

 Middle Miocene decapod crustacean assemblage from the Tuzla Basin (Tušanj, Bosnia and Herzegovina), with a description of two new species and comparison with coeval faunas from Slovenia
Middle Miocene decapod crustacean assemblage from the Tuzla Basin (Tušanj, Bosnia and Herzegovina), with a description of two new species and comparison with coeval faunas from Slovenia
Article number: 22.1.9
https://doi.org/10.26879/894
Copyright Paleontological Society, February 2019
Author biographies
Plain-language and multi-lingual abstracts
PDF version
Submission: 23 May 2018. Acceptance: 4 February 2019
{flike id=2430}
ABSTRACT
A decapod assemblage consisting of three species is described from the upper Langhian (lower Badenian) of the Tuzla Basin in Bosnia and Herzegovina. The assemblage is dominated by Retropluma minuta sp. nov. (Brachyura, Retroplumidae) and accompanied by Munidopsis salinaria sp. nov. (Anomura, Galatheoidea) and Portunus monspeliensis A. Milne Edwards, 1860 (Brachyura, Portunidae). Based on the calcareous nannoplankton, the studied assemblage is assigned to the upper part of the NN5 Zone. The studied assemblage is considered to have inhabited a shallow, near-shore depositional setting, as suggested by the microfossil association. Munidopsis, Portunus and Retropluma have previously been reported from Slovenia, the area closest to the Tuzla Basin with known decapod fossils. Contrary to shallow-marine settings of the localities studied herein, Munidopsis and Retropluma from Slovenian localities are reported from deeper-marine settings. The occurrence of near-complete individuals of Retropluma minuta sp. nov. suggests a wider bathymetric range of the genus in its evolutionary past than it has today.
Rok Gašparič. Oertijdmuseum Boxtel, Bosscheweg 80, 5293 WB Boxtel, the Netherlands. rok.gasparic@gmail.com
Matúš Hyžný. Department of Geology and Palaeontology, Faculty of Natural Sciences, Comenius University, Mlynská dolina G1, Ilkovičova 6, Bratislava 842 15, Slovakia; Geological-Paleontological Department, Natural History Museum Vienna, Burgring 7. A-1010 Vienna, Austria. hyzny.matus@gmail.com
Gordana Jovanović. Natural History Musem Belgrade, Njegoševa 51, 11000 Belgrade, Serbia. gordana.j@nhmbeo.rs
Stjepan Ćorić. Geologische Bundesanstalt, Neulinggasse 38, 1030 Wien, Austria. stjepan.coric@geologie.ac.at
Sejfudin Vrabac. Faculty of Mining, Geology and Civil Engineering Tuzla, Univerzitetska 2, 75000 Tuzla, Bosnia and Herzegovina. sejfudin.vrabac@untz.ba
Keywords: Malacostraca; fossil; new species; taphonomy; bathymetry; palaeoenvironment
Final citation: Gašparič, Rok, Hyžný, Matúš, Jovanović, Gordana, Ćorić, Stjepan, and Vrabac, Sejfudin. 2019. Middle Miocene decapod crustacean assemblage from the Tuzla Basin (Tušanj, Bosnia and Herzegovina), with a description of two new species and comparison with coeval faunas from Slovenia. Palaeontologia Electronica 22.1.9A 1-21. https://doi.org/10.26879/894
palaeo-electronica.org/content/2019/2430-decapods-from-the-tuzla-basin
Copyright: February 2019 Paleontological Society.
This is an open access article distributed under the terms of Attribution-NonCommercial-ShareAlike 4.0 International (CC BY-NC-SA 4.0), which permits users to copy and redistribute the material in any medium or format, provided it is not used for commercial purposes and the original author and source are credited, with indications if any changes are made.
creativecommons.org/licenses/by-nc-sa/4.0/
http://zoobank.org/D4051238-1D9D-4F40-9271-70C05A822967
INTRODUCTION
Almost a century has passed since Glaessnerʼs (1928) overview of Neogene decapods from what are now Austria and Slovenia, where he emphasised the need for studying fossil decapod associations in the neighbouring countries. At about the same time, a monograph on fossil decapods of the former Austro-Hungarian Empire was published (Lőrenthey and Beurlen, 1929). The beginning of modern research on fossil decapods from Austria is dated to the second half of the twentieth century with works of Bachmayer (e.g., Bachmayer, 1950, 1953, 1954, 1962; Bachmayer and Tollmann, 1953) and Müller (1984, 1998a, 1998b). Since the hallmark monograph on Badenian (= Langhian-early Serravallian) decapods by Müller (1984), the area once forming the Paratethys Sea (Rögl, 1998, 1999; Harzhauser et al., 2002, 2003) has received renewed attention at the beginning of the twenty-first century. A number of papers dedicated to Neogene decapod crustaceans of the northern part of the Pannonian Basin System include studies on taxonomy, taphonomy, biodiversity, and distribution (e.g., Hyžný and Müller, 2010; Hyžný and Schlögl, 2011; Hyžný et al., 2014, 2015, 2016; Hyžný, 2016). However, there is only limited information about fossil decapods from the southern part of the Pannonian Basin System (Mikuž, 2003; Gašparič and Hyžný, 2015; Gašparič and Ossó, 2016). This paper adds further data on Neogene decapod crustaceans of the latter area.
The earliest report of the Miocene fossil assemblage from the site Dolnja Tuzla in Bosnia and Herzegovina is that by Fuchs (1890), whereas Bittner (1892) reported briefly on a ‘small unidentified crab’ from the same locality. Later, Stevanović and Eremija (1960) reported several ‘well-preserved impressions of crabs’ in the marls of an immediate roof of a salt formation in Tušanj near Tuzla. This material has remained largely unstudied until recently. The present contribution aims to address this fossil decapod faunule in detail following a recent re-study of the material. The stratigraphic age was determined with the microfossil assemblage analysis presented herein and discussion on palaeoenvironmental and palaeobiogeographic implications is also provided.
GEOLOGICAL SETTING
The studied area is located in the Tuzla Basin, in the north-eastern part of Bosnia (Figure 1), which is  known for its rock-salt deposits. The Miocene infill, characterised by various lithologies, reaches a thickness of approximately 2300 m (Ćorić et al., 2007). The accessible exposures represent both siliciclastic and carbonate shallow-water sediments (Stevanović and Eremija, 1960). Middle Miocene (Badenian) sediments in the vicinity of the rock salt mine in Tuzla are exposed across a relatively large area with numerous fossil sites. Microfossil assemblages are dominated by foraminifers (Petrović, 1980; Vrabac and Mihajlović, 1990) and calcareous nannoplankton (Ćorić et al., 2007). Macrofossils are rare and consist of plant remains, foraminifera, pteropods, gastropods, bivalves, decapods, echinoids, and fish (Stevanović and Eremija, 1960; Vrabac, 1990).
known for its rock-salt deposits. The Miocene infill, characterised by various lithologies, reaches a thickness of approximately 2300 m (Ćorić et al., 2007). The accessible exposures represent both siliciclastic and carbonate shallow-water sediments (Stevanović and Eremija, 1960). Middle Miocene (Badenian) sediments in the vicinity of the rock salt mine in Tuzla are exposed across a relatively large area with numerous fossil sites. Microfossil assemblages are dominated by foraminifers (Petrović, 1980; Vrabac and Mihajlović, 1990) and calcareous nannoplankton (Ćorić et al., 2007). Macrofossils are rare and consist of plant remains, foraminifera, pteropods, gastropods, bivalves, decapods, echinoids, and fish (Stevanović and Eremija, 1960; Vrabac, 1990).
Because of salt mine exploration and for understanding of its fossil content, the age of the sedimentary filling of the Tuzla Basin has been discussed for a long time. The sediments were assigned to the middle Miocene (Badenian) based on the study of basin lithostratigraphy and foraminifera (Petrović, 1959; Stevanović and Eremija, 1960; Jovanović, 1980; Soklić et al., 1980; Vrabac et al., 1990). According to the most recent study by Vrabac and Ćorić (2008), the salt formation was deposited during the middle Miocene (early Badenian) in a gulf of the Central Paratethys Sea. Exploratory drilling in the roof of the salt formation in the Tuzla Basin indicates the presence of strata of all three parts of the Badenian stage with overlying Sarmatian sediments (Vrabac et al., 2013). Based on calcareous nannoplankton, the age of the roof of the salt formation from the Tuzla Basin was assigned to the lower Badenian NN5 Zone sensu Martini (1971) (Ćorić et al., 2007).
Decapod specimens described herein come from two localities in the Tuzla Basin: the ventilation shaft ‘Tušanj-2’ and the manhole in Simin Han at Dolnja Tuzla. The localities are positioned approximately 6 km from each other. At both sites, the presence of the roof of the salt formation is documented. Based on Bittner (1892), the initial exploratory salt shaft is dated to 1887; however, the ventilation shaft ‘Tušanj-2’, from which decapod specimens originate, was excavated in the second half of the twentieth century (1953-1967). The shaft ‘Tušanj-2’ is located approximately 30 m from the exploration borehole ‘IB-1’ with the GPS coordinates E 18°40'22" and N 44°32'57". The manhole to the salt mine in Simin Han at Dolnja Tuzla is located at E18°45'01" and N44°31'53".
