to Pliospalax. The species is also known from the Greek locality of Samos (Vasileiadou and Sylvestrou, 2009). However, the latter assemblage is poor, comprising only three upper molars. In addition, Black et al. (1980) referred to P. cf. sotirisi an m2 and an m3 from the island of Samos. The mesosinusid of the m2 reaches the anterior border of the molar, and the anterosinus and the mesosinus of the M1 become enamel islands in a similar wear stage (De Bruijn et al., 1970).
Pliospalax tourkobouniensis De Bruijn and Van der Meulen, 1975
De Bruijn and Van der Meulen (1975) described Pliospalax tourkobouniensis from the Greek locality of Tourkobounia-1 (MN 16). The youngest of the Pliospalax species is characterized by the presence of a mesoloph on the M1 molar and traces of the posteroloph. In addition, in advanced wear stages of the m1, the protosinusid disappears and the mesosinusid becomes shallow. The sinusid does not reach the crown basis.
Pliospalax complicatus Şen and Sarıca, 2011
The latest described Pliospalax species, P. complicatus, comes from Amasya (MN 13), in Turkey (Şen and Sarıca, 2011). It is characterized by the fusion of the m1 anteroconid into the anterolophid. The metaconid is not part of the anterolophid and it connects with the anteroconid. The anterosinusid is present, the metalophulid is double and the protoconid does not have a posterior arm. The m1 has a well-developed mesolophid and M1 lacks the mesoloph. P. complicatus differs from all the species of Pliospalax by the presence of an anterosinusid on the m1 (Şen and Sarıca, 2011).
RESULTS
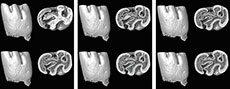
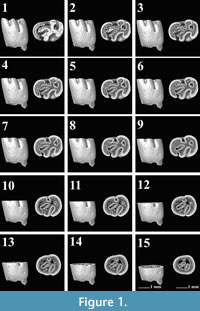 The m1 provides the highest number of characters in order to distinguish the different Pliospalax species (Şen and Sarıca, 2011, appendix 2). Therefore, we digitally mimicked the various wear stages of this element in order to recognize morphological differences and variations between the species above. Figure 1, Figure 2, Figure 3, Figure 4, Figure 5, Figure 6, Figure 7, Figure 8, Figure 9, Figure 10, Figure 11, Figure 12 give these digital wear stages, in which each subsequent stage is determined based on the depth where the different reentrant folds are closed, forming enamel islands, and where these islands disappear altogether.
The m1 provides the highest number of characters in order to distinguish the different Pliospalax species (Şen and Sarıca, 2011, appendix 2). Therefore, we digitally mimicked the various wear stages of this element in order to recognize morphological differences and variations between the species above. Figure 1, Figure 2, Figure 3, Figure 4, Figure 5, Figure 6, Figure 7, Figure 8, Figure 9, Figure 10, Figure 11, Figure 12 give these digital wear stages, in which each subsequent stage is determined based on the depth where the different reentrant folds are closed, forming enamel islands, and where these islands disappear altogether.
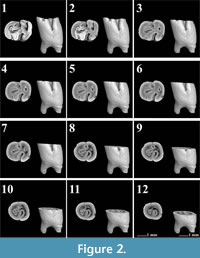 The first lower molars of Pliospalax macoveii from the locality of Çalta have four reentrant folds (Figure 1, Figure 2, Figure 3, Figure 4, representing ACA 858, ACA 859, ACA 881 right, and ACA 881 left, respectively) as Topachevski (1969) also described from the Romanian localities Mălușteni (type locality) and Bereşti. The anterior part of the P. macoveii molars varies in different individuals; the anterosinusid is absent in all of the specimens, but the left molar from ACA 881, has an anteriorly oriented opening between the metaconid and the anteroconid (Figure 4.1-4.3). The posterior metalophulid is either connected to anterolophulid (Figure 1.1-1.6) creating an enamel island, or it lacks this connection (Figure 2.1-2.3, Figure 3, Figure 4) leaving the anterior valley connected to protosinusid.
The first lower molars of Pliospalax macoveii from the locality of Çalta have four reentrant folds (Figure 1, Figure 2, Figure 3, Figure 4, representing ACA 858, ACA 859, ACA 881 right, and ACA 881 left, respectively) as Topachevski (1969) also described from the Romanian localities Mălușteni (type locality) and Bereşti. The anterior part of the P. macoveii molars varies in different individuals; the anterosinusid is absent in all of the specimens, but the left molar from ACA 881, has an anteriorly oriented opening between the metaconid and the anteroconid (Figure 4.1-4.3). The posterior metalophulid is either connected to anterolophulid (Figure 1.1-1.6) creating an enamel island, or it lacks this connection (Figure 2.1-2.3, Figure 3, Figure 4) leaving the anterior valley connected to protosinusid. 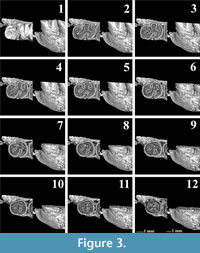 The anterior arm of the protoconid is well developed and in a later wear stage it connects to the labial anterolophid isolating the protosinusid. The mesolophid is short and either recognisable as a short ridge (Figure 1.1-1.3) or as a bulge on the hypolophulid (Figure 2.1-2.4, Figure 3.1-3.5, Figure 4.1-4.4). In both cases, with increased wear it fuses with the posterior metalophulid. In two out of the four molars (ACA 858, ACA 859), the posterosinusid is the first of the four reentrant folds to close. However, the number of enamel islands it creates varies.
