

 Heteractinid, hexactinellid and sphaeroclonid sponges as rare components of anthaspidellid-dominated reefs from the Ordovician of the Precordillera, western Argentina
Heteractinid, hexactinellid and sphaeroclonid sponges as rare components of anthaspidellid-dominated reefs from the Ordovician of the Precordillera, western Argentina
Article number: 28.1.a17
https://doi.org/10.26879/1351
Copyright Palaeontological Association, April 2025
Author biographies
Plain-language and multi-lingual abstracts
PDF version
Submission: 11 October 2023. Acceptance: 26 March 2025
ABSTRACT
Three new sponges have been discovered in the extensively studied reef mounds of the Lower Ordovician limestones of the Precordillera of western Argentina significantly increasing the diversity of the group in this region. These groups occur as solitary individuals and are unusual in reef ecosystems at this time. The sponges include a new species, Niquivilispongia asteria gen. et sp. nov., which is the second named heteractinid from Argentina, fragments of a reticulosan (? Mattaspongia sp.), and a sphaeroclone demosponge. These include sponge groups that are relatively common in Mesozoic reef settings, but rare in Ordovician reefs. This shows that a consortium of sponges was evolving to adapt to reef environments during the earlier part of the Ordovician, including some that only appear as rare components of the ecosystem. Those rare components were either adapted to specific but limited niches within the reef system, or were opportunistic colonizers from other habitats.
Marcelo G. Carrera. Cicterra (Conicet-Universidad Nacional de Córdoba), Facultad de Ciencias Exactas, Físicas y Naturales, Av. Vélez Sarsfield 1699, X5016GCA, Córdoba, Argentina. mcarrera@unc.edu.ar
Joseph P. Botting. Department of Natural Sciences, Amgueddfa Cymru - National Museum Wales, Cathays Park, Cardiff CF10 3NP, UK. acutipuerilis@yahoo.co.uk
and Nanjing Institute of Geology and Palaeontology, Chinese Academy of Sciences, Nanjing, China.
Fernando L. Cañas. Dpto. de Geología, Facultad de Cs Exactas, Fisico-Químicas y Naturales, Universidad Nacional de Rio Cuarto, Córdoba, Argentina (ICBIA - UNRC). fcanas@exa.unrc.edu.ar
Keywords: sponges; new genus; new species; Ordovician; reef ecology; evolution
Final citation: Carrera, Marcelo G., Botting, Joseph P., and Cañas, Fernando L. 2025. Heteractinid, hexactinellid and sphaeroclonid sponges as rare components of anthaspidellid-dominated reefs from the Ordovician of the Precordillera, western Argentina. Palaeontologia Electronica, 28(1):a17.
https://doi.org/10.26879/1351
palaeo-electronica.org/content/2025/5423-ordovician-reef-sponges
Copyright: April 2025 Palaeontological Association.
This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
creativecommons.org/licenses/by/4.0
https://zoobank.org/2E6F520E-FAEB-40F3-A313-681254BE1BB2
INTRODUCTION
The Ordovician reefs of the Precordillera of western Argentina have been extensively studied (Carrera, 1991; Cañas and Carrera, 1993, 2003; Cañas and Keller, 1993). More recently, detailed analysis was carried out in these Lower and Middle Ordovician reefs (Carrera et al., 2017; Cañas and Carrera 2023), with the reef structures found to be mainly formed by microbial boundstones and sponges. Among the sponges, the orchoclad demosponges (“lithistids”) are by far the most abundant group. Other significant components recorded to date include the genus Calathium (Carrera, 1991; Cañas and Carrera, 1993), previously considered to be a receptaculitid alga but now understood to be a calcified sponge (Li et al., 2015), and the stomatoporoid-like organism Zondarella (Keller and Flügel, 1996), which is now considered to be a laminated microbial structure (Cañas and Carrera, 2023).
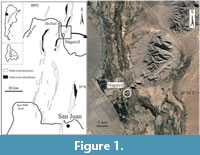 This paper reports the discovery of new sponges in reef mounds from the Lower Ordovician limestones (San Juan Formation) of the Precordillera of western Argentina (Figure 1), belonging to the heteractinid calcareans, hexactinellids (or reticulosans) and sphaeroclone orchoclads. The heteractinid was a relatively thin-walled sponge that represents an early branch within a group that later evolved to include many thick-walled and hypercalcified forms (Finks and Rigby, 2004a; Botting and Muir, 2018). Previous records of heteractinid sponges in Argentina are rare: there is a report of a thick-walled eifelliid (Chilcaia bimuralis Carrera, 1994) found in slightly younger levels (Darriwilian) of the same unit, and isolated hexaradiate spicules were also found by Beresi and Heredia (2000) in the Middle Ordovician Ponón Trehué area in southern Mendoza.
