

Carnian (Late Triassic) ostracods from the Sorgun Ophiolitic Mélange (Southern Turkey): Taxonomy, palaeoenvironment, and evidence of predation
Article number: 21.2.26A
https://doi.org/10.26879/852
Copyright Paleontological Society, July 2018
Author biographies
Plain-language and multi-lingual abstracts
PDF version
Submission: 18 January 2018. Acceptance: 28 June 2018
{flike id=2262}
ABSTRACT
The Mersin Ophiolitic Complex is situated in southern Turkey and contains the relics of the Palaeotethys and Neotethys Oceans and of the Huglu-Pindos basin. The succession exposed at the Tavusçayiri Block has been intensely studied but ostracods are still poorly documented. This paper illustrates and discusses the ostracods extracted from a radiolarian-rich limestone of the lower Tuvalian Spongotortilispinus moixi radiolarian Zone (Carnian, Late Triassic) intercalated with thick green tuffs. The moderately abundant assemblage (40 specimens) is composed of 29 species, 16 genera of seven families, including one new species: Bairdiacypris sorgunensis Forel sp. nov. This assemblage points to a moderately shallow subtidal environment submitted to short-lived environmental instabilities. In regards to the slightly older ostracods recently reported from the Tavusçayiri Block, the present record illustrates an upwards shallowing trend for the Carnian succession. All the taxa found are of typical Mesozoic-Cenozoic affinity, in contrast to the slightly older assemblage from the Julian of the Tavusçayiri Block (Kilek section), which also include Palaeozoic taxa. This different pattern could relate either to the disappearance of Palaeozoic taxa during the intervening interval or to their survival only in relatively deeper waters. Finally, we also report the oldest known record of a boring trace on the surface of an ostracod valve, probably of predatory origin. It represents the first evidence of predation on micro-benthos in the Triassic and is in line with several recent studies attesting the establishment of boring predation in the Late Triassic.
Marie-Béatrice Forel. CR2P, MNHN-CNRS-SU, Muséum national d’Histoire naturelle, 8 rue Buffon (CP38), 75005 Paris, France. marie-beatrice.forel@mnhn.fr
Péter Ozsvárt. MTA-MTM-ELTE, Research Group for Paleontology, P.O. Box 137, H 1431 Budapest, Hungary. ozsvart.peter@nhmus.hu
Patrice Moix. Rue de la Combe 55, 1969 Eison, Switzerland. patricemoix@gmail.com
Keywords: Ostracods; Carnian; southern Turkey; new species; paleoenvironment; predation
Final citation: Forel, Marie-Béatrice, Ozsvárt, Péter, and Moix, Patrice. 2018. Carnian (Late Triassic) ostracods from the Sorgun Ophiolitic Mélange (Southern Turkey): Taxonomy, palaeoenvironment, and evidence of predation. Palaeontologia Electronica 21.2.26A 1-23. https://doi.org/10.26879/852
palaeo-electronica.org/content/2018/2262-carnian-ostracods-from-turkey
Copyright: July 2018 Paleontology Society.
This is an open access article distributed under the terms of Attribution-NonCommercial-ShareAlike 4.0 International (CC BY-NC-SA 4.0), which permits users to copy and redistribute the material in any medium or format, provided it is not used for commercial purposes and the original author and source are credited, with indications if any changes are made.
creativecommons.org/licenses/by-nc-sa/4.0/
http://zoobank.org/BDBCC335-201C-4197-949A-81747E64B3B8
INTRODUCTION
The Triassic is a key period in the history of biodiversity and ecosystems because it is bounded by two mass extinctions, the end-Permian and the end-Triassic biotic crises, and it marks the transition from the Palaeozoic to the Meso-Cenozoic evolutionary faunas. Diversity patterns through space and time during this interval are pivotal to reconstruct the rate and mechanisms of biotic recovery and transition. In this discussion, ostracods, micro-crustaceans known from the Ordovician to present-day in all types of marine waters, recently provided challenging data which demonstrate the retention of primitive characteristics up to the Julian (Early Carnian), and thus reveal the protracted transitional interval following the end-Permian crisis in deep-waters (Forel et al., 2017). This stimulating discovery was made in the Mersin Ophiolitic Complex (MOC) in southern Turkey, which appears as a key area for the study of the Triassic biotic dynamics in marine environments.
The MOC is located in the South-Taurides belt in southern Turkey. It exhibits typical oceanic sediments of the Palaeotethys and Neotethys, as well as a typical series of the Huğlu-Pindos basin in the Late Cretaceous ophiolitic series and associated infra-ophiolitic mélange. Two major independent mélange units are recognized in the MOC: the Upper Cretaceous Sorgun Ophiolitic Mélange and the Middle to Upper Triassic Hacialani Mélange (Moix et al., 2011). One of the most important and well-developed tectonic blocks of the Sorgun Ophiolitic Mélange is the Tavusçayiri Block (Masset and Moix, 2004), which yielded the best-preserved and most diverse lower Tuvalian radiolarian fauna (Spongotortilispinus moixi radiolarian Zone) worldwide discovered in 2004 by Masset and Moix and partly described in a series of articles by Moix et al. (2007), Kozur et al. (2007a, 2007b, 2007c, 2009) and Ozsvárt et al. (2015, 2017a, 2017b). In this paper, we illustrate and discuss the silicified ostracods from the same radiolarian-rich layer (sample G11), which include 29 species and 16 genera. The environmental affinities of these taxa reflect deposition in moderately shallow subtidal environment with possible salinity and hydrodynamic fluctuations. These new conclusions suggest a shallowing upward trend at the Tavusçayiri Block, which is reported for the first time here. The ostracod assemblage from the sample G11 also differs significantly from the older assemblage of the Tavusçayiri Block (Tetraporobrachia haeckeli radiolarian Zone assemblage; Forel et al., 2017) by the absence of deep-water and typical Palaeozoic taxa. This pattern might either be due to the final disappearance of deep-sea Palaeozoic taxa during this interval, marking the end of the transitional interval following the end-Permian extinction, or to the survival of such taxa only in deeper water. We also report the occurrence of a drill hole on an ostracod valve, which is probably of predatory origin. This finding represents the oldest such record for ostracods worldwide. It further confirms the Mesozoic affinity of the present assemblage and ecosystem and the establishment of drilling predation in marine environments during the Late Triassic as indicated by other fossil groups (see Klompmaker et al., 2016 for details).
GEOLOGICAL SETTING, STRATIGRAPHY AND FACIES
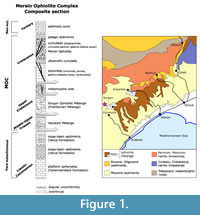 The studied area is located in the MOC, which belongs to the Taurides in Southern Turkey. It is situated northward from Erdemli city in Mersin area and is bounded by the villages of Gâvuruçtugu to the north and Sorgun to the south (Figure 1). The investigated section (coordinates 606066/4084886 in WGS84 system, zone 36S) belongs to the Tavusçayiri Block, which has been thoroughly described (Masset and Moix, 2004; Moix et al., 2007; Kozur et al., 2009; Ozsvárt et al., 2015). This succession occurs in the Upper Cretaceous Sorgun Ophiolitic Mélange (SOM), which is part of the MOC (Figure 1) occurring as blocks.
The studied area is located in the MOC, which belongs to the Taurides in Southern Turkey. It is situated northward from Erdemli city in Mersin area and is bounded by the villages of Gâvuruçtugu to the north and Sorgun to the south (Figure 1). The investigated section (coordinates 606066/4084886 in WGS84 system, zone 36S) belongs to the Tavusçayiri Block, which has been thoroughly described (Masset and Moix, 2004; Moix et al., 2007; Kozur et al., 2009; Ozsvárt et al., 2015). This succession occurs in the Upper Cretaceous Sorgun Ophiolitic Mélange (SOM), which is part of the MOC (Figure 1) occurring as blocks.
The base of the sequence is composed of breccia, which is unconformably overlain by polygenic conglomerates (Figure 2). The conglomerates are followed by black calciturbidites, which pass upward into a thin platform sequence, probably of early Carnian age (Moix et al., 2007; Kozur et al., 2009). The top of the platform represents a palaeo-relief locally rich in corals and filled with red pelagic limestones with abundant ammonoids indicative of the Trachyceras austriacum Zone of upper Julian, middle Carnian age (see Kozur et al., 2009 for details).  The red pelagic limestones (Hallstatt facies) are overlain by a volcanic unit (Pietra Verde-like tuffs) with interstratifications of pelagic and re-deposited limestones. They pass upward into a thick sequence of pelagic limestones intercalated with several debris-flows and calciturbidites. This sequence typically ends with a large breccia, but in a few places it is overlain by a well-developed “Ammonitico Rosso” facies (red calcareous facies rich in ammonoids) of middle Toarcian age. The uppermost part of the section is made of a breccia followed by radiolarites of late Dogger age.
The red pelagic limestones (Hallstatt facies) are overlain by a volcanic unit (Pietra Verde-like tuffs) with interstratifications of pelagic and re-deposited limestones. They pass upward into a thick sequence of pelagic limestones intercalated with several debris-flows and calciturbidites. This sequence typically ends with a large breccia, but in a few places it is overlain by a well-developed “Ammonitico Rosso” facies (red calcareous facies rich in ammonoids) of middle Toarcian age. The uppermost part of the section is made of a breccia followed by radiolarites of late Dogger age.
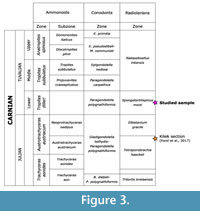 This succession corresponds to the Huğlu-type sequence developed farther to the west, which was first described by Özgül (1976) in the Bozkır Units and by Monod (1977) in the Beysehir-Hoyran Nappes. The stratigraphy of the Huğlu Unit was revised by Kozur (1997) and regional correlations have been proposed by Moix et al. (2013). All the ostracods reported here come from one sample (G11, Figure 2), which has been previously analysed for radiolarians (Kozur et al., 2009). The sampled horizon is located in a limestone unit, intercalated with thick green tuffs, and belongs to the lower Tuvalian Spongotortilispinus moixi radiolarian Zone (Moix et al., 2007; Figure 3). Although no microfacies information are available for this sample, other limestones intercalated within tuffites are radioalarian wackestones with abundant spicules, brachiopods, ammonoids, gastropods, ostracods, foraminifers, conodonts and pelagic crinoids.
