

 First record of Late Pleistocene holoplanktonic mollusks (Gastropoda, Euthecosomata, Pterotracheoidea) from northern Taiwan
First record of Late Pleistocene holoplanktonic mollusks (Gastropoda, Euthecosomata, Pterotracheoidea) from northern Taiwan
Article number: 28.2.a24
https://doi.org/10.26879/1535
Copyright Palaeontological Association, June 2025
Author biographies
Plain-language and multi-lingual abstracts
PDF version
Submission: 24 January 2025. Acceptance: 23 May 2025
ABSTRACT
Pelagic gastropods, including pteropods and heteropods, are cosmopolitan organisms recorded in Cenozoic deposits across the globe. Their fossils serve as valuable tools for paleoenvironmental reconstruction and enable precise stratigraphic correlations. Numerous Neogene and Quaternary assemblages of pelagic gastropods have been documented in the Indo-West Pacific region, spanning from Indonesia to central Japan. In Taiwan, molluscan fossil assemblages are abundant and diverse, predominantly comprising gastropods and bivalves. However, pteropod records from Taiwan are limited, with only two species previously reported from the Oligocene. This study investigates a previously undocumented Late Pleistocene assemblage of holoplanktonic pelagic gastropods from the Toukoshan Formation in northern Taiwan. The pteropod fauna includes nine taxa from the families Creseidae and Cavoliniidae, and one species of the heteropod family Atlantidae. Notably, the Toukoshan species Diacria cf. philippinensis and Diacavolinia cf. bandaensis represent the first Pleistocene records. Additionally, the heteropod Atlanta turriculata marks the first occurrence of this group in Taiwan. All identified species are epipelagic, corroborating earlier depth estimates for the fossil-bearing strata. The Toukoshan holoplanktonic assemblage shares similarities with other Pleistocene faunas from Okinawa and Honshu in Japan, highlighting regional paleoceanographic connections during this period.
Diana Osipova. Biodiversity Research Center, Academia Sinica, 128 Academia Road, Sec. 2, Nankang, Taipei 11529, Taiwan, Biodiversity Program, Taiwan International Graduate Program, Academia Sinica and National Taiwan Normal University, School of Life Science, National Taiwan Normal University, Heping E Rd, Da’an, Taipei 106308 Taiwan osipova.ds@outlook.com
Chien-Hsiang Lin. Biodiversity Research Center, Academia Sinica, 128 Academia Road, Sec. 2, Nankang, Taipei 11529, Taiwan. corresponding author kurosaki@gate.sinica.edu.tw
Keywords: Pteropoda; Toukoshan Formation; Indo-West Pacific; pelagic; taxonomy
Final citation: Osipova, Diana and Lin, Chien-Hsiang. 2025. First record of Late Pleistocene holoplanktonic mollusks (Gastropoda, Euthecosomata, Pterotracheoidea) from northern Taiwan. Palaeontologia Electronica, 28(2):a24.
https://doi.org/10.26879/1535
palaeo-electronica.org/content/2025/5549-pleistocene-holoplanktonic-mollusks-from-taiwan
Copyright: June 2025 Palaeontological Association.
This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
creativecommons.org/licenses/by/4.0
INTRODUCTION
Holoplanktonic gastropods, comprising pteropods (order Pteropoda Cuvier, 1804) and heteropods (superfamily Pterotracheoidea Rafinesque, 1814 of the order Littorinimorpha Golikov and Starobogatov, 1975), are ubiquitous in the world’s oceans (Janssen et al., 2019). Due to their aragonitic shell composition, these gastropods are prone to poor preservation unless deposited above the aragonite compensation depth (ACD), which is shallower than the calcite compensation depth (Gerhardt and Henrich, 2001; Janssen, 2017; Peijnenburg et al., 2020). Under high acidity, water becomes undersaturated with aragonite, which decreases ACD and promotes pteropod shells to dissolve even within their normal distribution depth. This makes them effective bioindicators of ocean acidification (Bednarsek et al., 2016). Moreover, their abundance and distribution in geological sequences have been extensively used for palaeoenvironmental reconstruction and stratigraphic correlations (Chen, 1969; Reiss et al., 1980; Shibata and Ishigaki, 1981; Buccheri, 1984). Assemblage analyses of pteropods are particularly useful for estimating water depth (Chen and Huang, 1990), climatic zones (Messenger et al., 2010; Keul et al., 2017; Hallenberger et al., 2022), and depositional environments (Buccheri et al., 1980; Wang et al., 1997; Hallenberger et al., 2022; Beccari et al., 2023).
Pteropods first appeared during the Late Mesozoic (Janssen and Goedert, 2016) and diversified rapidly during the Paleogene, continuing into the Quaternary (Bernasconi, 1982; Janssen and Peijnenburg, 2017). In Asia, records of pteropods date back to the Paleogene, with significant faunal changes observed across the boundary between the Eocene and Oligocene (Ando et al., 2009; Ando, 2011). Recently, early Eocene pteropods from the Zhepure Formation in Tibet have provided insights into the paleogeography of the Neo-Tethys residual basin and potential dispersal pathways to Asia (Li et al., 2020). Additional records of Neogene and Quaternary holoplanktonic gastropods are known from the Fiji Islands (Ladd, 1934; Janssen, 1999), Indonesia (Janssen, 1999), and South Australia (Janssen, 1989).
Neogene and Quaternary West Pacific occurrences of holoplanktonic gastropods are predominantly reported from Japan, with comprehensive analyses presented by Janssen (2007). These records range from southern Okinawa (Noda, 1972; Shibata and Ujihara, 1983) to central Honshu (Shibata, 1986). Diverse and abundant Pliocene assemblages have also been reported from Pangasinan, Luzon, in the Philippines (Janssen, 2007). In contrast, heteropod fossils remain less common, with sporadic reports from Japan, the Philippines, and Fiji, spanning the Miocene to Pleistocene (Shibata, 1977, 1984; Janssen and Grebneff, 2012).
Taiwan’s location in the West Pacific and its relatively young geological history, marked by the ongoing collision of the Eurasian and Philippine Sea plates, has resulted in significant tectonic uplift since the Miocene-Pliocene transition (Lin, A.-T. et al., 2003; Lin, C.-H. et al., 2022). This uplift exposed marine strata of various ages in the Western Foothills, where abundant fossils of marine invertebrates, including mollusks, crabs, ostracods, and foraminifera, have been discovered (e.g., Yabe and Hanzawa, 1930; Hu, 1984; Masuda and Huang, 1990). Among these, mollusks are the most diverse and extensively studied, with over 1,000 species recorded from the Oligocene to the Quaternary (Kanno and Chung, 1975; Huang et al., 1990; Hu, 1991–1995).
The first report of holoplanktonic gastropods in Taiwan came from Oligocene formations near Taipei, where two taxa were identified as belonging to the family Cavoliniidae (Clio sp.) (Chen, 1981). Subsequent studies described these specimens as Clio mai Chen and Huang, 1990, along with new Limacina lini Chen and Huang, 1990, which helped reconstruct the paleodepths of the Oligocene basin in northern Taiwan (Chen and Huang, 1990). However, the diversity and geological history of Taiwan’s holoplanktonic mollusks remain largely understudied, with no heteropod fossils reported from Taiwan to the best of our knowledge. This research aims to address this gap by providing new data and taxonomic descriptions of Late Pleistocene pelagic gastropod fossils from the Toukoshan Formation.
GEOLOGICAL SETTINGS
The Toukoshan Formation, consisting of Pleistocene strata, was introduced by Lin (1935) and Torii (1935). Its type locality is at the Touko mountain, Xinshe District, east of Fengyuan District in Taichung City (Chou, 1992). This formation overlies the Lower Pleistocene Cholan Formation (Kotaka, 1973; Chou, 1992; Nagel et al., 2013). Its thickness reaches over 3,000 m in the Taichung region (Hu, 1978; Chou, 1992; Liu et al., 2022).
Chang et al. (1965) initially divided the local Toukoshan Formation into the Wumeikeng, Nanwo, and Hutou members, in ascending order. However, most of the later studies classified the formation into two members: the lower Hsiangshan Member and the upper Huoyenshan Conglomerate Member (e.g., Chou, 1973), with fine-grained sandstone at the lower part and coarse-grained fluvial sediments dominating the upper part (Torii, 1935; Chang, 1948, 1955; Kotaka, 1973; Chen et al., 2001). Paleontological and sedimentary studies suggest that the Toukoshan Formation was deposited in environments ranging from shallow neritic and littoral to fluvial-deltaic from bottom to top (Hayasaka, 1933, 1934; Chou, 1973). In northwestern and west-central Taiwan, it is often unconformably overlain by terrace deposits of fluvial-lacustrine or marine-brackish facies (Liew, 1988).