This paper reports the discovery of new sponges in reef mounds from the Lower Ordovician limestones (San Juan Formation) of the Precordillera of western Argentina (Figure 1), belonging to the heteractinid calcareans, hexactinellids (or reticulosans) and sphaeroclone orchoclads. The heteractinid was a relatively thin-walled sponge that represents an early branch within a group that later evolved to include many thick-walled and hypercalcified forms (Finks and Rigby, 2004a; Botting and Muir, 2018). Previous records of heteractinid sponges in Argentina are rare: there is a report of a thick-walled eifelliid (Chilcaia bimuralis Carrera, 1994) found in slightly younger levels (Darriwilian) of the same unit, and isolated hexaradiate spicules were also found by Beresi and Heredia (2000) in the Middle Ordovician Ponón Trehué area in southern Mendoza.
The occurrence of a reticulosan (a probable stem-group hexactinellid sponge) is unusual in shallow Ordovician carbonate platforms, and extremely rare as a component of Lower Paleozoic reef environments. Only a few isolated hexactine spicules have been previously reported in the San Juan limestones of the Argentine Precordillera (Carrera, 2001; Mehl and Lehnert, 1997). In addition, a fragment of a sponge skeleton composed of sphaeroclone spicules has been recovered from the same reef mounds. Representatives of this particular orchoclad group are more commonly found in Upper Ordovician and Silurian deposits.
GEOLOGICAL SETTING
 The San Juan Formation is a fossiliferous muddy carbonate unit developed on top of a Cambrian to Lower Ordovician carbonate platform succession (Cañas, 1999). Skeletal wackestones and packstones intercalated with storm-generated intraclastic grainstones are the most conspicuous lithologies (Figure 2). Frequent centimeter-thick, lithoclastic-bioclastic grainstone and rudstone beds are interpreted as storm layers (Cañas, 1999), and usually have sharp, irregular erosive bases cut into the top of underlying wackestone layers. The frequency of these storm deposits through the sequence is uneven, being concentrated in this interval, which is interpreted as a middle ramp setting (Cañas, 1999). Above this interval, amalgamated medium-bedded lithoclastic-bioclastic grainstone, packstones or intraclastic rudstones characterize the Monorthis cumillangoensis Brachiopod Zone, associated with sponge-algal reef mounds; these are interpreted as inner ramp deposits (Cañas, 1999; Cañas and Carrera, 2003; 2023). The whole upper interval with reef structures has a variable thickness, developed within a diachronous transgressive systems tract with an erosive boundary at the base (Cañas, 1999; Keller, 1999).
The San Juan Formation is a fossiliferous muddy carbonate unit developed on top of a Cambrian to Lower Ordovician carbonate platform succession (Cañas, 1999). Skeletal wackestones and packstones intercalated with storm-generated intraclastic grainstones are the most conspicuous lithologies (Figure 2). Frequent centimeter-thick, lithoclastic-bioclastic grainstone and rudstone beds are interpreted as storm layers (Cañas, 1999), and usually have sharp, irregular erosive bases cut into the top of underlying wackestone layers. The frequency of these storm deposits through the sequence is uneven, being concentrated in this interval, which is interpreted as a middle ramp setting (Cañas, 1999). Above this interval, amalgamated medium-bedded lithoclastic-bioclastic grainstone, packstones or intraclastic rudstones characterize the Monorthis cumillangoensis Brachiopod Zone, associated with sponge-algal reef mounds; these are interpreted as inner ramp deposits (Cañas, 1999; Cañas and Carrera, 2003; 2023). The whole upper interval with reef structures has a variable thickness, developed within a diachronous transgressive systems tract with an erosive boundary at the base (Cañas, 1999; Keller, 1999).
The samples were collected from exposures of the San Juan Formation at the Niquivil locality (Figure 1). The material described herein is restricted to the base of Monorthis cumillangoensis Brachiopod Zone (Benedetto, 2002), which according to the associated conodonts can be referred to the B. navis Conodont Zone, of early Dapingian age (Lehnert and Keller, 1993; Albanesi et al., 2006; Mango and Albanesi 2020).