It is possible to establish the boundaries between the salt formation and its immediate roof, which is located at a depth of 266 m (Vrabac and Mihajlović, 1990). The laminated and massive marls with a dominant foraminiferan assemblage of Ammonia viennensis (d'Orbigny, 1846) and Nonion commune (d'Orbigny, 1846) overlie the salt formation. The first appearance of the foraminifer Uvigerina macrocarinata Papp and Turnovský, 1953, indicates the beginning of the early Badenian in the Central Paratethys (Cicha et al., 1998), and is recorded at about 15 m above the salt formation (Vrabac et al., 2013). The total thickness of these marls is about 150 m. In the Tetima salt deposit, located 10 km northeast of the salt deposits in Tušanj, A. viennensis (d 'Orbigny, 1846) is documented only 3 m above the salt formation (Vrabac et al., 2013). In Tušanj, similarly to the deposit in Tetima, in the first 3 m following the salt formation foraminifera are extremely rare and planktic forms are dominant. From this information, as well as from the borehole ‘IB-1’section, we conclude that the decapod specimens from the ventilation shaft in ‘Tušanj-2’ collected at a depth of 283 m belong to beds from the immediate proximity of the salt formation.
The investigated sediment from ‘Tušanj-2’ contained a rich association of calcareous nannofossils with Sphenolithus heteromorphus Deflandre, 1953. Accompanied nannofossils included Braarudosphaera bigelowii (Gran and Braarud, 1935) Deflandre, 1947; Coccolithus pelagicus (Wallich, 1877) Schiller, 1930; Helicosphaera carteri (Wallich, 1877) Kamptner, 1954; H. walbersdorfensis Müller, 1974; Micrantholithus sp.; Reticulofenestra haqii Backman, 1978; R. minuta Roth, 1970; Rhabdosphaera sicca (Stradner, 1963) Fuchs and Stradner, 1977; and Sphenolithus moriformis (Brönnimann and Stradner, 1960) Bramlette and Wilcoxon, 1967. Reworked Mesozoic specimens included Watznaueria barnesiae (Black in Black and Barnes, 1959) Perch-Nielsen, 1968; and W. fossacincta (Black, 1971) Bown in Bown and Cooper, 1989.
Based on the absence of Helicosphaera ampliaperta Bramlette and Wilcoxon, 1967, the sample can be attributed to the nannoplankton NN5 Zone sensu Martini (1971). The absence of Helicosphaera waltrans Theodoridis, 1984 allows the assignment of the assemblage to the upper part of the NN5 Zone, which can be correlated with the upper part of the Langhian and the upper part of the lower Badenian (Kováč et al., 2007; Piller et al., 2007).
Foraminifers are very rare in the sample from ‘Tušanj-2’ and planktic forms dominate. Several taxa were identified, including Globigerina bulloides d'Orbigny, 1826, Globigerina sp., Quinqueloculina sp., Cibicidoides sp., and Laevidentalina sp.
MATERIAL AND METHODS
Eight decapod specimens were collected from the middle Miocene marls of Tušanj. Seven of them were recovered from the ventilation shaft of the salt mine at a depth of 283 m, whereas a single specimen originates from Dolnja Tuzla. After preparation with fine needles, the specimens were photographed, measured, and further documented using CorelDRAW X5, Adobe Photoshop CC, and Statistica. Photographs were taken with digital cameras Nikon D810 and Nikon Coolpix P7800 under low angle light source conditions. We observed morphological details and measured specimens under stereomicroscope Leica EZ-4D. Because of a delicate nature of the studied specimens they were not whitened prior to photography.
Sieved material from the ‘Tušanj-2’ shaft with granulation 0.1-0.8 mm was used for determination of foraminifera. Samples were analysed by stereomicroscope Technical Carl Zeiss Jena with up to x100 magnification.
Repositories
Material studied herein. Specimens PS-02-08 are a part of the Petar Stevanović Collection housed in the Natural History Museum in Belgrade (Serbia). The specimen GBA 1892/005/0001 is housed in the collections of the Geological Survey in Vienna (Austria).
Comparative material. Retropluma minuta sp. nov. described herein was compared with several fossil Retropluma species. The material of these taxa is deposited in the following institutions: MCZ, Museo Civico “G. Zannato” di Montecchio Maggiore, Italy; MSM, Midtsønderjyllands Museum at Gram, Denmark; MSNM, Museo Civico di Storia Naturale di Milano, Italy; RGA/SMNH, Slovenian Museum of Natural History, Ljubljana, Slovenia.
Abbreviations
Parameters measured: L, carapace length excluding rostrum; LR, total carapace length including rostrum; LG, length of gastric region; LP, length of pleon; WM, maximum carapace width; WA, total width of anterior margin; OW, fronto-orbital width; WP, width of pleon. All measurements are in millimetres.
SYSTEMATIC PALAEONTOLOGY
Order DECAPODA Latreille, 1802
Infraorder ANOMURA MacLeay, 1838
Superfamily GALATHEOIDEA Samouelle, 1819
Family MUNIDOPSIDAE Ortmann, 1898
Genus MUNIDOPSIS Whiteaves, 1874
Type species. Munidopsis curvirostra Whiteaves, 1874, by monotypy.
Remarks. Munidopsis is one of the most speciose decapod genera and includes more than 220 species (Macpherson and Baba, 2011). It is highly diverse morphologically, which led some scholars to recognise several subgenera in the genus (for details see Ahyong et al., 2011 and Robins et al., 2013). Indeed, phylogenetic studies based both on molecular and morphological data clearly show that Munidopsis as currently understood is not monophyletic (Ahyong et al., 2011; Bracken-Grissom et al., 2013). Despite its speciose nature, the genus has a poor fossil record consisting of only a handful of taxa (Hyžný et al., 2014). For a discussion on differentiation of Munidopsis from other exclusively fossil genera, a reference is made to Robins et al. (2013) and references therein.
Munidopsis salinaria sp.nov.
Figure 2.1-2.2
zoobank.org/79B0063D-1425-421F-9970-F559C1C3C047
Diagnosis. Carapace with well-defined hepatic, epigastric, mesogastric, epibranchial and cardiac regions; epigastric regions circular; rostrum styliform, keeled; carapace surface covered with parallel striae, striae forming longer ridges in mesogastric region and posterior part of carapace.
 Etymology. The species epithet refers to the circumstances of the type specimen discovery which was recovered from the exploratory shaft of a salt mine. Latin fem. adj. salinaria, belonging to salt-works.
Etymology. The species epithet refers to the circumstances of the type specimen discovery which was recovered from the exploratory shaft of a salt mine. Latin fem. adj. salinaria, belonging to salt-works.
Holotype. GBA 1892/005/0001--isolated, slightly compressed carapace.
Measurements. L = 6.1; LR = 8.1; LG = 2.5; WM = 6.6; WA = 4.2; WM/L = 1.08.
Description. Carapace subsquare (length excluding rostrum/width is ~ 1); moderately convex transversely. Rostrum styliform, keeled; tip broken. Anterior margin sinuous. Lateral margins slightly convex, with two spines. Grooves well-developed. Circumgastric groove strong; slightly V-shaped in posteriormost part; hepatic and epibranchial branches distinct. Epigastric, hepatic, epibranchial, and cardiac regions well defined. Regions slightly inflated. Hepatic region quadrate with one anterolateral spine epigastric regions circular and inflated. Area of metagastric region not well preserved. Cardiac region broad, U-shaped posteriorly, straight posteriorly. Carapace ornamented with short to moderately long parallel transverse striae, striae forming transverse, squamous ridges in mesogastric and posterior portions of carapace. Posterior margin strongly rimmed, seemingly straight, not well preserved. Ventral surface and appendages not preserved.
Remarks. The only known specimen represents an incomplete dorsal carapace, broken at about mid-length; thus, the area of the metagastric region and posterior carapace margin cannot be described properly. Nevertheless, the specimen is complete enough to allow direct comparison with known fossil and extant congeners. From the area once covered by the Paratethys Sea, two additional species of Munidopsis are known (Hyžný et al., 2014). Munidopsis lieskovensis Hyžný and Schlögl, 2011, from the lower Miocene (Karpatian) of the Slovak part of the Vienna Basin, differs in having a narrower cardiac region and rather uniform carapace ornamentation. Munidopsis salinaria sp. nov. possesses strong squamous ridges on the posterior portions of the carapace; in this respect, it is close to Munidopsis palmuelleri Hyžný, Gašparič, Robins and Schlögl, 2014, from the middle Miocene (lower Badenian) of northern Slovenia. However, M. palmuelleri differs from the new species by its proportionally longer carapace, not well-marked cardiac region, and unkeeled rostrum. In general, the new species shares some characters with M. lieskovensis (keeled rostrum, circular epigastric regions) and other characters with M. palmuelleri (strong squamous ridges on posterior portions of the carapace). Munidopsis salinaria sp. nov. appears to be intermediate between the two taxa mentioned and, therefore, is considered a close relative of both discussed species. This is not surprising given both the geographic proximity of the known occurrences and stratigraphic overlap of all three taxa.
Munidopsis salinaria sp. nov. differs also from its fossil congeners from more distant parts of the world. Munidopsis scrabrosa Feldmann and Wilson, 1988, from the upper Eocene of Antarctica, and M. foersteri Feldmann, Tshudy and Thomson, 1993 from the Upper Cretaceous (Campanian) of Antarctica have a rounded base of the circumgastric groove, which seems to be slightly V-shaped in M. salinaria sp. nov. Moreover, M. foersteri has a pyriform carapace and the rostrum is nearly as long as the carapace (Feldmann et al., 1993), quite unlike many other munidopsids (Ahyong et al., 2011, figure 1). In fact, accommodation of M. foersteri within the family Chirostylidae would be more appropriate, for which the pyriform carapace with much elongated rostrum is typical (Baba et al., 2008, figure 1). Munidopsis scabrosa has scabrose ornamentation directed forwards; that of M. salinaria sp. nov. is much more ridge-like.