The fossil-bearing layer examined here is part of the uppermost Nanwo Member (also known as uppermost part of Hsiangshan Member) and comprises gray to dark gray sandstone and shale interbeds (Hu, 1992; Lee, 2000, figures A.1, A.2). This layer contains abundant fossils, including sea urchins, mollusks, crabs, and smaller fossils such as foraminifera, ostracods, and bryozoans (Kotaka, 1973; Hu, 1978, 1992). Comparing with recent foraminiferal data from the Taiwan Strait, Chow (1980) estimated water depths between 30 and 60 m for the fossil-bearing layer.
The age of the Toukoshan Formation remains controversial. Biostratigraphic studies on planktonic foraminifera indicate that the formation’s deposition began after the Olduvai event (Huang, 1984), with a time range from 1.24 to 0.46 Ma (Lee et al., 2002; Nigel et al., 2013). Based on vertebrate fossils, Lin (1963) proposed that its age might correspond to the Calabrian to Cromerian stages. More recent analyses, however, suggest a younger age, possibly younger than 0.4 to 0.5 Ma based on pollen evidence (Liew, 1988). Interestingly, radiocarbon dating of some fossil scallops (Wang and Peng, 1991) and echinoids (Ho et al., 2022) from nearby outcrops indicates ages of 30 to 50 ka, specifically Marine Isotope Stage (MIS) 3 of the last Glacial (Ho et al., 2022). This suggests the formation could be younger than expected, though some argue the dating materials may have been altered.
MATERIAL AND METHODS
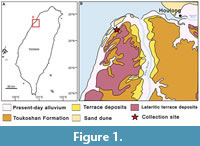 The sediment samples were collected at the outcrops of the Toukoshan Formation, Miaoli County, Xihu Township, Taiwan in the summer season of 2023 (24°35′51.66″N 120°43′32.63″E; Figure 1). The bulk sediments with a total weight of 45 kg were wet sieved through a 500-µm mesh and dried in a 40°C oven. Subsequently, all samples were examined under a microscope to isolate the shells of holoplanktonic gastropods for detailed analysis. A total of 77 specimens were recovered, of which 24 were analyzed in this study. Specimens were identified, described, measured (Table 1), and photographed. For pteropods with elongated conical shell width (WSh) and height (HSh) of the shells were measured. For species with globose shells four additional parameters were measured: aperture height (HA), aperture width (WA), joint width (WJ), and rostrum height (HR). For the heteropod specimen HSh, HA and WA were measured, as well as shell diameter (ShD). All specimens are deposited in the Biodiversity Research Museum, Academia Sinica, Taiwan, under the code ASIZF.
The sediment samples were collected at the outcrops of the Toukoshan Formation, Miaoli County, Xihu Township, Taiwan in the summer season of 2023 (24°35′51.66″N 120°43′32.63″E; Figure 1). The bulk sediments with a total weight of 45 kg were wet sieved through a 500-µm mesh and dried in a 40°C oven. Subsequently, all samples were examined under a microscope to isolate the shells of holoplanktonic gastropods for detailed analysis. A total of 77 specimens were recovered, of which 24 were analyzed in this study. Specimens were identified, described, measured (Table 1), and photographed. For pteropods with elongated conical shell width (WSh) and height (HSh) of the shells were measured. For species with globose shells four additional parameters were measured: aperture height (HA), aperture width (WA), joint width (WJ), and rostrum height (HR). For the heteropod specimen HSh, HA and WA were measured, as well as shell diameter (ShD). All specimens are deposited in the Biodiversity Research Museum, Academia Sinica, Taiwan, under the code ASIZF.
SYSTEMATIC PALEONTOLOGY
Class GASTROPODA Cuvier, 1795
Order LITTORINIMORPHA Golikov and Starobogatov, 1975
Superfamily PTEROTRACHEOIDEA Rafinesque, 1814
Family ATLANTIDAE Rang, 1829
Genus ATLANTA Lesueur, 1817
Type species. Atlanta peronii Lesueur, 1817, type by subsequent designation.
Atlanta turriculata d’Orbigny, 1836
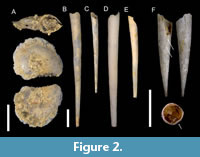
Figure 2A
*1836 Atlanta turriculata d’Orbigny, p. 173, pl. XX, figs. 5–11.
.1983 Atlanta turriculata d’Orbigny; Shibata and Ujihara, p. 157, pl. 47, fig. 4.
 Material. Miaoli County, Houlong Township; Toukoshan Formation, Late Pleistocene; 1 specimen, ASIZF0101021.
Material. Miaoli County, Houlong Township; Toukoshan Formation, Late Pleistocene; 1 specimen, ASIZF0101021.
Description. Shell very small, to 1.88 mm in diameter. Spire slender and high, protrudes over body whorls. Teleoconch consist of approximately 5 whorls (precise number cannot be counted due to preservation of specimen). Sculpture on whorls not observed. Keel well-developed and run string from 1,5 of penultimate whorl. Edge of keel broken. Umbilicus narrow, moderately deep. Aperture sub-circular, narrowing toward keel.
Remarks. The single complete specimen was recovered from the sediments in our examined locality. Additionally, a broken piece of the last whorl of the atlantid shell was found, however, it is impossible to identify the specimen.
From the recent fauna of the Indo-West Pacific region, two species are now recognized and share morphological similarity (Wall-Palmer et al., 2019), namely A. turriculta and Atlanta vanderspoeli Wall-Palmer, Hegmann and Peijnenburg, 2019. In addition to a narrower distribution range than that in A. turriculata, A. vanderspoeli also has a broader apical angle. However, the spire of our specimen is covered in sediments that hinder more precise identification. With all other features, the specimen appears to be A. turriculata. Additional specimens from the same locality will facilitate more confident identification.
The Toukoshan Formation appears to be the third locality of Quaternary age of the Indo-West Pacific from where A. turriculata was mentioned, following the occurrence in the Pleistocene deposits of the Noto Peninsula, central Honshu (Shibata and Ujihara, 1983; Shibata, 1986) and Okinawa (Shibata and Ujihara, 1983). This species can be quickly identified by its high and narrow spire and relatively small shell. The current specimen of A. turriculata is the first heteropod fossil recorded from Taiwan.
Order PTEROPODA Cuvier, 1804
Superfamily CAVOLINIOIDEA Gray, 1850 (1815)
Family CRESEIDAE Rampal, 1973
Genus CRESEIS Rang, 1828
Type species. Creseis acicula (Rang, 1828), type by subsequent designation.
Creseis acicula (Rang, 1828)
Figure 2B–E
*1828 Cleodora (Creseis) acicula Rang, p. 318, pl. 17, fig. 6.
.1912 Clio (Creseis) acicula (Rang); Yamakawa and Ishikawa, p. 2, pl. I, figs. 1a, 1b.
.1983 Creseis acicula acicula (Rang); Shibata and Ujihara, p. 159, pl. 44, fig. 1.
.1984 Creseis acicula forma acicula (Rang); Shibata, p. 78, pl. 23, figs. 9, 10.
.1990 Creseis acicula forma acicula (Rang); Ujihara, Shibata, and Saito, p. 312, pl. 1, fig. 1.
.1996 Creseis acicula forma acicula (Rang); Ujihara, p. 774, fig. 3.5.
.1999 Creseis acicula (Rang); Janssen, p. 186.
.2007 Creseis clava (Rang); Janssen, p. 68, fig. 7; pl. 2, figs. 9, 10; pl. 23, figs. 7, 8.
.2012 Creseis clava (Rang); Janssen and Grebneff, p. 20.
Material. Miaoli County, Houlong Township; Toukoshan Formation, Late Pleistocene; 4 specimens, ASIZF0101022–025.
Description. Shell elongated, needle-like. Gradually increase in width towards aperture. Possible uneven curvature along length. No additional structures on shell. Aperture circular. Embryonic shells absent in examined samples.
Remarks. This species appears to be the most abundant within the Toukoshan Formation (Table 1), accounting for more than 60 specimens. The preservation of this species is poor, and in almost all individuals protoconch is not preserved. Previously, the absence of an embryonic shell made it doubtful to assign a specific species name (Ujihara et al., 1990; Janssen, 1999), and identification was roughly done based on the shell outline (Janssen, 2007). However, presently, only three living species are recognized (WoRMS, 2025). Among them, C. acicula appears distinct by its slender, straight, and long shell. Within the Toukoshan formation, shells of Scaphopoda of similar size and shape can be distinguished from C. acicula by having non-transparent, thicker shells of white color with a glossy surface.