The studied sponges come from this last interval and were found enclosed in microbial boundstone reef mounds, associated with orchoclad demosponges and calathids, together with the stromatoporoid-like feature Zondarella (Keller and Flügel, 1996), now considered a microbial stromatolitic structure (Cañas and Carrera, 2023).
The sponge component in the construction of the Argentinean reefs is important, but constitutes a minor proportion compared with the primary microbial framework (Cañas and Carrera, 1993). However, in a recent study (Cañas and Carrera, 2023), the small reef mounds (kalyptra and kalyptrate reefs with a few isolated stromatolites) show abundant demosponges with entire sponge bodies or amorphous spiculitic boundstones forming scaffolding structures onto which small stromatolites or calcimicrobe crusts developed. This suggests that the proportion of sponges may be underestimated due to the lack of diagnostic features (Cañas and Carrera, 2023).
These reefs are primarily characterized as boundstones, where calcimicrobes and microproblematica, such as Girvanella, Rothpletzella and Allonema (formerly Wetheredella, Jarochowska and Munnecke, 2014), serve as the primary binders, fixing the sediment. Additionally, isolated domal to laminar stromatolites (Zondarella structures of Keller and Flügel, 1996), formed by the calcimicrobe Acuasiphonoria (Cañas and Carrera, 2023), further contribute to this sediment-binding process. Orchoclad demosponges (anthaspidellids) and calathids act as bafflers, trapping sediment or providing structural scaffolding.
Bioturbated wackestone occupies spaces within the mounds. The inter-reef facies are characterized by grainstones and rudstones with intraclasts and fragments of nautiloids, trilobites, gastropods and echinoderms.
MATERIAL
The sponges were found in three different decimeter-sized blocks of the reef mound that were collected for thin-section preparation. In one block there are at least two laminar to mound-shaped forms of the heteractinid, separated vertically by approximately 1.5 cm. The hexactinellid sponge is a 4-cm fragment included in a different block, and the sphaeroclonid was found in a thin section from the same reef mounds.
The reef structures described in previous studies by Cañas and Carrera (1993, 2023, and references therein) have been the subject of continuous sampling for over 30 years. Most of these samples were prepared as polished slabs of varying sizes and thin sections (primarily taken from a quadrate grid). Additionally, a significant volume of material was processed for acid dissolution and the recovery of insoluble microfossils. In recent years, as part of a research project and a Ph.D. program, another sampling effort was conducted, focusing on the upper reef mound interval. These structures, which are also dominated by microbial boundstones and lithistid orchoclad demosponges, contain the scarce sponges described in this contribution.
The material is housed under the prefix CEGH-UNC in the Centro de Investigaciones en Ciencias de la Tierra (CICTERRA), Universidad Nacional de Córdoba, Argentina.
SYSTEMATIC PALAEONTOLOGY
Class CALCAREA Bowerbank, 1864
Order HETERACTINIDA Hinde, 1888
Remarks. Heteractinids are a group of Palaeozoic sponges with anomalous spicules containing a non-actualistic number of rays (de Laubenfels, 1955; Rigby and Nitecki, 1975; Kozur et al., 2008). In almost all cases, the spicules are based on a hexaradiate pattern, sometimes with an additional pair of rays in the perpendicular plane (octactins), or additional radial rays (Kozur et al., 2008). The triradiate or hexaradiate symmetry mirrors that of calcite crystals, which appears to influence the growth form (Jones, 1970), although there is some evidence for the existence of a mineralogically distinct core, as in certain other early sponge groups (Botting and Butterfield, 2005). They are historically and currently regarded as representing the early stem group of Calcarea (Botting and Muir, 2018).
Family ?ASTRAEOSPONGIIDAE Miller, 1889
Remarks. The classification of heteractinids is currently difficult to apply consistently, with only four families listed by Finks and Rigby (2004a). Of these, the Eiffeliidae are thin-walled (normally a single layer of spicules), and probably plesiomorphic and paraphyletic (Botting and Muir 2018). The Wewokellidae have robust spicules including septiradiates, whereas the Nuchidae are normally aspiculate, with a perforated calcareous wall. The remaining family, Astraeospongiidae, is diverse and includes essentially all the species not included in the other families, but consists largely of thick-walled, bowl-shaped to vasiform sponges with robust, mostly octactine spicules that are often partly fused. As a result, the Astraeospongiidae includes most of the morphological features seen in the new species. This includes a shallow bowl-shaped to domal morphology in several genera, a thick wall with a disorganized spicule array and even the presence of fine monaxons in genera such as Ensiferites Reimann, 1945. In most astraeospongiid genera, at least the outer spicules are thickened to the point of becoming lumpy and irregular. In Blumenbachium Koenig, 1820, the spicules are dominantly four-rayed, and hexaradiate spicules are absent; however, this understanding is based only on the original illustrations (Finks and Rigby, 2004a), and the morphology of these spicules resembles receptaculitid meroms; it may, therefore, not be a sponge. Nonetheless, of the existing families within the Heteractinida, only the Astraeospongiidae is able to accommodate most aspects of the morphology and spiculation of the new species, and it is tentatively included there.