The anterior arm of the protoconid is well developed and in a later wear stage it connects to the labial anterolophid isolating the protosinusid. The mesolophid is short and either recognisable as a short ridge (Figure 1.1-1.3) or as a bulge on the hypolophulid (Figure 2.1-2.4, Figure 3.1-3.5, Figure 4.1-4.4). In both cases, with increased wear it fuses with the posterior metalophulid. In two out of the four molars (ACA 858, ACA 859), the posterosinusid is the first of the four reentrant folds to close. However, the number of enamel islands it creates varies.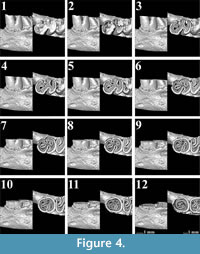 It has either a single, larger island (Figure 1.5-1.12) or two smaller ones (Figure 2.3-2.5). In the latter case, the lingual island is the smallest and the shallowest of the two. It disappears in later wear stages, before the labial island (Figure 2.6). The protosinusid is the second of the reentrant folds that closes (Figure 1.10, Figure 2.5). It becomes an enamel island and in a later wear stage it fades away. Two of the molars (ACA 881 right, ACA 881 left) have the protosinusid closing first followed by the posterosinusid (Figure 3.3-3.4, Figure 4.3-4.4). The closing of the deep mesosinusid follows after, persisting in later wear stages as an enamel island. The deepest of the reentrant folds is the sinusid. It closes in a very late wear stage forming an enamel island that reaches almost to the base of the molar (Figure 1.14-1.15, Figure 2.11-2.12, Figure 3.11-3.12, Figure 4.11-4.12).
It has either a single, larger island (Figure 1.5-1.12) or two smaller ones (Figure 2.3-2.5). In the latter case, the lingual island is the smallest and the shallowest of the two. It disappears in later wear stages, before the labial island (Figure 2.6). The protosinusid is the second of the reentrant folds that closes (Figure 1.10, Figure 2.5). It becomes an enamel island and in a later wear stage it fades away. Two of the molars (ACA 881 right, ACA 881 left) have the protosinusid closing first followed by the posterosinusid (Figure 3.3-3.4, Figure 4.3-4.4). The closing of the deep mesosinusid follows after, persisting in later wear stages as an enamel island. The deepest of the reentrant folds is the sinusid. It closes in a very late wear stage forming an enamel island that reaches almost to the base of the molar (Figure 1.14-1.15, Figure 2.11-2.12, Figure 3.11-3.12, Figure 4.11-4.12).
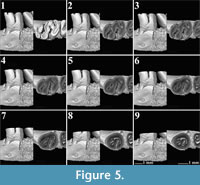 The m1 of Pliospalax tourkobouniensis is characterized by four reentrant folds (De Bruijn and Van der Meulen,1975; TB1-481 (holotype), Figure 5). However, one young specimen of P. tourkobouniensis (TB1-901; Figure 6) indicates the presence of a shallow anterosinusid (Figure 6.1), which separates the metaconid from the anterolophid. This additional sinusid increases the total number of reentrant folds to five. Following the wear stages of the molar, the posterosinusid forms either one (Figure 5.3-5.6) or two enamel islands (Figure 6.6-6.7);
The m1 of Pliospalax tourkobouniensis is characterized by four reentrant folds (De Bruijn and Van der Meulen,1975; TB1-481 (holotype), Figure 5). However, one young specimen of P. tourkobouniensis (TB1-901; Figure 6) indicates the presence of a shallow anterosinusid (Figure 6.1), which separates the metaconid from the anterolophid. This additional sinusid increases the total number of reentrant folds to five. Following the wear stages of the molar, the posterosinusid forms either one (Figure 5.3-5.6) or two enamel islands (Figure 6.6-6.7); the latter configuration resulting from the connection between the hypolophulid and the posterolophid. The anterosinusid is the first of the enamel folds to close with wear. The second fold to close, however, varies. The holotype of the species closes the posterosinusid and then, in a later stage of wear, the protosinusid (Figure 5.3), whereas in the second m1 the protosinusid closes first (Figure 6.1-6.4). The last and the deepest of the folds is the sinusid that stays open the longest (Figure 5.9, Figure 6.12). Between the hypolophulid and the posterior metalophulid we distinguish an additional fold of the enamel, indicating the presence of a short mesolophid that disappears quickly with wear (Figure 6.1-6.3). This character is not observed on the holotype.