From extant congeners, Munidopsis salinaria sp. nov. with its arrangement of carapace regions is similar to the Bathyankyristes group sensu Ahyong et al. (2011) represented by, e.g., M. (B.) levis Alcock and Anderson, 1894. The combination of the characters present in M. salinaria ap. nov., however, is unique and warrants an erection of a new species.
Occurrence. The species is known only from its type locality, i.e., Dolnja Tuzla in Bosnia and Herzegovina.
Infraorder BRACHYURA Latreille, 1802
Superfamily PORTUNOIDEA Rafinesque, 1815
Family PORTUNIDAE Rafinesque, 1815
Subfamily PORTUNINAE Rafinesque, 1815
Genus PORTUNUS Weber, 1795
Type species. Cancer pelagicus Linnaeus, 1758, by subsequent designation of Rathbun (1926).
Remarks. There are more than 40 exclusively fossil species of Portunus reported worldwide (Schweitzer et al., 2010); it must, however, be noted that subgenera are not commonly recognized in the palaeontological practice. As noted by Karasawa et al. (2008) and Spiridonov et al. (2014), the subgenera as recognized by Davie (2002) and Ng et al. (2008) appear to be not closely related, but rather represent distinct clades. In this respect, the fossil record of Portunus sensu lato is in need of revision.
Portunus is a common element in Neogene decapod assemblages, including those reported from the circum-Mediterranean area (Müller, 1984; Gašparič and Ossó, 2016; Díaz-Medina et al., 2018).
Portunus monspeliensis A. Milne Edwards, 1860
Figure 3.1-3.2
1860 Neptunus monspeliensis A. Milne Edwards, p. 232, pl. 4, fig. 1, pl. 5, fig. 1.
1  860 Neptunus granulatus A. Milne Edwards, p. 241, pl. 3, fig. 1, pl. 7, fig. 2.
860 Neptunus granulatus A. Milne Edwards, p. 241, pl. 3, fig. 1, pl. 7, fig. 2.
1929 Neptunus granulatus A. Milne Edwards; Lőrenthey and Beurlen, p. 188, pl. 13, figs. 3-4), pl. 14, figs. 1-4.
1956 Neptunus granulatus A. Milne Edwards; Comaschi Caria, p. 284, 288, pl. 1, figs. 1-7, pl. 2, figs. 1-6, pl. 3, figs. 1-2.
1968 Neptunus granulatus A. Milne Edwards; Stancu and Andreescu, p. 466, pl. 7, fig. 85.
1984 Portunus monspeliensis (A. Milne Edwards); Müller, p. 79, pl. 62, figs. 1-2.
1991 Portunus monspeliensis (A. Milne Edwards); Marras and Ventura, p. 108, pl. 1, figs. 1-4, pl. 2, figs. 1, 4, pl. 3, figs. 1-3.
1993 Portunus monspeliensis (A. Milne Edwards); Müller, p. 14-15, pl. 6, fig. G, pl. 7, fig. A.
2003 Portunus monspeliensis (A. Milne Edwards); Mikuž, p. 187-199, p. 1, figs. 1-5, pl. 2, figs. 1-8.
2007 Portunus monspeliensis (A. Milne Edwards); Artal and Gilles, p. 8, pl. 1, fig. C.
2009 Portunus monspeliensis (A. Milne Edwards); Marangon and De Angeli, p. 5-10, figs. 2-4.
2016 Portunus monspeliensis (A. Milne Edwards); Gašparič and Ossó, p. 55-66, pl. 2, figs. A-H.
2018 Portunus monspeliensis (A. Milne Edwards); Díaz-Medina, Ossó and Hyžný, p. 133, figs. 3A-E. (abbreviated synonymy)
Material. Single partially preserved carapace of a male individual, with preserved sternum and pleon (PS-08).
Measurements. L = 30.5; OW = 27.8; MW = 56.4 (estimated value); L/MW = 0.54.
Description. Carapace hexagonal, incomplete, and heavily weathered, with relative carapace measurements of approximately 1.85 times wider than long. Front protruding and slightly downturned. Fronto-orbital and anterolateral margins poorly preserved. Posterolateral margin presumably straight with concave depression in the last third of its length. Posterior margin broad.
Dorsal carapace surface covered densely by small granules, cuticle not preserved in its entirety. Carapace regions faintly defined; protogastric regions as semi-circular lobes with indistinct transverse ridge; mesogastric and metagastric regions trapezoid in outline and separated by faint transverse ridge. Cardiac region well-defined, pentagonal, somewhat swollen with central depression running longitudinally. Intestinal region faint and circular. Epibranchial region wide, separated from mesobranchial region by arcuate transverse ridge. Cervical groove distinct. Branchiocardiac groove well marked, along sides of cardiac region.
Thoracic sternum broad, oval, widening posteriorly, widest at thoracic sternite 6, with straight to slightly concave sutures between respective sternites. Thoracic sternites 1-2 not preserved, sternites 3-4 fused in trapezoidal plate, axial sulcus reaching anterior margin of sternite 3, transverse ridge in sternite 4 medially interrupted; sternites 5-7 transversely elongated, distally rounded; sternite 6 the longest; sternite 7 shorter than sternite 6; sternite 8 not recognizable; sternites 5-7 expanded laterally into episternites. Pleon broadly subtriangular; pleonites 1-4 not preserved, plenites 5-6 similarly long; telson subtriangular, poorly preserved.
Remarks. Despite the poor preservation of the studied specimen, it is attributed herein to Portunus monspeliensis based on the broadly hexagonal carapace outline, the presence of protogastric keels, and the characteristic sternum (Müller, 1984, pl. 62, figures 1-2; Marangon and De Angeli, 2009, figures 2-4; Gašparič and Ossó, 2016, pl. 1, figures A-H). Portunus monspeliensis is among the most widespread Miocene brachyuran crabs of the circum-Mediterranean area (Lőrenthey and Beurlen, 1929; Gašparič and Ossó, 2016; Díaz-Medina et al., 2018). This report represents the first occurrence of P. monspeliensis from the southern Pannonian Basin of the Central Paratethys.
Superfamily RETROPLUMOIDEA Gill, 1894
Family RETROPLUMIDAE Gill, 1894
Genus RETROPLUMA Gill, 1894
Type species. Archaeoplax notopus Alcock and Anderson, 1894, by monotypy.
Remarks. All species of Retropluma share a set of characteristic carapace features, including a flattened carapace with a more or less quadrangular outline and distinct transverse carinae, a truncate front, exposed orbits, a narrow single-lobed rostrum, pincer-shaped chelipeds, long slender pereiopods (P2-P4), and a reduced setose fifth pereiopod (P5) (McLay, 2006). The fossil species of Retropluma differ in their carapace width to length ratio, dorsal ornamentation, the position of anterolateral teeth, and the shape of the rostrum (Artal et al., 2006; Gašparič and Hyžný, 2015). Other distinctive features such as relative size of ocular peduncles (de Saint Laurent, 1989) are usually not present in the fossil material.
Retropluma is known from the Miocene of Europe, including Slovenia (Gašparič and Hyžný, 2015; Gašparič and Križnar, 2017), Slovakia (Hyžný et al., 2015), and Denmark (Fraaije et al., 2005). The material described below is the first report of the genus from Bosnia.
Retropluma minuta sp. nov.
Figure 4.1-4.6, Figure 5.1-5.4, Figure 6
zoobank.org/6063B8FC-0711-4087-9469-6B3F77B38670
Diagnosis. Carapace square in outline (W/L~1.0), with strong anterolateral tooth pointing slightly outwards. Rostrum rounded and spiny distally. Rounded, swollen bases of antennules extending over the sinuous anterior margin.
 Etymology. The species epithet refers to its minute size for retroplumid crabs. The new species represents the smallest fossil representative of the genus described to date.
Etymology. The species epithet refers to its minute size for retroplumid crabs. The new species represents the smallest fossil representative of the genus described to date.
Holotype. The holotype (PS-03) consists of an articulated near-complete individual preserved in ventral aspect.
Paratypes. PS-02, near-complete and articulated male individual in ventral aspect with preserved anterior; PS-04, near-complete and articulated male in ventral aspect with preserved bases of antennules; PS-06, near-complete and articulated female in ventral aspect with pleon and fifth pereiopods; PS-07, near-complete and articulated individual in ventral aspect with preserved front including rostrum.
Measurements. PS-02: L = 3.9; LP = 2.6; WM = 3.8; WA = 2.1; WP = 1.0; WM/L = 0.97. PS-03: L = 4.1; LP = 2.6; WM = 3.8; WA = 2.6; WP = 1.0; WM/L = 0.98. PS-04: L = 3.9; LP = 2.3; WM = 3.7; WA = 2.4; WP = 1.0; WM/L = 0.95. PS-05: L = 6.7 (estimated value); WM = 6.9 (estimated value); WM/L = 1.03. PS-06: L = 6.1; LP = 3.6; WM = 6.1; WP = 2.3; WM/L = 1.00. PS-07: L = 7.1; LP = 4.5; WM = 7.5; WA = 5.6 (estimated value); WP = 2.0; WM/L = 1.03.