NIQUIVILISPONGIA gen. nov.
zoobank.org/0BB8C6D5-E886-4BDA-A27D-457D4E479096
Diagnosis. As for the species, by monotypy.
Etymology. After the Niquivil locality, San Juan Province, Argentina.
Remarks. The closest similarity of the new genus is to Ensiferites Reimann, 1945. Niquivilispongia gen. nov. has comparable body morphology and both genera have fine monaxon-like spicules in addition to octactins. However, the new genus is differentiated by the absence of hypertrophied gastral rays in dermal-layer spicules and the presence of a conspicuous array of long, irregular, hypertrophied heteractinid rays and undulating monaxons, which may constitute a root tuft.
The unusual feature of an apparent root tuft may have helped in anchoring the sponge to the substrate. Such a feature is rare among heteractinids, with only Eiffelospongia Rigby and Collins, 2004, possessing a root tuft of sub-vertically aligned monaxons. However, Eiffelospongia requires a careful revision, for it shows so many cross-laid spicules that these could be in fact diagonal stauractins overlaid by longitudinal monaxons, as in Diagoniella (JB personal observation of the type material).
Niquivilispongia asteria sp. nov.
Figure 3-Figure 4
zoobank.org/3E1D53E0-0F1D-4D04-A84C-6FE47D989146
 Diagnosis. Domal to mound-shaped, encrusting heteractinid sponge with homogeneous, dense spicular net, mainly composed of hexaradiate heteractins (six rays in one plane) and small monaxons. A complex root tuft of irregular undulating monaxon-like spicules and hypertrophied heteractinid rays emerge from the base of the sponge body.
Diagnosis. Domal to mound-shaped, encrusting heteractinid sponge with homogeneous, dense spicular net, mainly composed of hexaradiate heteractins (six rays in one plane) and small monaxons. A complex root tuft of irregular undulating monaxon-like spicules and hypertrophied heteractinid rays emerge from the base of the sponge body.
Etymology. From Greek, asteria, meaning stars, referring to the star-like appearance of the heteractinid spicules.
Holotype. CEGH-UNC 27593, a section through the sponge in a thin section.
Paratypes. CEGH-UNC 27594-27595; additional thin sections cutting through sponges, from the type locality.
Type locality. Niquivil, Precordillera of Argentina; Middle Ordovician (early Dapingian; T laevis -B navis Conodont zones, Monorthis cumillangoensis Brachiopod Zone).
Description. Domal to mound-shaped encrusting heteractinid sponge 4 to 5 cm in maximum diameter (measured at the base). No evidence of central cavity or canals are seen within the homogeneous, dense spicular net, which is mainly composed of hexaradiate heteractins (six rays in one plane) and small monaxons.
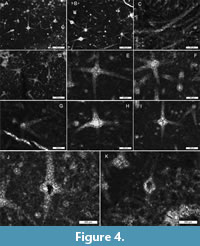 The heteractine spicules are 1 to 1.5 mm in maximum diameter, and typically 60–80 μm in basal ray diameter. No size ranks are observed, although traces of numerous finer spicules are visible in the background. The heteractins are octactine where visible, with six horizontal (in one plane) slightly curved rays and two perpendicular rays, arising from the axis. All rays equal in size and thickness.
The heteractine spicules are 1 to 1.5 mm in maximum diameter, and typically 60–80 μm in basal ray diameter. No size ranks are observed, although traces of numerous finer spicules are visible in the background. The heteractins are octactine where visible, with six horizontal (in one plane) slightly curved rays and two perpendicular rays, arising from the axis. All rays equal in size and thickness.