This succession corresponds to the Huğlu-type sequence developed farther to the west, which was first described by Özgül (1976) in the Bozkır Units and by Monod (1977) in the Beysehir-Hoyran Nappes. The stratigraphy of the Huğlu Unit was revised by Kozur (1997) and regional correlations have been proposed by Moix et al. (2013). All the ostracods reported here come from one sample (G11, Figure 2), which has been previously analysed for radiolarians (Kozur et al., 2009). The sampled horizon is located in a limestone unit, intercalated with thick green tuffs, and belongs to the lower Tuvalian Spongotortilispinus moixi radiolarian Zone (Moix et al., 2007; Figure 3). Although no microfacies information are available for this sample, other limestones intercalated within tuffites are radioalarian wackestones with abundant spicules, brachiopods, ammonoids, gastropods, ostracods, foraminifers, conodonts and pelagic crinoids.
MATERIAL AND METHODS
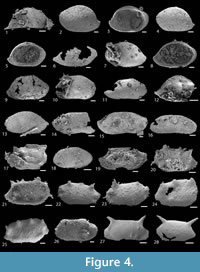 The sample G11 was processed using dilute acetic acid for radiolarian extraction (Masset and Moix, 2004; Moix et al., 2007; Kozur et al., 2007a, 2007b, 2007c) and yielded moderately abundant silicified ostracods characterized by their delicate lace-like preservation. This assemblage is composed of 29 species belonging to 16 genera and seven families (Table 1). Due to the relatively poor preservation and low quantity of the material, clear identification of diagnostic characters was limited so most species are kept in open nomenclature, but all are illustrated for later comparisons (Figure 4-Figure 5). For the sake of brevity, the following systematic palaeontology chapter only includes the full description of the new species Bairdiacypris sorgunensis Forel sp. nov., as well as the synonymy and remarks on previously known species. Following Maddocks (2015), the degree of slope of the antero-dorsal and postero-dorsal margins are measured on external lateral views of the carapace or valve, with 0 being horizontal. The size convention is as follows: <400 μm very small, 400-500 μm small, 500-700 μm medium, 700-1000 μm large, >1000 μm very large.
The sample G11 was processed using dilute acetic acid for radiolarian extraction (Masset and Moix, 2004; Moix et al., 2007; Kozur et al., 2007a, 2007b, 2007c) and yielded moderately abundant silicified ostracods characterized by their delicate lace-like preservation. This assemblage is composed of 29 species belonging to 16 genera and seven families (Table 1). Due to the relatively poor preservation and low quantity of the material, clear identification of diagnostic characters was limited so most species are kept in open nomenclature, but all are illustrated for later comparisons (Figure 4-Figure 5). For the sake of brevity, the following systematic palaeontology chapter only includes the full description of the new species Bairdiacypris sorgunensis Forel sp. nov., as well as the synonymy and remarks on previously known species. Following Maddocks (2015), the degree of slope of the antero-dorsal and postero-dorsal margins are measured on external lateral views of the carapace or valve, with 0 being horizontal. The size convention is as follows: <400 μm very small, 400-500 μm small, 500-700 μm medium, 700-1000 μm large, >1000 μm very large.
The illustrated specimens and types are deposited in the collections of the Hungarian Natural History Museum, Budapest, under the numbers INV 2018.1 to INV 2018.31.
Abbreviations. L, length; H, height; DB, dorsal border; VB, ventral border; ADB, antero-dorsal border; AVB, antero-ventral border; PDB, postero-dorsal border; PVB, postero-ventral border.
SYSTEMATIC PALAEONTOLOGY
The present taxonomy follows the classifications of Horne et al. (2002), Moore (1961) and Becker (2002). The taxonomy of Cytheroidea is based on the emendations of Whatley and Boomer (2000) and that of Bythocytheridae follows Schornikov (1990). Species left in open nomenclature, except for exceptional genera or species, are omitted.
Class OSTRACODA Latreille, 1806
Subclass PODOCOPA Müller, 1894
Order PODOCOPIDA Müller, 1894
Superfamily BAIRDIOIDEA Sars, 1887
Family BAIRDIIDAE Sars, 1887
Genus BAIRDIA McCoy, 1844
Type species. Bairdia curta McCoy, 1844, by subsequent designation of Ulrich and Bassler (1923, p. 320).
Bairdia sp. 4
Figure 4.7, 4.8
Material examined. One complete left valve (INV 2018.1).
Dimensions. L = 811 μm, H = 491 μm.
Range. Spongotortilispinus moixi radiolarian Zone, lower Tuvalian, Upper Carnian (this work), Late Triassic.
Occurrence. Sample G11, Killik Formation, Tavusçayiri Block, Sorgun Ophiolitic Mélange, southern Turkey (606066/4084886), Huğlu Tuffite (this work).
Remarks. Bairdia sp. 4 is very close to Bairdia sp. 4 in Forel et al., 2017 from the Tetraporobrachia haeckeli radiolarian Zone, Julian, Middle Carnian, Late Triassic of the Kilek section, Tavusçayiri Block, Sorgun Ophiolitic Mélange (Forel et al., 2017). Both species are only known by an isolated left valve with straight hinge line underlain by two pronounced lists, a calcified inner lamella moderately developed at AB and a raised posterior end. However, Bairdia sp. 4 differs from Bairdia sp. 4 in Forel et al., 2017 by its more elongate posterior margin, longer hinge line, poorly developed selvage along ventral margin and moderately developed calcified inner lamella at AB bordered by a thin laterally compressed area. Although kept in open nomenclature, this species is of high importance as it bears the oldest known drill hole on an ostracod valve, as discussed below.
Genus BAIRDIACYPRIS Bradfield, 1935
Type species. Bairdiacypris deloi Bradfield, 1935 by original designation.
Bairdiacypris sorgunensis Forel sp. nov.
Figure 4.14, 4.15
zoobank.org/ED721A32-3CFE-40E1-88CE-DF2AECE78AA9
v. 2017 Bairdiacypris aequisymmetrica Mette, Honigstein and Crasquin; Forel, Tekin, Okuyucu, Bedi, Tuncer and Crasquin, p. 11, figs. 10R-T.
Etymology. The species name refers to the Sorgun Ophiolitic Mélange from which the material was derived.
Type material. Holotype: one right valve, adult, from the Kilek section, P6M4075 (Figure 4.15, from Forel et al., 2017); Paratype 1: one right valve, stage A-2, from the Kilek section, P6M4073 (figure 10R in Forel et al., 2017); Paratype 2: one right valve, stage A-3, from the Kilek section, P6M4074 (figure 10S in Forel et al., 2017).
Additional material examined. One broken right valve, INV 2018.2 (Figure 4.14); 4 complete valves from the Kilek section (Forel et al., 2017).
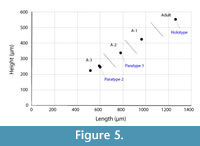 Dimensions. L = 517-1259 μm, H = 223-558 μm, H/L = 0.40-0.44 (Figure 5, all specimens from Kilek section; Forel et al., 2017).
Dimensions. L = 517-1259 μm, H = 223-558 μm, H/L = 0.40-0.44 (Figure 5, all specimens from Kilek section; Forel et al., 2017).
Diagnosis. A Bairdiacypris species with an elongate lateral outline, sub-symmetrical anterior and posterior margins and ventral margin marked by a thin lip.
Description. Carapace very large, elongate, sub-rectangular in lateral view with Hmax around mid-L and Lmax at/or slightly above mid-H; dorsal margin long with distinct PDB, DB and ADB separated by distinct antero-dorsal and postero-dorsal corners; DB long (about 33-40% of Lmax) and straight, ADB straight and sloping toward the anterior margin with an angle of about 25°, PDB shorter and concave upward; anterior margin with relatively large radius of convexity, maximum located above mid-H, AVB broadly rounded and strongly inclined toward AB; ventral margin long (ranging from 49 to 53% of Lmax) with concavity located in front of mid-L, marked by a thin lip running all along ventral length; posterior margin nearly symmetrical to anterior margin, but more sharply pointed upward and with a smaller radius of concavity located at the same level than anterior one; overlap not observed; surface smooth.
Range. Tetraporobrachia haeckeli radiolarian Zone, Julian, Middle Carnian (Forel et al., 2017); Spongotortilispinus moixi radiolarian Zone, lower Tuvalian, Upper Carnian (this work), Late Triassic.
Occurrence. Kilek section, Tavusçayiri Block, Sorgun Ophiolitic Mélange, southern Turkey, Huğlu Tuffite (Forel et al., 2017); sample G11, Killik Formation, Tavusçayiri Block, Sorgun Ophiolitic Mélange, southern Turkey (606066/4084886), Huğlu Tuffite (this work).
Remarks. Bairdiacypris sorgunensis Forel sp. nov. is related to Bairdiacypris aequisymmetrica Mette, Honigstein and Crasquin, 2014 from the Middle Anisian (Reifling Formation) of the Northern Calcareous Alps (Austria; Mette et al., 2014) due to its elongate morphology in lateral view and its sub-symmetrical anterior and posterior margins. However, Bairdiacypris aequisymmetrica Mette, Honigstein and Crasquin, 2014, has less tapered and pointed anterior and posterior ends and lacks the thin lip along the ventral margin. Bairdiacypris sorgunensis Forel sp. nov. also differs from Bairdia anisica Kozur, 1970a from the Upper Anisian of Germany (Kozur, 1970a) in being more elongate with a longer and lower dorsal border, a higher anterior margin with shorter antero-ventral border and in possessing a thin lip along ventral margin. It is further different from Bairdiacypris anisica Kozur, 1971a, from the Upper Anisian of Hungary (Kozur, 1971a) by its more pronounced medio-ventral concavity, more rectangular outline in lateral view, more pointed anterior and posterior ends and thin lip along ventral margin. Six specimens from the Julian of the Kilek section (Forel et al., 2017) were erroneously attributed to Bairdiacypris aequisymmetrica Mette, Honigstein and Crasquin, 2014, as shown in the synonym list. They plot into at least four ontogenetic stages as shown in Figure 5, from A-3 to Adult. The specimen extracted from the sample G11 is 350 μm in height, which corresponds to the upper size range of the A-2 stage or to the lower size range of the A-1 stage.
Genus ISOBYTHOCYPRIS Apostolescu, 1959
Type species. Isobythocypris unispinata Apostolescu, 1959, by original designation.
Isobythocypris ? cf. postera (Herrig, 1979a) in Forel et al., 2017
Figure 4.18
v. 2017 Isobythocypris? cf. postera (Herrig, 1979a); Forel, Tekin, Okuyucu, Bedi, Tuncer and Crasquin, p. 17, 19, figs. 10R-T.