 Description. Carapace subsquare (WM/L = 0.97-1.03). Rostrum bent slightly downward, rounded and spiny distally. Supraorbital margin sinuous, with indistinct supraorbital tooth and reduced orbits. Postorbital tooth strong and triangular, pointed forwards. Rounded, swollen bases of antennules extending over the anterior margin and filling entire orbital area beneath the rostrum. Anterolateral margin straight till anterior carina. Lateral and posterolateral margins slightly convex. Posterior margin strong and rimmed, with weakly developed re-entrant for fifth pereiopods (P5).
Description. Carapace subsquare (WM/L = 0.97-1.03). Rostrum bent slightly downward, rounded and spiny distally. Supraorbital margin sinuous, with indistinct supraorbital tooth and reduced orbits. Postorbital tooth strong and triangular, pointed forwards. Rounded, swollen bases of antennules extending over the anterior margin and filling entire orbital area beneath the rostrum. Anterolateral margin straight till anterior carina. Lateral and posterolateral margins slightly convex. Posterior margin strong and rimmed, with weakly developed re-entrant for fifth pereiopods (P5).
Morphological characters of dorsal carapace only partially preserved. Three transverse carinae discernible on lateral portions of carapace. Carinae forming ridges at intersection with lateral margin. Carapace widest mid-length at median carina. Surface of thoracic sternites pitted and finely granulose, with enlarged granules laterally and on ridges.
Female pleon broadly triangular, with straight margins, narrowing anteriorly, somites becoming narrower and longer anteriorly, telson longer than wide and terminally rounded. Male pleon narrower than that of female. Somite 2 wide, with strong sinuous transverse ridge. Posterior margin straight, anterior margin strongly concave. Lateral margins almost triangular. Male pleon widest at transverse ridge of somite 2. Somites 3-5 fused, sutures 3/4 and 4/5 visible. Lateral margins of fused 3-5 segment straight, narrowing distally. Somite 6 longer than wide, with sinuous lateral margins, convex posteriorly and concave anteriorly, posterolateral corners bluntly rounded. Distinct transverse ridge in anterior part of somite 6, anterior margin with telson slightly convex and centrally rimmed. Telson narrow and long, with rounded termination. Female sternal plate in outline more round than that of male, sternites 5-7 long, convex and ridged. Sternite 8 reduced and covered by pleon.
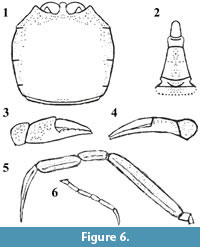 Chelipeds (P1) long, slender and distinctly unequal. Right cheliped slender, propodus and dactylus curved, narrowing distally, both fingers long and forceps-like, lateral surfaces without ornamentation. Occlusal margin of right cheliped fingers straight and unarmed. Left cheliped noticeably more robust than right cheliped. Fingers stouter, both with several large blunt teeth. Carpus and merus stout and adorned with tubercles. P2-P4 achelate, much longer than chelipeds, P3 longest, surface finely granulose and flattened, with central groove running along the whole length, ending in long blade-like dactyli. P5 strongly reduced and thin, length about one third of P3.
Chelipeds (P1) long, slender and distinctly unequal. Right cheliped slender, propodus and dactylus curved, narrowing distally, both fingers long and forceps-like, lateral surfaces without ornamentation. Occlusal margin of right cheliped fingers straight and unarmed. Left cheliped noticeably more robust than right cheliped. Fingers stouter, both with several large blunt teeth. Carpus and merus stout and adorned with tubercles. P2-P4 achelate, much longer than chelipeds, P3 longest, surface finely granulose and flattened, with central groove running along the whole length, ending in long blade-like dactyli. P5 strongly reduced and thin, length about one third of P3.
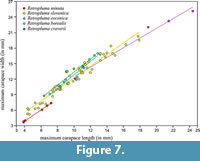
Remarks. The studied material consists of six articulated specimens preserved in ventral aspect. Original cuticle, although preserved, is covered by coarse mineral grains which obliterate finer details. Although dorsal carapace features are incomplete, there are enough distinguishing characters to warrant the erection of a new species. Retropluma minuta sp. nov. differs from its fossil congeners, i.e., Retropluma borealis Fraaije, Hansen and Hansen, 2005; R. craverii (Crema, 1895); R. eocenica Vía Boada, 1959; and R. slovenica Gašparič and Hyžný, 2014, not only by its minute size, but more importantly, by a distinctly square carapace outline, and the smallest width to length ratio (Figure 7; for raw data see Appendix 1). Retropluma gallica Artal, Van Bakel and Castillo, 2006, was not included in the comparison because of lack of sufficient data (i.e., available carapace measurements). Additionally, Retropluma minuta sp. nov. is characterised by a sinuous anterior margin with reduced orbits and enlarged, rounded bases of antennules and a unique male pleon with strong ridge on somite 2.
 Of the previously described fossil representatives of Retropluma, R. minuta sp. nov. is similar to Retropluma slovenica (Figure 5.5-5.6) from the early Miocene of Slovenia (Gašparič and Hyžný, 2015) and the middle Miocene of Slovakia (Hyžný et al., 2015). The new species differs from R. slovenica in having a subsquare carapace (WM/L = 0.98-1.03) whereas it is wider in R. slovenica (WM/L = 1.24), and having a smaller rounded and spinous rostrum. On the anterior margin of R. minuta sp. nov. are visible greatly enlarged rounded bases of antennules (present also among extant species, e.g., McLay, 2006, figure 3A) and reduced ocular orbits, unlike the specimens of R. slovenica. Additionally, R. minuta sp. nov. displays a uniquely developed and strongly ridged male pleonal somite 2 and possesses distinctly unequal chelipeds, with a much stronger left cheliped (preserved in three studied specimens) with a dentate occlusal margin, whereas the chelipeds in R. slovenica are of sub-equal size with left cheliped being somewhat slenderer. Pereiopods 2-4 are longer compared to carapace size in R. minuta sp. nov. and the blade-like dactyli are also markedly longer. Retropluma minuta sp. nov. also has a more convex posterior margin than R. slovenica.
Of the previously described fossil representatives of Retropluma, R. minuta sp. nov. is similar to Retropluma slovenica (Figure 5.5-5.6) from the early Miocene of Slovenia (Gašparič and Hyžný, 2015) and the middle Miocene of Slovakia (Hyžný et al., 2015). The new species differs from R. slovenica in having a subsquare carapace (WM/L = 0.98-1.03) whereas it is wider in R. slovenica (WM/L = 1.24), and having a smaller rounded and spinous rostrum. On the anterior margin of R. minuta sp. nov. are visible greatly enlarged rounded bases of antennules (present also among extant species, e.g., McLay, 2006, figure 3A) and reduced ocular orbits, unlike the specimens of R. slovenica. Additionally, R. minuta sp. nov. displays a uniquely developed and strongly ridged male pleonal somite 2 and possesses distinctly unequal chelipeds, with a much stronger left cheliped (preserved in three studied specimens) with a dentate occlusal margin, whereas the chelipeds in R. slovenica are of sub-equal size with left cheliped being somewhat slenderer. Pereiopods 2-4 are longer compared to carapace size in R. minuta sp. nov. and the blade-like dactyli are also markedly longer. Retropluma minuta sp. nov. also has a more convex posterior margin than R. slovenica.
Retropluma is represented by seven extant species, mostly from the Indo-Pacific region (Gašparič and Hyžný, 2015, table 4). Retropluma minuta sp. nov. resembles closely the extant Retropluma laurentae McLay, 2006, in general outline of the carapace, possession of a small rounded rostrum, a forward pointing triangular postorbital tooth and swollen, rounded bases of antennules. The carapace outline of R. minuta sp. nov. is subsquare, whereas it is rectangular in R. laurentae and the anterior margin of R. minuta sp. nov. is more sinuous with noticeable supraorbital tooth.
Occurrence. The species is known only from its type locality, i.e., the ventilation shaft of the salt mine Tušanj-2, Bosnia and Herzegovina.
DISCUSSION
Decapod Taphonomy
Decapod crustaceans have a relatively low fossilisation potential in comparison to more calcified mollusc shells (Kidwell and Flessa, 1995; Stempien, 2005). Consequently, decapods are subject to rapid decomposition, and the fragile exoskeleton is often fragmented before burial takes place (Plotnick et al., 1988; Stempien, 2005; Krause et al., 2011; Klompmaker et al., 2017).
All examined specimens of Retropluma minuta sp. nov. are preserved in ventral aspect. Their relaxed position (appendages aligned along the carapace sides and chelipeds closed; cf. Bishop, 1986, p. 331) suggests that they represent corpses, which indicates that the retroplumid specimens are autochthonous. No carapace displacement could be observed, which is otherwise characteristic for preservation of crab moults, in which the pleon is not connected to the carapace anymore (Schäfer, 1972; Bishop, 1986). However, the moulted carapace may flip back (Schäfer, 1972), especially in a low-energy environment, so the resulting remains are impossible to distinguish from the corpse. Thus, we cannot say for sure whether the specimens represent corpses or moults. Whatever the real nature of the specimens, the preservation of delicate appendages still attached to the crab body does not suggest post-mortem transport.
Studied decapod specimens from the Tuzla Basin show two basic modes of preservation. The squat lobster Munidopsis and the brachyuran crab Portunus are preserved as isolated carapaces without appendages, whereas retroplumid crabs are complete and virtually intact. This observation has further implications: retroplumid crabs are to be considered autochthonous, whereas isolated carapace of Munidopsis and insufficiently preserved specimen of Portunus monspeliensis might have been transported prior to final burial. However, such transport had to be rather short; otherwise fragile carapaces would not be preserved at all.