At a first glance the thin sections appear to show a small number of hexactin-type spicules, with four equal rays in one plane, of the same size as the heteractine spicules, and with similar morphology of evenly tapered, sometimes curved rays. Rays in one plane are generally orthogonal, but some show deviations, including possibly interposed rays bisecting the space between two others (Figure 3G). However, this is almost certainly an artefact of the angle of section through standard octactins (see remarks).
Internal structure of the spicules is generally replaced by coarse calcite cement showing typical drusy texture with increasing crystallite size towards the center. Some spicules show an axial hollow, either irregular (typical of incomplete infill of cavity following spicule dissolution) or with a sharp, circular boundary. In the latter case, the hollow is approximately 100 μm in diameter. In some of the entirely calcitized spicules, there is a circular, axial discontinuity within the structure, of similar dimensions (Figure 4 J-K).
A complex root tuft or similar structure emerges from the base of the sponge body in one paratype, the spicules protruding from the base of the sponge either as a continuation or extension of the main spicule rays, or as simple juxtaposed monaxon-like spicules (Figure 3I-K; Figure 4C). Some of the spicules included in this skeletal base appear to be fine-rayed heteractins or similar, with shortened lateral rays and extended, undulating basal rays.
Remarks. The possible presence of hexactins is a critical feature for the interpretation of the affinities of the new sponge (Botting and Butterfield, 2005) and needs to be considered carefully. Such features can be especially challenging in thin sections; thin-walled heteractinids like Eiffelia are preserved on rock surfaces, where orientations are consistent, and spicule morphology is easier to assess. For thick-walled heteractinids, there is much more scope for variability in orientation, leading to misleading appearance due to the angle of section. Unfortunately, the small size and the lack of mineralogical contrast of the spicules versus the host rock prevent the use of MicroCT to generate a three-dimensional view. To address this, we considered all possible sectioning planes through an octactine spicule, as some orientations can show the presence of four rays in a plane (appearing as a hexactin or derivative), and we compared this with the observed frequency of such spicules. If the spicules were all octactins, then there is only one orientation of section that would give six visible rays, but three (along each of the coplanar axes) that would make them appear as triaxons (i.e., hexactins/stauractins). Therefore, with random orientation of spicules, we would need to see significantly more than three times as many ‘hexactins’ as hexaradiate spicules, in order to assume that genuine triaxons were present. Instead, we found that the proportion of apparent hexactins is very low, with at least 20 hexaradiate views for each four-rayed view within a given field of view. This is much lower than expected for a random orientation of octactins and indicates a preferred orientation with the six coplanar rays approximately parallel to the angle of section (which appears to be parallel to the body wall). Overall, it is most likely that the apparent four-rayed spicules are in fact octactins cut through one of three possible planes.
Differentiation from other genera of the family was discussed under the genus heading. Distinction between species of the genus, if more are discovered, should be based on minor skeletal features, such as spicule net arrangement, types and proportions of spicules, or their size and arrangement. Development of canals could also be taken into account for species differentiation, together with any consistent difference in body form.
Class ?HEXATINELLIDA Sollas, 1887
“RETICULOSA”
Remarks. Reticulosans are a complex group of hexactinellid-like sponges with a thin wall of loose hexactins and derivatives. However, current understanding suggests that most of them fall into the stem groups of Hexactinellida and Silicea, and probably also the stem group of Porifera (Botting and Butterfield, 2005; Botting and Muir, 2018). These sponges usually dominated Cambrian to Silurian offshore sponge communities, below storm wave base (Muir and Botting 2015), but they declined dramatically thereafter. An exception is the Dictyospongiidae, a distinctive family in which the skeleton developed into a tough, woven structure that was able to survive turbulent, shallow-water conditions, and which thrived during the Devonian and Carboniferous in particular (Hall and Clarke, 1899). Some dictyosponges possessed some hexactinellid-like microscleres (Kling and Reif, 1969) and probably fall into the late stem group of Hexactinellida. The current taxon, however, remains unresolved as to whether it should be placed within the total group of Hexactinellida, or in a deeper position.