the latter configuration resulting from the connection between the hypolophulid and the posterolophid. The anterosinusid is the first of the enamel folds to close with wear. The second fold to close, however, varies. The holotype of the species closes the posterosinusid and then, in a later stage of wear, the protosinusid (Figure 5.3), whereas in the second m1 the protosinusid closes first (Figure 6.1-6.4). The last and the deepest of the folds is the sinusid that stays open the longest (Figure 5.9, Figure 6.12). Between the hypolophulid and the posterior metalophulid we distinguish an additional fold of the enamel, indicating the presence of a short mesolophid that disappears quickly with wear (Figure 6.1-6.3). This character is not observed on the holotype.
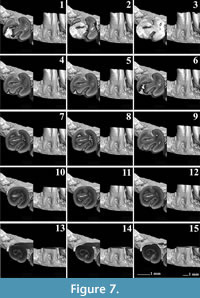 The m1 of Pliospalax sotirisi (Figure 7) has a distinct and short ridge that starts from the hypolophulid and meets the posterior metalophulid. Using the micro-CT photos, we distinguish the border between the enamel layers of the two crests (Figure 7.3-7.6). The ridge is similar to the hypolophid folds, present in older spalacine species like Debruijnia tintinnabulus (De Bruijn et al. 2023, figure 1). The longitudinal ridge in P. sotirisi is not a mesolophid but rather an anterior extension of the hypolophulid. In a later wear stage, it connects to the posterior metalophulid. The posterosinusid of the m1 is already closed on the only specimen available (Figure 7.1). The micro-CT pictures clearly show the connection between the enamel wall of the hypolophulid and posterolophid (Figure 7.2-7.6). This contact forms two enamel islands that will disappear in later wear stages. A similar morphology can be seen in P. tourkobouniensis (Figure 6.6). The closure of the posterosinusid is followed by that of the protosinusid (Figure 7.7-7.8). The mesosinusid comes next (Figure 7.10), with the sinusid, as the deepest of the folds, being the last that closes in a later stage of wear (Figure 7.14). However, since this specimen is the only m1 in the fossil record, it is not possible to distinguish whether these characters varied, like in Tourkobounia-1, or whether they are the rule.
The m1 of Pliospalax sotirisi (Figure 7) has a distinct and short ridge that starts from the hypolophulid and meets the posterior metalophulid. Using the micro-CT photos, we distinguish the border between the enamel layers of the two crests (Figure 7.3-7.6). The ridge is similar to the hypolophid folds, present in older spalacine species like Debruijnia tintinnabulus (De Bruijn et al. 2023, figure 1). The longitudinal ridge in P. sotirisi is not a mesolophid but rather an anterior extension of the hypolophulid. In a later wear stage, it connects to the posterior metalophulid. The posterosinusid of the m1 is already closed on the only specimen available (Figure 7.1). The micro-CT pictures clearly show the connection between the enamel wall of the hypolophulid and posterolophid (Figure 7.2-7.6). This contact forms two enamel islands that will disappear in later wear stages. A similar morphology can be seen in P. tourkobouniensis (Figure 6.6). The closure of the posterosinusid is followed by that of the protosinusid (Figure 7.7-7.8). The mesosinusid comes next (Figure 7.10), with the sinusid, as the deepest of the folds, being the last that closes in a later stage of wear (Figure 7.14). However, since this specimen is the only m1 in the fossil record, it is not possible to distinguish whether these characters varied, like in Tourkobounia-1, or whether they are the rule.
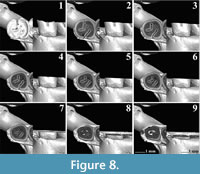 The m1s of an extant Spalax microphthalmus from Romania (REG 22726) belong to the same individual (Figure 8-Figure 9). For our analyses, we used both the right and the left m1. Both of them have a short posterior metalophulid ending freely in the mesosinusid.
The m1s of an extant Spalax microphthalmus from Romania (REG 22726) belong to the same individual (Figure 8-Figure 9). For our analyses, we used both the right and the left m1. Both of them have a short posterior metalophulid ending freely in the mesosinusid.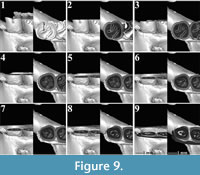 The mesolophid connects to the entoconid creating an enamel island between them (Figure 8.1-8.3, Figure 9.1-9.6). On the right m1, the latter disappears rapidly with wear, but it persists on the left one, almost to the base of the molar. Both m1s have two reentrant folds. The mesosinusid closes first (Figure 8.2, Figure 9.2-9.3). Towards the base of the molar, it is subdivided into two enamel islands (Figure 8.6, Figure 9.5). The sinusid is the deepest of the folds and it closes last (Figure 8.5, Figure 9.5).