Environment Preferences of Decapods
Although the squat lobster Munidopsis is considered a typical deep-water genus (Macpherson, 2007), squat lobsters in general inhabit a wide range of habitats from the intertidal zone to bathyal depths (Baba et al., 2008) and are a common element of benthic communities on coral reefs or rocky and muddy bottoms (Lowrich and Thiel, 2011). Moreover, there are fossil representatives of Munidopsis reported from shallow marine settings (Feldmann and Wilson, 1988; Feldmann et al., 1993).
Many extant portunids, such as Portunus pelagicus (Linnaeus, 1758), are found in the Indo-Pacific waters and even in the eastern margin of the Mediterranean Sea (Lai et al., 2010). Judging from the lithologies (sandstones and sandy limestones interbedded with marls) bearing fossils of Portunus monspeliensis (Gašparič and Ossó, 2016), and comparing to extant P. pelagicus (Chande and Mgaya, 2003; Kunsook et al., 2014), it can be concluded that P. monspeliensis inhabited a wide range of habitats, but preferred sublittoral algal and sea grass meadows on both sandy and muddy substrate. Based on the cluster analysis, Müller (1984, table 4) estimated the bathymetry of sedimentary settings with abundant Portunus remains as deposited in a shallow marine environment with 20-30 m water depth.
Extant representatives of Retropluma preferentially inhabit muddy or sandy bottoms at a depth of 70 to 450 m (de Saint Laurent, 1989; McLay, 2006; see also Gašparič and Hyžný, 2015, table 4), where they spend most of the time buried in soft substrate (Ahyong, 2008). Retropluma slovenica from the early Miocene of Slovenia was recorded from settings interpreted to have been deposited in waters deeper than 125 m (Gašparič and Hyžný, 2015), and a similar depth was estimated for the palaeoenvironment at the middle Miocene locality of Slovakia where R. slovenica was also found (Hyžný et al., 2015).
Palaeoenvironment and Bathymetry
The middle Miocene transgression occurred in the area of what is now northern Bosnia and Herzegovina during the early Badenian, having its base at 14.91 Ma (Ćorić et al., 2009; Pezelj et al., 2013; see also Kováč et al., 2017a, 2017b) as a result of the increase in global sea level. The gastropod Sveltia cf. dertovaricosa (Sacco, 1894) and the pteropod Vaginella austriaca, Kittl, 1886, were found in the ventilation shaft ‘Tušanj-2’ at 283 m depth, together with the decapod remains and internal moulds of cancellariid snails. The mollusc assemblage suggests fully marine settings. The foraminiferal assemblages of ‘Tušanj-2’ indicate that the investigated marls of Tušanj were deposited in very shallow sublittoral settings with the water depth not exceeding 10 m. For comparison, numerous miliolid foraminifers inhabit the Gulf of Florida today, in association with nonionids and rotalid foraminifera, at depths of up to 3 m (Moore, 1957). Quinqueloculina with Ammonia are characteristic for shallow inner shelf settings (Haq and Boersma, 1998). Similarly, the absence of discoasters and the dominance of Reticulofenestra minuta, Helicosphaera carteri, and Coccolithus pelagicus in the nannoplankton assemblage point to a nutrient-rich, shallow-marine environment (Ćorić et al., 2009).
Interestingly, the investigated decapod assemblage from the Tuzla Basin consists of taxa not commonly found in shallow-marine settings. Besides Portunus, a typical inhabitant of shallow waters, there are also specimens of Retropluma. The occurrence of articulated specimens of the latter genus in sublittoral settings is rather surprising. Although some Eocene retroplumoids have been reported from shallow-marine settings (Beschin et al., 1996; Khodaverdi et al., 2016), these taxa represent genera other than Retropluma, whereas Eocene representatives of Retropluma are known from outer-shelf habitats (Artal et al., 2006).
Comparison with Miocene Decapod Assemblages from Slovenia
Paratethyan decapod faunas are rather homogeneous across respective basins (Hyžný, 2016); in fact, the Paratethys has been shown to represent a compact area without further biogeographic fragmentation (Harzhauser et al., 2002, 2003). Thus, not surprisingly, the decapod assemblage from the Tuzla Basin of Bosnia and Herzegovina described herein is very close taxonomically to decapod associations from the geographically proximal area of Slovenia. All three genera, Munidopsis, Portunus, and Retropluma, have been previously reported from Slovenian early and middle Miocene localities (Hyžný et al., 2014; Gašparič and Hyžný, 2015; Gašparič and Ossó, 2016). There are, however, distinct differences in the inferred palaeoenvironment of localities with respective decapod taxa (Table 1). Although in all cases muddy sea bottoms with predominantly siliciclastic depositional settings were documented, the water depth apparently was not similar. Whereas Retropluma slovenica and Munidopsis palmuelleri are considered to derive from a deep water environment (Hyžný et al., 2014; Gašparič and Hyžný, 2015), the association from the Tuzla Basin dominated by Retropluma minuta sp. nov. occurred in a shallow-water, nutrient rich palaeoenvironment.
CONCLUSIONS
A fossil decapod assemblage consisting of three species is described from the upper Langhian (lower Badenian) of the Tuzla Basin. The occurrence is the first formal report of fossil decapod crustaceans from Bosnia and Herzegovina. The decapod assemblage is dominated by Retropluma minuta sp. nov. (Brachyura, Retroplumidae) and accompanied by Munidopsis salinaria sp. nov. (Anomura, Galatheoidea) and Portunus monspeliensis (Brachyura, Portunidae). Based on the microfossil association, the studied assemblage is considered to have once inhabited a shallow, near-shore depositional setting. The occurrence of near-complete individuals of Retropluma in the Miocene shallow-marine settings of the Tuzla Basin suggests wider bathymetric range of the genus than it displays today. All three genera reported from the Tuzla Basin, namely Munidopsis, Portunus and Retropluma, have previously been reported from Slovenia, the most proximal area with known decapod fossils to that of the Tuzla Basin in Bosnia and Herzegovina. Contrary to shallow-marine settings of the localities of the Tuzla Basin, Munidopsis and Retropluma from Slovenian localities are reported from deeper-marine settings.
ACKNOWLEDGEMENTS
We wish to thank I. Zorn (Geologische Bundesanstalt, Wien) for help and information on material deposited at the GBA, V. Mikuž (Faculty of Natural Sciences and Engineering, University of Ljubljana) and M. Križnar (Museum of Natural History, Ljubljana) for their support, A. Žibrat Gašparič (Faculty of Arts, University of Ljubljana) for her linguistic support and proofreading of the manuscript. The manuscript benefitted from constructive criticism of three anonymous reviewers. The research of MH was supported by the Austrian Science Fund (FWF; Lise Meitner Program M 1544-B25), Slovak Research and Development Agency under contract nos. APVV-16-0121 and APVV-17-0555, and by Hungarian Scientific Research Fund (OTKA K112708).
REFERENCES
Ahyong, S.T. 2008. Deepwater crabs from seamounts and chemosynthetic habitats off eastern New Zealand (Crustacea: Decapoda: Brachyura). Zootaxa, 1708:1-72.
Ahyong, S.T., Andreakis, N., and Taylor, J. 2011. Mitochondrial phylogeny of the deep-sea squat lobsters, Munidopsidae (Galatheoidea). Zoologischer Anzeiger, 250:367-377. https://doi.org/10.1016/j.jcz.2011.06.005
Alcock, A. and Anderson, A.R. 1894. Natural History notes from H. M. Indian Marine Survey Steamer “Investigator”, Commander C.F. Oldham, R.N., commanding. Series II, No. 14. An account of a recent collection of deep-sea Crustacea from the Bay of Bengal and Laccadive Sea. Journal of the Asiatic Society of Bengal, 63:141-185.
Artal, P. and Gilles, A. 2007. New Miocene crabs from Pignon (southeast France). Memorie della Societa Italiana di Scienze Naturali e del Museo Civico di Storia Naturale di Milano, 35:8-11.
Artal, P., Van Bakel, B., and Castillo, J. 2006. Retropluma Gill, 1894 (Crustacea, Decapoda) from the Eocene of the eastern Pyrenees (Spain, France). Cainozoic Research, 5:65-71.
Baba, K., Macpherson, E., Poore, G.C.B., Ahyong, S.T., Bermudez, A., Cabezas, P., and Lin, Ch.-W. 2008. Catalogue of squat lobsters of the world (Crustacea: Decapoda: Anomura-families Chirostylidae, Galatheidae and Kiwaidae). Zootaxa, 1905:1-220.
Bachmayer, F. 1950. Neue Dekapoden aus dem österreichischen Tertiär. Annalen des Naturhistorischen Museums in Wien, 57:133-140.
Bachmayer, F. 1953. Die Dekapodenfauna des tortonischen Leithakalkes von Deutsch-Altenburg (Niederösterreich). Mitteilungen der Geologischen Gesellschaft in Wien, 44 (for 1951):237-262.
Bachmayer, F. 1954. Zwei bemerkenswerte Crustaceen-Funde aus dem Jungtertiär des Wiener Beckens. Sitzungsberichte der Mathematisch-Naturwissenschaftlichen Klasse der Kaiserlichen Akademie der Wissenschaften, Wien, 163(1-2):63-70.
