

 Paleoecology and taphonomy of spatangoid echinoid-produced burrows (Scolicia) in slope and basin floor deposits from the Eocene of Cuba
Paleoecology and taphonomy of spatangoid echinoid-produced burrows (Scolicia) in slope and basin floor deposits from the Eocene of Cuba
Article number: 28.1.a4
https://doi.org/10.26879/1271
Copyright Paleontological Society, January 2025
Author biographies
Plain-language and multi-lingual abstracts
PDF version
Submission: 20 January 2023. Acceptance: 20 December 2024.
ABSTRACT
The morphological variability in the preservation of the trace fossil Scolicia is mainly taphonomic and results from different factors. However, only a few papers have discussed this issue using a taphonomical approach. In addition, although Scolicia is commonly present in deep-sea and shelf ichnoassemblages, its record in slope deposits is scarce. Herein, variations in the preservation of Scolicia are discussed in slope and basin floor contexts from the Eocene Capdevila Formation, western Cuba. The studied specimens come from two outcrops located in the Artemisa and Pinar del Río regions. The burrows were assigned to Scolicia prisca, Scolicia isp. var. vertebralis, and Scolicia isp. var. laminites. The observed variations in the diagnostic features of Scolicia reflect the wide variation expected in burrows made by spatangoid echinoids. Furthermore, we demonstrated that S. vertebralis could be a preservational variant of S. prisca and that S. prisca could be produced as furrows by extant echinoids, and not necessarily represent eroded burrows. In the Artemisa section, Scolicia is associated with basin floor deposits in bathyal context and form part of the Nereites ichnofacies. In the Pinar de Río section, Scolicia occur in shallower slope deposits and is included in the impoverished Cruziana ichnofacies. Their presence in slope deposits could be due to different factors, such as the predominance of sandy substrates, high sedimentation rate, and good oxygenation, which also led to an absence of trace fossils of the Zoophycos ichnofacies, which is typical of slope settings.
Jorge Villegas-Martín. 1 Technological Institute of Paleoceanography and Climatic Changes – itt OCEANEON, Unisinos University, Av. Unisinos, 950, 93022-000, São Leopoldo, Rio Grande do Sul Brazil; jvillegasmartin@gmail.com and Ichnos Research Group, Geology Graduate Program, Unisinos University, Av. Unisinos 950, 93022-000, São Leopoldo, Rio Grande do Sul Brazil. (Corresponding author)
Renata G. Netto.Ichnos Research Group, Geology Graduate Program, Unisinos University, Av. Unisinos 950, 93022-000, São Leopoldo, Rio Grande do Sul Brazil. nettorg@unisinos.br
Reinaldo Rojas-Consuegra. Centro de Investigación del Petróleo. Churruca No 481, Cerro, La Habana, Cuba, CP 12000. rojas@ceinpet.cupet.cu
Jordi M. de Gibert. Deceased on September 23, 2012.
Keywords: trace fossils; Scolicia preservational variations; Capdevila Formation; early Eocene
Final citation: Villegas-Martín, Jorge, Netto, Renata G., Rojas-Consuegra, Reinaldo, de Gibert, Jordi M. 2025. Paleoecology and taphonomy of spatangoid echinoid-produced burrows (Scolicia) in slope and basin floor deposits from the Eocene of Cuba. Palaeontologia Electronica, 28(1):a4.
https://doi.org/10.26879/1271
palaeo-electronica.org/content/2025/5428-spatangoid-echinoid-burrows
Copyright: January 2025 Paleontological Society
This is an open access article distributed under the terms of Attribution-NonCommercial-ShareAlike 4.0 International (CC BY-NC-SA 4.0), which permits users to copy and redistribute the material in any medium or format, provided it is not used for commercial purposes and the original author and source are credited, with indications if any changes are made.
creativecommons.org/licenses/by-nc-sa/4.0/
INTRODUCTION
Spatangoids constitute a large group of irregular echinoids and key bioturbators that rework the substrate during deposit-feeding activity and locomotion in modern shallow to deep marine settings (e.g., Bromley et al., 1997; Thompson and Riddle, 2005). The burrowing activity of extant spatangoid echinoids is well-known from natural observations and aquarium experiments (Nichols, 1959; Chesher, 1963; Howard et al., 1974; Bromley et al., 1997; Belaústegui et al., 2017), resulting in distinctive bioturbation structures that characterize the ichnogenera Scolicia (De Quatrefages, 1849), Cardioichnus (Smith and Crimes, 1983), Bichordites (Plaziat and Mahmoudi, 1988), and Sursumichnus (Uchman et al., 2022). Scolicia is interpreted as the grazing activity of spantagoid echinoids with double drainage tubes (e.g., Ward and Lewis, 1975; Bromley and Asgaard, 1975; Smith and Crimes, 1983; Bromley et al., 1997; Belaústegui et al., 2017), while Bichordites represents those produced by spantagoids with a single drainage tube (e.g., Plaziat and Mahmoudi, 1988; Villegas-Martín et al., 2014; Belaústegui et al., 2017). Cardioichnus is a heart-shaped depression (e.g., Smith and Crimes, 1983; Kappus and Lucas, 2019; Naimi et al., 2021), indicating a resting behavior, and can occur associated with Bichordites and Scolicia, forming compound trace fossils (e.g., Smith and Crimes, 1983; Mayoral and Muñiz, 2001; Bernardi et al., 2010; Belaústegui et al., 2017; Kappus and Lucas, 2019). Sursumichnus is also related to resting traces associated with Scolicia burrows (Uchman et al., 2022).
The ichnogenus Scolicia includes a variety of morphologies that were originally attributed to distinct ichnogenera and, posteriorly, re-assigned to it after careful ichnotaxonomic revisions; examples include specimens assigned to Psammichnites Torell, 1870, Subphyllocorda Götzinger and Becker, 1932, and Palaeobullia Götzinger and Becker, 1932, among others (e.g., Häntzchel, 1975; Ksiażkiewicz, 1970, 1977; Smith and Crimes, 1983; Plaziat and Mahmoudi, 1988; Uchman, 1995; Belaústegui et al., 2017). Nevertheless, some complex morphological features can still be recognized as different preservational variations of Scolicia. This variability in the Scolicia preservation is mainly taphonomic and results from factors such as the burrow position within the sediment bed, erosion, and lithological variability (Plaziat and Mahmoudi, 1988; Uchman, 1995; Gibert and Goldring, 2008). However, only a few papers discussed this topic with respect to taphonomy (Bromley and Asgaard, 1975; Smith and Crimes, 1983; Plaziat and Mahmoudi, 1988; Uchman, 1995; Fu and Werner, 2000).
Distinct preservational variations of Scolicia represent the most common trace fossil recorded in upper slope and basin floor (bathyal context) deposits of the Eocene Capdevila Formation, in western Cuba (Villegas-Martín, 2009; Villegas-Martín et al., 2014). Although Scolicia has been registered exclusively in marine settings, ranging from shelf to deep sea areas (e.g., Tchoumatchenco and Uchman, 2001; Wetzel, 2008; Giannetti, 2010; Riahi et al., 2014; Villegas-Martin et al., 2014; Carmona et al., 2020), its occurrence in slope deposits is uncommon so far (Fu and Werner, 2000; Wetzel, 2008; Demircan and Uchman, 2017). The presence of Scolicia in both shallower slope and basin floor deposits in western Cuba opens a window to discuss the triggers that potentially control the distribution of spatangoid burrows in distinct marine benthic regions. Thus, the aim of this paper is twofold: (i) to discuss the taphonomic control in the variability of Scolicia morphologies; and (ii) to evaluate the paleoecological parameters that favor spatangoid distribution throughout the sea bottom.