The mesolophid connects to the entoconid creating an enamel island between them (Figure 8.1-8.3, Figure 9.1-9.6). On the right m1, the latter disappears rapidly with wear, but it persists on the left one, almost to the base of the molar. Both m1s have two reentrant folds. The mesosinusid closes first (Figure 8.2, Figure 9.2-9.3). Towards the base of the molar, it is subdivided into two enamel islands (Figure 8.6, Figure 9.5). The sinusid is the deepest of the folds and it closes last (Figure 8.5, Figure 9.5).
 The current research includes two m1 molars of Spalax microphthalmus from Hungary (REG 22729), which belong to the same individual (Figure 10, Figure 11). The short posterior metalophulid is preserved better on the left m1 (Figure 11.1-11.4). The right m1 has a short mesolophid (Figure 10.1-10.3) and an even shorter ectomesolophid (sensu De Bruijn et al. 2023, figure 1; Figure 10.1-10.2). The left m1 lacks both characters. The molars have two open reentrant folds, the mesosinusid and the sinusid. However, the wide posterior enamel island indicates the presence of an additional crest, the posterolophid
The current research includes two m1 molars of Spalax microphthalmus from Hungary (REG 22729), which belong to the same individual (Figure 10, Figure 11). The short posterior metalophulid is preserved better on the left m1 (Figure 11.1-11.4). The right m1 has a short mesolophid (Figure 10.1-10.3) and an even shorter ectomesolophid (sensu De Bruijn et al. 2023, figure 1; Figure 10.1-10.2). The left m1 lacks both characters. The molars have two open reentrant folds, the mesosinusid and the sinusid. However, the wide posterior enamel island indicates the presence of an additional crest, the posterolophid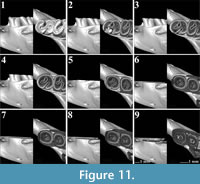 (Figure 10.1-10.5, Figure 11.1-11.3). When the mesosinusid is closed, the enamel island separates into two that disappear in a later wear stage (Figure 10.9, Figure 11.6-11.7). The sinusid closes last and it continues almost to the base of the molar.
(Figure 10.1-10.5, Figure 11.1-11.3). When the mesosinusid is closed, the enamel island separates into two that disappear in a later wear stage (Figure 10.9, Figure 11.6-11.7). The sinusid closes last and it continues almost to the base of the molar.
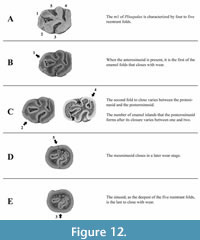 Studying the morphological changes on the occlusal surface of the m1 in different Spalacinae species, we distinguish four wear stages for Pliospalax (Figure 12, Table 1). The unworn stage is where the m1 is characterized by four to five enamel folds (Figure 12A). These are the anterosinusid (missing in some species), the protosinusid, the mesosinusid, the sinusid and the posterosinusid. In case an anterosinusid is present, it is the first fold to close with wear (Figure 12B). The next stage varies with species closing the protosinusid as the second and the posterosinusid as third fold or vice versa (Figure 12C). As we noted, both sequences can be found within one species. During this stage, the posterosinusid forms one or two enamel islands, depending on the connection between the hypolophulid and the posterolophid of the molar. The mesosinusid is the fourth fold that closes in a later wear stage (Figure 12D) with the sinusid, as the deepest of the five, being the last, closing near the base of the molar (Figure 12E). The studied Spalax species suggests that this genus follows the same wear stages but starting from an unworn stage (A) lacking the anterosinusid and protosinusid.
Studying the morphological changes on the occlusal surface of the m1 in different Spalacinae species, we distinguish four wear stages for Pliospalax (Figure 12, Table 1). The unworn stage is where the m1 is characterized by four to five enamel folds (Figure 12A). These are the anterosinusid (missing in some species), the protosinusid, the mesosinusid, the sinusid and the posterosinusid. In case an anterosinusid is present, it is the first fold to close with wear (Figure 12B). The next stage varies with species closing the protosinusid as the second and the posterosinusid as third fold or vice versa (Figure 12C). As we noted, both sequences can be found within one species. During this stage, the posterosinusid forms one or two enamel islands, depending on the connection between the hypolophulid and the posterolophid of the molar. The mesosinusid is the fourth fold that closes in a later wear stage (Figure 12D) with the sinusid, as the deepest of the five, being the last, closing near the base of the molar (Figure 12E). The studied Spalax species suggests that this genus follows the same wear stages but starting from an unworn stage (A) lacking the anterosinusid and protosinusid.
Applying X-ray microtomography on left and right molars from the same individuals allowed us to distinguish additional morphological variations other than those between species. Throughout the different wear stages, the development of the mesolophid varies between different species as well as between individual molars (e.g., Figure 1, Figure 2, Figure 11, Figure 12). In Spalax microphthalmus we even noted the absence of a mesolophid and ectomesolophid on the left molar. Notably, this mirrors the pattern of more advanced wear in the right m1 of the same individual.