Material examined. One complete right valve (INV 2018.3).
Dimensions. L = 511 μm, H = 237 μm.
Range. Tetraporobrachia haeckeli radiolarian Zone, Julian, Middle Carnian (Forel et al., 2017); Spongotortilispinus moixi radiolarian Zone, lower Tuvalian, Upper Carnian (this work), Late Triassic.
Occurrence. Kilek section, Tavusçayiri Block, Sorgun Ophiolitic Mélange, southern Turkey, Huğlu Tuffite (Forel et al., 2017); sample G11, Killik Formation, Tavusçayiri Block, Sorgun Ophiolitic Mélange, southern Turkey (606066/4084886), Huğlu Tuffite (this work).
Remarks. This species is closely related to Isobythocypris? postera (Herrig, 1979) described from the Lower Jurassic of Germany (Herrig, 1979a), which has been documented from the Hettangian to the Pliensbachian of Germany, France and Ireland (Apostolescu, 1959; Herrig, 1979a, b; Rutherford and Ainsworth, 1989; Harloff, 1993; Harloff and Jäger, 1994; Monostori, 1996). However, we lack complete material to clearly attribute our material to Isobythocypris? postera (Herrig, 1979a) or to define precisely the diagnostic characters of the potential new species.
Genus MIRABAIRDIA Kollmann, 1963
Type Species. Mirabairdia pernodosa Kollmann, 1963 by original designation.
Mirabairdia longispinosa Kristan-Tollmann, 1978
Figure 4.20, 4.21
v. 1978 Mirabairdia longispinosa Kristan-Tollmann, p. 94-96, pl. 3, fig. 1-3, pl. 7, fig. 5.
Material examined. One complete left valve (INV 2018.4).
Dimensions. L = 640 μm, H = 350 μm (not considering spines).
Range. Cordovelian, Early Carnian, (Kristan-Tollmann, 1978); Spongotortilispinus moixi radiolarian Zone, lower Tuvalian, Upper Carnian (this work), Late Triassic.
Occurrence. Cassian beds, South Tyrol, Italy (Kristan-Tollmann, 1978); sample G11, Killik Formation, Tavusçayiri Block, Sorgun Ophiolitic Mélange, southern Turkey (606066/4084886), Huğlu Tuffite (this work).
Remarks. The present record is the youngest occurrence of Mirabairdia longispinosa Kristan-Tollmann, 1978, which was previously only known from the base of the Carnian (Cordovelian; Kristan-Tollmann, 1978). It is also the first record of this species outside Italy.
Genus PETASOBAIRDIA Chen in Chen and Shi, 1982
Type species. Petasobairdia bicornuta Chen in Chen and Shi (1982) by original designation.
Remarks. The species Ceratobairdia crassispinosa Monostori and Tóth, 2014, has been reported from the Ladinian, Middle Triassic of the Transdanubian Central Range, Hungary (Monostori and Tóth, 2014). However, the type material of Ceratobairdia crassispinosa Monostori and Tóth, 2014, does not possess the ventrolateral alate ridge and flat ventral surface, which are diagnostic characters of the genus Ceratobairdia Sohn, 1954. Ceratobairdia crassispinosa Monostori and Tóth, 2014, is also characterized by its anterior and posterior margins, which are strongly compressed laterally. For these reasons, this species is removed from Ceratobairdia and assigned to Petasobairdia, which shows laterally compressed anterior and posterior margins and ridge, nodes, denticulation or horn-like spines at the dorsal margin of the LV.
Petasobairdia longispinosa (Kozur, 1971b)
Figure 4.27, 4.28, Figure 6.1
v. 1971b Ceratobairdia longispinosa Kozur; 4; pl. 1, fig. e.
Non 1984 Ceratobairdia longispinosa Kozur; Salaj and Jendrejáková, pl. 1, fig. 12, pl. 3, figs. 1-4.
 cf. 1996 Ceratobairdia longispinosa Kozur; Crasquin-Soleau and Grădinaru, pl. 5, fig. 6.
cf. 1996 Ceratobairdia longispinosa Kozur; Crasquin-Soleau and Grădinaru, pl. 5, fig. 6.
Non 2013 Ceratobairdia longispinosa Kozur; Sebe, Crasquin and Grădinaru, p. 520; pl. 3, fig. 11.
v. 2014 Petasobairdia longispinosa (Kozur); Mette, Honigstein and Crasquin, p. 81, pl. 3, figs. 7-9.
v. 2017 Petasobairdia longispinosa (Kozur); Forel, Tekin, Okuyucu, Bedi, Tuncer and Crasquin, figs. 12L-M.
Material examined. Five complete left valves (INV 2018.5.1-INV 2018.5.5) and one complete right valve (INV 2018.5.6).
Dimensions. L = 466-883 μm, H = 247-400 μm (this work only, not considering the spines).
Range. Upper Anisian, Illyrian, Middle Triassic (Kozur, 1971b); Middle Anisian, Middle Triassic (Mette et al., 2014); Tetraporobrachia haeckeli radiolarian Zone, Julian, Middle Carnian, Late Triassic (Forel et al., 2017); Spongotortilispinus moixi radiolarian Zone, lower Tuvalian, Upper Carnian, Late Triassic (this work).
Occurrence. Slovakia (Kozur, 1971b); Reifling Formation, Northern Calcareous Alps, Austria (Mette et al., 2014); Kilek section, Tavusçayiri Block, Sorgun Ophiolitic Mélange, southern Turkey, Huğlu Tuffite (Forel et al., 2017); sample G11, Killik Formation, Tavusçayiri Block, Sorgun Ophiolitic Mélange, southern Turkey (606066/4084886), Huğlu Tuffite (this work).
Remarks. Based on the illustrations in Salaj and Jendrejáková (1984), their specimens are re-attributed to the genus Mirabairdia. Similarly, the material from the lower Anisian of Dobrogea, Romania (Sebe et al., 2013) is rejected from Petasobairdia longispinosa (Kozur 1971b) because it lacks the dorsal spines or even scars of broken spines. Petasobairdia longispinosa (Kozur, 1971b), therefore, ranges from the Middle Anisian to the lower Tuvalian, Upper Carnian and this species is not recorded from the Middle Triassic of North Dobrogea. In the current state of knowledge, this species is restricted to the northern margins of the Meliata and Küre oceans (see for example Stampfli, 2000) during the Anisian and occurs at their southern margins in the Carnian. Petasobairdia crassispinosa (Monostori and Tóth, 2014) from the Ladinian of Hungary (Monostori and Tóth, 2014) differs from Petasobairdia longispinosa (Kozur, 1971b) only by the basal morphology of their dorsal spines, which are broader compared to the type material of Petasobairdia longispinosa. Several specimens of Petasobairdia longispinosa (Kozur, 1971b) possess this type of spine morphology so that this character is not sufficient to establish a new species. The two specimens of Petasobairdia crassispinosa illustrated by Monostori and Tóth (2013, 2014) are rather characterized by a relatively short and high carapace and strongly compressed anterior and posterior margins. These characters are more pronounced in the material of Monostori and Tóth (2013, 2014) compared to the type material of Petasobairdia longispinosa (Kozur, 1971b). But the two specimens of Monostori and Tóth (2013, 2014) are not sufficient to distinguish possible transitional forms between the two morphologies. Until more material can be studied, we keep these specimens in different species.
Genus PTYCHOBAIRDIA Kollmann, 1960
Type species. Ptychobairdia kuepperi Kollmann, 1960 by original designation.
Ptychobairdia kristanae Kollmann, 1960
Figure 6.2
v. 1960 Ptychobairdia kristanae Kollmann; p. 99, 100, pl. 25, figs. 6-9.
v. 1960 Ptychobairdia medwenitschi Kollmann; p. 100, pl. 26, figs. 1-11.
1963 Ptychobairdia kristanae Kollmann; Kollmann, p. 181.
Material examined. One complete right valve (INV 2018.6).
Dimensions. L = 670 μm, H = 490 μm.
Range. Late Triassic (Kollmann, 1960, 1963); Spongotortilispinus moixi radiolarian Zone, lower Tuvalian, Upper Carnian, Late Triassic (this work).
Occurrence. Austria (Kollmann, 1960, 1963); sample G11, Killik Formation, Tavusçayiri Block, Sorgun Ophiolitic Mélange, southern Turkey (606066/4084886), Huğlu Tuffite (this work).
Superfamily CYTHEROIDEA Baird, 1850
Family CYTHERURIDAE Müller, 1894
Subfamily CYTHERURINAE Müller, 1894
Genus JUDAHELLA Sohn, 1968
Type species. Judahella tsorfatia Sohn, 1968, by original designation.
Judahella fortenodosa (Urlichs, 1972)
Figure 6.7
v. 1972 Monoceratina fortenodosa Urlichs; p. 676, pl. 1, figs. 6-8.
v. 1980 Monoceratina fortenodosa Urlichs; Kristan-Tollmann, Tollmann and Hamedani, p. 187, pl. 8, fig. 10.
v. 1988 Triceratina fortenodosa (Urlichs); Kristan-Tollmann, figs. 6, 10.
v. 1991 Triceratina fortenodosa (Urlichs); Kristan-Tollmann, Haas and Kovács, pl. 8, figs. 1, 2.
v. 1999 Triceratina fortenodosa (Urlichs); Mette and Mohtat-Aghai, pl. 6, fig. 7.
v. 2012 Triceratina fortenodosa (Urlichs); Mette, Elsler and Korte, p. 70.
2013 Tuberoceratina fortenodosa (Urlichs); Hillebrandt, Krystyn, Kürschner, Bonis, Ruhl, Richoz, Schobben, Urlichs, Kment, McRoberts, Simms, and Tomãsových, p. 70.
v. 2014 Judahella fortenodosa (Urlichs); Monostori and Tóth, p. 29, pl. 3, figs. 3, 4.
Material examined. One complete right valve (INV 2018.7).
Dimensions. L = 904 μm, H = 365 μm.
Range. Norian-Rhaetian, Late Triassic (Urlichs, 1972); Rhaetian, Late Triassic (Kristan-Tollmann, 1988; Kristan-Tollmann et al., 1980); Norian-Rhaetian, Late Triassic (Kristan-Tollmann, et al. 1991); Rhaetian, Late Triassic (Mette and Mohtat-Aghai, 1999); Rhaetian, Late Triassic (Mette et al., 2012); Rhaetian, Late Triassic (Hillebrandt et al., 2013); Ladinian, Middle Triassic (Monostori and Tóth, 2014); Spongotortilispinus moixi radiolarian Zone, lower Tuvalian, Upper Carnian, Late Triassic (this work).