GEOLOGICAL SETTING
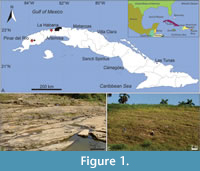 The Scolicia specimens studied herein come from two sections of the Capdevila Formation: i) the Artemisa section (near La Habana, Artemisa Province), which is part of the Havana-Matanzas Anticlinal infill; and ii) the Pinar del Río section (Pinar del Río Province), which is part of the sedimentary infill of Los Palacios and Bahía Honda basins (e.g., Brönnimann and Rigassi, 1963; Albear and Iturralde-Vinent, 1985; Bralower and Iturralde-Vinent, 1997; García-Delgado and Torres-Silva, 1997; Gordon et al., 1997) (Figure 1). Foraminiferal associations and nannofossil zonations (NP-11 and NP-12 ODP zones, Site 865) place the Capdevila Formation deposits mainly in the Eocene (Brönnimann and Stradner, 1960; Brönnimann and Rigassi, 1963; Bralower and Iturralde-Vinent, 1997).
The Scolicia specimens studied herein come from two sections of the Capdevila Formation: i) the Artemisa section (near La Habana, Artemisa Province), which is part of the Havana-Matanzas Anticlinal infill; and ii) the Pinar del Río section (Pinar del Río Province), which is part of the sedimentary infill of Los Palacios and Bahía Honda basins (e.g., Brönnimann and Rigassi, 1963; Albear and Iturralde-Vinent, 1985; Bralower and Iturralde-Vinent, 1997; García-Delgado and Torres-Silva, 1997; Gordon et al., 1997) (Figure 1). Foraminiferal associations and nannofossil zonations (NP-11 and NP-12 ODP zones, Site 865) place the Capdevila Formation deposits mainly in the Eocene (Brönnimann and Stradner, 1960; Brönnimann and Rigassi, 1963; Bralower and Iturralde-Vinent, 1997).
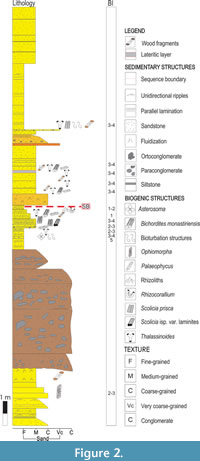 The Capdevilla Formation mainly consists of mudstones, limestones, siltstones, and massive and parallel-laminated fine to very coarse-grained sandstones, which can be intercalated with marls, carbonates, and matrix- or grain-supported conglomerates (e.g., Brönnimann and Rigassi, 1963; Piotrowski, 1987; Brust et al., 2011; Villegas-Martín et al., 2014). Tuffites and carbonatic concretions have also been reported at some localities (Piotrowski, 1987; Brust et al., 2011). Scolicia is preserved in medium- to fine-grained sandstones in the middle part of the Artemisa section. In the Pinar del Río section, Scolicia occurs in medium- to fine-grained parallel stratified sandstones (Figure 2) that characterizes the middle and upper part of the sedimentary succession (Villegas-Martín et al., 2014).
The Capdevilla Formation mainly consists of mudstones, limestones, siltstones, and massive and parallel-laminated fine to very coarse-grained sandstones, which can be intercalated with marls, carbonates, and matrix- or grain-supported conglomerates (e.g., Brönnimann and Rigassi, 1963; Piotrowski, 1987; Brust et al., 2011; Villegas-Martín et al., 2014). Tuffites and carbonatic concretions have also been reported at some localities (Piotrowski, 1987; Brust et al., 2011). Scolicia is preserved in medium- to fine-grained sandstones in the middle part of the Artemisa section. In the Pinar del Río section, Scolicia occurs in medium- to fine-grained parallel stratified sandstones (Figure 2) that characterizes the middle and upper part of the sedimentary succession (Villegas-Martín et al., 2014).
Microfossils, corals, echinoderm fragments, bivalves, brachiopods, and small fragments of macroscopic plants and red algae encompass the paleontological record of the Capdevila Formation (Cushman and Bermudez, 1949; Brönnimann and Stradner, 1960; Brönnimann and Rigassi, 1963; Piotrowski, 1987; Bralower and Iturralde-Vinent, 1997; Brust et al., 2011). Preliminary ichnological studies in the Artemisa region revealed the existence of a diverse trace fossil assemblage including Cosmorhaphe isp., Helminthorhaphe isp., Ophiomorpha isp., Paleodictyon isp., Planolites isp., ? Psammichnites isp., Scolicia isp., and Thalassinoides isp. (Villegas-Martín, 2009; Rojas-Consuegra et al., 2018). These deposits have been assumed to be a bathyal marine sequence formed during an active tectogenesis period. Turbiditic currents carried massive amounts of terrigenous materials into the basin, forming a flysch deposits (Albear and Iturralde-Vinent, 1985; Lex, 2013).
The deposits of the Capdevila Formation in the Pinar del Río section, otherwise, represent upper slope to shelf settings, based on sedimentological, micropaleontological, and ichnological data (Brust et al., 2011; Villegas-Martín et al., 2014). The trace fossil assemblage is dominated by Scolicia prisca and S. isp. var. laminites, with subordinate Asterosoma isp., Bichordites monastiriensis, Ophiomorpha isp., Palaeophycus isp., Rhizocorallium isp., and Thalassinoides isp. (Figure 2). Dense indistinct bioturbation was observed on some beds. Rhizobioturbation on the top of sandy beds bearing Bichordites attests to shallower marine conditions (Villegas-Martín et al., 2014; Villegas-Martín and Netto, 2017).
MATERIALS AND METHODS
The specimens analyzed in this study mainly occur in the sandstone beds of the Capdevila Formation from western Cuba (Villegas-Martín, 2009; Rojas-Consuegra and Villegas-Martín, 2009; Villegas-Martín et al., 2014) (Figure 1). These sandstone beds crop out on La Habana-Pinar del Río national highway, around 4.5 km N to the Guanajay village (22º57'57.83" N, 82º41'23.13" W), in the Artemisa Province (La Habana-Matanzas Anticlinal), and at the northeast from the Pinar del Río city (Pinar del Río Province), close to the Pinar fault (22º23'16.8" N, 83º47'10.4" W), which constituted the tectonic boundary between the Guaniguanico mountains and the Los Palacios Basin.
Part of the analyzed material comprises specimens observed exclusively in the field (mostly from the Pinar del Río section) and previously studied by Villegas-Martín et al. (2014). Most of the studied specimens from the Artemisa section, previously studied by Villegas-Martín (2009) and Rojas-Consuegra and Villegas-Martín (2009), are housed in the paleontological collection of the National Museum of Natural History (MNHNCu) in La Habana, Cuba (samples MNHNCu-96.001287, MNHNCu-96.001291, MNHNCu-96.001292, MNHNCu-96.001294, MNHNCu-96.001295, MNHNCu-96.001296, MNHNCu-96.001297, MNHNCu-96.002933, MNHNCu-96.003567, MNHNCu-96003689). Those assigned to Scolicia without an ichnospecific definition by Villegas-Martín (2009) were revised in this work.
 The analyzed trace fossils were characterized through direct observations of the specimens in the collection and in situ at the outcrops in the study area. The distinct morphologies were grouped based on their morphology and toponomy, using the classification proposed by Seilacher (1964) and Savrda (2007). The ichnotaxonomical approach followed the revisions made by Smith and Crimes (1983), Plaziat and Mahmoudi (1988), and Uchman (1995). Furthermore, the terminology used to describe the Scolicia specimens is based on the descriptions by Plaziat and Mahmoudi (1988) and Uchman (1995) (see Figure 3).
The analyzed trace fossils were characterized through direct observations of the specimens in the collection and in situ at the outcrops in the study area. The distinct morphologies were grouped based on their morphology and toponomy, using the classification proposed by Seilacher (1964) and Savrda (2007). The ichnotaxonomical approach followed the revisions made by Smith and Crimes (1983), Plaziat and Mahmoudi (1988), and Uchman (1995). Furthermore, the terminology used to describe the Scolicia specimens is based on the descriptions by Plaziat and Mahmoudi (1988) and Uchman (1995) (see Figure 3).