DISCUSSION
According to von Koenigswald (1993), there are five ontogenetic phases in the development of the molar. The first two (A and B) create the occlusal surface (primary and secondary occlusal surface), the third one (C) the side walls, the fourth (D) the base of the crown, and the last one (E) the roots of the molar. However, the timing of each phase varies. Different heterochronies are observed between the low-crowned and hypsodont molars. The latter develop an extended C phase with shorter A and B phases. As a result, with wear, many morphological characters formed in the first two phases will rapidly disappear. The results of the current research largely confirm the observations of von Koenigswald (1993) on the hypsodont molars. Both the A and B phases of Pliospalax, form the occlusal surface of its molars. However, unlike the rest of the hypsodont species where the A and B phases are accelerated, in Pliospalax only the first is shortened. The B phase, responsible for the formation of the secondary occlusal surface of the molar, is prolonged, over a similar period or in some cases longer (e.g., P. sotirisi) than the C phase. The deep sinus in the m1s signifies the prolongation of the B phase. Thanks to these heterochronies the morphology of the occlusal surface of Pliospalax changes rapidly with wear. Different characters are affiliated to the different ontogenetic phases. The anterosinusid in the m1, when it is present, is the first of the enamel folds to disappear (Figure 12B) due to wear. This indicates that the character is related to the first ontogenetic phase (A), which rapidly gives its place to the more prolonged second phase (B) (von Koenigswald, 1993, figure 2). During the B phase, the protosinusid and the posterosinusid close first, although the order may vary (Figure 12C). The mesosinusid closes in a later wear stage with the sinusid, as the deepest of the five, closing last, signaling the end of the B phase (Figure 12D, 12E).
Different researchers studying the Spalacinae specified the number of the m1’s reentrant folds (e.g., De Bruijn and Van der Meulen, 1975; Kuss and Storch, 1978; Ünay 1996; Şen and Sarıca, 2011). Pliospalax is characterized by four of them (Sarıca and Şen, 2003). This character separates it from the genus Spalax, which has only three (Ünay 1996; 1999; Sarıca and Şen, 2003). However, a young specimen of Spalax nehringi Satunin, 1898 from the Greek island of Kalymnos was found to have four folds, including the protosinusid (Kuss and Storch, 1978). Similar divergences are observed in Pliospalax. One of the characters that distinguish Pliospalax complicatus from all the species of Pliospalax is the presence of the anterosinusid on the m1 (Şen and Sarıca, 2011). However, based on the current research, it is also present on the young representatives of P. tourkobouniensis. This character is present in older genera like Vetusspalax De Bruijn, Markovic and Wessels, 2013, Debruijnia Ünay, 1996 and Heramys Klein Hofmeijer and De Bruijn, 1985. Sinapospalax species lack the anterosinusid in the m1. However, Şen and Sarıca (2011) mention the presence of a notch between the m1 anteroconid and metaconid in S. canakkalensis (Ünay, 1981, plate 2, figure 2). The major evolutionary lineage begins with species having five m1 reentrant folds with Vetusspalax, Debruijnia and Heramys, then four with Sinapospalax and Pliospalax, and lastly three with Spalax. However, wear dependency seems to even influence the genus characters, with young specimens showing apparent atavisms like the reappearance of the m1’s anterosinusid and the ectomesolophid in the young individuals of Pliospalax and Spalax species. Alternatively, very worn specimens may lose a sinus on their wear surface which could lead to misidentifications. We believe this is illustrated by the presumed presence of Spalax in the Early Pliocene of Greece indicated by De Bruijn (1984). Spalax odessanus is known from the late Early Pleistocene of Ukraine (Rekovets and Nadachowski, 1995), but was originally dated to the Pliocene of Ukraine (Topachevski, 1969). De Bruijn (1984) placed his specimens from Karaburun in this species, extending its range to Late Ruscinian. However, this identification was based on worn specimens. The Greek locality includes two m1s. Based on their illustrations (De Bruijn, 1984, figure 1) we recognize a late phase C, of the wear stages, on the m1 from figure 1a and phase D for figure 1d. As we have seen, the protosinusid of Pliospalax has already disappeared at these stages. The late wear stage of the available m1s is insufficient to safely determine the species of the locality, even at the genus level. Taking into account the dependency of the dental morphology from the wear stage and the age of the locality, it seems likely that the Karaburun fossils belong to Pliospalax, although the species cannot be determined on the basis of the material.
The taxonomic problems of the genus Pliospalax begin from its type species, P. macoveii. From the drawings of Simionescu, it is obvious that the molars are in an advanced stage of wear (Simionescu, 1930, figure 31). Simionescu did not provide any diagnosis of the species, but limited himself to a comparison between the holotype and Prospalax rumanus Simionescu, 1930 from the same locality. Recently, Crespo et al. (2023) reviewed the Pliospalax specimens from Mălușteni and Bereşti. All the material belongs to a single species, Pliopalax macoveii and thus, Prospalax rumanus is considered a junior synonym of P. macoveii (Crespo et al., 2023).