Occurrence. Kössen Beds, Austria (Urlichs, 1972); Iran (Kristan-Tollmann, 1988; Kristan-Tollmann et al., 1980); Tyrol (Kristan-Tollmann, et al. 1991); Kössen Formation, Northern Calcareous Alps, Austria (Mette and Mohtat-Aghai, 1999); Kössen Formation, Eiberg Member, Northern Calcareous Alps, Austria (Mette et al., 2012); Kössen Formation, Eiberg Member, Kuhjoch section, Austria (Hillebrandt et al., 2013); Hungary (Monostori and Tóth, 2014); sample G11, Killik Formation, Tavusçayiri Block, Sorgun Ophiolitic Mélange, southern Turkey (606066/4084886), Huğlu Tuffite (this work).
Remarks. As shown by the synonym list, this species has a complex taxonomic history in regards to its generic placement. It was originally attributed to the genus Monoceratina Roth, 1928, by Urlichs (1972) and later alternatively re-attributed to the genera Triceratina Upson, 1933, and Tuberoceratina Gründel and Kozur, 1972. Recently, Whatley and Boomer (2000, p. 144) reviewed the systematics of the early Cytheruridae and concluded that this species is a member of the genus Judahella Sohn, 1968. The present record is the oldest occurrence of Judahella fortenodosa (Urlichs, 1972).
Order PLATYCOPIDA Sars, 1866
Subclass MYODOCOPA Sars, 1866
Order MYODOCOPIDA Sars, 1866
Superfamily POLYCOPOIDEA Sars, 1866
Family POLYCOPIDAE Sars, 1866Genus POLYCOPE Sars, 1866
Type species. Polycope orbicularis Sars, 1866 by original designation.
Polycope cincinnata Apostolescu, 1959
Figure 6.11
v. 1970b Polycopsis hungarica Kozur; p. 407, pl. 3, fig. 12.
v. 1971 Polycopsis cincinnata (Apostolescu); Bunza and Kozur, p. 15, pl. 2, figs. 10, 11.
v. 1972 Polycope cincinnata Apostolescu; Urlichs, p. 696, pl. 4, fig. 13.
v. 2013 Polycope cincinnata Apostolescu; Monostori and Tóth, p. 307, pl. 1, fig. 3.
v. 2014 Polycope cincinnata Apostolescu; Mette, Honigstein and Crasquin, pl. 1, fig. 2.
v. 2017 Polycope cincinnata Apostolescu; Forel, Tekin, Okuyucu, Bedi, Tuncer and Crasquin, p. 36, 37, fig. 24E, F.
Material examined. One complete right valve (INV 2018.8).
Dimensions. L = 460 μm, H = 470 μm.
Range. Middle Anisian, Middle Triassic (Kozur, 1970b); Anisian, Middle Triassic (Bunza and Kozur, 1971); upper Norian, Upper Triassic (Urlichs, 1972); Ladinian, Middle Triassic (Monostori and Tóth, 2013); Middle Anisian, Middle Triassic (Mette et al., 2014); Tetraporobrachia haeckeli radiolarian Zone, Julian, Middle Carnian, Late Triassic (Forel et al., 2017); Spongotortilispinus moixi radiolarian Zone, lower Tuvalian, Upper Carnian, Late Triassic (this work).
Occurrence. Felsöörs, Hungary (Kozur, 1970b); Austria (Bunza and Kozur, 1971); Kössen, Austria (Urlichs, 1972); Litér quarry, Hungary (Monostori and Tóth, 2013); Riefling Formation, Austria (Mette et al., 2014); Kilek section, Tavusçayiri Block, Sorgun Ophiolitic Mélange, southern Turkey, Huğlu Tuffite (Forel et al., 2017); sample G11, Killik Formation, Tavusçayiri Block, Sorgun Ophiolitic Mélange, southern Turkey (606066/4084886), Huğlu Tuffite (this work).
Remarks. The species Polycope cincinnata Apostolescu, 1959 is common and widespread in the Jurassic. For the sake of brevity, we follow Forel et al. (2017) and do not report all of its Jurassic occurrences but rather focus on its Triassic roots.
OSTRACOD DIVERSITY AND STRATIGRAPHIC RANGE
The sample G11 was processed for radiolarian analysis (Moix et al., 2007; Kozur et al., 2007a, 2007b, 2007c, 2009; Ozsvárt et al., 2015; 2017a, 2017b) and provided a moderately abundant assemblage of silicified ostracods belonging to 29 species, 16 genera and seven families (Table 1). Bairdiidae strongly dominate the assemblage with 23 species and 10 genera (Acratia, Bairdia, Bairdiacypris, Dicerobairdia, Isobythocypris, Medwenitschia, Mirabairdia, Nodobairdia, Petasobairdia, Ptychobairdia). Each of the six remaining families is represented by one monotypic genus: Bythocytheridae (genus Nemoceratina), Cavellinidae (genus Bektasia), Cytherellidae (genus Leviella), Cytheruridae (genus Judahella), Healdiidae (genus Ogmoconcha) and Polycopidae (genus Polycope). The genus Microcheilinella is mentioned by Masset and Moix (2004) as part of the assemblage in G11, without illustration, and is not found here.
The present assemblage records the oldest known occurrence of the genus Medwenitschia that was previously documented from the Upper Norian (Kristan-Tollmann, 1971) to the Rhaetian (Kollmann, 1960, 1963). It is additionally the first Carnian record of Nemoceratina (Nemoceratina), which was reported from the Anisian-Ladinian and Norian (Kozur, 1970b; Kozur and Mostler, 1971; Bunza and Kozur, 1971). Two of the species reported here were only known from the ostracod assemblage extracted from the Kilek section of Julian age, Tetraporobrachia haeckeli Zone, which is also located in the Tavusçayiri Block: Bairdiacypris sorgunensis Forel sp. nov. (Figure 4.11) and Isobythocypris ? cf. postera (Herrig, 1979a) in Forel et al., 2017 (Figure 4.14). Five other species were previously known from the Triassic interval and outside Turkey: Mirabairdia longispinosa, Petasobairdia longispinosa, Ptychobairdia kristanae, Judahella fortenodosa, Polycope cincinnata. The present assemblage records the youngest occurrences of Petasobardia longspinosa and Mirabairdia longispinosa, as well as the oldest occurrence of Ptychobairdia kristanae. Although Judahella fortenodosa is known from the Ladinian through the Rhaetian, we report here the first occurrence of this species in the Carnian and in Turkey.
PALAEOENVIRONMENTAL SETTING
In order to evaluate the environmental conditions and water depth at which the present ostracod assemblage developed, we use the model established by Lethiers and Raymond (1991), based on the proportions of the following ostracod groups: (1) deep-sea taxa developed in cold-water, low-energy environments, which differ morphologically from the neritic forms by their thin tests and/or delicate or long spines, (2) Bairdia sensu lato that are inhabitants of euhaline thermospheric waters, (3) other neritic species that are associated with more nearshore conditions. When deep-sea species are more than 50% of the assemblage, the association is very probably reflecting deposition in the bathyal to abyssal zone and Bairdia are generally less than 15% of the species. Between 50 and 12% of deep-sea species, the assemblage suggests the outer shelf-upper slope zone, where Bairdia are between 15 and 22% of the species. When the deep-sea taxa comprise less than 12% of the species, Bairdia are between 15 and 50% and suggestive of a marine, relatively warm and oxygenated platform with euhaline conditions. Where Bairdia are from 0 to 15% of the species, the environment is likely more euryhaline and more or less oxygenated (Lethiers and Raymond, 1991). It is worth noting that this model was established using Devonian assemblages, corresponding to different climatic conditions and different ostracod assemblages from the Triassic period. However, increasing data document the persistence of Palaeozoic taxa up to the end of the Middle Triassic (e.g., Crasquin-Soleau and Grădinaru, 1996; Forel and Crasquin, 2011; Sebe et al., 2013) and even to the Late Triassic (Forel et al., 2017). These residual taxa include Palaeocopida and typical deep-sea taxa originally described from most of the Palaeozoic and used for the establishment of the Lethiers and Raymond (1991) model. Their subsistence through the end-Permian extinction and up to the Late Triassic is a strong argument for the stability of the deep-water conditions over this interval, in spite of the drastic climatic modifications. However, the precise conditions, which allowed for such survival, are still obscure. As a consequence, we chose to use the Lethiers and Raymond (1991) model, supplemented with information on the typical shallow-water Triassic taxa available from previous works.
In the present contribution, deep-sea species are not abundant: Nemoceratina (Nemoceratina) sp. which is the only representative of the typically deep-sea Bythoceratidae, Acratia sp. and Bairdia sp. 8 which possess characteristic spinose posterior ends. The Bairdia sensu lato group includes all smooth and thin-shelled Bairdiidae: Bairdia sp. 1-7, 9, all Bairdiacypris and Isobythocypris species, in total 11 species. The group of other neritic taxa is composed of 15 species: all thick-shelled sculptured Bairdiidae, Judahella, Ogmoconcha, Bektasia, Leviella and Polycope.
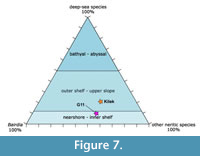 As a first observation, the extreme abundance of radiolarian specimens indicates blooming in the pelagic environment. The studied limestone level was originally described as a radiolarian micritic limestone intercalated within tuffites. The tuffites represent re-sedimentation, which might be consistent with the occurrence of Bouma sequences. However, until now locally, only calciturbidites (calcarenites) have been observed on the field. Regarding ostracods taxa, deep-sea taxa are 10% of the species, whereas Bairdia lato sensu and other neritic species are, respectively, 38% and 52% of the assemblage extracted from the sample G11 (Figure 7). The presence of shallow-water taxa such as the genus Bektasia, which has been described as tolerating important salinity variations, and the species Judahella fortenodosa, (Kristan-Tollmann and Hamedani, 1973; Kristan Tollmann et al., 1980), indicates a rather moderately shallow subtidal environment submitted to environmental variations. The occurrence of strong-shelled sculptured Bairdiidae further indicates the influence of hydrodynamism. In modern environments, the genus Polycope is interpreted as an opportunistic group indicating organic-rich, fine-grained sediment and high productivity (Karanovic and Brandão, 2016). Although not abundant in the studied sample, the occurrence of this genus tends to indicate that it was deposited under the influence of the open ocean with high productivity. A previous estimate indicated a water-depth of about 150-200 m for the studied assemblage (Masset and Moix, 2004; Moix et al., 2007). Based on the observations and interpretations presented here, this estimate might be slightly high.