THE ICHNOGENUS SCOLICIA AT THE CAPDEVILA FORMATION
Except for Bichordites, recorded by Villegas-Martín et al. (2014) in Pinar del Río section, all the Scolicia group morphologies found in the Capdevila Formation deposits can be assigned to the ichnogenus Scolicia. These show bilobate and trilobate morphology, a meniscate infill (backfill structure), two parallel sediment strings at each side of the burrow floor, and a flat or concave up ridge between the strings (e.g., Smith and Crimes, 1983; Plaziat and Mahmoudi, 1988; Uchman, 1995; Belaústegui et al., 2017). Three different morphologies prevail among these specimens and can be assigned to: Scolicia prisca, Scolicia vertebralis, and Scolicia isp. var laminites. In the case of the Scolicia vertebralis, we demonstrated that this ichnospecies is a preservational variant de Scolicia, and thus named as Scolicia isp. var vertebralis in this work.
Scolicia prisca de Quatrefagues, 1849
The specimens ascribed to Scolicia prisca are the most abundant among the studied material (MNHNCu-96.001292, MNHNCu-96.001294, MNHNCu-96.001297, MNHNCu-96.002933, MNHNCu-96.003567), and occur in fine- to medium-grained sandstone beds in the Artemisa and Pinar de Río (Figure 2) sections and carbonate beds in the Artemisa section.
 The structures are characterized by an epichnial trilobate furrow in a longitudinal section formed by two convex lateral slopes and a bottom. Slightly inclined convex asymmetric ribs compose the backfill structure that covers the slopes, more noticeable in some portions. The upper margins of the lateral slopes are elevated above the bedding surface, in general. The bottom is characterized by a convex central ridge-like structure usually covered by asymmetric fine transverse ribs perpendicular to the long axis and showing two sediment parallel strings/rows at each side (Figure 4). The convex ridge varies in width and height (e.g., Smith and Crimes, 1983; Plaziat and Mahmoudi, 1988; Uchman, 1995). The furrows show a straight to slightly meandering trajectory, with a predominance of the latter. The furrow width varies along the structure and the length ranges from 157.9 mm to 198.7 mm among the analyzed specimens. The width varies from 29.2 mm to 31.6 mm. The ribs are variable in size, with a length between 9.5 mm and 16.4 mm and a width between 1.8 mm and 3.5 mm. The bottom is 2.0-3.8 mm wide.
The structures are characterized by an epichnial trilobate furrow in a longitudinal section formed by two convex lateral slopes and a bottom. Slightly inclined convex asymmetric ribs compose the backfill structure that covers the slopes, more noticeable in some portions. The upper margins of the lateral slopes are elevated above the bedding surface, in general. The bottom is characterized by a convex central ridge-like structure usually covered by asymmetric fine transverse ribs perpendicular to the long axis and showing two sediment parallel strings/rows at each side (Figure 4). The convex ridge varies in width and height (e.g., Smith and Crimes, 1983; Plaziat and Mahmoudi, 1988; Uchman, 1995). The furrows show a straight to slightly meandering trajectory, with a predominance of the latter. The furrow width varies along the structure and the length ranges from 157.9 mm to 198.7 mm among the analyzed specimens. The width varies from 29.2 mm to 31.6 mm. The ribs are variable in size, with a length between 9.5 mm and 16.4 mm and a width between 1.8 mm and 3.5 mm. The bottom is 2.0-3.8 mm wide.
In some specimens, part of the furrow bottom is characterized by a single groove (Figure 4D). In others, only part of the furrow is formed by the convex central ridge and the two sediment parallel strings; the slopes are not identified (Figure 4E). Smith and Crimes (1983) assigned the latter morphology to Scolicia isp. Some of these specimens were assigned to S. prisca previously by Rojas-Consuegra and Villegas-Martín (2009) and Villegas-Martín et al. (2014). In both cases, considering their partial preservation, they were herein re-assigned to S. cf. prisca (sensu Uchman, 1995).
Scolicia isp. var. vertebralis Villegas-Martín et al., 2024
 The specimens assigned to Scolicia isp. var. vertebralis are bilobate furrows preserved as concave epirelief and occur exclusively in the flysch deposits of the Artemisa section (MNHNCu-96.001296). The furrows occur in fine- to medium-grained sandstone beds, are horizontal to the bedding plane, and show straight to slightly meandering trajectories (Figure 5C-D). They are straight to curved wide V-shaped furrows with elevated slopes in cross-section (Figure 5C). The slopes are oblique with asymmetric arcuate ribs (backfill infill) that extend to the margin (Figure 5A-B). The furrow displays a bottom composed of a single central groove of sediment (e.g., Uchman, 1998). The furrows are 29.2-31.6 mm wide. The ribs are 9.5-16.4 mm long and 1.8-3.5 mm wide. The single central groove is 2.0-3.8 mm wide.
The specimens assigned to Scolicia isp. var. vertebralis are bilobate furrows preserved as concave epirelief and occur exclusively in the flysch deposits of the Artemisa section (MNHNCu-96.001296). The furrows occur in fine- to medium-grained sandstone beds, are horizontal to the bedding plane, and show straight to slightly meandering trajectories (Figure 5C-D). They are straight to curved wide V-shaped furrows with elevated slopes in cross-section (Figure 5C). The slopes are oblique with asymmetric arcuate ribs (backfill infill) that extend to the margin (Figure 5A-B). The furrow displays a bottom composed of a single central groove of sediment (e.g., Uchman, 1998). The furrows are 29.2-31.6 mm wide. The ribs are 9.5-16.4 mm long and 1.8-3.5 mm wide. The single central groove is 2.0-3.8 mm wide.
This morphology has been assigned to the S. vertebralis ichnospecies, which included, in general, furrows with slopes covered by asymmetric ribs and a bottom composed of a single central sediment row. As suggested for the Scolicia vertebralis ichnospecies, the single central row of sediment constituted the original space occupied by the dorsal ridge and parallel strings (Figure 4D, Figure 5). Therefore, the morphology that characterizes S. vertebralis can also be observed in parts of S. prisca. For instance, one specimen shows S. prisca grading to S. vertebralis due to the erasure of the dorsal ridge (Figure 4D). Therefore, we identified that the S. vertebralis morphology can be produced as result of erosion of the convex central ridge and the two parallel strings in S. prisca. Thus, we considered S. vertebralis as a preservational variant, named as Scolicia isp. var vertebralis in this work.
Initially, these specimens here assigned to Scolicia isp. var vertebralis were previously ascribed to ?Psammichnites isp. by Villegas-Martín (2009) and Scolicia cf. prisca by Rojas-Consuegra and Villegas-Martín (2009).
Scolicia isp. var. laminites Uchman, 1995
 Scolicia isp. var. laminites was described in samples collected at the Artemisa section (MNHNCu-96.001295, MNHNCu-96003689), and in situ in deposits at the Pinar del Río section (Figure 2). These burrows are preserved as convex epireliefs at the top of medium-grained sandstone and carbonate beds (Figure 6), are parallel to the bedding plane, and show straight to slightly meandering trajectories. The specimens are burrows with discrete biserial menisci (backfill structure) (Figure 6A-D). A median groove can be observed in cross-section in some burrows (Figure 6C-D). The burrows are 25.0-30.6 mm wide.