Family MATTASPONGIIDAE Rigby, 1970
Genus MATTASPONGIA Rigby, 1970
Remarks. Rigby (1970) erected the genus Mattaspongia and the family Mattaspongiidae because there were no comparable genera within what he considered to be the closest family, the Dictyospongiidae Hall and Clarke, 1899. The visible characters of Mattaspongia did not fit well with the definition of the Dictyospongiidae, as the genus lacks a woven meshwork and the spicules are effectively loose within soft tissues (the lyssacine condition). However, Finks and Rigby (2004b) refined the definition of the Dictyospongiidae to include six subfamilies, with the genus Mattaspongia included in the Subfamily Thysanodictyinae, Hall and Clarke, 1899, which involves “Dictyospongids with coarse ridgelike, quadrate mesh that may represent radially erect lamellae, spicule bundles, coarse pentactins or a combination thereof” (Finks and Rigby, 2004b: 398). This still does not reasonably accommodate Mattaspongia, as it possesses a quadrate mesh but this reticulation lacked any coarse, ridge-like development, or even overlapping spicule rays. However, it is distinct from earlier reticulosans in being dominated by a single size of hexactin, rather than having subdivided quadrules. We therefore follow Rigby’s (1970) original interpretation, including Mattaspongia tentatively within its own family, and rejecting an assignment to the dictyosponges.
? MATTASPONGIA sp.
(Figure 5A-D)
Material. One fragment from a reef mound block at Niquivil locality Middle Ordovician (Dapingian), CEGH-UNC 27596.
 Description. Thin-walled sponge, apparently tubular and with a faintly visible osculum at the top. The sponge fragment is 4 cm long and approximately 2 cm wide. Main body spicules are coarse, predominantly aligned hexactin-type forms, mostly of the same size. This main layer consists of large hexactins arranged in moderately regular quadrate disposition, with rays aligned to form parallel rows. Vertical rays are intercalated to form small quadrules and irregular, somewhat alternating rows. Vertical rows of spicules are commonly between 2 and 3 mm apart. Horizontal spicule rays generally extend below vertical rays of adjacent spicules. Small monaxons (0.1–0.3 mm long) are irregularly distributed between the primary rays, most of them horizontally disposed, but some appear perpendicular or with variable angles.
Description. Thin-walled sponge, apparently tubular and with a faintly visible osculum at the top. The sponge fragment is 4 cm long and approximately 2 cm wide. Main body spicules are coarse, predominantly aligned hexactin-type forms, mostly of the same size. This main layer consists of large hexactins arranged in moderately regular quadrate disposition, with rays aligned to form parallel rows. Vertical rays are intercalated to form small quadrules and irregular, somewhat alternating rows. Vertical rows of spicules are commonly between 2 and 3 mm apart. Horizontal spicule rays generally extend below vertical rays of adjacent spicules. Small monaxons (0.1–0.3 mm long) are irregularly distributed between the primary rays, most of them horizontally disposed, but some appear perpendicular or with variable angles.
Main spicules are coarse-rayed hexactins, with equal ray size of approximately 2 mm long. Basal ray diameter is 0.3 mm, tapering rapidly to a pointed tip. Dermal ray with a button-like apex or may appear to be absent, possibly due to breakage or awkward angle of preservation. Primary hexactins, seen in a transverse polished section at the base, apparently show a shorter perpendicular ray than the horizontal ones.
Remarks. Interpretation of sponges from fragments is challenging, especially in sectional view. Nonetheless, the main characteristics of Mattaspongia (i.e., a primary layer of coarse hexactins forming a quadrate mesh, vertical rays overlapping to form small quadrules in irregular and somewhat alternating rows, and small monaxons irregularly distributed among quadrules) are seen in our material. However, the presence of conspicuous parietal gaps (or internal canals) in Mattaspongia may indicate separation of the specimen described here from the genus erected by Rigby (1970); this is only likely to be resolved with additional material.
Class DEMOSPONGIAE Sollas, 1875
Order ORCHOCLADINA Rauff, 1895
Family ASTYLOSPONGIIDAE Zittel, 1877
ASTYLOSPONGIIDAE indet.
(Figure 5E-F)
Material. A single thin section from the San Juan reef mound block at Niquivil locality Middle Ordovician (Dapingian), CEGH-UNC 27597.
Description. A single thin section from a reef mound slab shows a fragmented skeleton of a sponge composed of fused sphaeroclones that form an isodictyal net with triangular interspaces in all directions. The sphaeroclones are six-armed anapodal desmas (dichotriders) with terminal expansions (zygosis).
The spicule net shows an open structure, at least as far as is visible in the small area preserved, with sphaeroclones being anaxial desmas with five to six ray-like arms extended from one side of a globular centrum and showing knobby or spiny terminations. The globular centrum of the sphaeroclones in the specimen is variable in size (0.4 to 0.8 mm) and formed by the fusion of the zygome endings of the rays (arms), which are generally slender and long (0.7 to 1.1 mm), forming an open triangular structure. In well-preserved sphaeroclones, or incompletely fused centra, the arms are composed of a cladome with a simple bifurcated zygome; the zygomes of multiple spicules interlock to form the globular center.