As a first observation, the extreme abundance of radiolarian specimens indicates blooming in the pelagic environment. The studied limestone level was originally described as a radiolarian micritic limestone intercalated within tuffites. The tuffites represent re-sedimentation, which might be consistent with the occurrence of Bouma sequences. However, until now locally, only calciturbidites (calcarenites) have been observed on the field. Regarding ostracods taxa, deep-sea taxa are 10% of the species, whereas Bairdia lato sensu and other neritic species are, respectively, 38% and 52% of the assemblage extracted from the sample G11 (Figure 7). The presence of shallow-water taxa such as the genus Bektasia, which has been described as tolerating important salinity variations, and the species Judahella fortenodosa, (Kristan-Tollmann and Hamedani, 1973; Kristan Tollmann et al., 1980), indicates a rather moderately shallow subtidal environment submitted to environmental variations. The occurrence of strong-shelled sculptured Bairdiidae further indicates the influence of hydrodynamism. In modern environments, the genus Polycope is interpreted as an opportunistic group indicating organic-rich, fine-grained sediment and high productivity (Karanovic and Brandão, 2016). Although not abundant in the studied sample, the occurrence of this genus tends to indicate that it was deposited under the influence of the open ocean with high productivity. A previous estimate indicated a water-depth of about 150-200 m for the studied assemblage (Masset and Moix, 2004; Moix et al., 2007). Based on the observations and interpretations presented here, this estimate might be slightly high.
THE OLDEST DRILLING TRACE ON OSTRACODS
Ostracods constitute a minor component of the diet of diverse organisms, such as bivalves, gastropods, echinoids, turtles, amphibians, fishes, annelids and other crustaceans including other ostracods (e.g., Lowndes, 1930; Harding, 1962; Reyment, 1966; Robertson, 1988; Penchaszadeh et al., 2004; Costa et al., 2006; Ghioca-Robrecht and Smith, 2008; Leal, 2008; Alcalde et al., 2010; Rossi et al., 2011; Vandekerkhove et al., 2012). Identifiable evidence for the predatory pressure imposed on ostracods in the fossil record is associated with drilling gastropods of the families Naticidae and Muricidae (e.g., Maddocks, 1988; Reyment and Elewa, 2002). The oldest record of such activity on ostracods is of Early Albian, Early Cretaceous age (Maddocks, 1988). Because of the small size of ostracods, they provide an important food source for juvenile gastropods and an alternative nutritive resource when regular prey are lacking (e.g., Reyment, 1967; Maddocks, 1988). Predation on ostracods is most of the time disregarded, but these characteristics make them potential indicators of the dynamics of juvenile predators and of periods of prey impoverishment.
Among the specimens recovered from the sample G11, one complete left valve of the species Bairdia sp. 4 displays a drill hole located in the postero-ventral portion of the lateral surface (Figure 4.5). The drill hole is circular in outline and represents the ichnogenus Oichnus Bromley, 1981. It was inflicted starting from the outside of the valve and is oriented approximately perpendicular to the surface. The hole is not straight-sided but is parabolic, corresponding to the ichnotaxon Oichnus paraboloides Bromley, 1981. The outer borehole diameter (OBD) is 51.4 μm, and the inner borehole diameter (IBD) is 18 μm. The IBD/OBD ratio is of 0.35, which indicates a rather “non-functional” hole when compared to modern drilling activity (Kitchell et al., 1986; see Grey et al., 2005 for discussion on IBD/OBD ratio variations among naticid species). In modern marine environments (e.g., Ansell and Morton, 1987) and laboratory experiments (e.g., Kowalewski, 2004; Dietl and Kelley, 2006), it has been recognized that the ichnospecies O. paraboloides is left by predatory gastropods.
There are several possible origins for the observed drill hole. We exclude the hypothesis of organisms seeking for domicile (domichnia) because the boreholes left by such activities should rather be cylindrical, complete with multiple drill holes (Klompmaker et al., 2016). Boring organisms in hard substrates are also not favoured, as they should leave cylindrical and complete boreholes, which may penetrate the shell at an oblique angle. Oichnus paraboloides Bromley, 1981, might also be of parasitic origin, but in this scenario multiple holes of small size perpendicular to the surface should be observed (Klompmaker et al., 2016). Ostracods are known as hosts of parasites in recent environments as well as in the fossil record, for other crustaceans (Recent: Vannier and Abe, 1993), ciliates (Early Triassic: Weitschat and Guhl, 1994) or Pentastomida (Silurian: Siveter et al., 2015). The parasitic nature of the ciliates found on the inner part of an ostracod carapace from the Early Triassic of Spitzberg has been discussed (see Klompmaker and Boxshall, 2015). However, the parabolic morphology of the borehole on Bairdia sp. 4 excludes the action of a parasite located inside the carapace. Among crustacean parasites, some are known to develop internally while others attach externally but leave minimal lesions of a few microns in diameters. The incomplete borehole on the surface of Bairdia sp. 4 and the absence of attachment scars further tend to reject a parasitic origin.
The OBD and IBD of the drill hole is consistent with records from ostracods of younger assemblages (Table 2). To summarize, both the morphology and size of the borehole on Bairdia sp. 4 in Forel et al. (2017) from the G11 sample of the Tavusçayırı Block are consistent with a predatory origin. However, no potential predator has been found from this sample, which has only yielded siliceous sponges, radiolarians (Moix et al., 2007; Kozur et al., 2009) and ostracods (this work). Therefore, if a predator produced this trace, it remains unknown, but additional sampling may clarify this issue. At this time, we hypothesize that the origin of this borehole is predatory, as several elements are in line with this possibility. Two preliminary conclusions can be made: first, owing to the size of the drill hole on Bairdia sp. 4, it is likely that the culprit was a juvenile; second, since Bairdiidae are epibenthic organisms, the culprit might have been hunting at the sediment surface.
During the Mesozoic, drilling activity is sporadic until the end of the Cretaceous, and it subsequently increased in relation to the rise of Muricidae and Naticidae gastropods (Harper, 2006). The oldest indubitable group that includes drillers is reported from the Valanginian, Early Cretaceous (Kaim, 2004). Triassic occurrences of drill holes are rare, with only unquestionable occurrences in the Carnian and Norian (see summary and discussion in Klompmaker et al., 2016). The present report is a new addition to the Triassic record of drill holes and the very first known clue of Triassic drilling activity on micro-organisms.
MESOZOIC VERSUS PALAEOZOIC AFFINITY?
Following the end-Permian extinction that deeply affected marine ostracods worldwide (see Crasquin and Forel, 2014 for a review), the Triassic period is a turning point in their evolutionary history as it shows the shift from the Palaeozoic fauna to the Meso-Cenozoic or modern one. This transition extends from the oldest Meso-Cenozoic representatives in the Wuchiapingian (Late Permian) to the very last Palaeozoic taxa recently discovered from the Julian (Middle Carnian) of the Sorgun Ophiolitic Mélange (Tetraporobrachia haeckeli radiolarian Zone; Forel et al., 2017). Palaeocopida were major components of the Palaeozoic ostracod fauna in all types of marine environments (e.g., Becker and Wang, 1992; Olempska and Blazsyk, 1996). They had been severely hit by the end-Permian extinction and are only residual taxa during the Triassic up to their latest record in the Julian (Middle Carnian) of the Kilek section (Forel et al., 2017). In parallel, the Triassic saw the radiation of the Cypridoidea, Platycopida, sculptured Bairdiidae and Cytheroidea, which were mainly represented by the Bythocytheridae and Editiidae (e.g., Knüpfer, 1967; Gramm and Egorov, 1986; Olempska, 1999) in the Palaeozoic, (see Crasquin and Forel, 2014 and references therein).
The ostracod assemblage obtained from the sample G11 is in line with most of the neritic assemblages of the Late Triassic, in being largely dominated by Meso-Cenozoic taxa. Meso-Cenozoic genera represent 62% of the assemblage (17 species): they include 10 sculptured and two, typically Triassic, Bairdiidae genera (Bairdia (Urobairdia), Dicerobairdia, Isobythocypris, Medwenitschia, Mirabairdia, Nodobairdia, Petasobairdia, Ptychobairdia) and five typical Triassic or Mesozoic monotypic genera (Nemoceratina (Nemoceratina), Judahella, Ogmoconcha, Bektasia, Leviella). Palaeozoic-Mesozoic (panchronic) taxa are represented by 10 species: eight Bairdia species and two Bairdiacypris species. Acratia is the only Palaeozoic-Mesozoic genus, which is a common component of both deep and neritic water assemblages in the Early-Middle Triassic (e.g., Kozur, 1970b; Kristan-Tollmann, 1991; Forel and Crasquin, 2011; Forel, 2012; Forel and Grădinaru, 2018) and in the Carnian as well (Forel et al., 2017; Crasquin et al., 2018). Therefore, the G11 assemblage clearly differs from the assemblage of the slightly older Kilek section (Tetraporobrachia haeckeli radiolarian Zone) at the Tavusçayiri Block, where Palaeozoic taxa are abundant, witnessing a deep-sea refuge for Palaeozoic taxa up to the Carnian (Forel et al., 2017). One explanation for this discrepancy is that most of the last residual Palaeozoic taxa might have gone extinct between the Julian, Tetraporobrachia haeckeli Zone sampled in Kilek, and the Tuvalian, Spongotortilispinus moixi Zone studied here (Figure 3). A second possibility is that the G11 assemblage lacks Palaeozoic survivors because of the shallower water depths compared to the Kilek assemblage. The Julian Kilek assemblage developed in the outer platform-upper slope zone (Forel et al., 2017), representing a slightly greater water depth compared to the G11 assemblage of Tuvalian age. Only high resolution and large-scale sampling will help to clarify these questions and understand if the deep-water refuge was restricted to this peculiar geographical area or a rather widespread phenomenon in the southern part of a Neotethyan sub-basin.