Topachevski (1969) provided a detailed description of P. macoveii, but based in part on specimens from the localities of Bereşti in Romania and a mandible from Novopetrovka of Odesa in Ukraine, next to the material from the type locality of Mălușteni (Şen and De Bruijn, 1977). Given the taxonomic problems of the group, mixing material from localities far apart for a description is a risky enterprise. A complicating factor is the generally small size of spalacine assemblages, which implies that previous authors had to make identifications based on whatever wear stages were available in a particular locality.
Pliospalax macoveii was also described from the Pliocene locality of Çalta in Turkey (Şen, 1977). This well-defined assemblage became, given the problems with the type material, the main source of comparison between the different species and P. macoveii (e.g., Popov, 2004; Sarıca and Şen, 2003; Şen and Sarıca, 2011). In the character state matrix of Şen and Sarıca (2011), P. macoveii from the type locality of Mălușteni and that from Çalta have overall similar character states (Şen and Sarıca, 2011, appendix 2-3). Notably, these character states are also found in P. sotirisi from Maritsa. These similarities include the labial position of the anteroconid on m1 that connects through a lophid to the metaconid, as well as their relative size, with the metaconid being slightly bigger than the anteroconid (Şen and Sarıca, 2011, appendix 3, n. 1-4). In addition, in all these assemblages the mesolophid of the m1 is absent or weakly developed, the anterolophulid is complete, and the protosinusid present (Şen and Sarıca, 2011, appendix 3, n. 6-7, 9 and 13). The only morphological characters that differ are observed in the connection between the protoconid and the anteroconid, and the independence of the anterolophid, on the m1, the occurrence of the posterosinusid on the m2, the presence of the protosinusid and the occurrence of the anterosinusid on the m3 (Şen and Sarıca, 2011, appendix 3, n. 5, 8, 25, 27, 30). In Maritsa, the protoconid of the m1 connects to the anteroconid only through the anterolophulid, compared to the other two localities where it connects by the anterolophid or the labial anterolophid. Furthermore, in Çalta the anterolophid is either distinct or fused to the protoconid, while in Mălușteni and Maritsa it is distinct. However, the results from the current research from P. macoveii from Çalta, show that the morphology of the anterior part of the m1 varies between the different individuals. The posterosinusid of the m2 is open in all three localities, but in Çalta, it can likewise be an enamel island. Even though our research is focused on the first lower molars of the different Pliospalax species, the wear dependency of characters is the same for all elements. Thus, the posterosinusid, like every reentrant fold, transforms into an enamel island in later wear stages. Furthermore, the protosinusid and the anterosinusid on the m3 are absent in Mălușteni but present in Çalta. Unfortunately, these characters are not possible to be observed in Maritsa’s material, since no m3 has been recovered. De Bruijn and Van der Meulen (1975) stated that “two species of Pliospalax are known: P. macoveii and P. sotirisi. These species are closely related and may be synonymous”. Based on the work of Şen and Sarıca (2011) and the current research, which underline the morphological change of the occlusal surface with wear, and the importance of the morphological variation in the Pliospalax genus, we consider P. sotirisi as a younger synonym of P. macoveii. Observing its wear stage, the m1 from Maritsa belongs to an early C phase that does not provide a safe determination of the species. Its sinus is deeper than P. macoveii; which indicates a longer ontogenetic B phase. However, these observations do not warrant separating the Maritsa assemblage as an independent species.
CONCLUSION
Using micro-CT, wear stages can be reconstructed in hypsodont molars, as is shown for the spalacine species Pliospalax macoveii, P. sotirisi, P. tourkobouniensis, and Spalax microphthalmus. The different wear stages can in part be correlated to the ontogenetic phases in hypsodont molars proposed by von Koenigswald (1993), except for the B phase. The formation of sinuses and sinusids can be interpreted as a continuation of the B phase, while in other parts of the molar the side walls (C phase) are formed.
Comparison of the different wear stages shows a difference in the timing of the different sinuses in different species. Overall, this leads to a reduction of the number of sinusids over time, from five in the older genera to four in Sinapospalax and Pliospalax and three in the extant Spalax (s.l.). Young specimens can show apparent atavisms, showing traces of a sinusid that disappear quickly with wear. This was noted for the presence of an anterosinusid in a relatively unworn specimen of P. tourkobouniensis and the presence of four folds in a Spalax nehringi from Kalymnos.
Using virtual wear stages provides important data for taxonomy. This holds especially true for spalacines, which, with a few exceptions, are found in very limited numbers in fossil assemblages. Virtual reconstruction can give a better impression of the variation and allows us to compare material in similar wear stages. Based on our results, we consider P. sotirisi as a younger synonym of P. macoveii.