Bachmayer, F. 1962. Die Calappiden (Crustacea, Decapoda) aus den tortonischen Ablagerungen des Wiener Beckens. Annalen des Naturhistorischen Museums in Wien, 65:39-46.
Bachmayer, F. and Tollmann, A. 1953. Die Crustaceen-Fauna aus dem tortonischen Leithakalk (Steinbrüche der Firma Fenk) bei Groß-Höflein im Burgenland, p. 308-314. In (editor unknown) Kober-Festschrift 1953, Skizzen zum Antlitz der Erde. Geologische Arbeiten, herausgegeben aus Anlaβ des 70. Geburtstages von Prof. Dr. L. Kober, Universität Wien, Gesellschaftsbuchdruckerei Brüder Hollinek, Wien.
Backman, J. 1978. Late Miocene-Early Pliocene nannofossil biochronology and biogeography in the Vera Basin, SE Spain. Stockholm Contributions in Geology, 32:93-114.
Beschin, C., Busulini, A., De Angeli, A., and Tessier, G. 1996. Retroplumoidea (Crustacea, Brachyura) nel Terziario del Vicentino (Italia settentrionale). Lavori-Società Veneziana di Scienze Naturali, 21:83-102.
Bishop, G. A. 1986. Taphonomy of the North American decapods. Journal of Crustacean Biology, 6:326-355. https://doi.org/10.1163/193724086X00190
Bittner, A. 1892. A Petrefakten des marinen Neogens von Dolnja Tuzla in Bosnien. Verhandlungen der k.k. Geologischen Reichsanstalt, 7:189-183.
Black, M. 1971. Coccoliths of the Speeton Clay and Sutterby Marl. Proceedings of the Yorkshire Geological Society, 38:381-424. https://doi.org/10.1144/pygs.38.3.381
Black, M. and Barnes, B. 1959. The structure of coccoliths from the English Chalk. Geological Magazine, 96:321-328. https://doi.org/10.1017/S0016756800062294
Bown, P.R. and Cooper, M.K.E. 1989. Jurassic, p.82-115. In Bown, P.R. (ed.), Calcareous Nannofossil Biostratigraphy. British Micropalaeontological Society Publication Series. Chapman & Hall, London.
Bracken-Grissom, H.D., Cannon, M.E., Cabezas, P., Feldmann, R.M., Schweitzer, C.E., Ahyong, S.T., Felder, D.L., Lemaitre, R., and Crandall, K.A. 2013. A comprehensive and integrative reconstruction of evolutionary history for Anomura (Crustacea: Decapoda). BMC Evolutionary Biology, 13:128 (28 pages). https://doi.org/10.1186/1471-2148-13-128
Bramlette, M.N. and Wilcoxon, J.A. 1967. Middle Tertiary calcareous nannoplankton of the Cipero section, Trinidad, W.I. Tulane Studies in Geology and Paleontology, 5:93-131.
Brönnimann, P. and Stradner, H. 1960. Die Foraminiferen- und Discoasteriden-zonen von Kuba und ihre interkontinentale Korrelation. Erdoel-Zeitschrift, 76(10):364-369.
Chande, A.I. and Mgaya, Y.D. 2003. The fishery of Portunus pelagicus and species diversity of portunid crabs along the coast of Dar es Salaam, Tanzania. Western Indian Ocean Journal of Marine Science, 2/1:75-84. https://doi.org/10.4314/wiojms.v2i1.28431
Cicha, I., Rögl, F., Rupp, C., and Čtyroká, J. 1998. Oligocene-Miocene foraminifera of the Central Paratethys. Abhandlungen der Senckenbergischen Naturforschenden Gesellschaft, 549:1-325.
Comaschi Caria, I. 1956. I Crostacei miocenici della Sardegna. Bollettino del Servizio Geologico d’Italia, 78:283-290.
Ćorić, S., Pavelić, D., Rögl, F., Mandić, O., Vrabac, S., Avanić, R., Jerković, L., and Vranjković, A. 2009. Revised Middle Miocene datum for initial marine flooding of North Croatian Basins (Pannonian Basin System, Central Paratethys). Geologia Croatica, 62:31-43.
Ćorić, S., Vrabac, S., Ferhatbegović, Z., and Ðulović, I. 2007. Biostratigraphy of Middle Miocene Sediments from the Tuzla Basin (North-eastern Bosnia) Based on Foraminifera and Calcareous Nannoplankton. Joannea Geologie und Paläontologie, 9:21-23.
Crema, C. 1895. Sopra alcuni decapodi terziarii del Piemonte. Atti Reale Accademia delle Scienze di Torino, 30:664-681.
Davie, P.J.F. 2002. Crustacea: Malacostraca: Eucarida (Part 2): Decapoda--Anomura, Brachyura, p. 1-641. In Wells, A. and Houston, W.W.K. (eds.), Zoological Catalogue of Australia. Vol. 19.3B. CSIRO Publishing, Melbourne.
De Angeli, A., Garassino, A., and Pasini, G. 2011. Retropluma craverii (Crema, 1895) (Crustacea, Decapoda, Brachyura, Retroplumidae) from the Pliocene of Reggio Emilia (N Italy). Atti della Società Italiana di Scienze Naturali e del Museo Civico di Storia Naturale di Milano, 152:37-44.
Deflandre, G. 1947. Braarudosphaera nov. gen., type d'une famille nouvelle de Coccolithophorides actuels a elements composites. Comptes Rendus Hebdomadaires des Séances de l'Académie des Sciences, 225:439-441.
Deflandre, G. 1953. Hétérogénéité intrinsèque et pluralité des éleménts dans les coccolithes actuels et fossiles. Comptes Rendus Hebdomadaires des Séances de l'Académie des Sciences Paris, 237:1785-1787.
de Saint Laurent, M. 1989. La nouvelle superfamille des Retroplumoidea Gill, 1894 (Decapoda, Brachyura): systématique, affinités et évolution. Mémoire du Muséum national d’Histoire Naturelle, 144:103-179.
Díaz-Medina, G., Ossó, À., and Hyžný, M. 2018. A middle Miocene decapod faunule from Granada (Spain), with remarks on distribution pattern of the crab Portunus monspeliensis. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen, 288:129-141. https://doi.org/10.1127/njgpa/2018/0728
Feldmann, R.M., Tshudy, D.M., and Thomson, M.R.A. 1993. Late Cretaceous and Paleocene decapod crustaceans from James Ross Basin, Antarctic Peninsula. The Paleontological Society Memoir, 28:1-41.
Feldmann, R.M. and Wilson, M.T. 1988. Eocene decapod crustaceans from Antarctica. Memoirsof Geological Society of America, 169:465-488.
Fraaije, R.H.B., Hansen, J., and Hansen, T. 2005. Late Miocene decapod fauna from Gram, Denmark. Palaeontos, 7:51-61.
Fuchs, R. and Stradner, H. 1977. Über Nannofossilien im Badenien (Mittelmiozan) der zentralen Paratethys. Beiträge zur Paläontologie von Österreich, 2:1-58.
Fuchs, Th. 1890. Einsendungen von Petrefacten aus Bosnien von Dolnja Tuzla. Annalen des k. k. Naturhistorischen Hofmuseums, Notizen, 5:86.
Gašparič, R. and Hyžný, M. 2015 (online 2014). An early Miocene deep-water decapod crustacean faunule from the Slovenian part of the Styrian Basin, and its palaeoenvironmental and palaeobiogeographical significance. Papers in Palaeontology, 1:141-166. https://doi.org/10.1002/spp2.1006
Gašparić, R. and Križnar, M. 2017. Early Miocene decapod Retropluma slovenica Gašparič & Hyžný, 2014 from Govce beds of Tunjice Hills (Central Slovenia). Geologija, 59:55-66. https://doi.org/10.5474/geologija.2017.006
Gašparić, R. and Ossó, A. 2016. New reports of decapod Portunus monspeliensis A. Milne Edwards, 1860 from Miocene beds of eastern Slovenia with notes on palaeoecology and palaeobiogeography. Geologija, 60:77-85. http://doi.org/10.5474/geologija.2016.005
Gill, T. 1894. A new bassalian type of crabs. American Naturalist, 28:1034-1045.
Glaessner, M.F.1928. Die Dekapodenfauna des österreichischen Jungtertiärs. Jahrbuch der Geologischen Bundesanstalt, 78:161-219.
Gran, H.H. and Braarud, T. 1935. A quantitative study of the phytoplankton in the Bay of Fundy and the Gulf of Maine (including observations on hydrography, chemistry and turbidity). Journal of the Biological Board of Canada, 1:279-467. https://doi.org/10.1139/f35-012
Haq, B.U. and Boersma, A. 1998. Introduction to Marine Micropaleontology. Elsevier, Amsterdam and Tokyo.
Harzhauser, M., Mandic, O., and Zuschin, M. 2003. Changes in Paratethyan marine molluscs at the Early/Middle Miocene transition: diversity, palaeogeography and palaeoclimate. Acta Geologica Polonica, 53:323-339.
Harzhauser, M., Piller, W.E., and Steininger, F.F. 2002. Circum-Mediterranean Oligo-Miocene biogeographic evolution - the gastropods’ point of view. Palaeogeography, Palaeoclimatology, Palaeoecology, 183:103-133. https://doi.org/10.1016/S0031-0182(01)00464-3
Hyžný, M. 2016. Diversity and distribution patterns of the Oligocene and Miocene decapod crustaceans (Crustacea: Malacostraca) of the Western and Central Paratethys. Geologica Carpathica, 67:471-494. https://doi.org/10.1515/geoca-2016-0030
Hyžný, M., Gašparič, R., Robins, C.M., and Schlögl, J. 2014. Miocene squat lobsters (Decapoda, Anomura, Galatheoidea) of the Central Paratethys - a review, with description of a new species of Munidopsis. Scripta Geologica, 147:241-267.