CONCLUSIONS
Silicified ostracods of lower Tuvalian, Late Carnian age (Spongotortilispinus moixi radiolarian Zone) have been extracted from the sample G11 of the Tavusçayiri Block in the Mersin Mélange located in southern Turkey. This sample previously provided the best-preserved and most diverse radiolarian fauna of that age worldwide. The ostracod assemblage is composed of 29 species, which represent 16 genera and seven families. The species Bairdiacyris sorgunensis Forel sp. nov. is newly described. The environmental affinities of the recovered taxa suggest a moderately shallow subtidal environment subjected to environmental variations indicative of a shallowing upward trend from the Julian to the Tuvalian at the Tavusçayiri Block. We describe and discuss the occurrence of a drill hole at the left valve of Bairdia sp. 4, which is likely of predatory origin. It is the oldest known record of predatory activity on marine ostracods. All ostracods from the sample G11 are typical for the Triassic period worldwide and Palaeozoic taxa are missing. This observation implies that the Palaeozoic taxa from the Tavusçayiri Block sequence were either already extinct in the late Carnian or survived only in deeper waters.
ACKNOWLEDGEMENTS
The present study is MTA-MTM-ELTE Paleo Contribution No. 262. We are grateful to two anonymous reviewers and the editors who greatly improved an earlier version of this contribution.
REFERENCES
Alcalde, L., Derocco, N.N., and Rosset, S.D. 2010. Feeding in syntopy: Diet of Hydromedusa tectifera and Phrynops hilarii (Chelidae). Chelonian Conservation and Biology, 9:33-44. https://doi.org/10.2744/ccb-0794.1
Ansell, A.D. and Morton, B. 1987. Alternative predation tactics of a tropical naticid gastropod. Journal of Experimental Marine Biology and Ecology, 111:109-119. https://doi.org/10.1016/0022-0981(87)90049-9
Apostolescu, V. 1959. Ostracodes du Lias du Bassin de Paris. Revue de l’Institut Français du Pétrole, 14:795-826.
Baird, W. 1850. The Natural History of the British Entomostraca. Ray Society, London. https://doi.org/10.5962/bhl.title.1807
Becker, G. 2002. Contributions to Palaeozoic Ostracod Classification [POC], No. 24, Palaeozoic Ostracoda: the standard classification scheme. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen, 226:165-228. https://doi.org/10.1127/njgpa/226/2002/165
Becker, G. and Wang, S. 1992. Kirkbyacea and Bairdiacea (Ostracoda) from the Palaeozoic of China. Palaeontographica, Abteilung A, 224:1-54.
Bhatia, S.B., Sirjana, S., and Sunila, B. 1989. Predation by gastropod drills on early Lutetian ostracods from Kutch, Western India. Geobios, 22:387-394. https://doi.org/10.1016/s0016-6995(89)80138-x
Bradfield, H.H. 1935. Pennsylvanian Ostracoda of Ardmore Basin, Oklahoma. Bulletin of American Paleontology, 22:1-145.
Bromley, R.G. 1981. Concepts in ichnotaxonomy illustrated by small round holes in shells. Acta Geológica Hispánica, 16:55-64.
Bunza, G. and Kozur, H. 1971. Beiträge zur Ostracodenfauna der tethyalen Trias. Geologisch-Paläontologische Mitteilungen Innsbruck, 1:1-76.
Chen, D.Q. and Shi, C.G. 1982. Latest Permian Ostracoda from Nantong, Jiangsu and from Miannyang, Hubei. Bulletin of Nanjing Institute of Geology and Palaeontology, Academia Sinica, 4:105-152. (In Chinese with English abstract)
Costa, P.F., Oliveira, R.F., and da Fonseca, L.C. 2006. Feeding ecology of Nereis diversicolor (O. F. Müller) (Annelida, Polychaeta) on estuarine and lagoon environments in the southwest coast of Portugal. Pan-American Journal of Aquatic Sciences, 1:114-126.
Crasquin, S. and Forel, M.B. 2014. Ostracods (Crustacea) through Permian-Triassic events. Earth-Science Reviews, 137:52-64. https://doi.org/10.1016/j.earscirev.2013.01.006
Crasquin, S., Scuito, F., and Reitano, A. 2018. Carnian ostracods (Crustacea) of the Mufara Formation (Monte Scalpello, Central-Eastern Sicily, Italy). Annales de Paléontologie, 104:129-142. https://doi.org/10.1016/j.annpal.2018.03.001
Crasquin-Soleau, S. and Grădinaru, E. 1996. Early Anisian ostracode fauna from the Tulcea unit (Cimmerian North Dobrogean orogeny, Romania). Annales de Paléontologie, 82:59-116.
Dietl, G.P. and Kelley, P.H. 2006. Can naticid gastropod predators be discriminated by the holes they drill? Ichnos, 13:1-6. https://doi.org/10.1080/10420940600848889
Dumitrica, P., Tekin, U.K., and Bedi, Y. 2013. Taxonomic study of the tetrahedral, pentagonal and hexagonal spongy spumellarian Radiolaria from the middle Carnian (Late Triassic) of the Koseyahya nappe (Elbistan, SE Turkey) and other Triassic localities. Paläontologische Zeitschrift, 87:311-343. https://doi.org/10.1007/s12542-012-0160-2
Forel, M.B. 2012. Ostracods (Crustacea) associated with microbialites across the Permian-Triassic boundary in Dajiang (Guizhou Province, South China). European Journal of Taxonomy, 19:1-34. https://doi.org/10.5852/ejt.2012.19
Forel, M.B. and Crasquin, S. 2011. In the aftermath of Permian-Triassic boundary mass extinction: new ostracod (Crustacea) genus and species from South Tibet. Geodiversitas, 33:247-256. https://doi.org/10.5252/g2011n2a3
Forel, M.B. and Grădinaru, E. 2018. First report of Ostracods (Crustacea) associated with Bithynian (Anisian, Middle Triassic) Tubiphytes -microbial reef in the North Dobrogean Orogen (Romania). Papers in Palaeontology, 4:211-244. https://doi.org/10.1002/spp2.1103
Forel, M.B., Tekin, U.K., Okuyucu, C., Bedi, Y., Tuncer, A., and Crasquin, S. 2017. Discovery of a long-term refuge for ostracods (Crustacea) after the end-Permian extinction: a unique Carnian (Late Triassic) fauna from the Mersin Melange, southern Turkey. Journal of Systematic Palaeontology. https://doi.org/10.1080/14772019.2017.1391342
Gallet, Y., Besse, J., Krystyn, L., and Marcoux, J. 1994. Magnetostratigraphy of the Mayerling section (Austria) and Erenkolu Mezarlık (Turkey) section: improvement of the Carnian (Late Triassic) magnetic polarity time scale. Earth and Planetary Science Letters, 125:173-191. https://doi.org/10.1016/0012-821x(94)90214-3
Ghioca-Robrecht, D.M. and Smith, L.M. 2008. Feeding ecology of polymorphic larval barred tiger salamanders in playas of the Southern Great Plains. Canadian Journal of Zoology, 86:554-563. https://doi.org/10.1139/z08-027
Gramm, M.N. and Egorov, G.I. 1986. Early Carboniferous Editiidae (Ostracoda) and remarks on phylogeny of Cytheracea. Paleontologiheskij Zurnal, 2:50-60. (In Russian)
Grey, M., Boulding, E.G., and Brookfield, M.E. 2005. Shape differences among boreholes drilled by three species of naticid gastropods. Journal of Molluscan Studies, 71:253-256. https://doi.org/10.1093/mollus/eyi035
Gründel, J. and Kozur, H. 1972. Zur Taxonomie der Bythocytheridae und Tricorninidae (Podocopida, Ostracoda). Monatsberichte der Deutschen Akademie der Wissenschaften zu Berlin, 13(for 1971):907-937.
Harding, J.P. 1962. Mungava munda and four other new species of ostracod crustaceans from fish stomachs. The Natural History of Rennell Island, British Solomon Islands, 4:51-62.
Harloff, J. 1993. Ostracoden des Unter-Pliensbachiums in Baden-Württemberg. Stuttgarter Beiträge zur Naturkunde, Serie B, Geologie und Paläontologie, 191:1-214.
Harloff, J. and Jäger, R. 1994. Ostracoden aus dem Lias der Kalkalpen Bayerns und Nordtirols. Stuttgarter Beiträge zur Naturkunde, Serie B, Geologie und Paläontologie, 205:1-63.
Harper, E.M. 2006. Dissecting post-Palaeozoic arms races. Palaeogeography, Palaeoclimatology, Palaeoecology, 232:322-343. https://doi.org/10.1016/j.palaeo.2005.05.017
Herrig, E. 1979a. Ostrakoden aus dem Lias von Thüringen: Die Gattungen Bairdia (Teil II), Fabalicypris und Bairdiacypris. Zeitschrift für Geologische Wissenschaften, 7:763-782.
Herrig, E. 1979b. Weitere glattschalige Ostrakoden aus dem Lias von Thüringen. Zeitschrift für Geologische Wissenschaften, 7:1343-1361.
Hillebrandt, A.V., Krystyn, L., Kürschner, W.M., Bonis, N.R., Ruhl, M., Richoz, S., Schobben, M.A.N., Urlichs, M., Kment, K., McRoberts, C.A., Simms, M., and Tomãsových, A. 2013. The Global Stratotype Sections and Point (GSSP) for the base of the Jurassic System at Kuhjoch (Karwendel Mountains, Northern Calcareous Alps, Tyrol, Austria). Episodes, 36:162-198.
Horne, D.J., Cohen, A., and Martens, K. 2002. Taxonomy, morphology and biology of Quaternary and living Ostracoda, p. 5-36. In Holmes, J.A. and Chivas, A. (eds.), The Ostracoda: Applications in Quaternary Research. Geophysical Monograph, American Geophysical Union, Washington, D.C. https://doi.org/10.1029/131gm02
Hornung, T., Brandner, R., Krystyn, L., Joachimski, M.M., and Keim, L. 2007. Multistratigraphic constraints on the NW Tethyan ‘Carnian Crisis’. New Mexico Museum of Natural History and Science Bulletin, 41:59-67.
Hussain, S.M., Ravi, G., Mohan, S.P., and Rajeshwara Rao, N. 2004. Recent Benthic Ostracoda from the inner shelf off Chennai, southeast coast of India-implication of microenvironments. Environmental Micropaleontology, Microbioloogy and Meiobenthology, 1:105-121.
Kaim, A. 2004. The evolution of conch ontogeny in Mesozoic open sea gastropods. Acta Palaeontologica Polonica, 62:1-183.
Karanovic, I. and Brandão, S.N. 2016. The genus Polycope (Polycopidae, Ostracoda) in the North Atlantic and Arctic: Taxonomy, distribution, and ecology. Systematics and Biodiversity, 14(2):198-223. https://doi.org/10.1080/14772000.2015.1131756
Kitchell, J.A., Boggs, C.H., Rice, J.A., Kitchell, J.F., Hoffman, A., and Martinell, J. 1986. Anomalies in naticid predatory behavior: a critique and experimental observations. Malacologia, 27:291-298.