The stratigraphic range of C. acicula in Cenozoic sediments of the western Pacific starts from the Miocene in Japan (Shibata, 1980; Ujihara et al., 1990) and Fiji (Janssen, 1999; Janssen and Grebneff, 2012). Very abundant C. acicula was described from multiple Pliocene formations in Japan (Shibata and Ishigaki, 1981; Shibata, 1984, 1986; Ujihara et al., 1990; Ujihara, 1996), and maintained abundance trend until Pleistocene (Shibata and Ishigaki, 1981; Shibata and Ujihara, 1983; Shibata, 1984).
Genus STYLIOLA Gray, 1847
Type species. Cleodora recta Blainville, 1827 accepted as Styliola subula (Quoy and Gaimard, 1827), type by monotypy.
Styliola subula (Quoy and Gaimard, 1827)
Figure 2F
*1827 Cleodora subula Quoy and Gaimard, p. 233–234, pl. 8D, figs. 1–3.
1912 Clio (Styliola) subula (Quoy and Gaimard); Yamakawa and Ishikawa, p. 4, pl. I, figs. 4a, 4b.
.1972 Creseis acicula (Quoy and Gaimard, non Creseis acicula (Rang, 1828)); Noda, p. 481, pl. 57, figs. 1–4.
.1983 Styliola subula (Quoy and Gaimard); Shibata and Ujihara, p. 161, pl. 44, fig. 6.
.1984 Styliola subula (Quoy and Gaimard); Shibata, p. 79, pl. 24, figs. 8, 9.
.1989 Styliola subula (Quoy and Gaimard); Janssen, p. 32, pl. 5, figs. 13–19; pl. 6, figs. 1-9.
.1990 Styliola subula (Quoy and Gaimard); Ujihara, Shibata, and Saito, p. 313, pl. 1, fig. 2.
.1996 Styliola subula (Quoy and Gaimard); Ujihara, p. 774, figs. 3.6–3.9.
.1999 Styliola subula (Quoy and Gaimard); Janssen, p. 180, pl. I, figs. 1a, 1b.
.2007 Styliola subula (Quoy and Gaimard); Janssen, p. 73, pl. 3, figs. 1–4; pl. 24, figs. 2, 3.
.2012 Styliola subula (Quoy and Gaimard); Janssen and Grebneff, p. 20.
Material. Miaoli County, Houlong Township; Toukoshan Formation, Late Pleistocene; 1 specimen, ASIZF0101026.
Description. Shell straight and conical, slightly curved. Barely visible longitudinal groove become more apparent closer to aperture. Aperture broken. Depression made by groove slightly compress circular transverse section. Protoconch elliptical and pointed. Protoconch tip of single specimen broken, visually divided from postembryonic shell by circular groove (depression).
Remarks. This species is represented by only a single specimen of a small size (2.3 mm). Styliola subula can be easily identified by having a groove that runs across the whole shell from the apex towards the aperture, and the shell being wide starting from the contraction at the protoconch.
In the western Pacific sediment sequences, Styliola subula appeared during the Miocene in southern Australia (Janssen, 1989), Indonesia (Janssen 1999), and southern and central Japan (Shibata, 1986; Ujihara et al., 1990). During the Pliocene-Pleistocene, the distribution of this species ranged from Indonesia (Janssen, 1999; Janssen and Grebneff, 2012) and the Philippines (Janssen, 2007) to central Honshu, on both western Noto and eastern Boso Peninsulas (Noda, 1972; Shibata and Ishigaki, 1981; Shibata and Ujihara, 1983; Shibata, 1984; Ujihara et al., 1990; Ujihara, 1996).
Family CAVOLINIIDAE Gray, 1850 (1815)
Subfamily CAVOLINIINAE Gray, 1850 (1815)
Genus CAVOLINIA Abildgaard, 1791
Type species. Cavolinia natans Abildgaard, 1791 accepted as Cavolinia tridentata (Forsskål, 1775), type by monotypy.
Cavolinia globulosa Gray, 1850
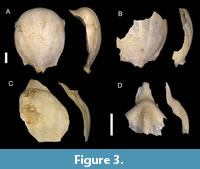
Figure 3A–B
*1850 Cavolinia globulosa Gray, p. 8.
1912 Cavolinia (s. str.) globulosa Gray; Yamakawa and Ishikawa, p. 20, pl. VI, figs. 12a-12c.
.1983 Cavolinia globulosa Gray; Shibata and Ujihara, p. 164, pl. 45, fig. 3.
.1984 Cavolinia globulosa Gray; Shibata, p. 85, pl. 26, figs. 1, 2.
.1986 Cavolinia globulosa Gray; Shibata, Ishigaki, and Ujihara, p. 50, pl. 8, fig. 9.
 Material. Miaoli County, Houlong Township; Toukoshan Formation, Late Pleistocene; 2 specimens, ASIZF0101027–028.
Material. Miaoli County, Houlong Township; Toukoshan Formation, Late Pleistocene; 2 specimens, ASIZF0101027–028.
Description. Shell comparatively big in size. Ventral part absent in all specimens. Apical spine narrow and bended dorsally, but mostly broken. Lateral spines small, directed to caudal side. Distance between their tops smaller than shell width in middle. Dorsal part mostly rounded, except for caudal part. Five dorsal ribs can be distinguished. Lateral ribs wide, middle ribs separated by not deep grooves. Dorsal apertural lip bent ventrally, creating hump.
Remarks. In our samples, this species is represented by only the dorsal parts of two individuals. Shells of Cavolinia globulosa differ from other Cavolinia species by having small lateral spines almost oriented backwards, and the shell having its greatest width at its middle part. Shibata (1984) noted that his fossil specimens differ slightly from the recent counterpart by having a more curved dorsal lip and a more ovate-elongated shape.
Within the West Pacific region as a fossil this species was recorded only from deposits in Japan, ranging from the Okinawa islands (Shibata and Ujihara, 1983) up to the central Honshu (e.g., Shibata et al., 1986), with the first appearance during the Miocene (Shibata, 1984) and ranges from Neogene to Quaternary (Shibata and Ishigaki, 1981; Shibata and Ujihara, 1983; Shibata, 1984; Shibata et al., 1986).
Cavolinia cf. gibbosa d’Orbigny, 1835
Figure 3C
?1912 Cavolinia (s. str.) gibbosa (d’Orbigny); Yamakawa and Ishakawa, p. 21, pl. VI, figs. 13a–13c.
Material. Miaoli County, Houlong Township; Toukoshan Formation, Late Pleistocene; 1 specimen, ASIZF0101029.
Description. Shell comparatively big, rounded. On the dorsal side at least five radial ribs present, all of the same size. Outer hump absent. Rostrum appear wide and flat.
Remarks. In our material, only a single fragment was recovered, representing only the dorsal part with a rostrum. Based on the relatively big size of the shell, wide outer hump and inner hump being not strong, it is possible current specimen belongs to the species Cavolinia gibbosa (d’Orbigny, 1835). But due to the lack of other important features, we cannot assign more precise name. It differs from C. globulosa from the Toukoshan Formation by not having distinct outer hump.
Fossil specimens of this species were recovered only from the Japanese Neogene and Quaternary sediments. The oldest records appear to be incomplete shells of Cavolinia gibbosa from the Chiba Prefecture of the “Neogene” sediments (Yamakawa, 1912). Shibata and Ujihara (1983) and Shibata (1986) reported this species from multiple formations in central Honshu of the Pleistocene age. Additionally, species reported as Cavolinia okinawana Noda, 1972 bears high resemblance to C. gibbosa, however, more detailed comparison of type specimens is required.
Cavolinia cf. tridentata (Forsskål, 1775)
Figure 3D
Material. Miaoli County, Houlong Township; Toukoshan Formation, Late Pleistocene; 1 specimen, ASIZF0101030.
Description. Dorsal lip triangular in shape, with rather broad rostrum. Small lip gutter, but no constriction. One evident central dorsal radial rib does not continue to the rostrum. Outer hump not steep. Inner hump cannot be observed.
Remarks. This specimen is hard to identify due to the limited features that are preserved. The only preserved dorsal lip has one distinct feature that represents a central dorsal rib that ends before outer hump and does not continue to the rostrum. This characteristic can also be observed in Cavolinia tridentata (Forsskål, 1775) and Cavolinia shibatai Janssen, 2007. However, comparing the Toukoshan specimen to Pagasinan species (Janssen, 2007), C. shibatai has no inner hump, and C. tridentata has a comparatively wider rostrum. Additionally, our studied specimen also resembles Diacavolinia triangulata Van der Spoel, Bleeker and Kobayasi, 1993 in the lateral view; however, in the original description, the feature of dorsal ribs was not specified. The main difference between Cavolinia and Diacavolinia Van der Spoel, 1987 species lies in the development of the shell, with Cavolinia species retaining the protoconch. Combining that our specimen lacks the caudal part and the rostrum is long and not wide, we decided to assign it to the species Cavolinia tridentata.
Genus DIACAVOLINIA Van der Spoel, 1987
Type species. Hyalaea longirostris Blainville, 1821 accepted as Diacavolinia longirostris (Blainville, 1821), type by original designation.