Remarks. The characteristics exhibit by the spicular net seen in the thin section are not fully comparable with, or are difficult to relate to, described astylospongiids. Similar characteristics are seen in the Upper Ordovician and Silurian genera Astylostroma (Rigby and Webby, 1988) and Astylospongiella (Rigby and Lenz, 1978). In particular, Astylostroma has similar zygome junctions; however, the lack of fine detail in the spicule preservation, and the limited material, prevent a more comprehensive comparison.
Most of the described genera of the family have a wide distribution but are restricted to the Upper Ordovician and Silurian, with only a few occurrences having been recorded from Middle Ordovician rocks (see Finks and Rigby, 2004c). It therefore appears that the early Middle Ordovician (Dapingian) age of our material may represent the oldest record of the family.
DISCUSSION
Spiculate Sponges as Main Metazoan Builders in Lower-Middle Ordovician Reefs
Immediately following the demise of the early Cambrian calcimicrobial-archaeocyathan reefs, metazoans were uncommon as reef builders (Rowland and Gangloff, 1988; Rowland and Shapiro, 2002; Adachi et al., 2015). Many reefs of the post-extinction interval are composed purely of microbialites (Rowland and Shapiro, 2002; Webby, 2002; Lee et al., 2015). By the late Cambrian to Early Ordovician, the spiculate lithistids (orchoclad demosponges) had become the main metazoan components within reefs that were still dominantly microbial (Webby, 2002; Carrera and Rigby, 1999, 2004; Carrera and Botting, 2008). These sponges, which were clearly the dominant metazoans in reefs of Lower and Middle Ordovician age on all continents, are also commonly found together with classical forms now considered hypercalcified sponges, such as pulchrilaminids and calathids.
Many Ordovician demosponge-dominated reefs have been discovered since the early 1970s, most of them located in North America (Church, 1974; Toomey and Nitecki, 1979; Rigby et al., 1995; Carrera and Rigby, 1999, 2004; and references therein), and western Argentina in South America (Cañas and Carrera, 1993, 2003, 2023). More recently, important occurrences have been recorded in Asia, primarily in China and Korea (see Adachi et al., 2009; Li et al., 2015; Choh, et al., 2013 Lee and Riding 2018 and references therein). In the majority of these reefs, cosmopolitan genera of orchoclad demosponges (“lithistids”) occur in various proportions (Carrera and Rigby, 2003; Choh, et al., 2013).
The first complete fossil lithistids have been found in middle Cambrian rocks, but the group only became common in upper Cambrian and Lower Ordovician shallow-water limestones (Carrera and Botting 2008). Interlocking spicules in isodictyal nets, and cemented attachment structures, were their main adaptations for inhabiting shallow environments, including reef settings (Finks, 2003). In the Cambrian and Early Ordovician, anisotropic triangulated skeletons (with triangles parallel to the surface) predominate, as in the orchoclad lithistids, particularly the Anthaspidellidae (Finks, 2003). Such a structure was able to maintain rigidity in agitated waters, even in anthaspidellids with elongate tubular or conical shapes. These sponges invaded reef environments in the late Cambrian (Carrera and Botting, 2008).
A diversification of the orchoclad lithistids occurred during the Early Ordovician (Carrera and Rigby, 2004), at species, genus and family level. This led to occupation of a variety of shallow- to deep-platform environments during the Lower and Middle Ordovician. The trend toward more resistant skeletons, associated with colonization of more turbulent environments through the Cambrian–Ordovician, appears to have been one of the key aspects of early sponge evolution (Carrera and Botting, 2008).
Although this is a key interval for lithistid diversification and evolutionary adaptation, other sponge groups do not obviously show a synchronous evolutionary history in the same aspects. Hexactinellida, for example, are a deep-water clade that was beginning to diversify in extremely offshore environments during the Cambrian and Ordovician (Del Mouro et al. 2024). The hexactinellid record in the Palaeozoic is complex, and very confused with that of reticulosans (which also include stem-group hexactinellids, as well as earlier-branching groups; Botting and Muir, 2018), from which they cannot easily be separated.