Klompmaker, A.A. and Boxshall, G.A. 2015. Fossil crustaceans as parasites and hosts. Advances in Parasitology, 90:233-289. https://doi.org/10.1016/bs.apar.2015.06.001
Klompmaker, A.A., Nützel A., and Kaim A. 2016. Drill hole convergence and a quantitative analysis of drill holes in mollusks and brachiopods from the Triassic of Italy and Poland. Palaeogeography, Palaeoclimatology, Palaeoecology, 457:342-359. https://doi.org/10.1016/j.palaeo.2016.06.017
Knüpfer, J. 1967. Editia germanica sp. n. eine neue Ostracodenart aus dem Unterkarbon im Norden der DDR und zur Taxonomie der Familie Bythocytheridae Sars 1926. Berichte der Deutschen Gesellschaft für Geologische Wissenschaften, Reihe A (Geologie und Paläontologie), 12:193-200.
Kollmann, K. 1960. Ostracoden aus der alpinen Trias. I. Parabairdia n. g. und Ptychobairdia n. g. (Bairdiidae). Jahrbuch der Geologischen Bundesanstalt, 5:79-105.
Kollmann, K. 1963. Ostracoden aus der alpinen Trias. II. Weitere Bairdiidae. Jahrbuch der Geologischen Bundesanstalt, 106:121-203.
Kowalewski, M. 2004. Drill holes produced by the predatory gastropod Nucella lamellosa (Muricidae): palaeobiological and ecological implications. Journal of Molluscan Studies, 70:359-370. https://doi.org/10.1093/mollus/70.4.359
Kozur, H. 1970a. Neue Ostracoden aus der germanischen Mittel- und Obertrias. Geologie, 19:434-455.
Kozur, H. 1970b. Neue Ostracoden-Arten aus dem obersten Anis des Bakonyhochlandes (Ungarn). Berichte des Naturwissenschaftlichen Vereins Innsbruck, 58:384-428.
Kozur, H. 1971a. Die Bairdiacea der Trias. Teil 3: Einige neue Arten triassischer Bairdiacea und Bemerkungen zur Herkunft der Macrocyprididae (Cypridacea). Geologisch Paläontologische Mitteilungen, Innsbruck, 1(6):1-18.
Kozur, H. 1971b. Die Bairdiacea der Trias. Teil 1: Skulpturierte Bairdiidae aus mitteltriassischen Flachwasserablagerungen. Geologisch-Paläontologische Mitteilungen Innsbruck, 1(3):1-27.
Kozur, H. 1997. Late Paleozoic and Early Mesozoic opening of Tethyan oceanic and continental basins in Turkey, p. 77. In De Wever, P. and Caulet, J.P. (eds.), InterRad VIII abstracts, Laboratoire de Géologie du Muséum Paris, Paris/Bierville, France, 8-13 Septembre 1997. (Publisher unknown).
Kozur, H. 2003. Integrated ammonoid, conodont and radiolarian zonation of the Triassic and some remarks to stage/substage subdivision and the numeric age of the Triassic stages. Albertiana, 28:57-74.
Kozur, H., Moix, P., and Ozsvárt, P. 2007a. Characteristic Nassellaria of the lower Tuvalian (Upper Triassic) Spongotortilispinus moixi Zone of the Huğlu Unit in the Mersin Mélange, southeastern Turkey. Bulletin de la Société Vaudoise des Sciences Naturelles, 90(3):151-173.
Kozur, H., Moix, P., and Ozsvárt, P. 2007b. Stratigraphically important Spumellaria and Entactinaria from the lower Tuvalian (Upper Triassic) of the Huğlu Unit in the Mersin Mélange, south-eastern Turkey. Bulletin de la Société Vaudoise des Sciences Naturelles, 90(3):175-195.
Kozur, H., Moix, P., and Ozsvárt, P. 2007c. Further new Nassellaria of the lower Tuvalian (Upper Triassic) Spongotortilispinus moixi Zone of the Huğlu Unit in the Mersin Mélange. Bulletin de la Société Vaudoise des Sciences Naturelles, 90(4):197-215.
Kozur, H., Moix, P., and Ozsvárt, P. 2009. New Spumellaria (Radiolaria) from the Early Tuvalian Spongotortilispinus moixi Zone of Southeastern Turkey, with some Remarks on the Age of this Fauna. Jahrbuch der Geologischen Bundesanstalt, 149:25-59.
Kozur, H. and Mostler, H. 1971. Holothurien-Sklerite und Conodonten aus der Mittel- und Obertrias von Köveskál (Balatonhochland, Ungarn). Geologisch-Paläontologische Mitteilungen Innsbruck, 1(10):1-36.
Kozur, H. and Mostler, H. 1994. Anisian to Middle Carnian radiolarian zonation and description of some stratigraphically important radiolarians. Geologisch-Paläontologische Mitteilungen Innsbruck, Sonderband 3:39-255.
Kozur, H. and Mostler, H. 1996. Longobardian (Late Ladinian), Oertlispongidae (Radiolaria) from the Republic of Bosnia-Hercegovina and the stratigraphic value of advanced Oertispongidae. Geologisch-Paläontologische Mitteilungen Innsbruck, Sonderband 4:105-193.
KristanTollmann, E. 1971. Weitere Beobachtungen an skulptierten Bairdiidae (Ostrac.) der alpinen Trias. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen, 139:57-81.
Kristan-Tollmann, E. 1978. Bairdiidae (Ostracoda) aus den obertriadischen Cassianer Schichten der Ruones-Wiesen bei Corvara in Südtirol. Schriftenreihe der Erdwissenschaftlichen Kommissionen, Österreichische Akademie der Wissenschaften, 4:77-104.
Kristan-Tollmann, E. 1988. Unexpected microfaunal communities within the Triassic Tethys. Geological Society of London, Special Publication, 37:213-223. https://doi.org/10.1144/gsl.sp.1988.037.01.13
Kristan-Tollmann, E. 1991. Ostracods from the Middle Triassic Sina Formation (Aghdarband Group) in NE-Iran. Abhandlungen der Geologischen Bundesanstalt, 38:195-200.
Kristan-Tollmann, E., Haas, J., and Kovács, S. 1991. Karnische Ostracoden und Conodonten der Bohrung Zsamek-14 im Transdanubischen Mittelgebirge (Ungarn). Jubiläumsschrift 20 Jahre Geologische Zusammenarbeit Österreich-Ungarn, 1:193-219.
Kristan-Tollmann, E. and Hamedani, A. 1973. Eine spezifische Mikrofaunen- Vergesellschaftung aus den Opponitzer Schichten der Oberkarn der niederoesterreichischen Kalkvoralpen. Neues Jahrbuch für Geologie und Paläontologie. Abhandlungen, 143(2):193-222.
Kristan-Tollmann, E., Tollmann, A., and Hamedani, A. 1980. Beiträge zur Kenntnis der Trias von Persien. II Zur Rhätfauna von Bagerabad bei Isfahan (Korallen, Ostracoden). Mitteilungen der Österreichischen Geologischen Gesellschaft, 73:163-235.
Latreille, P.A. 1806. Genera Crustaceorum et Insectorum: Secundum Ordinem Naturalem in Familias Disposita, Iconibus Exemplisque Plurimis Explicata. Tomus 1. Koenig, Paris. https://doi.org/10.5962/bhl.title.4819
Leal, J.H. 2008. A remarkable new genus of carnivorous, sessile bivalves (Mollusca: Anomalodesmata: Poromyidae) with descriptions of two new species. Zootaxa, 1764:1-18.
Lethiers, F. and Raymond, D. 1991. Les crises du Dévonien supérieur par l’étude des faunes d’ostracodes dans leur cadre paléogéographique. Palaeogeography, Palaeoclimatology, Palaeoecology, 88:133-146. https://doi.org/10.1016/0031-0182(91)90019-n
Lowndes, A.G. 1930. Living ostracods in the rectum of a frog. Nature, 126:958. https://doi.org/10.1038/126958c0
Maddocks, R.F. 1988. One hundred million years of predation on ostracods: the fossil record in Texas, p. 637-657. In Hanai, T., Ikeya, N., and Ishizaki, K. (eds.), Evolutionary Biology on Ostracoda, Proceedings of the Ninth International Symposium on Ostracoda. Kodansha Ltd., Tokyo, Japan. https://doi.org/10.1016/s0920-5446(08)70212-7
Maddocks, R.F. 2015. New and poorly known species of Bairdoppilata and Paranesidea (Bairdiidae, Ostracoda) from French Frigate Shoals and O’ahu, the Hawaiian Islands. Zootaxa, 4059:277-317. https://doi.org/10.11646/zootaxa.4059.2.3
Masset, O. and Moix, P. 2004. Les Mélanges de L’ophiolite de Mersin (Turquie du Sud). Unpublished MSc Thesis, Université de Lausanne, Switzerland.
McCoy, F. 1844. A Synopsis of the Characters of the Carboniferous Limestone Fossils of Ireland. Dublin University Press, Dublin. https://doi.org/10.5962/bhl.title.11559
Mette, W., Elsler, A., and Korte, C. 2012. Palaeoenvironmental changes in the Late Triassic (Rhaetian) of the Northern Calcareous Alps: Clues from stable isotopes and microfossils. Palaeogeography, Palaeoclimatology, Palaeoecology, 350-352:62-72. https://doi.org/10.1016/j.palaeo.2012.06.013
Mette, W., Honigstein, A., and Crasquin, S. 2014. Deep-water ostracods from the Middle Anisian (Reifling Formation) of the Northern Calcareous Alps (Austria). Journal of Micropalaeontology, 34:71-91. https://doi.org/10.1144/jmpaleo2014-009
Mette, W. and Mohtat-Aghai, P. 1999. Ostracods and foraminifera from Upper Triassic intrashelf basin deposits in the Northern Calcareous Alps. Geologisch Paläontologische Mitteilungen Innsbruck, 24:45-77.
Moix, P., Beccaletto, L., Masset, O., Kozur, H., Dumitrica, P., Vachard, D., Martini, R., and Stampfli, G. 2011. Geology and correlation of the Mersin Mélanges, Southern Turkey. Turkish Journal of Earth Sciences, 20:57-98.
Moix, P., Kozur, H., Stampfli, G.M., and Mostler, H. 2007. New palaeontological, biostratigraphical and palaeogeographial results from the Triassic of the Mersin mélange, SE Turkey, p. 282-311. In Lucas, S.G. and Spielmann, J.A. (eds.), The Global Triassic. New Mexico Museum of Natural History and Science Bulletin 41. New Mexico Museum of Natural History and Science, Albuquerque.