Hyžný, M., Hudáčková, N., and Szalma, Š. 2015. Taphonomy and diversity of Middle Miocene decapod crustaceans from the Novohrad-Nógrad Basin, Slovakia, with remarks on palaeobiography. Acta Geologica Slovaca, 7:139-154.
Hyžný, M., Mandic, O., Harzhauser, M., and Ledvák, P. 2016. Euryhaline preferences of the decapod crab Mioplax socialis enabled it to survive the Badenian/Sarmatian extinction (Miocene) in the Central Paratethys. Geologica Carpathica, 67:165-176. https://doi.org/10.1515/geoca-2016-0011
Hyžný, M. and Müller, P.M. 2010. The first fossil record of the genus Callichirus (Decapoda, Axiidea, Callianassidae) from the middle Miocene of Hungary, with description of a new species. Bulletin of the Mizunami Fossil Museum, 36:37-43.
Hyžný, M. and Schlögl, J. 2011. An early Miocene deep-water decapod crustacean faunule from the Vienna basin (Western Carpathians, Slovakia). Palaeontology, 54:323-349. https://doi.org/10.1111/j.1475-4983.2011.01033.x
Jovanović, Č. 1980. Geneza pretortonskih miocenskih sedimenata na prostoru između Drine i Une. Geološki Glasnik, 15:1-159.
Kamptner, E. 1954. Untersuchungen über den Feinbau der Coccolithen. Anzeiger. Österreichische Akademie der Wissenschaften. Mathematische-Naturwissenschafliche Klasse, 87:152-158.
Karasawa, H., Schweitzer, C.E., and Feldmann, R.M. 2008. Revision of Portunoidea Rafinesque, 1815 (Decapoda: Brachyura) with emphasis on the fossil genera and families. Journal of Crustacean Biology, 28:82-127. https://doi.org/10.1651/07-2882R.1
Khodaverdi Hassan-vand, M., Bahrami, A., Yazdi, M., Ossó, À., Safari, A., Martínez, J.L., and Vega, F.J. 2016. Occurrence of Retrocypoda almelai Via Boada, 1959 (Decapoda: Retroplumidae) in the Eocene of Central Iran. Paleontología Mexicana, 5:21-31.
Kidwell, S.M. and Flessa, K.W. 1995. The quality of the fossil record: populations, species, and communities. Annual Review of Earth and Planetary Sciences, 24:433-464. https://doi.org/10.1146/annurev.earth.24.1.433
Kittl, E. 1886. Ueber die Miozanen Pteropoden von Oesterreich- Ungarn mit Berücksihtigung verwandter Vorkommnise der Nachbarnländer. Annalen des k. k. Naturhistorischen Hofmuseums Wien, 1:47-74.
Klompmaker, A.A., Portell, R.W., and Frick, M.G. 2017. Comparative experimental taphonomy of eight marine arthropods indicates distinct differences in preservation potential. Palaeontology, 60:773-794. https://doi.org/10.1111/pala.12314
Kováč, M., Andreyeva-Grigorovich, A., Bajraktarević, Z., Brzobohatý, R., Filipescu, S., Fodor, L., Harzhauser, M., Nagymarosy, A., Oszczypko, N., Pavelić, D., Rögl, F., Saftić, B., Sliva, Ľ., and Studencka, B. 2007. Badenian evolution of the Central Paratethys Sea: paleogeography, climate and eustatic sea-level changes. Geologica Carpathica, 58:579-606.
Kováč, M., Hudáčková, N., Halásová, E., Kováčová, M., Holcová, K., Oszczypko-Clowes, M., Báldi, K., Less, G., Nagymarosy, A., Ruman, A., Klučiar, T., and Jamrich, M. 2017a. The Central Paratethys palaeoceanography: a water circulation model based on microfossil proxies, climate, and changes of depositional environment. Acta Geologica Slovaca, 9:75-114.
Kováč, M., Márton, E., Oszczypko, N., Vojtko, R., Hók, J., Králiková, S., Plašienka, D., Klučiar, T., Hudárková, N., and Oszczypko-Clowes, M. 2017b. Neogene palaeogeography and basin evolution of the Western Carpathians, Northern Pannonian domain and adjoining areas. Global and Planetary Change, 155:133-154. https://doi.org/10.1016/j.gloplacha.2017.07.004
Krause, R.A. Jr., Parsons-Hubbard, K., and Walker, S.E. 2011. Experimental taphonomy of a decapod crustacean: Long-term data and their implications. Palaeogeography, Palaeoclimatology, Palaeoecology, 312:350-362. https://doi.org/10.1016/j.palaeo.2011.03.020
Kunsook, C., Gajaseni, N., and Paphavasit, N. 2014. The feeding ecology of the blue swimming crab, Portunus pelagicus (Linnaeus, 1758), at Kung Krabaen Bay, Chanthaburi Province, Thailand. Tropical Life Sciences Research, 25:13-27.
Lai, J.C.Y., Ng, P.K.L., and Davie, P.J.F. 2010. A Revision of the Portunus pelagicus (Linneus, 1758) species complex (Crustacea: Brachyura: Portunidae), with the recognition of four species. Raffles Bulletin of Zoology, 58:199-237.
Larghi, C. 2003. First record of Oligocene retroplumd crab (Crustacea: Decapoda: Brachyura) from Italy. Bulletin of the Mizunami Fossil Museum, 30:57-60.
Latreille, P.A. 1802. Histoire Naturelle, Général et Particulière, des Crustacés et des Insectes, Volume 3. Familles naturelles des genres. Dufart, Paris.
Linnaeus, C. 1758. Systema Naturae per Regna Tria Naturae, Secundum Classes, Ordines, Genera, Species, cum Characteribus, Differentiis, Synonymis, Locis. Editio Decima, Reformata. Laurentius Salvius, Holmiae.
Lőrenthey, E. and Beurlen, K. 1929. Die fossilen Dekapoden der Länder der ungarischen Krone. Geologica Hungarica, Series Palaeontologica, 3:1-420.
Lowrich, G.A. and Thiel, M. 2011. Ecology, physiology, feeding and trophic role of squat lobsters, p. 183-222. In Poore, G.C.B., Ahyong, S.T., and Taylor, J. (eds.), The Biology of Squat Lobsters. CSIRO Publishing, Melbourne.
MacLeay, W.S. 1838. On the brachyurous decapod Crustacea brought from the Cape by Dr. Smith, p. 53-71. In Smith, A. (ed.), Illustrations of the Zoology of South Africa, Consisting Chiefly of Figures and Descriptions of the Objects of Natural History Collected during an Expedition into the Interior of South Africa, in the Years 1834, 1835, and 1836; Fitted Out by "The Cape of Good Hope Association for Exploring Central Africa": Together with a Summary of African Zoology, and an Inquiry Into the Geographical Ranges of Species in that Quarter of the Globe, Volume 5. Smith, Elder and Co., London.
Macpherson, E. 2007. Species of the genus Munidopsis Whiteaves, 1784 from the Indian and Pacific Oceans and reestablishment of the genus Galacantha A. Milne-Edwards, 1880 (Crustacea, Decapoda, Galatheidae). Zootaxa, 1417:1-135.
Macpherson, E. and Baba, K. 2011. Taxonomy of squat lobsters, p. 39-71. In Poore, G.C.B., Ahyong, S.T., and Taylor, J. (eds), The Biology of Squat Lobsters. CSIRO Publishing, Melbourne.
Marangon, S. and De Angeli, A. 2009. Exceptionally preserved specimens of Portunus monspeliensis (A. Milne Edwards, 1860) (Brachyura, Portunidae) from the Miocene of Sardinia (Italy). Atti della Società Italiana di Scienze Naturali e del Museo Civico di Storia Naturale di Milano, 150:3-12.
Marras, G. and Ventura, G. 1991. Crostacei decapodi del Miocene di Sassari (Sardegna nord occidentale). Bollettino della Società Sarda di Scienze Naturali, 28:105-119.
Martini, E. 1971. Standard Tertiary and Quartenary calcareous nannoplankton zonation, p. 739-785. In Farinacci, A. (ed), Proceedings of the 2nd Planktonic Conference. Tecnoscienza, Roma.
McLay, C.L. 2006. Retroplumidae (Crustacea, Decapoda) from the Indo-Malayan archipelago (Indonesia, Philippine) and the Melanesian arc islands (Solomon Islands, Fiji and New Caledonia), and paleogeographical comments. Mémoires du Muséum National d’Histoire Naturelle, 193:375-391.
Mikuž, V. 2003. Miocenske rakovice iz okolice Šentilja v Slovenskih Goricah. (The Miocene crabs from vicinity Šentilj in Slovenske Gorice, Slovenia). Razprave IV. razreda SAZU, 44:187-199.
Milne Edwards, A. 1860. Histoire des Crustacés podophthalmaires fossiles et Monographie des Décapodes Macroures de la famille des Thalassinens. Annales des Sciences Naturelles, 4e Série, 14:129-357.
Moore, W.E. 1957. Ecology of recent foraminifera in Northern Florida Keys. AAPG Bulletin, 41:727-741.
Müller, C. 1974. Nannoplankton aus dem Mittel-Miozän von Walbersdorf (Burgenland). Senckenbergiana Lethaea, 55:389-405.