Scolicia isp. var. laminites was described in samples collected at the Artemisa section (MNHNCu-96.001295, MNHNCu-96003689), and in situ in deposits at the Pinar del Río section (Figure 2). These burrows are preserved as convex epireliefs at the top of medium-grained sandstone and carbonate beds (Figure 6), are parallel to the bedding plane, and show straight to slightly meandering trajectories. The specimens are burrows with discrete biserial menisci (backfill structure) (Figure 6A-D). A median groove can be observed in cross-section in some burrows (Figure 6C-D). The burrows are 25.0-30.6 mm wide.
According to Uchman (1995), this morphology represents the preservation of the upper part of echinoid burrows with similar morphology to Bichordites and Scolicia. Some studied specimens were assigned to Scolicia isp. var. laminites based on the identification of the convex central ridge and the two sediment rows/strings at the bottom (Figure 6C-F). The names Scolicia var. laminites or Scolicia isp. have been used to characterize burrows showing similar morphology (the former “Laminites”) without referring to the presence of the parallel strings (Poiré et al., 2003; Rebata et al., 2006). The attribution of trace fossils to a non-specific ichnospecies is a common practice in ichnology, especially when clear diagnostic features are lacking. However, the identification of the parallel strings and/or convex central ridge should be crucial for attributing the trace fossil to Scolicia.
THE EOCENE SCOLICIA TRACEMAKER
Natural observations and experiments in aquaria demonstrated that the ichnogenus Scolicia (as the morphologies described herein) corresponds to the burrows resulted from the grazing activity of spantagoid echinoids with double drainage tubes (e.g., Ward and Lewis, 1975; Howard et al., 1974; Bromley and Asgaard, 1975; Smith and Crimes, 1983; Bromley et al., 1997; Belaústegui et al., 2017). These large meniscate traces are typically produced by backfilling when the spatangoid plows through the sediment (Smith and Crimes, 1983; Kanazawa, 1992, 1995; Bromley et al., 1997). The animal excavates the sediment in the front, which is transported to the posterior part of the burrow, where it is packed with mucus. The open burrow is limited to the volume immediately surrounding the echinoid test. The two open drains (related to sediment strings) are kept behind by action of the subanal tuft spines and the corresponding tube feet, while periodically, a respiratory shaft may be constructed for aeration purposes or to feed upon surface detritus (Smith and Crimes, 1983; Plaziat and Mahmoudi, 1988; Kanazawa, 1992, 1995; Bromley et al., 1997).
It is well-known that many echinoids from different fossil and extant families of the Order Spantangoida possess the morphology to produce Scolicia -like burrows (e.g., Bromley and Asgaard, 1975, Bromley et al., 1995, 1997; Kanazawa, 1995; Asgaard and Bromley, 2007). Fossil species of different families, such as Schizasteridae (Brisaster iheringi) and Hemiasteridae (Hemiaster expergitus), were observed within or associated with Scolicia traces (Fu and Werner, 2000; Carmona et al., 2020).
The fossil record of spatangoid in Cuba is extensive and includes specimens of different families, including the Schizasteridae and Hemiasteridae families. In addition, genera such as Hemiaster, Schizaster, and Linthia have been identified in Eocene deposits (e.g., Sanchez Roig, 1926, 1949; Kier, 1984). Thus, these taxa might be potential tracemakers of the Scolicia specimens studied herein. However, no body fossil of spatangoid echinoids were reported in the studied sections bearing Scolicia, although undetermined fragments of echinoids are known in the marine deposits of the Capdevila Formation (Brust et al., 2011). Therefore, further detailed studies on the echinoderms found in the Capdevila Formation are necessary to try to infer the Scolicia tracemakers more accurately.
TAPHONOMY INSIGHTS
The complex morphological features that characterize the ichnogenus Scolicia represent distinct preservational variations used to define the different ichnospecies (e.g., Uchman, 1995, 1998). Thus, the variability in the Scolicia preservation seems to be mostly taphonomic and results from factors such as the burrow position within the sediment bed, superimposition, erosion, and lithological diversity (e.g., Plaziat and Mahmoudi, 1988; Uchman, 1995; Gibert and Goldring, 2008). Morphological variations are observed in the Scolicia specimens from the Capdevila Formation. These variations include features concerning the ribbed slopes, the preservation of the parallel strings, and the convex central ridge.
 The ribs of the slopes are less evident in the specimens of Scolicia prisca and Scolicia isp. var vertebralis preserved in medium-grained sandstone beds in the Capdevila Formation (Figure 4, Figure 5). In S. prisca, the ribbed slopes are partially preserved or absent in some parts of the furrows (Figure 4D, E). These furrows show only the convex central ridge and the two parallel strings/rows. These preservational variations were previously reported by Plaziat and Mahmoudi (1988, fig. 10) in the deposits of the Cuisian Basin, Arro (Spain). At the Capdevilla Formation, this typical morphology is a consequence of a deep erosion that erased most parts (slopes) of the burrow, preserving only its bottom (Figure 4E). In beds with abundant Scolicia, slopes are often partially or totally destroyed by burrows’ overlap (Figure 7), a feature also observed previously (Fu and Werner, 2000).
The ribs of the slopes are less evident in the specimens of Scolicia prisca and Scolicia isp. var vertebralis preserved in medium-grained sandstone beds in the Capdevila Formation (Figure 4, Figure 5). In S. prisca, the ribbed slopes are partially preserved or absent in some parts of the furrows (Figure 4D, E). These furrows show only the convex central ridge and the two parallel strings/rows. These preservational variations were previously reported by Plaziat and Mahmoudi (1988, fig. 10) in the deposits of the Cuisian Basin, Arro (Spain). At the Capdevilla Formation, this typical morphology is a consequence of a deep erosion that erased most parts (slopes) of the burrow, preserving only its bottom (Figure 4E). In beds with abundant Scolicia, slopes are often partially or totally destroyed by burrows’ overlap (Figure 7), a feature also observed previously (Fu and Werner, 2000).
The typical parallel strings of Scolicia prisca are only preserved in some specimens in the Artemisa section, being the two sediment parallel rows predominant in the specimens studied (Figure 4A-F). These sediment rows are well reported for S. prisca (e.g., Bromley and Asgaard, 1975; Ksiażkiewicz, 1977; Smith and Crimes, 1983; Plaziat and Mahmoudi, 1988, Uchman, 1995, 1998) and constitute the proof that the parallel strings had existed. The preservation of the parallel strings is either rare or occasional (Smith and Crimes, 1983; Plaziat and Mahmoudi, 1988) because they are empty tunnels that eventually can collapse due to the sediment pressure (Kanazawa, 1995), or due to animal displacement (Smith and Crimes, 1983). Backfill lamination may be preserved in the external surface of the parallel strings (Smith and Crimes, 1983), and this feature has been seen in a few specimens from the Capdevilla Formation in the Artemisa section (Figure 4B). In some S. prisca, the sediment parallel rows can be tapered due to superimposition of the lateral slopes over the margins of the convex central ridge (Figure 4F). This characteristic has already been mentioned for S. prisca by Donovan et al. (2005), and had been discussed by Plaziat and Mahmoudi (1988). According to Uchman (1995, 1998), a similar situation can originate when the burrow occupies a deep position in the sandy portion of the beds, and the lateral slopes rest over the convex central ridge. In most S. prisca specimens examined, the bottom ridge is covered by laminated surface (Figure 4 A-C, E-F). In a few specimens, however, the convex central ridge presents a smooth surface (Figure 4D). In general, and according to the studied material, the width of the central ridge and the height of the lateral slopes can vary within the same specimen. However, it can also remain constant in some specimens. Uchman (1995) also observed similar features in Scolicia specimens from Miocene deposits of Italy. In other specimens, the typical morphology of S. prisca loses the slope, preserving the two parallel strings and the central ridge, only (Figure 4E). This change in morphology is caused by erosion (as explained above), reflecting the change in the position of the tracemaker from a shallower level to a deeper one into the substrate.