ACKNOWLEDGMENTS
This research has been partly financed by the Synthesys+ (FR-TAF_Call4_072) grant. We would like to thank Ş. Şen and G. Billet from Muséum national d'Histoire naturelle in Paris for their hospitality and assistance during the visit of the first author in the rich collections of the Museum. We express our gratitude to the Department of Geology of Utrecht University in The Netherlands and especially to W. Wessels for her constant help by giving us access to the rich collections of her department. Finally, we thank R. Langelaan from Naturalis Biodiversity Center for his help to scan the material of the current research.
REFERENCES
Andrews, P. 1990. Small mammal taphonomy. In Lindsay, E.H., Fahlbusch, V., and Mein, P. (ed.), European Neogene Mammal Chronology. NATO Advanced Study Institute Series Life Science, New York, A 180:487–494.
https://doi.org/10.1007/978-1-4899-2513-8_28
Black, C.C., Krishtalka, L., and Solounias, N. 1980. Mammalian fossils of Samos and Pikermi. Part 1. The Turolian rodents and insectivores of Samos. Annals of Carnegie Museum, 49(2): 359–378.
Crespo, V. D., Vasile, Ș., Petculescu, A., Rățoi, B.G., and Haiduc, B.S. 2023. The Early Pliocene small mammals (Eulipotyphla, Rodentia, Lagomorpha) from Berești and Mălușteni (eastern Romania): a fresh look at old collections. Earth and Environmental Science Transactions of the Royal Society of Edinburgh, 114:49–63.
https://doi.org/10.1017/S175569102200024X.
De Bruijn, H. 1984. Remains of the mole-rat Microspalax odessanus Topachevski, from Karaburun (Greece, Macedonia) and the family Spalacidae. Proceedings Koninklijke Nederlandse Akademie van Wetenschappen, B 87(4):417–425.
De Bruijn, H. and Van der Meulen, A.J. 1975. The early Pleistocene rodents from Tourkobounia-1 (Athens, Greece). Proceedings Koninklijke Nederlandse Akademie van Wetenschappen, B 78:314–338.
De Bruijn, H., Dawson, M.R., and Mein, P. 1970. Upper Pliocene Rodentia, Lagomorpha and Insectivora (Mammalia) from the isle of Rhodes (Greece). Proceedings of the Koninklijke Nederlandse Akademie van Wetenschappen, B 73:525–584.
De Bruijn, H., Marković, Z., and Wessels, W. 2013. Late Oligocene rodents from Banovići (Bosnia and Herzegovina). Palaeodiversity, 6:63–105.
De Bruijn, H., Bosma, A.A., and Wessels, W. 2015. Are the Rhizomyinae and the Spalacinae closely related? Contradistinctive conclusions between genetics and palaeontology. Palaeobiodiversity and Palaeoenvironments, 95:257–269.
https://doi.org/10.1007/s12549-015-0195-y
De Bruijn, H., Marković, Z., Wessels, W., and Weerd, A.A. van de 2023. On the antiquity and status of the Spalacidae, new data from the late Eocene of south-East Serbia. Palaeobiodiversity and Palaeoenvironments, 103(2):433–445.
https://doi.org/10.1007/s12549-022-00529-z
Güldenstädt, A.J. 1770. Spalax, novum glirium genus. Novi Commentarii. Academiae Scientiarum Imperialis Petropolitanae, 14(1):409–440.
He, Y., Hu, S., Ge, D., Yang, Q., Connor, T., and Zhou, C. 2019. Evolutionary history of Spalacidae inferred from fossil occurrences and molecular phylogeny. Mammal Review, 50(1):11–24.
https://doi.org/10.1111/mam.12170
Illiger, C. 1811. Prodromus systematis mammalium et avium additis terminis zoographicis utriusque classis, eorumque versione germanica. Salfeld C. Berlin, Germany, p. 301.
Klein Hofmeijer, G. and De Bruijn, H. 1985. The mammals from the Lower Miocene of Aliveri (island of Evia, Greece). Part 4. The Spalacidae and Anomalomyidae. Proceedings Koninklijke Nederlandse Akademie van Wetenschappen, B 88(2):185–198.
Kormos, T. 1932. Neue pliozäne Nagetiere aus der Moldau. Paläontologische Zeitschrift, 14:193–200.
Kuss, S.E. and Storch, G. 1978. Eine Säugetierfauna (Mammalia: Artiodactyla, Rodentia) des älteren Pleistozäns von der Insel Kalymnos (Dodekanes, Griechenland). Neues Jahrbuch für Geologie und Paläontologie, Monatshefte, 4:206–227.
Lopatin, A.V. 2003. The revision of the Early Miocene beavers (Castoridae, Rodentia, Mammalia) from the North Aral Region. Russian Journal of Theriology, 2(1):15–25.