Diacavolinia longirostris (Blainville, 1821)
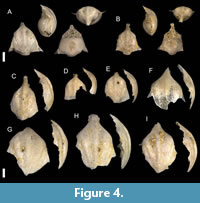
Figure 4
*1821 Hyalaea longirostris Blainville, p. 81.
1912 Cavolinia (s. str.) longirostris (Blainville); Yamakawa and Ishikawa, p. 19, pl. VI, figs. 11a–11c.
.1983 Cavolinia longirostris longisrostris (Blainville); Shibata and Ujihara, p. 164, pl. 45, fig. 5.
.1984 Cavolinia longirostris forma longirostris (Blainville); Shibata, p. 86, pl. 25, figs. 5, 6.
.1993 Diacavolinia longirostris (Blainville); Van der Spoel, Bleeker. And Kobayashi, p. 132, fig. 3.
 Material. Miaoli County, Houlong Township; Toukoshan Formation, Late Pleistocene; 9 specimens, ASIZF0101031–039.
Material. Miaoli County, Houlong Township; Toukoshan Formation, Late Pleistocene; 9 specimens, ASIZF0101031–039.
Description. Shell medium in size, triangular from the dorsal view. The greatest width of the shell at the lateral spines. Ventral part moderately swollen and convex. Growth lines visible along all ventral part. Dorsal side smooth and glossy, slightly convex with five longitudinal ribs, three of them in the center the most evident. Middle longitudinal rib continues to the rostrum. Outer hump absent. Lateral ribs wide. Growth lines evident on the dorsal side. Dorsal lip bent ventrally and considerably protrudes over the ventral part of the shell. Second lock mechanism present. Apical spine shaded, leaving a caudal joint.
Remarks. Representatives of this species are among the most abundant in the studied locality (Table 1). All studied specimens align with the so-called Diacavolinia longirostris group, which has been the subject of taxonomic debate (Burridge et al., 2019) and includes D. longirostris and closely related taxa. The slight variation in form and shape of our specimens can be observed. Some of them, which are represented by only the dorsal part of the shell (Figure 4E–I), differ from others only by the dorsal lip that is only slightly bent to the ventral side compared to the rest of the specimens (Figure 4A–D). These specimens may belong to the species Diacavolinia vanutrechti Van der Spoel, Bleeker and Kobayasi, 1993. Another difference lies in two specimens with a complete shell, with one specimen being more convex with lateral spines bent outwards (Figure 4A), which is characteristic of D. longirostris. The other specimen (Figure 4B) has a generally smaller and less convex shell with straight lateral spines. Generally, these variation lies within the D. longirostris, but the incompleteness of most of the specimens and a small number of individuals hinder a more precise identification. At the same time, recent phylogenetic and morphological study (Burridge et al., 2019) suggests that D. longirostris might be restricted only to the Atlantic Ocean.
Within the Neogene and Quaternary deposits in the Indo-West Pacific region, this species was reported only from Japan, with the southernmost occurrence in the Okinawa islands (Noda, 1972; Shibata and Ujihara, 1983) up to the central Honshu (Yamakawa, 1912; Shibata and Ujihara, 1983). The earliest occurrence of Diacavolinia longirostris is of Miocene from the Shizuoka Prefecture (Shibata, 1984) and noted to be abundant in the early Pleistocene deposits (Shibata and Ishigaki, 1981).
Key distinguishing features include the degree of convexity in the dorsal and ventral parts, the shape and characteristics of the rostrum, the configuration of lateral spines, and the presence of outer and inner humps (Van der Spoel et al., 1993). Most of the Diacavolinia longirostris specimens from the Toukoshan Formation are poorly preserved, with broken dorsal parts that obscure these critical diagnostic features. Some variation in the dorsal lip was observed, with certain specimens resembling D. vanutrechti, which is characterized by a less ventrally tilted dorsal lip. However, due to insufficient preservation, confidently distinguishing between these species is challenging. Consequently, all these specimens are provisionally identified as D. longirostris.
Diacavolinia cf. bandaensis Van der Spoel, Bleeker, and Kobayasi, 1993
Figure 5A
*1993 Diacavolinia bandaensis Van der Spoel, Bleeker and Kobayasi; p. 144, figs. 19A–B, pl. II, fig. 24.
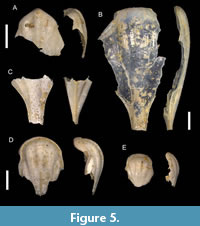 Material. Miaoli County, Houlong Township; Toukoshan Formation, Late Pleistocene; 1 specimen, ASIZF0101040.
Material. Miaoli County, Houlong Township; Toukoshan Formation, Late Pleistocene; 1 specimen, ASIZF0101040.
Description. Dorsal side of the shell smooth and glossy, not convex. Five longitudinal ribs, three of them in the center the most evident, lateral spines wide and continue to lateral spines. Dorsal lip triangular in shape, with rostrum. Outer hump evident. Rostrum broken, but the basement suggests that it rater broad. Apertural margin of the dorsal side triangular in shape with central radial rib being the longest. Growth lines evident on the dorsal side.
Remarks. The single broken specimen representing only part of the dorsal side differs from other cavoliniids from the Toukoshan Formation. The presence of the hump that comes before rostrum, makes it different from Diacavolinia longirostris. To our knowledge, this species has no fossil record and current specimen may represent the first Pleistocene occurrence of D. bandaensis.
This species was first described by Van der Spoel et al. (1993) from the Banda Sea in Indonesia. They noted that this species is closely related to Diacavolinia angulosa (Gray, 1850) and Diacavolinia pacifica Van der Spoel, Bleeker, and Kobayasi, 1993, but the latter two species are smaller, and the outer hump has slightly arched shape compared to more triangular in D. bandaensis.
Subfamily DIACRIINAE Rampal, 2019
Genus DIACRIA Gray, 1840
Type species. Hyalaea trispinosa Blainville, 1821 accepted as Diacria trispinosa (Blainville, 1821), type by subsequent designation.
Diacria cf. philippinensis Janssen, 2007†
Figure 5B–C
*2007 Diacria philippinensis Janssen†; p. 100, pl. 7, fig. 3; pl. 8, fig. 6; pl. 25, fig. 1.
.2012 Diacria philippinensis Janssen†; Janssen and Grebneff, p. 24, figs. 16a–c.
Material. Miaoli County, Houlong Township; Toukoshan Formation, Late Pleistocene; 2 specimens, ASIZF0101041–042.
Description. Dorsal part of the compressed shell not swollen with three evident central ribs. Lateral ribs wide, mostly broken. The remaining part of the caudal spine indicates its long length. Dorsal aperture edge almost straight, broken.
Remarks. From the studied locality, this species is represented by two fragmentary specimens, comprising one with preserved only the dorsal part, and one with the base of the apical spine. The characteristic feature of the latter specimen is the presence of three radial dorsal ribs and remains of the apical spine, resembling species Diacria philippinensis, but lacking transverse ornamentation at the dorsal apertural margin. However, species Diacria trispinosa (Blainville, 1821) (specifically, forma trispinosa) also possesses similar features. Unlike D. philippinensis, shells of D. trispinosa are bigger, less convex, and widen sharply in the posterior part at the base of the apical spine. However, our specimens lack important characteristics of the ventral side and lateral spines that hinder more precise identification.
The species Diacria philippinensis was described based on material from the type locality in Luzon, Philippines of Pliocene age (Piacenzian) (Janssen, 2007). Occurrence from the sediments of the same age is known in the Viti Levi Island, Fiji Archipelago (Janssen and Grebneff, 2012). It is worth noting that both records differ by the convexity and the shape of lateral spines; in Fiji’s specimens are placed wide apart. Additionally, Janssen (2007) mentioned that the apical spine is shed in all specimens from the Philippines and replaced by a septum. However, Fiji specimens (Janssen and Grebneff, 2012) retained their apical spine. Our studied material resembles shells described from the Fiji Archipelago.
Genus TELODIACRIA Rampal, 2019
Type species. Hyalaea quadridentata Blainville, 1821 accepted as Telodiacria quadridentata (Blainville, 1821), type by original designation.
Telodiacria quadridentata (Blainville, 1821)
Figure 5D–E
*1821 Hyalaea quadridentata Blainville, p. 81.
.1912 Cavolinia (Diacria) quadridentata (Blainville); Yamakawa and Ishikawa, p. 18, pl. VI, figs. 10a–10c.
.1983 Diacria quadridentata erythra Van der Spoel; Shibata and Ujihara, p. 163, pl. 44, figs. 10, 11.
Material. Miaoli County, Houlong Township; Toukoshan Formation, Late Pleistocene; 2 specimens, ASIZF0101043–044.