 Scolicia prisca and Scolicia isp. var vertebralis mainly correspond to burrows made in the sand/mud interface. It is well-accepted that these morphologies are a consequence of the recent or sub-recent rock weathering, when the upper part of the full burrow is eroded, preserving only the lower portion excavated in the sand (e.g., Plaziat and Mahmoudi, 1988; Uchman, 1995). However, extant spatangoid echinoids can produce structures similar in morphology to S. prisca and Scolicia isp. var vertebralis at the substrate surface in extant shallow marine settings (Figure 8). Although the preservation potential of these furrows is low, due to the continuous action of erosive processes in these settings, they demonstrated that both morphologies can be made on top of the substrate.
Scolicia prisca and Scolicia isp. var vertebralis mainly correspond to burrows made in the sand/mud interface. It is well-accepted that these morphologies are a consequence of the recent or sub-recent rock weathering, when the upper part of the full burrow is eroded, preserving only the lower portion excavated in the sand (e.g., Plaziat and Mahmoudi, 1988; Uchman, 1995). However, extant spatangoid echinoids can produce structures similar in morphology to S. prisca and Scolicia isp. var vertebralis at the substrate surface in extant shallow marine settings (Figure 8). Although the preservation potential of these furrows is low, due to the continuous action of erosive processes in these settings, they demonstrated that both morphologies can be made on top of the substrate.
Furrows similar to S. prisca were also observed in fine-grained sediments from modern deep-sea depositional settings (3.451 m) in the southeastern Indian Ocean, where irregular echinoids plow a furrow as wide as its body in fine sediment when searching for food (Hollister et al., 1975, figure 21.18). In both cases (deep-sea and shallow marine settings), these furrows can be preserved mainly at the top bed and do not always represent eroded burrows. Furthermore, these extant morphologies resembling to S. prisca present the sediment rows at the bottom of the furrow and the sediment parallel strings are not observed. In this sense, Smith and Crimes (1983) suggested that although some Scolicia appear to be formed by shallow-burrowing echinoids at an interface of mud overlying sand, others are probably formed by epifaunal echinoids plowing half-buried through the sediment-water interface, leaving a partially backfilled furrow. In the case of Scolicia isp. var. laminites specimens, the preservation of a bilobate top, as observed in some of the studied specimens, suggests the erasing of the uppermost part of the burrow. The laminated backfill is better visualized in burrows preserved in carbonate rocks, probably due to the early cementation process during the diagenesis. In contrast, it is subtle in burrows preserved in medium-grained sandstones (Figure 6A-D). The vertical cross-cutting of these specimens shows the bilobate top, the laminated backfill, and the two parallel strings at the burrow bottom (Figure 6E-F).
PALEOECOLOGICAL INSIGHTS
Artemisa Section
The Artemisa section deposits, which contain Scolicia prisca, Scolicia isp. var. vertebralis, and Scolicia isp. var. laminites were accumulated in a basin floor setting impacted by turbidity currents, probably in a bathyal context (Albear and Iturralde-Vinent, 1985; Lex, 2013). Scolicia is commonly registered in deep-sea turbiditic beds since the Mesozoic (e.g., Uchman, 1995, 1998; Tchoumatchenco and Uchman, 2001; Wetzel and Uchman, 2001; Rodríguez-Tovar et al., 2010; Buatois et al., 2024). Graphoglyptid trace fossils, such as Cosmorhaphe isp., Helminthoraphe isp., and Paleodyction isp., also occur associated with Scolicia in the Artemisa section (Rojas-Consuegra and Villegas-Martín, 2009; Rojas-Consuegra et al., 2018). Although the ichnotaxonomy of some of these trace fossils needs revision (A. Uchman, pers. comm.), the ichnological assemblage includes winding, meandering, and network structures typical of the Nereites ichnofacies commonly found in post-Paleozoic deep-sea turbiditic setting (e.g., Rodríguez-Tovar et al., 2010; Buatois and Mángano, 2011; Uchman and Wetzel, 2011). Rojas-Consuegra and Villegas-Martín (2009) suggested the occurrence of the Nereites ichnofacies, in the Artemisa section based on the presence of these graphoglyptid burrows.
The abundance of Scolicia morphologies at the top of the sandstone beds in the Artemisa section suggests post-event colonization, a common strategy in deep-sea beds (e.g., Uchman, 1995; Buatois et al., 2001; Rodríguez-Tovar et al., 2010). There is a consensus that when the gravity flows ceased, these burrows are produced at the sand/mud interface in shallow tiers (e.g., Bromley and Asgaard, 1975; Werner and Wetzel, 1982). Posterior erosion will only preserve the basal portion of the spatangoid burrow, which are recognized as S. prisca and S. vertebralis (S. isp. var. vertebralis in this work) in the fossil record (e.g., Uchman, 1995). However, spatangoids also produce locomotion furrows similar to S. prisca on the seabed surface not only in shallow waters (Figure 8A) but also in deep-sea settings (Hollister et al., 1975, figures 21.10, 21.18). Contrary to shallow marine settings, the potential for preserving superficial furrows in deep-sea substrates is greatly enhanced by the muddy composition and the low hydrodynamic energy, which favors sediment aggregation by biofilms (the bioglue principle sensu Goldring and Seilacher, 1971) and substrate compaction. Hence, S. prisca and S. isp. var. vertebralis might represent original furrows produced at the sand/mud interface at the top of a fresh turbidite bed. Once the gravitational flow currents cease, the mud particles decant, forming a thin mud layer (turbidite Te layer) atop of the sand/silt beds (Tc/Td layers). Pelagic material decays in the mud, enriching the marine substrate with organic matter and microbial activity. These processes open the colonization window (sensu Pollard et al., 1993) for spatangoid forage, leaving furrows. The bottom quiescence supports biofilm formation and enhances EPS (extracellular polymeric substance) into the substrate, allowing the furrows to be preserved (the bioglue property of EPS, as explained by Goldring and Seilacher, 1971). Upon rock exposure and weathering, they appear as epichnial grooves on the top of turbidite sandstone beds. This hypothesis could explain the presence of some S. prisca and S. isp. var. vertebralis as epichnial furrows in the flysch deposits of the Artemisa section. However, additional taphonomical studies on Scolicia traces in deep-sea are necessary to test this hypothesis.
Pinar del Río Section
The Pinar del Río section (Palacios Basin) has been interpreted as deposited in upper slope settings based on the sedimentary facies association, the prevalence of a Cruziana ichnofacies trace fossil assemblage; and the lack of typical traces of the Nereites ichnofacies (see Villegas-Martín et al., 2014). An upper slope context for these deposits was also suggested by Brust et al. (2011) based on micropaleontological and sedimentological data. Indeed, the geomorphology of the Los Palacios Basin during the early Eocene allowed the development of a short platform and a shallower slope in the Pinar del Río area (Sommer, 2009; Villegas-Martín et al., 2014, figure 8).
Although Scolicia is not commonly registered in ancient slope deposits, a few records are known (Fu and Werner, 2000; Demircan and Uchman, 2017). The presence of spatangoid echinoids producing Scolicia on a shallower slope can be favored by better substrate oxygenation, high sedimentation rates, and the prevalence of sandy substrates (Villegas-Martín et al., 2014). Extant Scolicia tracemakers (Spatangus group, Smith and Crimes, 1983) live in benthic settings from shelf to deep slope, preferring sandy substrates (Kanazawa, 1992; Kroh and Hansson, 2013). Fu and Werner (2000) identified abundant Scolicia- like burrows in fine-grained sandy substrates from slope to deep-sea zones (400 to 3.800 m) in the NE Atlantic Ocean, inferring that sand granulometry, bottom currents, and higher sedimentation rates control their distribution more than the water depth. Furthermore, echinoids that produce Scolicia- type burrows are more tolerant to the increase in sedimentation rates (Vermeij, 1978). Thus, the scarcity of Scolicia record in slope deposits might be due to the predominance of muddy sediments and the low oxygenation and sedimentation rates typical of deeper slope settings.