Musser, G.G. and Carleton, M.D. 2005. Superfamily Muroidea. p. 894–1599. In Wilson, D.E. and Reeder, D.M. (ed.), Mammal species of the world. Johns Hopkins University Press, 3rd ed., Baltimore (MD).
Nevo, E., Filippucci, M.G., Redi, C., Simson, S., Heth, G., and Beiles, A. 1995. Karyotype and genetic evolution in speciation of subterranean mole rats of the genus Spalax in Turkey. Biological Journal of Linnean Society, 54:203–229.
https://doi.org/10.1111/j.1095-8312.1995.tb01034.x
Palmer, T.S. 1903. Some new generic names of mammals. Science, 17:873.
Popov, V. 2004. Pliocene small mammals (Mammalia, Lipotyphla, Chiroptera, Lagomorpha. Rodentia) from Muselievo (North Bulgaria). Geodiversitas, 26:403–491.
Rekovets, L. and Nadachowski, A. 1995. Pleistocene voles (Arvicolidae) of Ukraine. Paleontologia i Evolució 28:145–245.
Sarıca, N. and Şen, S. 2003. Spalacidae, p. 141–163. In Fortelius, M., Kappelman, J., Şen. S., and Bernor, R.L. (ed.), Geology and paleontology of the Miocene Sinap Formation, Turkey. Columbia University Press, New York.
Şen, S. 1977. La faune de rongeurs pliocènes de Çalta (Ankara, Turquie). Bulletin du Muséum National d’Histoire Naturelle, Sciences de la Terre. 61:89–171.
Şen, S. and De Bruijn, H. 1977. Comparaison des Rongeurs du Néogène terminal de Maritsa, Çalta et Tourkobounia 1, Méditerranée orientale. Proceedings of the Koninklijke Nederlandse Akademie van Wetenschappen, Amsterdam, B 80(3):222–227.
Şen, S. and Sarica, N. 2011. Middle-Late Miocene Spalacidae (Mammalia) From Western Anatolia, and the Phylogeny of the Family. Bulletin of the Earth Sciences Application and Research Centre of Hacettepe University. Yerbilimleri. 32(1):21–50.
Simionescu, I. 1930. Vertebratele Pliocene de la Malusteni (Covurlui). Les Vertébrés Pliocène de Malusteni (Roumanie). Academia Romana. Publicatiunile Fondului Vasile Adamachi. București: Cultura Nationala. 9(5):83–151.
Thomas, O. 1896. On the genera of rodents: an attempt to bring up to date the current arrangement of the order. Proceedings of the Zoological Society of London, p. 1012–1028.
Topachevski, V.O. 1969. Fauna of the USSR: Mammals, mole rats, Spalacidae. Akademia Nauk SSSR. 99:1–247. (In Russian)
Ünay, E. 1981. Middle and Upper Miocene rodents from the Bayraktepe section (Çanakkale, Turkey). Proceedings of the Koninklijke Nederlandse Akademie van Wetenschappen, B 84:217–238.
Ünay, E. 1996. On fossil Spalacidae (Rodentia), p. 246–252. In Bernor, R.L., Falbusch, V., and Mittmann, H.W. (ed.), The Evolution of Western Eurasian Neogene Mammal Faunas. Columbia University Press, New York.
Ünay, E. 1999. Family Spalacidae. p. 421–425. In Rössner, G.E. and Heissig, K. (ed.), The Miocene land mammals of Europe, Verlag Dr. F. Pfeil, München.
Van Dam, J.A., Fortuny, J., Ruijven, L.J. van 2011. MicroCT-scans of fossil micromammal teeth: re-defining hypsodonty and enamel proportion using true volume. Palaeogeography, Palaeoclimatology, Palaeoecology, 311:103–110.
https://doi.org/10.1016/j.palaeo.2011.08.010
van Weers, D.J. 2005. A taxonomic revision of the Pleistocene Hystrix (Hystricidae, Rodentia) from Eurasia with notes on the evolution of the family. Contributions to Zoology, 74(3-4):301–312.
https://doi.org/10.1163/18759866-0740304007
Vasileiadou, K., Koufos, G.D., and Syrides, G.E. 2003. Silata, a new locality with micromammals from the Miocene/Pliocene boundary of the Chalkidiki peninsula, Macedonia, Greece. Deinsea, 10:549–562.
Vasileiadou, K. and Sylvestrou, I.A. 2009. The Late Miocene mammal faunas of the Mytilinii Basin, Samos Island, Greece: new collection. 4. Micromammals. Beiträge zur Paleaontologie, 31:189–205.
von Koenigswald, W. 1993. Heterochronies in Morphology and Schmelzmuster of hypsodont molars in the Muroidea (Rodentia). Quaternary International, 19:57–61.
https://doi.org/10.1016/1040-6182(93)90023-9
von Satunin, K. 1898. Spalax nehringi nov. spec. Zoologischer Anzeiger, 21:314.