Moix, P., Vachard, D., Allibon, J., Martini, R., Wernli, R., Kozur, H., and Stampfli, G.M. 2013. Palaeotethyan, Neotethyan and Huglu-Pindos series in the Lycian Nappes (SW Turkey): geodynamical implications, p. 401-444. In Tanner, L.H., Spielmann, J.A., and Lucas, S.G. (eds.), The Triassic System. New Mexico. New Mexico Museum of Natural History and Science Bulletin 61. New Mexico Museum of Natural History and Science, Albuquerque.
Monod, O. 1977. Recherches Géologiques dans le Taurus Occidental au Sud de Beysehir (Turquie). Ph.D. thesis, Université de Paris-Sud, Orsay, France.
Monostori, M. 1996. Pliensbachian ostracod fauna from condensed limestones of the Bakony Mts. (Transdanubian Central Range, Hungary). Fragmenta Mineralogica et Palaeontologica, 18:31-61.
Monostori, M. and Tóth, E. 2013. Ladinian (Middle Triassic) silicified ostracod faunas from the Balaton Highland (Hungary). Rivista Italiana di Paleontologia e Stratigrafia, 119:303-323.
Monostori, M. and Tóth, E. 2014. Additional Middle to Upper Triassic ostracod faunas from the boreholes of Transdanubian Central Range (Hungary). Hantkeniana, 9:21-43.
Moore, R.C. (ed.) 1961. Treatise on Invertebrate Paleontology. Part Q. Arthropoda 3, Crustacea, Ostracoda. Geological Society of America and University of Kansas Press, Boulder and Lawrence, 1-442.
Müller, G.W. 1894. Die Ostracoden des Golfes von Neapel und der angrenzenden Meeres Abschnitte. Fauna und Flora Neapel, 21:1-404. https://doi.org/10.5962/bhl.title.7419
Olempska, E. 1999. Silicified shallow-water ostracodes from the Early Carboniferous of South China. Acta Palaeontologica Polonica, 44:383-436.
Olempska, E., and Blaszyk, J. 1996. Ostracods from Permian of Spitsbergen. Polish Polar Research, 17:3-20.
Özer, E., Koç, H. and Özsayar, T.Y. 2004. Stratigraphical evidence for the depression of the northern margin of the Menderes-Tauride Block (Turkey) during the Late Cretaceous. Journal of Asian Earth Sciences, 22:401-412. https://doi.org/10.1016/s1367-9120(03)00084-1
Özgül, N. 1976, Toroslarýn bazı temel jeoloji özellikleri. Türkiye Jeoloji Kurumu Bülteni, 19:65-78.
Ozsvárt, P., Dumitrica, P., Hungerbühler, A., and Moix, P. 2017a. Mono- and dicyrtid Nassellaria (Radiolaria) from the Upper Carnian of the Sorgun Ophiolitic Mélange, Southern Turkey and Kopría Mélange, Rhodes, Greece. Revue de Micropaléontologie, 60:137-160. https://doi.org/10.1016/j.revmic.2016.11.004
Ozsvárt, P., Dumitrica, P., and Moix, P. 2017b. New Early Tuvalian (Carnian, Triassic) radiolarians from the Huglu-Pindos succession in the Sorgun Ophiolitic Mélange, Southern Turkey. Ofioliti, 42:55-67.
Ozsvárt, P., Moix, P., and Kozur, H. 2015. New Carnian (Upper Triassic) radiolarians from the Sorgun Ophiolitic Mélange, Southern Turkey. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen, 277:337-352. https://doi.org/10.1127/njgpa/2015/0509
Parlak, O. 1996. Geochemistry and Geochronology of the Mersin Ophiolite Within the Eastern Mediterranean Tectonic Frame (Southern Turkey). Ph.D. Thesis, Université de Genève, Genève, Switzerland.
Penchaszadeh, P E., Bigatti, G., and Miloslavich, P. 2004. Feeding of Pseudechinus magellanicus (Philippi, 1857) (Echinoidea: Temnopleuridae) in the SW Atlantic Coast (Argentina). Ophelia, 58:91-99. https://doi.org/10.1080/00785326.2004.10410216
Reyment, R.A. 1963. Bohrlöcher bei Ostracoden. Paläontologische Zeitschrift, 37:283-291. https://doi.org/10.1007/bf02987919
Reyment, R.A. 1966. Preliminary observations on gastropod predation in the Western Niger Delta. Palaeogeography, Palaeoclimatology, Palaeoecology, 2:81-102. https://doi.org/10.1016/0031-0182(66)90010-1
Reyment, R.A. 1967. Paleoethology and fossil drilling gastropods. Transactions of the Kansas Academy of Science, 70:33-50. https://doi.org/10.2307/3627612
Reyment, R.A. and Elewa, A.M.T. 2002. Predation by drills on Ostracoda. Predator - prey interactions in the fossil record. Topics in Geobiology, 20:93-111. https://doi.org/10.1007/978-1-4615-0161-9_5
Reyment, R.A., Reyment, E.R., and Honigstein, A. 1987. Predation by boring gastropods on Late Cretaceous and Early Palaeocene ostracods. Cretaceous Research, 8:189-209. https://doi.org/10.1016/0195-6671(87)90021-8
Robertson, A.I. 1988. Abundance, diet and predators of juvenile banana prawns, Penaeus merguiensis, in a tropical mangrove estuary. Australian Journal of Marine and Freshwater Research, 39:467-478. https://doi.org/10.1071/mf9880467
Rossi, V., Benassi, G., Belletti, F., and Menozzi, P. 2011. Colonization, population dynamics, predatory behaviour and cannibalism in Heterocypris incongruens (Crustacea: Ostracoda). Journal of Limnology, 70:102-108. https://doi.org/10.4081/jlimnol.2011.102
Roth, R. 1928. Monoceratina: a new genus of Ostracoda from the Pennsylvanian of Oklahoma. Journal of Paleontology: 2,15-19.
Ruiz, F., Abad, M., Gonzalez-Regalado, M.L., Civis, J., Gonzalez-Delgado, J.A., Mara García, E.X., and Toscano, A. 2010. Predation on Neogene ostracods of southwestern Spain. Rivista Italiana di Paleontologia e Stratigrafia, 116:253-260.
Ruiz, F., Abad, M., Mara García, E.X., Toscano, A., Prudencio, M.I., Dias, M.I., and Galán, E. 2011. Predation on ostracod populations of two North African lagoons. Crustaceana, 84: 1537-1545. https://doi.org/10.1163/156854011x605846
Rutherford, M.M. and Ainsworth, N.R. 1989. Micropalaeontological and stratigraphical recognition of the Triassic-Jurassic boundary in the North Celtic Sea and Fastenit Basins, p. 45-69. In Batten, D.J. and Keen, M.C. (eds.), North-west European Micropalaeontology and Palynology. Ellis Horwood Ltd, Chichester.
Salaj, J. and Jendrejáková, O. 1984. Ecology and facial relation of some groups of Triassic foraminifers and ostracods of stratigraphic importance. Geologickÿ Zbornĭk-Geologica Carpathica, 35:231-240.
Sars, G.O. 1866. Oversigt af marine Ostracoder. Norske Videnskaps-Akademi, Förhandlingar, 1865:1-130.
Sars, G.O. 1887. Nye bidrag til kundskaben om middelhavets invertebrafauna: 4. Ostracods mediterranea (sydeuropaeiske ostracoder). Archiv for Mathematik og Naturvidenskab, 12:173-324. https://doi.org/10.5962/bhl.title.10252
Schornikov, E.I. 1990. Evolution and classification of Bythocytheridae. Courier Forschungsinstitut Senckenberg, 123:291-302.
Sebe, O.G., Crasquin, S., and Grădinaru, E. 2013. Early and Middle Anisian deep-water ostracods (Crustacea) from North Dobrogea (Romania). Revue de Paléobiologie, 32:509-529.
Siveter, D.J., Briggs, D.E.G., Siveter, D.J., and Sutton, M.D. 2015. A 425-million-year-old Silurian pentastomid parasitic on ostracods. Current Biology, 25:1632-1637. https://doi.org/10.1016/j.cub.2015.04.035
Sohn, I.G. 1954. Ostracodes from the Permian of the Glass Mountains, Texas. United States Geological Survey Professional Paper, 264-A:1-24.
Sohn, I.G. 1968. Triassic ostracodes from Makhtesh Ramon, Israel. Bulletin of the Geological Survey of Israel, 44:1-71.
Stampfli, G.M. 2000. Tethyan oceans, p. 1-23. In Bozkurt, E., Winchester, J.A., and Piper, J.D.A. (eds.), Tectonics and Magmatism in Turkey and Surrounding Area. Geological Society of London Special Publication 173. The Geological Society of London, London.
Ulrich, E.O. and Bassler, R.S. 1923. Paleozoic Ostracoda: their morphology, classification and occurrence. Maryland Geological Survey, Silurian, 9:271-391.
Upson, M.E. 1933. The Ostracoda of the Big Blue Series in Nebraska. Bulletin, Nebraska Geological Survey, Second Series, 8:1-64.
Urlichs, M. 1972. Ostracoden aus den Kössener Schichten und ihre Abhängigkeit von der Ökologie. Mitteilungen der Gesellschaft der Geologie- und Bergbaustudenten in Österreich, 21:661-710.
Vandekerhove, J., Namiotko, T., Hallmann, E., and Martens, K. 2012. Predation by macroinvertebrates on Heterocypris incongruens (Ostracoda) in temporary ponds: impacts and responses. Fundamental and Applied Limnology, 181:39-47. https://doi.org/10.1127/1863-9135/2012/0310
Vannier, J. and Abe, K. 1993. Functional morphology and behavior of Vargula hilgendorfii (Ostracoda: Myodocopida) from Japan, and discussion of its crustacean ectoparasites: preliminary results from video recordings. Journal of Crustacean Biology, 13:51-76. https://doi.org/10.2307/1549122
Weitschaft, W. and Guhl, W. 1994. Erster Nachweis fossiler Ciliaten. Paläontologische Zeitschrift, 68:17-31. https://doi.org/10.1007/bf02989430
Whatley, R. and Boomer, I.D. 2000. Systematic review and evolution of the early Cytheruridae (Ostracoda). Journal of Micropalaeontology, 19:139-151. https://doi.org/10.1144/jm.19.2.139