Description. Shell medium in size (3 mm), rather higher than wide. Ventral part absent in examined specimens. Dorsal side is moderately convex. Thickened dorsal apertural margin has brownish color in inner and outer sides, closer to aperture margin transverse crests are present. Five dorsal radial ribs separated by distinct grooves, three ribs in the middle are the most distinct. Apical spine absent. Lateral spines not recognizable as lateral parts of the shell broken.
Remarks. This species from the Toukoshan Formation comprises only the dorsal parts of two specimens. Based on the Yamakawa (1912) record from the Chiba Prefecture in central Japan, this species started its appearance in the West Pacific during the Neogene. However, the only other record from Pleistocene deposits is Shibata (1986). Shibata and Ujihara (1983) identified this taxon as Diacria quadridentata erythra Van der Spoel, 1971, but their specimen differs from D. q. erythra (currently accepted as Diacria erythra Van der Spoel, 1971) by being comparatively smaller with less distinct radial ribs, and, thus, represents T. qudridentata.
DISCUSSION
Taxonomic Composition
The Toukoshan Formation has revealed a diverse yet relatively sparse fauna of holoplanktonic gastropods. This assemblage predominantly comprises at least nine pteropod taxa and a single heteropod species. Among these, the most abundant is Creseis acicula, whose fragile shells are well-preserved but lacking embryonic parts. The next most common species is Diacavolinia longirostris. However, the species affiliation needs to be confirmed by a more comprehensive review of this species within the Cenozoic sediments in the Indo-West Pacific, as it was recently suggested that this species is restricted only to the Atlantic Ocean (Burridge et al., 2019).
Other pteropod species are represented by only a few fragmentary shells with low abundance. Notably, the typically abundant and common species from the Pleistocene deposits (e.g., Shibata, 1984), Styliola subula, is nearly absent from our study. Atlanta turriculata is the only heteropod species found in the Toukoshan Formation. However, this single specimen is the first record of heteropod from the Pleistocene deposits in Taiwan.
Placing the Toukoshan Holoplanktonic Fauna in Time and Place
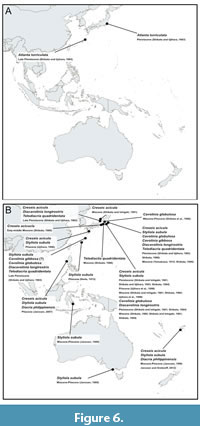 The fossil record indicates that the modern holoplanktonic fauna in the West Pacific began to establish during the Pliocene-Pleistocene, with most species having their first appearance during the Miocene (Bernasconi, 1982; Shibata, 1986; Wang et al., 1997). The most comprehensive fossil records of holoplanktonic gastropods are from Japan, where deposits of various ages have been reported (Figure 6). However, the nearly absence of records from other parts of the West Pacific strongly hinders our understanding of the origin and development of the fauna during the entire Neogene and Quaternary in the region, although this limitation can also be applied to other marine faunas (Lin et al., 2021; Mediodia et al., 2024). Further paleontological investigations could significantly enhance our understanding of holoplanktonic mollusk distributions.
The fossil record indicates that the modern holoplanktonic fauna in the West Pacific began to establish during the Pliocene-Pleistocene, with most species having their first appearance during the Miocene (Bernasconi, 1982; Shibata, 1986; Wang et al., 1997). The most comprehensive fossil records of holoplanktonic gastropods are from Japan, where deposits of various ages have been reported (Figure 6). However, the nearly absence of records from other parts of the West Pacific strongly hinders our understanding of the origin and development of the fauna during the entire Neogene and Quaternary in the region, although this limitation can also be applied to other marine faunas (Lin et al., 2021; Mediodia et al., 2024). Further paleontological investigations could significantly enhance our understanding of holoplanktonic mollusk distributions.
Pteropods in South China Sea are thought to have originated in the open waters of the western Pacific and subsequently dispersed into the sea during the Late Quaternary via the Bashi Strait, located between the southern tip of Taiwan and Luzon Island (Wang et al., 1997). It is plausible that the Toukoshan fauna in the northern Taiwan Strait shares this origin. Alternatively, the connection between the Taiwan Strait and the northern West Pacific might have been facilitated by the China Coastal Current (Ho et al., 2022).
Nearly all pteropod species from the Toukoshan Formation are extant and have been documented in Indo-West Pacific deposits since the Miocene (Figure 6). Exceptions include Diacria cf. philippinensis and Diacavolinia cf. bondaensis. The known stratigraphic range of D. philippinensis is the Pliocene (Janssen, 2007; Janssen and Grebneff, 2012), suggesting it likely evolved during the Miocene but lacks living representatives. As the present study identified specimens as D. philippinensis, its stratigraphic range has been extended into the Pleistocene, being the first recorded Quaternary occurrence of this species. In contrast, D. bandaensis lacks fossil records from the Pacific region and is only known as a living species (Van der Spoel et al., 1993; Burridge et al., 2019). This study provides a potential first Quaternary record for this taxon. However, due to the high morphological similarity between D. bandaensis and Diacavolinia angulosa –a species widely reported in Indo-West Pacific Neogene and Quaternary deposits–a thorough comparison with type specimens and new fossil records could reveal additional occurrences of this species.
The holoplanktonic gastropods from the Toukoshan Formation predominantly represent tropical-subtropical species, which are characteristic of the circumglobal warm-water region (Be and Gilmer, 1977, and references therein; Wall-Palmer et al., 2016). Most of the pteropod species live in wide range of temperatures ranging from 15 to 28°C (Chen and Be, 1964; Van der Spoel, 1967; Williams, 1972), with an exception of T. quadridentata that can tolerate only narrow range of warm water temperature (Williams, 1972). Based on the radiocarbon dating (e.g., Wang and Peng, 1990; Ho et al., 2022), the age of the Toukoshan Formation correlated to the MIS 3, and characterized by milder glacial temperatures (Ho et al., 2022). The cooler conditions may influence the decline in abundance and number of species of warm-water pteropods at that time. This is possibly reflected in the presence of a high number of eurythermal C. acicula, along with a low abundance of stenothermic T. quadridentata. The contrast in number of taxa is observed from the Szekou Formation, another locality studied by us, that are considered to represent warmer MIS 5e, and several formations in central Japan (Shibata and Ujihara, 1983), yielding greater diversity and occurrence of both pteropods and heteropods. A similar situation was observed in cores from the Red Sea (Almogi-Labin, 1982), where diversity peaked during interglacial stage 5, in contrast to the reduction of diversity towards glacial maxima at stage 2. However, the discrepancies in the interpreted age of the Toukoshan Formation complicate precise correlations with other records, permitting only generalized comparisons.
During the deposition, a shallow-water condition is proposed, with a depth of around 60 m (Chow, 1980). The holoplanktonic fauna reported in the present paper is epipelagic and typically inhabits depths above 200 m (Tesch, 1948; Wormelle, 1962), usually with pronounced diurnal migration (Bé and Gilmer, 1977; Wall-Palmer et al., 2018). An exception may be represented by C. acicula, which is known to inhabit only the upper 50 m (Myers, 1968). Similarly, the higher abundance of this species compared to others indicates conditions shallower than 100 m and may support previous interpretations of depositional conditions for the fossil-bearing layer (Chow, 1980). However, it should also be noted that, due to their buoyant shells, pteropods may have been transported over long distances at the surface before sinking and being deposited, potentially in waters shallower than their original habitat.
The current study represents a resumption of studies on fossil mollusks in Taiwan and aims to contribute to the scarce records of holoplanktonic gastropods in the West Pacific. It might improve our understanding of their development and establishment in this area after its dispersal from the Tethys residual basin (Li et al., 2020). A thorough investigation on fossil holoplanktonic gastropods is planned within Taiwan and adjacent territories.
ACKNOWLEDGMENTS
We thank C.-H. Hsu for his valuable comments and corrections for the geological description of the studied locality. We are also grateful reviewers for their constructive suggestions, which greatly improved the quality of the manuscript. We acknowledge H.-W. Liu and members of the Marine Paleontology Laboratory, Biodiversity Research Center, Academia Sinica, for their support during fieldwork and sample preparation. This study was supported by the Intramural funding of Academia Sinica, Taipei, Taiwan and National Science and Technology Council, Taiwan (Grant No. 112-2116-M-001-017-MY3).
REFERENCES
Abildgaard, P.C. 1791. Nyere efterretning om det skaldyr fra Middelhavet, som Forskål har beskrevet under navn af Anomia tridentata. Skrifter af Naturhistorie-Selskabet, 1(2):171–175. (In Danish)
Almogi-Labin, A. 1982. Stratigraphic and paleoceanographic significance of Late Quaternary pteropods from deep-sea cores in the Gulf of Aqaba (Elat) and northernmost Red Sea. Marine Micropaleontology, 7(1):53–72.
https://doi.org/10.1016/0377-8398(82)90015-9
Ando, Y. 2011. Oligocene pteropods (Gastropoda: Thecosomata) from the Kishima Formation. Revista Mexicana de Ciencias Geológicas, 28(2): 245–253.