The Scolicia -bearing trace fossil assemblage in the Pinar del Río section is interpreted as an impoverished expression of the Cruziana ichnofacies due to the prevalence of burrows produced by detritus-feeding marine invertebrates and its considerably low ichnodiversity (Villegas-Martin et al., 2014). Except for a few records (e.g., Savrda et al., 2001), Cruziana ichnofacies assemblages in slope deposits are uncommon. These deposits are mainly characterized by Zoophycos ichnofacies, particularly in post-Paleozoic rocks, having Zoophycos and Chondrites as ichnoguilds (e.g., Bromley, 1996; Buatois and Mángano, 2011). Besides Scolicia has been reported with Zoophycos and Chondrites in slope settings, when interbedded muddy and sandy substrates occur (Fu and Werner, 2000), the typical features of the Zoophycos ichnofacies, such as the dominance of complex feeding deep-tier structures with spreite made by deposit feeders or farmers (Buatois and Mángano, 2011), are not observed in the Pinar del Río section.
Thus, the predominance of sandy deposits and the high sedimentation rates in the slope settings represented by Pinar del Río deposits probably inhibited the activity of Zoophycos and Chondrites tracemakers, which prefer muddy substrates (e.g., Fu and Werner, 2000; Buatois and Mángano, 2011). Despite the high frequency of the erosive events that characterize the Pinar del Río section slope deposits (Villegas-Martín et al., 2014), erosion does not play a role in the absence of Zoophycos and Chondrites in the Scolicia -dominated assemblage. These burrows normally occur deeper into the substrate than Scolicia (e.g., Bromley, 1996; Buatois and Mángano, 2011); thus, if their absence was the result of erosion, the Scolicia specimens would also be absent. Furthermore, the absence of deeper tiers might have favored the abundance of Scolicia in the Pinar del Río section (e.g., Bromley and Asgaard, 1975; Werner and Wetzel, 1982; Gibert and Goldring, 2008).
CONCLUSIONS
The bilobate and trilobate trace fossils preserved as concave and convex epirelief in the sedimentary rocks of the Capdevila Formation (western Cuba) can be assigned to the ichnogenus Scolicia, representing Scolicia prisca, S. cf. prisca, S. isp. var. vertebralis, and S. isp. var. laminites. Scolicia isp. var. vertebralis is considered a preservational variant of Scolicia prisca in this work. The observed variations in the diagnostic features (dorsal ridge, laminate backfill, and parallel strings) reflect the wide variation expected in the burrows made by spatangoid echinoderms.
Some Scolicia morphologies might not represent exclusively the basal portion of eroded burrows but also furrows produced by spatangoids on the surface of sea bottoms, mainly in deep sea settings. The predominance of sandstone beds deposited by turbidity currents and the high sedimentation rates favored the colonization of spatangoid echinoderms that generated Scolicia in shallower slope settings and precluded the establishment of a typical Zoophycos ichnofacies assemblage in the slope deposits of the Pinar del Río section.
ACKNOWLEDGEMENTS
The authors thank the Museo Nacional de Historia Natural de La Habana for the access to samples studied in this work, Paulo Paim, Ubiratan Faccini, and Elvio Bosetti for the critical comments that helped to improve this manuscript, and The Brazilian Research and Technological Development Council (CNPq) for the PEC-PG grant 190620/2011-0 to JVM, and grants 451731/2013-1 and 303863/2016-1 to RGN. To the Instituto de Ecología y Sistemática (IES) de La Habana for the support to JVM. Roberto Pillon is thanked for providing the picture that illustrates Figure 8A. Our sincere gratitude goes to the handling editor Noelia Carmona and to Zain Belaústegui (University of Barcelona) and one anonymous reviewer for their insightful constructive comments, which have helped to improve the manuscript.
REFERENCES
Albear, J.F. and Iturralde-Vinent, M.A. 1985. Estratigrafía de las provincias de La Habana, p. 12–54. In Arroyo, A. (ed.), Contribución a la geología de las provincias de La Habana y Ciudad de La Habana. Editorial Cientifíco-Técnica, La Habana.
Asgaard, U. and Bromley, R.G. 2007. Co-occurrence of Schizasterid echinoids and the trace fossil Scolicia, Pleistocene, Greece: Facts, myths, and fascioles, p. 87–95. In Bromley, R.G., Buatois, L.A., Mángano, G., Genise, J.F., and Melchor, R.N. (eds.), Sediment-Organism Interactions: A Multifaceted Ichnology. SEMP Special publication, Oklahoma.
Belaústegui, Z., Muñiz, F., Nebelsick, J.H., Domènech, R., and Martinell, J. 2017. Echinoderm Ichnology: Bioturbation, bioerosion and related processes. Journal of Paleontology, 91:643–661.
https://doi.org/10.1017/jpa.2016.146
Bernardi, M., Boschele, S., Ferretti, P., and Avanzini, M. 2010. Echinoid burrow Bichordites monastiriensis from the Oligocene of NE Italy. Acta Palaeontologica Polonica, 55:479–486.
https://doi.org/10.4202/app.2009.0064
Bralower, T.J. and Iturralde-Vinent, M.A. 1997. Micropaleontological dating of the collision between the North American and Caribbean plates in western Cuba. Palaios, 12:133–150.
https://doi.org/10.2307/3515303
Bromley, R.G. 1996. Trace Fossils. Biology, Taphonomy and Applications, second ed. Chapman and Hall, London.
Bromley, R.G. and Asgaard, U. 1975. Sediment structures produced by a spatangoid echinoid: a problem of preservation. Bulletin of the Geological Society of Denmark, 24:261–281.
Bromley, R.G., Jensen, M., and Asgaard, U. 1995. Spatangoid echinoids: deep-tier trace fossils and chemosymbiosis. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, 195:25–35.
https://doi.org/10.1127/njgpa/195/1995/25
Bromley, R.G., Asgaard, U., and Jensen, M. 1997. Experimental study of sediment structures created by a spatangoid echinoid, Echinocardium mediterraneum. Proceedings of the Geologists' Association, 108:183–189.
https://doi.org/10.1016/S0016-7878(97)80026-0
Brönnimann, P. and Rigassi, D. 1963. Contribution to the geology and palaeontology of the area of the city of La Habana, Cuba and its surroundings. Eclogae Geologicae Helvetiae, 56(1):193–430.
Brönnimann, P. and Stradner, H. 1960. Die Foraminiferen und- discoasteridenzonen von Kuba und ihre interkontinentale korrelation. Erdoel-Zeitschrift, 76:364–369.
Brust, J., Hüneke, H., Meschede, M., and Sommer. M. 2011. Facies and provenance of basin-margin deposits in the Los Palacios Basin (Capdevila Formation, Cuba). Facies, 57:73–92.
https://doi.org/10.1007/s10347-010-0233-1
Buatois, L.A. and Mángano, M.G. 2011. Ichnology: Organism-substrate interactions in space and time. Cambridge University Press, Cambridge.
Buatois, L., Mángano, M.G., and Sylvester. Z. 2001. A diverse deep-marine ichnofauna from the Eocene Tarcau Sandstone of the Eastern Carpathians, Romania. Ichnos, 8(1):23–62.
https://doi.org/10.1080/10420940109380172
Buatois, L.A., Mángano, M.G., and Carmona, N.B. 2024. Scolicia, ichnotaxonomic practices, and the limits of behavioural convergence. Ichnos, 30(4):338–361.
https://doi.org/10.1080/10420940.2024.2325468
Carmona, N.B., Gabriela, M., Buatois, L., Bromley, R., Ponce, J.J., Asgaard, U., and Bellosi, E. 2020. Scolicia and its producer in shallow-marine deposits of the Miocene Chenque Formation (Patagonia, Argentina): functional morphology and implications for understanding burrowing behavior. Ichnos, 27:290–299.
https://doi.org/10.1080/10420940.2020.1744589
Chesher, R.H. 1963. The morphology and function of the frontal ambulacrum of Moira atropos (Echinoidea: Spatangoida). Bulletin of Marine Science of the Gulf and Caribbean, 13:549–573.