Müller, P. 1984. Decapod Crustacea of the Badenian. Geologica Hungarica, Series Palaeontologica, 42:3-317.
Müller, P. 1993. Neogene Decapod Crustaceans from Catalonia. Scripta Musei Geologici Seminarii Barcinonensis, 225:1-39.
Müller, P. 1998a: Crustacea Decapoda, p. 1-48. In Flügel, H.W. (ed.), Catalogus Fossilium Austriae. Verlag der Österreichischen Akademie der Wissenschaften, Wien.
Müller, P. 1998b. Decapode Crustacea aus dem Karpat des Korneuburger Beckens (Unter-Miozän, Niederösterreich). Beiträge zur Paläontologie, 23:273-281.
Ng, P.K.L., Guinot, D., and Davie, P.J.F. 2008. Systema Brachyurorum: Part I. An annotated checklist of extant brachyuran crabs of the world. The Raffles Bulletin of Zoology, Supplement 17:1-286.
d'Orbigny, A. D. 1826. Tableau méthodique de la classe des Céphalopodes. Annales des Sciences Naturelles, 7:96-169, 245-314.
d'Orbigny, A.D. 1846. Die fossilen foraminiferen des tertiären Beckens von Wien. Foraminifères fossiles du bassin tertiaire de Vienne. Gide, Paris.
Ortmann, A.E. 1898. Gliedierfüssler: Arthropoda, p. 1057-1168. In Gerstäcker, A. and Ortmann, A.E. (eds.), Dr. H.G. Bronn’s Klassen und Ordnungen des Thier-Reichs, wissenschaftlich dargestellt in Wort und Bild. Bdand V, Abteilung. 2 (Crustacea), 2 Hälfte (Malacostraca). C.F. Winter, Leizig.
Papp, A. and Turnovsky, K. 1953. Die Entwicklung der Uvigerinen im Vindobon (Helvet und Torton) des Wiener Beckens. Jahrbuch der Geologischen Bundesanstalt, 96:117-142.
Perch-Nielsen, K. 1968. Der Feinbau und die Klassifikation der Coccolithen aus dem Maastrichtien von Danemark. Biologiske Skrifter. Kongelige Danske Videnskabernes Selskab, 16:1-96.
Petrović, M. 1980. Biostratigrafski značaj srednjemiocenskih foraminifera iz bušotina Tuzlanskog basena. Geološki anali Balkanskog Poluostrva, 43-44:155-203.
Petrović, V.M. 1959. Prilog poznavanju tortonsko-sarmatskih foraminifera iz Tuzle i uže okoline. Geološki anali Balkanskog Poluostrva, 26:195-202.
Pezelj, Đ., Mandić, O., and Ćorić, S. 2013. Paleoenvironmental dynamics in the southern Pannonian Basin during initial Middle Miocene marine flooding. Geologica Carpathica, 64:81-100.
Piller, W.E., Harzhauser, M., and Mandic, O. 2007. Miocene Central Paratethys stratigraphy - current status and future directions. Stratigraphy, 4:151-168.
Plotnick, R., Baumiller, T., and Wetmore, K.L. 1988. Fossilization potential of the mud crab, Panopeus (Brachyura: Xanthidae) and temporal variability in crustacean taphonomy. Palaeogeography, Palaeoclimatology, Palaeoecology, 63:27-43. https://doi.org/10.1016/0031-0182(88)90089-2
Rafinesque, C.S. 1815. Analyse de la Nature, ou Tableau de l’Univers et des Corps Organises. J. Barravecchia, Palermo.
Robins, C.M., Feldmann, R.M., and Schweitzer, C.E. 2013. Nine new genera and 24 new species of the Munidopsidae (Decapoda: Anomura: Galatheoidea) from the Jurassic Ernstbrunn Limestone of Austria, and notes on fossil munidopsid classification. Annalen des Naturhistorischen Museums in Wien, 115A:167-251.
Rögl, F. 1998. Palaeogeographic considerations for Mediterranean and Paratethys seaways (Oligocene to Miocene). Annalen des Naturhistorischen Museums in Wien, 99A:279-310.
Rögl, F. 1999. Mediterranean and Paratethys. Facts and hypotheses of an Oligocene to Miocene Paleogeography (short overview). Geologica Carpathica, 59:339-349.
Roth, P.H. 1970. Oligocene calcareous nannoplankton biostratigraphy. Eclogae Geologicae Helvetiae, 63:799-881.
Sacco, F.1894. I molluschi del terreni terziarii del Piemonte e dellaLiguria,16 (Cancellariidae) H.e A. Adams 1853. Bolettino dei Musei di Zoologiae di Anatomia Comparata della Reale Universita di Torino, 171: 68-70.
Samouelle, G. 1819. The Entomologist’s Useful Compedium; or an Introduction to the Knowledge of British Insects, Comprising the Best Means of Obtaining and Preserving Them, and a Description of the Apparatus Generally Used; Together with the Genera of Linné, and the Modern Method of Arranging the Classes Crustacea, Myriapoda, Spiders, Mites and Insects, from Their Affinities and Structure, According to the Views of Dr. Leach. Also an Explanation of the Terms Used in Entomology; a Calendar of the Times of Appearance and Usual Situations of Near 3,000 Species of British Insects; with Instructions for Collecting and Fitting Up Objects for the Microscope. Thomas Boys, London.
Schäfer, W. 1972. Ecology and Palaeoecology of Marine Environments (Aktuo-paläontologie nach Studien in der Nordsee, Translated by I. Oertel, Edited by G.Y. Craig). Oliver and Boyd, Edinbourgh.
Schiller, J. 1930. Coccolithineae, p.89-267. In Rabenhorst, L. (ed.), Kryptogamen-Flora von Deutschland, Österreich und der Schweiz. Akademische Verlagsgesellschaft, Leipzig.
Schweitzer, C.E., Feldmann, R.M., Garassino, A., Karasawa, H., and Schweigert, G. 2010. Systematic list of fossil decapod crustacean species. Crustaceana Monographs, 10:1-222. https://doi.org/10.1163/ej.9789004178915.i-222
Soklić, I., Atanacković, M., Jerković, L., and Petrović, M. 1980. Biostratigrafski aspekti za utvrđivanje geneze i starosti tuzlanskog sonog ležišta. Simpozij iz Regionalne Geologije i Paleontologije, 1980:307-316.
Spiridonov, V.A., Neretina, T.V., and Schepetov, D. 2014. Morphological characterization and molecular phylogeny of Portunoidea Rafinesque, 1815 (Crustacea Brachyura): Implications for understanding evolution of swimming capacity and revision of the family-level classification. Zoologischer Anzeiger, 253:404-429. https://doi.org/10.1016/j.jcz.2014.03.003
Stancu, I. and Andreescu, E. 1968. Fauna tortoniana din regiunea Rugi-Delinesti (Bazanul Caransebesului). Studii și Cercetări de Geologie, Geofizică, Geografie: Seria Geologie, 2:455-471.
Stempien, J.A. 2005. Brachyuran taphonomy in a modern tidal-flat environment: Preservation potential and anatomical bias. Palaios, 20:400-410.
Stevanović, P. and Eremija, M. 1960. Miocen Donje Tuzle. Geološki Anali Balkanskog Poluostrva, 27:45-102.
Stradner, H. 1963. New contributions to Mesozoic stratigraphy by means of nannofossils. Proceedings of the Sixth World Petroleum Congress, 1(4):167-183.
Theodoridis, S. 1984. Calcareous nannofossil biostratigraphy of the Miocene and revision of the helicoliths and discoasters. Utrecht Micropaleontological Bulletin, 32:1-271.
Vía Boada, L. 1959. Decápodos fósiles del Eoceno espanol. Boletin del Instituto Geológico y Minero de Espana, 70:1-72.
Vrabac, S. 1990. Asocijacije makrofaune i njihov stratigrafski značaj u krovinskim sedimentima ležišta soli Tuzlanskog basena. Zbornik XII. Kongresa Geologa SFRJ, 1:302-311.
Vrabac, S. and Ćorić, S., 2008. Revizija „karpata“ Tuzlanskog bazena sa osvrtom na stratigrafski položaj sone formacije. Geološki Glasnik, 37:71-81
Vrabac, S. and Mihajlović, Đ. 1990. Paleontološke i biostratigrafske odlike krovinskih sedimenata ležišta kamene soli Tušanj u Tuzli. Geološki anali Balkanskog Poluostrva, 53:297-303.
Vrabac, S., Ćorić, S., Ferhatbegović, Z., and Đulović, I. 2013. Stratigraphy of the middle Miocene salt deposit Tetima near Tuzla (The Central Paratethys, north Bosnia), p. 207. In Çağatay, N. and Zabci, C. (eds.), Regional Committee on Mediterranean Neogene Stratigraphy 14 Congress, Neogene to Quaternary Geological Evolution of Mediterranean, Paratethys and Black Sea. Book of Abstracts. Istanbul Technical University, Istanbul.
Wallich, G.C. 1877. Observations on the coccosphere. Annals and Magazine of Natural History, 19:342-350.
Weber, F. 1795. Nomenclator entomologicus secundum entomologiam systematicam ill. Fabricii, adjectis speciebus recens detectis et varietatibus. Bohn, C.E. Hamburg, London.
Whiteaves, J.F. 1874. On recent deep-sea dredging operations in the Gulf of St. Lawrence. American Journal of Science, Series 3, 7:210-219.