Ando, Y., Ujihara, A., and Ichihara, T. 2009. First occurrence of Paleogene pteropods (Gastropoda; Thecosomata) from Japan. The Journal of the Geological Society of Japan, 115(4):187–190.
https://doi.org/10.5575/geosoc.115.187
Bé, A.W.H. and Gilmer, R.W. 1977. A Zoogeographic and taxonomic Review of Euthecosomatous Pteropoda, p. 733–808. In Ramsay, A.T. (ed.), Oceanic Micropaleontology. Academic Press, London.
Beccari, V., Almogi-Labin, A., Basso, D., Panieri, G., Makovsky, Y., Hajdas, I., and Spezzaferri, S. 2023. Late Holocene pteropod distribution across the base of the south-eastern Mediterranean margin: the importance of the > 63 µm fraction, Journal of Micropalaeontology, 42, 13-29.
https://doi.org/10.5194/jm-42-13-2023
Bednaršek, N., Harvey, C.J., Kaplan, I.C., Feely, R.A., and Možina, J. 2016. Pteropods on the edge: cumulative effects of ocean acidification, warming, and deoxygenation. Progress in Oceanography, 145:1–24.
https://doi.org/10.1016/j.pocean.2016.04.002
Bernasconi, M.P. and Ronna, E. 1982. The Thecosomatous Pteropods: a contribution toward the Cenozoic Tethyan paleobiogeography. Bollettino Della Societa Paleontologica Italiana, 21(2-3):211–222.
Blainville, H.M.D. de. 1821. Hyale, Hyalæa (Malacoz.), p. 65–83. In Cuvier F. (ed.), Dictionnaire des Sciences Naturelles. Levrault, Strasbourg and Le Normant, Paris. (In French)
Blainville, H.M.D. de. 1825–1827. Manuel de Malacologie et de Conchyliologie. Levrault, Paris. (In French)
Bouchet, P., Rocroi, J.P., and Frýda, J. 2005. Classification and nomenclator of gastropod families. Malacologia, 47(1–2):1–397.
Bouchet, P., Rocroi, J.P., Hausdorf, B., Kaim, A., Kano, Y., Nützel, A., Parkhaev, P., Schrödl, M., and Strong, E.E. 2017. Revised classification, nomenclator and typification of Gastropod and Monoplacophoran families. Malacologia, 61(1–2):1–526.
https://doi.org/10.4002/040.061.0201
Buccheri, G. 1984. Pteropods as climatic indicators in Quaternary sequences: A Lower-Middle Pleistocene sequence outcropping in Cava Puleo (Ficarazzi, Palermo, Italy). Palaeogeography, Palaeoclimatology, Palaeoecology, 45:75–86.
https://doi.org/10.1016/0031-0182(84)90110-x
Buccheri, G., Catalano, R., and Heezen, B. 1980. Pleistocene pteropod limestones dredged from the East Yucatan Scarf. Palaeogeography, Palaeoclimatology, Palaeoecology, 30:97–105.
https://doi.org/10.1016/0031-0182(80)90051-6
Chang, S.S.-L., Chiu, H.-T., Hsieh, S.-H., and Kuan, M.-Y. 1965. Case history of Tiechenshan gas field, Taiwan, China. Third Petroleum Symposium, ECAFE, 1–13. (In Chinese)
Chang, L.-S. 1948. The revision of stratigraphy categories of Taiwan (continued). Geological Review, 13:291–310. (In Chinese)
Chang, L.-S. 1955. The strata of Taiwan. Quarterly Journal of the Taiwan Bank, 7:26–49. (In Chinese)
Chang, J.-C., Teng, K.-H., and Liu, M.-C. 1998. A Geomorphological Study on River Terraces in Miaoli Hills. Geographical Research, 29:97–112.
https://doi.org/10.6234/JGR
Chen, C. 1969. Pteropods in the hot brines sediments of the Red Sea, p. 313–316. In Degens, E. T. and Ross, D. A. (eds.), Hot Brines and Recent Heavy Metal Deposits in the Red Sea. Springer, Berlin.
https://doi.org/10.1007/978-3-662-28603-6_31
Chen, C. and Be, A. W. H. 1964. Seasonal distributions of euthecosomatous pteropods in the surface waters of five stations in the western North Atlantic. Bulletin of Marine Science of the Gulf and Caribbean, 14(2):185–220.
https://doi.org/10.1016/0011-7471(65)90465-1
Chen, C. and Huang, C.-Y. 1990. Two new Oligocene pteropods with paleoenvironmental implications. Proceedings of the Geological Society of China, 33(3):223–238.
Chen, P.-H., Huang, T.-C., Huang, C.-Y., Jiang, M.-J., Lo, S.-L., and Kuo, C.-L. 1977. Paleomagnetic and coccolith stratigraphy of Plio-Pleistocene shallow marine sediments, Chuhuangkeng, Miaoli. Petroleum Geology of Taiwan, 14:219–240.
Chen, W.-S., Ridgway, K.D., Horng, C.-S., Chen, Y.-G., Shea, K.-S., and Yeh, M.-G. 2001. Stratigraphic architecture, magnetostratigraphy, and incised-valley systems of the Pliocene- Pleistocene collisional marine foreland basin of Taiwan. GSA Bulletin, 113:1249–1271.
https://doi.org/10.1130/0016-7606(2001)113<1249:samaiv>2.0.co;2
Cheng, Y.-M. 1981. Notes On Euthecosomatous Pteropods Clio sp. in the Oligocene formations of Northern Taiwan. Ti-Chih, 3:157–160. (In Chinese)
Chou, J.-T. 1973. Sedimentology and paleogeography of the Upper Cenozoic system of western Taiwan. Proceeding of the Geological Society of China, 16:111–143.
Chou, J.-T. 1992. Cenozoic formations of the foothills region in Western Taiwan. Species Publication of the Central Geological Survey, 6:85–134.
Chow, J. 1980. Late Neogene paleoenvironments of the Mucha section in Kaohsiung. Petroleum Geology of Taiwan, 17:89–98.
Cuvier, G. 1795. Second mémoire sur l’organisation et les rapports des animaux à sang blanc, dans lequel on traite de la structure des mollusques et de leur division en ordre, lu à la société d’Histoire Naturelle de Paris, le 11 prairial an troisième. Magazin Encyclopédique, Ou Journal Des Sciences, Des Lettres et Des Arts, 2:433–449. (In French)
Cuvier, G. 1804. Mémoire concernant l’animal de l’ Hyale, un nouveau genre de mollusques nus, intermédiaire entre l’ Hyale et le Clio, et l’établissement d’un nouvel ordre dans la classe des mollusques. Annales Du Muséum National d’Histoire Naturelle, 4(21):223–234. (In French)
Forskål, P. 1775. Descriptiones Animalium, Avium, Amphibiorum, Piscium, Insectorum, Vermium; quae in Itinere Orientali Observavit Petrus Forskål. Post Mortem Auctoris editit Carsten Niebuhr. Adjuncta est materia Medica Kahirina. Hafniae. (in Latin)
Gerhardt, S. and Henrich, R. 2001. Shell preservation of Limacina inflata (Pteropoda) in surface sediments from the Central and South Atlantic Ocean: A new proxy to determine the aragonite saturation state of water masses. Deep-Sea Research Part I: Oceanographic Research Papers, 48:2051–2071.
https://doi.org/10.1016/s0967-0637(01)00005-x
Golikov, A.N. and Starobogatov, Y.I. 1975. Systematics of prosobranch gastropods. Malacologia, 15(1):185–232.
Gray, J.E. 1847. A list of the genera of recent Mollusca, their synonyma and types. Proceedings of the Zoological Society of London, 15:129–219.
Gray, J.E. 1850. Catalogue of the Mollusca in the Collection of the British Museum. Part II. Pteropoda. Edward Newman, London.