Cushman, J.A. and Bermúdez. P.J. 1949. Some Cuban species of Globorotalia: Cushman. Foraminifera. Research Contribution, 25(2):26–45.
de Quatrefagues, M.A. 1849. Note sur la Scolicia prisca (A. de Q) annelide fossile de la Craie. Annales des Sciences Naturelles, 3 sér., Zoologie, 12:265–266.
Demircan, H. and Uchman, A. 2017. Short distance variability of trace fossils in submarine slope and proximal basin plain deposits: a case study from the Ceylan Formation (upper Eocene), Gelibolu Peninsula, NW Turkey. Bollettino della Società Paleontologica Italiana, 56:253–275.
https://doi.org/10.4435/BSPI.2017.22
Donovan, S.K., Willem, R., and Pickerill, R.K. 2005. The ichnofossil Scolicia prisca de Quatrefages from the Paleogene of Eastern Jamaica and fossil echinoids of the Richmond Formation. Caribbean Journal of Science, 41(4):876–881.
Fu, S. and Werner, F. 2000. Distribution, ecology and taphonomy of the organism trace, Scolicia, in northeast Atlantic deep-sea sediments. Palaeogeography, Palaeoclimatology, Palaeoecology, 156:289–300.
https://doi.org/10.1016/S0031-0182(99)00146-7
García-Delgado, D.E. and Torres Silva, A. 1997. Sistema Paleógeno, p. 115–140. In Furrazola-Bermúdez, G.F., and Núñez-Cambra, K. (eds.), Estudios sobre geología de Cuba. Centro Nacional de información geológica, La Havana.
Giannetti, A. 2010. Influence of climate, sea-level changes and tectonics on ichnoassemblages distribution in a carbonate-dominated, deep-marine environment (Upper Paleocene, Zumaya section). Palaeogeography, Palaeoclimatology, Palaeoecology, 285:104–118.
https://doi.org/10.1016/j.palaeo.2009.11.001
Gibert, J.M.de. and Goldring, R. 2008. Spatangoid-produced ichnofabrics (Bateig Limestone, Miocene, Spain) and the preservation of spatangoid trace fossils. Palaeogeography, Palaeoclimatology, Palaeoecology, 270:299–310.
https://doi.org/10.1016/j.palaeo.2008.01.031
Goldring, R. and Seilacher, A. 1971. Limulid undertracks and their sedimentologic implications. Neues Jahrbuch für Geologie und Paläontologie, 137:422–442.
Gordon, M.B., Mann, P., Cáceres, D., and Flores, R. 1997. Cenozoic tectonic history of the North American-Caribbean plate boundary zone in western Cuba. Journal Geophysical Research, 102:10055–10082.
https://doi.org/10.1029/96JB03177
Götzinger, G. and Becker, H. 1932. Zur geologischen Gliederung des Wienerwald flysches (Neue Fossilfunde). Jahrbuch der Geologische Bundesanstalt, 82:343–396.
Häntzschel, W. 1975. Trace fossils and Problematica, p. T1–T269. In Teichert, C. (ed.), Treatise on Invertebrate Paleontology. Part W, Miscellanea, Supplement 1. Geological Society of America, Boulder, Colorado and University of Kansas Press, Kansas.
Hollister, C.D., Heezen, B.C., and Nafe, K.E. 1975. Animal traces on the deep-sea floor, p. 493–510. In Frey, R.W. (ed.), The study of trace fossils. Springer-Verlag New York Inc.
Howard, J.D., Reineck, H.E., and Rietschel, S. 1974. Biogenic sedimentary structures formed by heart urchins. Senckenbergiana Maritima, 6:185–205.
Kanazawa, K. 1992. Adaptation of test shape for burrowing and locomotion in spatangoid echinoids. Palaeontology, 35:733–750.
Kanazawa, K. 1995. How spatangoids produce their traces relationship between burrowing mechanism and trace structure. Lethaia, 28(21):1–219.
https://doi.org/10.1111/j.1502-3931.1995.tb01424.x
Kappus, E.J. and Lucas, S.G. 2019. Ichnologic note a new ichnospecies of Cardioichnus from the Cretaceous (Albian) of New Mexico. Ichnos, 26:127–133.
https://doi.org/10.1080/10420940.2018.1482215
Kier, P.M. 1984. Fossil spatangoid echinoids of Cuba. Smithsonian contributions to paleobiology, 55:1–336.
Kroh, A. and Hansson, H. 2013. Spatangus purpureus O.F. Müller, 1776. In Kroh, A. and Mooi, R. (eds.), World Echinoidea Database. Accessed through: World Register of Marine Species.
https://www.marinespecies.org/echinoidea/aphia.php?pp=takdetails&id=124418
Książkiewicz, M. 1970. Observations on the ichnofauna of the Polish Carpathians, p. 283–322. In Crimes, T.P. and Harpers, J.C. (eds.), Trace fossils. Seel House Press, Liverpool.
Ksiażkiewicz, M. 1977. Trace fossils in the flysch of the Polish Carpathians. Palaeontologica Polonica, 36:1–208.
Léxico Estratigráfico de Cuba (Lex). 2013. Instituto de Geología y Paleontología. Servicio Geológico de Cuba/Ministerio de Energía y Minas. Tercera Versión, La Habana.
Mayoral, E. and Muñiz, F. 2001. New ichnospecies of Cardioichnus from the Miocene of the Guadalquivir Basin, Huelva, Spain. Ichnos, 8:69–76.
https://doi.org/10.1080/10420940109380174
Naimi, M.N., Cherif, A., and Belaid, M. 2021. The trace fossil Cardioichnus planus from the lower Miocene of Algeria: the first record from Africa and a probable endemic tracemaker. Geologia Croatica, 74:121–126.
Nichols, D. 1959. Changes in the chalk heart-urchin Micraster interpreted in relation to living forms. Philosphical Transactions of the Royal Society B, 242:347–437.
Piotrowski, J. 1987. Nuevos datos sobre los sedimentos Cretácico Superior tardío y el Paleógeno en la zona estructo-facial de San Diego los Baños, p. 185–196. In Arroyo, A. (ed.), Contribución a la Geología de la provincia de Pinar del Río. Edición Científico-Técnicas, La Habana.
Plaziat, J.C. and Mahmoudi, M.1988. Trace fossils attributed to burrowing echinoids: a revision including new ichnogenus and ichnospecies. Geobios, 21:209–233.
https://doi.org/10.1016/S0016-6995(88)80019-6
Poiré, D.G., Spalletti, L.A., and Del Valle, A. 2003. The Cambrian-Ordovician siliciclastic platform of the Balcarce Formation (Tandilia System, Argentina): facies, trace fossils, palaeoenvironments and sequence stratigraphy. Geologica Acta, 1:41–60.
https://doi.org/10.1344/105.000001591
Pollard, J.E., Goldring, R., and Buck, S.G. 1993. Ichnofabrics containing Ophiomorpha: significance in shallow-water facies interpretation. Journal of the Geological Society, 150:149–164.
https://doi.org/10.1144/gsjgs.150.1.0149
Rebata, L.A., Gingras, M.K., Matti, E., Räsänem, M.E., and Barberi, M. 2006. Tidal-channel deposits on a delta plane from the Upper Miocene Nauta Formation, Marañon Foreland Sub-basin, Peru. Sedimentology, 53:971–1013.