Hallenberger, M., Reuning, L., Takayanagi, H., Iryu, Y., Keul, N., Ishiwa, T., and Yokoyama, Y. 2022. The pteropod species Heliconoides inflatus as an archive of late Pleistocene to Holocene environmental conditions on the Northwest Shelf of Australia. Progress in Earth and Planetary Science 9, 49.
https://doi.org/10.1186/s40645-022-00507-1
Hayasaka, I. 1933. Notes on the Toukoshan Formation, Taiwan. Journal of Geology, 40(477):364–366. (In Japanese)
Hayasaka, I. 1934. Geology and fossil mollusks from Peisa village, Shinchu area. Proceedings of Earth Sciences in Taiwan, 5:37–42. (In Japanese)
Heh, K., and Hsiao, P.-T. 1957. Geology of the Tunghsiao anticline, Miaoli District. Symposium of Petroleum Geology of Taiwan, 128–141. (In Chinese)
Ho, S.-L., Wang, J.-K., Lin, Y.-J., Lin, C.-R., Lee, C.-W., Hsu, C.-H., Chang, L.-Y., Wu, T.-H., Tseng, C.-C., Wu, H.-J., John, C.-M., Oji, T., Liu, T.-K., Chen, W.-S., Li, P., Fang, J.-N., and Lin, J.-P. 20220. Changing surface ocean circulation caused the local demise of echinoid Scaphechinus mirabilis in Taiwan during the Pleistocene-Holocene transition. Scientific Reports, 12(8204):1–9.
https://doi.org/10.1038/s41598-022-11920-3
Hu, C.-H. 1978. Studies on Ostracodes from the Toukoshan Foramtion (Pleistocene), Miaoli District, Taiwan. Petroleum Geology of Taiwan, 15:127–166.
Hu, C.-H. 1981. New ostracod faunas from the Maanshan Mudstone of Hengchun Peninsula, southern Taiwan. Pertoleum Geology of Taiwan, 81:81–109.
Hu, C.-H. 1984. Cenozoic Crab Fossils from Taiwan Island. Petroleum Geology of Taiwan, 20:181–198.
Hu, C.-H. 1992. Miaoli County Longgang local Tongxiao layer (Pleistocene), p. 689–753. In Hu, C.-H. (ed.), Mollusk Fauna of Taiwan. National Museum of Natural Science, Taichung, Taiwan.
Hu, C.-H. (ed.). 1991−1995. Mollusk Fauna of Taiwan. National Museum of Natural Science, Taichung, Taiwan.
Huang, C.-Y. 1988. Forameniferal paleoecology of a Late Pleistocene lagoonal sequence of the Szekou Formation Peninsula. Proceeding of the Geological Society of China, 31:181–206.
Huang, C.-Y., Masuda, K., and Wu, L.-C. 1990. Miocene Pelecypoda in the Western Foothills of Northern Taiwan (Part 1: Stratigraphy, Faunal Occurences and Paleoceanographic Significance). Bulletin of the National Museum of Natural Science, 2:119–139.
Huang, T. 1984. Planktic foraminiferal biostratigraphy and datum planes in the Neogene sedimentary sequence in Taiwan. Palaeogeography, Palaeoclimatology, Palaeoecology, 46:97–106.
Janssen, A.W. 1989. Cainozoic Pteropoda from Australia. Scripta Geologica, 91:1–76.
Janssen, A.W. 1999. Euthecosomatous gastropods (Mollusca Heterobranchia) from Buton, Fiji: systematics, biostratigraphy. Geologie En Mijnbouw, 78:179–189.
Janssen, A.W. 2007. Holoplanktonic Mollusca (Gastropoda: Pterotracheoidea, Janthinoidea, Thecosomata and Gymnosomata) from the Pliocene of Pangasinan (Luzon, Philippines). Scripta Geologica, 135:29–177.
Janssen, A.W. and Grebneff, A. 2012. Notes on the systematics, morphology and biostratigraphy of fossil holoplanktonic Mollusca, 22. Further pelagic gastropods from Viti Levu, Fiji Archipelago. Basteria, 76(1–3):15–30.
Janssen, A.W. and Goedert, J.L. 2016. Notes on the systematics, morphology and biostratigraphy of fossil holoplanktonic Mollusca, 24. First observation of a genuinely Late Mesozoic thecosomatous pteropod. Basteria, 80(1–3):59–63.
Janssen, A.W. and Peijnenburg, K.T.C.A. 2017. An overview of the fossil record of Pteropoda (Mollusca, Gastropoda, Heterobranchia). Cainozoic Research, 17(1):3–10.
Janssen, A.W., Bush, S.L., and Bednaršek, N. 2019. The shelled pteropods of the northeast Pacific Ocean (Mollusca: Heterobranchia, Pteropoda). Zoosymposia, 13:305–346.
https://doi.org/10.11646/zoosymposia.13.1.22
Kanno, S. and Chung, C.-T. 1975. Tertiary formations and their molluscan faunas from the Central Range and foothills areas of northern Taiwan. Geology and Palaeontology of Southeast Asia, 15:363–391.
Keul, N., Peijnenburg, K.T.C.A., Andersen, N., Kitidis, V., Goetze, E., and Schneider, R.R. 2017. Pteropods are excellent recorders of surface temperature and carbonate ion concentration. Scientific Reports, 7(1):12645.
https://doi.org/10.1038/s41598-017-11708-w
Kotaka, T. 1973. Faunal analysis of the Upper Miaoli Group in the Tunghsiao area, Miaoli province, Formosa, China: Part 1: Statistic analysis. Science Reports of the Tohoku University. 2nd Series, Geology. Special Volume, 6:81–96.
Ladd, H.S. 1934. Geology of Vitilevu, Fiji. Bernice P. Bishop Museum Bulletin, 119:1–263.
Lee, Q.-L. 2000. Chronostratigraphy and Sedimentary Environment of the Toukoshan Formation in the Baishatun Area, Miaoli. M.S. Thesis. Institute of Geophysics, National Central University, Taoyuan, Taiwan.
Lee, C.-L., Huang, T., Shieh, K.-S., and Chen, Z.-H. 2002. The chronostratigraphy and sedimentary environments of the Toukoshan Fm. in Beishatun area, Miaoli. Annual Report of Central Geological Survey, MOEA, 1999–2000, 17–20.
Lesueur, C.A. 1817. Mémoire sur deux nouveaux genres de mollusques, Atlante et Atlas. Journal de Physique, de Chimie et d’Histoire Naturelle, 85:390–393. (in French)
Li, X.-F., Li, G.-B., Garvie, C.L., Wang, T.-Y., and Zhao, J. 2020. First report of the early Eocene pteropods from the Zhepure Formation in Yadong, southern Tibet, China. Journal of Paleontology, 94(5):819–828.
https://doi.org/10.1017/jpa.2020.28
Liew, P.-M. 1988. Quaternary stratigraphy in Western Taiwan: palynological correlation. Proceedings of the Geological Society of China, 31(1):169–180.
Lin, A.-T., Watts, A.B., and Hesselbo, S.P. 2003. Cenozoic stratigraphy and subsidence history of the South China Sea margin in the Taiwan region. Basin Research, 15(4):453–478.
https://doi.org/10.1046/j.1365-2117.2003.00215.x
Lin, C.-C. 1935. Stratigraphical studies on the younger Tertiary and Pleistocene formations of the Toyohara district, Taityu prefecture, Taiwan. Memoirs of the Faculty of Science and Agriculture, Taihoku University, 13(3):13–30.
Lin, C.-C. 1963. Quaternary Geology of Taiwan. Taiwan Wen-Hsien, 14(1):1–53.
Lin, C.-H., Chien, C.-W., Lee, S.-W., and Chang, C.-W. 2021. Fish fossils of Taiwan: a review and prospection. Historical Biology, 33(9):1362–1372.
https://doi.org/10.1080/08912963.2019.1698563
Lin, C.-H., Ou, H.-Y., Lin, C.-Y., and Chen, H.-M. 2022. First Skeletal Fossil Record of the Red Seabream Pagrus major (Sparidae, Perciformes) from the Late Pleistocene of Subtropical West Pacific, Southern Taiwan. Zoological Studies, 61:1–14.
https://doi.org/10.6620/ZS.2022.61-10
Lin, J.-P., Lin, C.-H., Chu, W.-C., and Chang, C.-H. 2021. Introduction to the special issue about new advances on stratigraphy and paleontology in Taiwan. Terrestrial, Atmospheric and Oceanic Sciences, 32(6):1047–1050.
https://doi.org/10.3319/TAO.2021.12.29.01
Liu, S.-H., Hebenstreit, R., and Böse, M. 2022. Late Quaternary landform evolution and sedimentary successions in the Miaoli Tableland, northwestern Taiwan. E and G Quaternary Science Journal, 71(1):1–22.
https://doi.org/10.5194/egqsj-71-1-2022
Masuda K. and Huang C.-Y. 1990. Check List of Tertriary and Quarteenary Mollusca in Taiwan. Acta Geologica Taiwanica, 28:31–76.
Mediodia, D., Castro, A., Tablizo, M., Policarpio, D., Calvelo, J., Baguio, J.P., Borja, A., Lin, C.-H., and Fernando, A.G. 2024. Paleoichthyology in the Philippines: A review of Cenozoic fish fossils with insights on its current status and future opportunities. Geobios, 1–12.
https://doi.org/10.1016/j.geobios.2024.02.006
Messenger, R.W., Hart, M.B., Smart, C.W., Leng, M.J., Lock, E.J., and Howard, A.K. 2010. Pteropod faunas as indicators of Late Pleistocene climate change in the Caribbean Sea. Micropalaeontology, Sedimentary Environments and Stratigraphy: a Tribute to Dennis Curry (1912-2001), 4:18–28.
https://doi.org/10.1144/TMS004.3
Myers, T.D. 1968. Horizontal and vertical distribution of thecosomatous pteropods off Cape Hatteras. Ph.D. dissertation. Duke University, Durham, North Carolina, USA.