https://doi.org/10.1111/j.1365-3091.2006.00795.x
Riahi, S., Uchman, A., Stow, D., Soussi, M., and Lattrache, K.B.I. 2014. Deep-sea trace fossils of the Oligocene-Miocene Numidian Formation, northern Tunisia. Palaeogeography, Palaeoclimatology, Palaeoecology, 414:155–177.
https://doi.org/10.1016/j.palaeo.2014.08.010
Rodríguez-Tovar, F.J., Uchman, A., Aitor Payros, A., Orue-Etxebarria, X., Apellaniz, E., and Molina, E. 2010. Sea-level dynamics and palaeoecological factors affecting trace fossil distribution in Eocene turbiditic deposits (Gorrondatxe section, N Spain). Palaeogeography, Palaeoclimatology, Palaeoecology, 285:50–65.
https://doi.org/10.1016/j.palaeo.2009.10.022
Rojas-Consuegra, R.R. and Villegas-Martín, J. 2009. Icnofósiles e Icnofacies en algunas formaciones geológicas cubanas. VIII Congreso de Geología, GEOCIENCIAS´2009. Memorias en CD-ROM, GEO2-P14, La Habana, p. 397–424. ISBN 978-959-7117-19-3.
Rojas-Consuegra, R.R., Villegas-Martín, J., and García Sánchez, R. 2018. Presencia de los icnogéneros Zoophycos Massalongo, 1855 y Paleodictyon Meneghini, 1850 en el Paleógeno de Cuba. IX Jornadas Científico Técnicas Ceinpet 2018, La Havana, p. 29.
Sánchez Roig, M. 1926. Contribución a la Paleontología Cubana: Los Equinodermos Fósiles de Cuba. Boletin de Minas, 10:1–179.
Sánchez Roig, M. 1949. Los equinodermos fósiles de Cuba. Paleontología de Cuba, 1:1–302.
Savrda, S. 2007. Taphonomy of trace fossils, p. 92–109. In Miller, W. III (ed.), Trace Fossils: Concepts, Problems, Prospects, Elsevier Science, Oxford.
Savrda, C.E., Krawinkel, H., McCarthy, F.M.G., McHugh, C.M.G., Olson, H.C., and Mountain, G. 2001. Ichnofabrics of a Pleistocene slope succession, New Jersey margin: relations to climate and sea-level dynamics. Palaeogeography, Palaeoclimatology, Palaeoecology, 171:41–61.
https://doi.org/10.1016/S0031-0182(01)00266-8
Seilacher, A. 1964. Sedimentological classifications and nomenclature of trace fossils. Sedimentology, 3:253–256.
https://doi.org/10.1111/j.1365-3091.1964.tb00464.x
Seilacher, A. 2008. Biomats, biofilms, and bioglue as preservational agents for arthropod trackways. Palaeogeography, Palaeoclimatology, Palaeoecology, 270:252–257.
https://doi.org/10.1016/j.palaeo.2008.07.011
Smith, A.B. and Crimes, T.P. 1983. Trace fossils formed by heart urchins - a estudy of Scolicia and related traces. Lethaia, 16:79–92.
https://doi.org/10.1111/j.1502-3931.1983.tb02001.x
Sommer, M. 2009. Late Cretaceous to Miocene tectonic reconstruction of the northwestern Caribbean-Regional analysis of Cuban geology. Unpublished Ph.D. dissertation, University of Greifswald, Greifswald, Germany.
Tchoumatchenco, P. and Uchman, A. 2001. The oldest deep-sea Ophiomorpha and Scolicia and associated trace fossils from the Upper Jurassic-Lower Cretaceous deep-water turbidite deposits of SW Bulgaria. Palaeogeography, Palaeoclimatology, Palaeoecology, 169:85–99.
https://doi.org/10.1016/S0031-0182(01)00218-8
Thompson, B.A.W. and Riddle. M.J. 2005. Bioturbation of the spatangoid urchin Abatus ingens in Antarctic marine sediments. Marine Ecology Progress Series, 290: 135–143.
https://www.jstor.org/stable/24868023
Torell, O.M. 1870. Petrificata Suecana Formationis Cambricae. Acta Universitets Lundensis, Lunds Universit Årsskrift, 2:1–14.
Tunis, G. and Uchman, A. 1993. Trace fossils in the "Flysch del Grivo" (Ypresian) in the Julian Pre-Alps, NE Italy: preliminary observations. Gortania, 14:71–104.
Tunis, G. and Uchman, A. 1998. Ichnology of Eocene flysch deposits in the Carnian Pre-Alps (North-Eastern Italy). Gortania, 20:41–58.
Uchman, A. 1991. Trace fossils from stress environments in Cretaceous-Paleogene of Polish Outer Carpathians. Annales Societatis Geologorum Poloniae, 61:207–220.
Uchman, A. 1995. Taxonomy and palaeoecology of flysch trace fossils: the Marnoso-arenacea Formation and associated facies (Miocene, Northern Apennines, Italy). Beringeria, 15:1–115.
Uchman, A. 1998. Taxonomy and ethology of flysch trace fossils: a revision of the Marian Książkiewicz collection and studies of complementary material. Annales Societatis Geologorum Poloniae, 68:105–218.
Uchman, A. and Wetzel, A. 2011. Deep-Sea ichnology: The relationships between depositional environment and endobenthic organisms, p. 517–556. In Hüneke, H. and Mulder, T. (eds), Developments in Sedimentology, Elsevier Science, Amsterdam.
Uchman, A., Lebanidze, Z., Kobakhidze, N., Beridze, T., Makadze, D., Koba, L., Khutsishvili, S., Chagelishvili, R., Koiava, K., and Khundadze, N., 2022. Unusual echinoid resting trace records change in the position of the redox boundary (Palaeogene of the Lesser Caucasus in Georgia). Acta Geologica Polonica, 72:317–330.
Vermeij, G.J. 1978. Biogeography and adaptation: Patterns of marine life. Harvard University Press, Cambridge, Massachusetts.
Villegas-Martín, J. 2009. Los icnofósiles de Cuba: icnotaxonomía, tafonomía y paleoambiente. Unpublished Graduate Dissertation, Universidad de la Habana, La Habana, Cuba.
Villegas-Martín, J. and Netto, R.G. 2017. Bichordites from the early Eocene of Cuba: significance in the evolutionary history of the spatangoids. Journal of South American Earth Sciences, 80:404–410.
https://doi.org/10.1016/j.jsames.2017.10.008
Villegas-Martín, J., Netto, R.G., Lavina, E.L.C., and Rojas-Consuegra, R. 2014. Ichnofabrics of the Capdevila Formation (early Eocene) in the Los Palacios Basin (western Cuba): Paleoenvironmental and paleoecological implications. Journal of South American Earth Sciences, 56:214–227.
https://doi.org/10.1016/j.jsames.2014.09.006
Ward, D.M. and Lewis, D.W. 1975. Paleoenvironmental implications of storm-scoured ichnofossiliferous mid-Tertiary limestones, Waihao district, South Canterbury, New Zealand. New Zealand Journal of Geology and Geophysics, 18:881–908.
https://doi.org/10.1080/00288306.1975.10423533
Werner, F. and Wetzel, A. 1982. Interpretation of biogenic structures in oceanic sediments. Bulletin de l' Institut de Géologie du Bassin d'Aquitaine, 31:275–288.
Wetzel, A. 2008. Recent bioturbation in the deep south China Sea: a uniformitarian ichnologic approach. Palaios, 23:601–615.
https://www.jstor.org/stable/27670546
Wetzel, A. and Uchman, A. 2001. Sequential colonization of muddy turbidites in the Eocene Beloveža Formation, Carpathians, Poland. Palaeogeography, Palaeoclimatology, Palaeoecology, 168:171–186.
https://doi.org/10.1016/S0031-0182(00)00254-6