Nagel, S., Castelltort, S., Wetzel, A., Willett, S.D., Mouthereau, F., and Lin, A.-T. 2013. Sedimentology and foreland basin paleogeography during Taiwan arc continent collision. Journal of Asian Earth Sciences, 62:180–204.
https://doi.org/10.1016/j.jseaes.2012.09.001
Noda, H. 1972. Some fossil Pteropoda from Miyazaki and Okinawa prefectures, southwest Japan. Transactions and Proceedings of the Palaeontological Society of Japan, New Series, 88:472–484.
Orbigny, A.D. d’. 1834–1847. Voyage dans l’Amérique Méridionale (le Brésil, la République Orientale de l’Uruguay, la République Argentine, la Patagonie, la République du Chili, la République de Bolivia, la République du Pérou), Exécuté Pendant les Années 1826, 1827, 1828, 1829, 1830, 1831, 1832 et 1833: Tome 5(3) Mollusques. Chez Pitois-Levrault et ce, Paris. (In French)
Peijnenburg, K.T.C.A., Janssen, A.W., Wall-Palmer, D., Goetze, E., Maas A.E., Todd, J.A., and Marlétaz, F. 2020. The origin and diversification of pteropods precede past perturbations in the Earth’s carbon cycle. PNAS. 117(41):25609–25617.
https://doi.org/10.1073/pnas.1920918117
Quoy, J.R.C. and Gaimard, J.P. 1827. Observations zoologiques faites à bord de l’Astrolabe, en mai 1826, dans le Détroit de Gibraltar. Annales Des Sciences Naturelles, 10:225–239. (In French)
Rafenisque, C.S. 1814. Précis des Découvertes et Travaux Somiologiques de Mr. C. S. Rafinesque-Schmalz Entre 1800 et 1814. Royale typographie militaire, Palerme. (In French)
Rampal, J. 1973. Phylogénie des Ptéropodes Thécosomes d’après la structure de la coquille et la morphologie du manteau. Comptes Rendus Des Séances de l’Académie Des Sciences de Paris. Ser. D, 277:1345–1348. (In French)
Rampal, J. 2019. Euthecosomata (Mollusca: Gastropoda: Thecosomata). A taxonomic review. Bollettino Malacologico, 55:145–186.
https://doi.org/10.1101/098475
Rang, S. 1828. Notice sur quelques mollusques nouveaux appartenant au genre Cléodore, et etablissement et monographie des sous-genre Créseis. Annales Des Sciences Naturelles, 13:302–319. (In French)
Rang, S. 1829. Manuel de l’Histoire Naturelle des Mollusques et de Leurs Coquilles, Ayant pour Base de Classification Celle de M. le Baron Cuvier. Roret, Paris. (In French)
Reiss, Z., Luz, B., Almogi-Labin, A., Helicz, E., Winter, A., Wolf, M., and Ross, D.A. 1980. Late Quaternary paleoceanography of the Gulf of Aqaba (Elat), Red Sea. Quaternary Research, 14:294–308.
https://doi.org/10.1016/0033-5894(80)90013-7
Shibata, H. 1980. Pteropods from the Early Miocene (Kurami and Saigo Groups) of the Kakegawa District and the Early to Middle Miocene (Yatsuo Formation) of the Yatsuo District, central Japan. Bulletin of the Mizunami Fossil Museum, 7:59–68.
Shibata, H. 1984. Pteropods and heteropods from the Upper Cenozoic of Kakegawa, Shizuoka Prefecture, Japan. Bulletin of the Mizunami Fossil Museum, 11:73–91.
Shibata, H. 1986. Development of the Late Cenozoic pteropod fauna in central Japan. Palaeontological Society of Japan Species Papers, 29:119–126.
Shibata, H. and Ishigaki, T. 1981. Heteropodous and pteropodous biostratigraphy of Cenozoic strata of Chubu Province, Japan. Bulletin of the Mizunami Fossil Museum, 8:55–70.
Shibata, H. and Ujihara, A. 1983. Middle and Late Pleistocene heteropods and pteropods from Chiba, Noto Peninsula and Kikaijima, Japan. Bulletin of the Mizunami Fossil Museum, 10:151–169.
Spoel, S. van der. 1967. Euthecosomata: a Group with Remarkable Developmental Stages (Gastropoda, Pteropoda). J. Noorduyn en Zoon, N.V., Gorinchem.
Spoel, S. van der. 1971. New forms of Diacria quadridentata (de Blainville, 1821), Cavolinia longirostris (de Blainville, 1821) and Cavolinia uncinata (Rang, 1829) from the Red Sea and the East Pacific Ocean (Mollusca, Pteropoda). Beaufortia, 19(243):1–20.
Spoel, S. van der. 1987. Diacavolinia nov. gen. separated from Cavolinia (Pteropoda, Gastropoda). Bulletin Zoologisch Museum, 11(9):77–79.
Spoel, S. van der, Bleeker, J., and Kobayasi, H. 1993. From Cavolinia longirostris to twenty-four Diacavolinia taxa, with a phylogenetic discussion (Mollusca, Gastropoda). Bijdragen Tot de Dierkunde, 62(3):127–166.
Teng, K.-H. 1979. A quantitative study on the landforms of lateritic gravel tablelands in northwestern Taiwan. The College of Chinese Culture Institute of Geography Science Reports, 3:113–186.
Tesch, J. J. 1948. The thecosomatous pteropods. II. The Indo-Pacific. Dana Report, 5(30):1–45.
Torii, K. 1935. Explanatory text of the geological map of Taiwan, Tosei Sheet. Government-General of Taiwan, 732.
Torii, K. and Yoshida, K. 1931. Report on the geology of the oil fields of Miaoli and Chuntung, Hsinchu. Government-General of Taiwan, 1–81.
Ujihara, A. 1996. Pteropods (Mollusca, Gastropoda) from the Pliocene Miyazaki Group, Miyazaki Prefecture, Japan. Journal of Paleontology, 70:771–788.
Ujihara, A., Shibata, H., and Saito, T. 1990. Pteropods from the Sagara Group (Mio-Pliocene), Shizuoka Prefecture, Japan. Venus, 49:306–329.
Wall-Palmer, D., Smart, C.W., Kirby, R., Hart, M.B., Peijnenburg, K.T.C.A., and Janssen, A.W. 2016. A review of the ecology, palaeontology and distribution of atlantid heteropods (Caenogastropoda: Pterotracheoidea: Atlantidae). Journal of Molluscan Studies, 82(2):221–234.
https://doi.org/10.1093/mollus/eyv063
Wall-Palmer, D., Metcalfe, B., Leng, M.J., Sloane, H.J., Ganssen, G., Vinayachandran, P.N., and Smart, C.W. 2018. Vertical distribution and diurnal migration of atlantid heteropods. Marine Ecology Prgress Series, 587:1-15.
https://doi.org/10.3354/meps12464
Wang, C.-H., Peng, T.-R., and Chen, P.-F. 1991. Oxygen and Carbon isotopic compositions of mollusks from the Late Pleistocene Szekou Formation, Southern Taiwan. Proceedings of the National Science Council, Republic of China (A), 15(5):455–464.
Wang, L., Jian, Z., and Chen, J. 1997. Late Quaternary pteropods in the South China Sea: carbonate preservation and paleoenvironmental variation. Marine Micropaleontology, 32:115–126.
Williams, S.W. 1972. The temporal and spatial variation of selected thecosomatous pteropods from the Florida Middle Ground. M.S. thesis. Florida State University, Florida, USA.
Wormelle, R.L. 1962. A survey of the standing crop of plankton of the Florida current. VI. A study of the distribution of the pteropods of the Florida current. Bulletin of Marine Science of the Gulf and Caribbean, 12(1):93–136.
WoRMS Editorial Board. 2025. World Register of Marine Species. Creseis Rang, 1828. Available from https://www.marinespecies.org at VLIZ. Accessed 24 January 2025.
https://doi.org/10.14284/170
Yabe, H. and Hanzawa, S. 1930. Tertiary foraminiferous rocks of Taiwan (Formosa). Science Reports of the Tohoku Imperial University. 2nd Series, Geology, 14(1):1–46.
Yamakawa, G. and Ishikawa, M. 1912. Some pteropods from the Neogene of Semata (I and II). Journal of the Geological Society of Tokyo, 19:1–8, 17–24.