The main associations of Cambrian spiculate sponges are in siliciclastic deposits from middle to deep muddy shelf and basinal environments. These relatively deep environments yield only simple asconoid-type reticulosan sponges, protomonaxonid demosponges (now Ascospongiae Botting, 2021), and a few heteractinids (see Carrera and Rigby 1999; 2003; Carrera and Botting 2008). This differential distribution persisted through the Lower and Middle Ordovician (see Carrera and Rigby 1999, 2004; Botting et al. 2021). Some lineages of presumed total-group hexactinellids such as brachiosponges became adapted to carbonate platforms (Rigby, 1970) and other inshore, turbulent environments (Botting, 2005) during the Middle to Late Ordovician, but are not typical components of reefs. A major trend towards reef environments among hexactinellids is not seen until the Mesozoic.
Ordovician polyactinellids and heteractinids are associated with carbonate platform facies in the Argentine Precordillera (Carrera, 1994; Mehl and Lehnert, 1997). These groups show very few changes in their distribution and abundance through the Cambrian–Ordovician transition. Only during the Middle and Late Ordovician do the heteractinids show a significant increase in the number of genera and families (Rigby, 1991; Carrera and Rigby, 2004).
The heteractinids show a clear trend towards robust construction and occupation of shallow-water environments, from thin-walled and delicate ancestors (Rigby and Nitecki, 1975; Botting and Butterfield, 2005). In particular, some Ordovician and Silurian heteractinids had robust skeletons of coarse spicules that are often well preserved, and demonstrate an unambiguous calcitic composition (Mehl and Reitner, 1996; Rhebergen, 2007), despite an apparent biminerallic composition of the ancestral taxa, which also shared hexactins with the Silicea (Botting and Butterfield, 2005). The group developed through the Palaeozoic towards thicker body walls, a more strongly fused construction, and habitation of more turbulent environments such as reefs (Rigby and Nitecki, 1975), so that the appearance of a heteractinid in the San Juan Formation reefs is not surprising. A remaining problem is the question of why such sponges, if present, were not more successful and common. The skeleton of the new heteractinid is complex and resembles those of the Silurian and Devonian heteractinids (see Rigby and Nitecki 1975; Rigby 1991; Finks and Rigby 2004a). In this sense the finding of a Lower-Middle Ordovician astraeospongiid slightly predates the oldest first occurrences of the family and is certainly older than their main diversification in the Silurian.
Mesozoic sponges, especially hexactinellids and lithistids, have an essentially modern aspect. Among lithistids those with tetraclone (not known in the Paleozoic) and rhizoclone desmas are the most common (Rigby, 1991; Pisera and Bodzioch, 1991; Pisera, 2004). Among Hexactinellida the earliest representatives of the Lychniscosa occur in the Triassic (although hexactinosans are known from the Devonian; Rigby et al., 2001; Nose et al., 2013) and both are equally common and highly diversified (Krautter, 2002). A much deeper origin is indicated by the presence of diagnostic sceptrulophoran microscleres from the base of the Ordovician (Carrera and Maletz, 2014), and it is likely that these lineages had already diversified in deep ocean environments that are not generally preserved. During the widespread and sheltered epeiric seas of the Mesozoic, these groups were able to colonize shallow water and became visible in the normal fossil record. Lychniscosan hexactinellids, in particular, maintain a high diversity during the Mesozoic (Pisera, 1997; 2004), but the Cenozoic record is much more sparse. The complex three–dimensional skeleton of clearly fused spicules of the Hexactinosa and Lychniscosa may possibly have been a preadaptation that allowed hexactinellids to colonize shallow carbonate platforms and reefs. There are several examples of hexactinellids as part of Mesozoic reefs (Palmer and Fursich, 1981; Werner et al., 1994; Leinfelder, et al., 1996; Senowbary-Daryan and Amirhassankhani, 2012; Tomás, et. al., 2019), and there are also modern reef-building hexactinellids in deeper water (Conway et al., 2001). In shallow water, hexactinellids are currently limited to cold or very sheltered environments such as caves (Vacelet et al., 1994). The existence of hexactinellids in warm, shallow conditions during the Mesozoic is therefore anomalous, and may be related to sheltered conditions or relatively low levels of turbulence. This is similar to the presence of Mattaspongia ? sp. in the Ordovician Argentine reefs, as reported here; it was not a natural reef-builder in terms of the robustness of its skeleton, and must have required sheltered conditions that were at least locally available, perhaps in a similar way to the late-surviving ascosponge Neochoiaella with the reticulosan Ammonella in reef-bounded lagoonal conditions in the Jurassic (Keupp and Schweigert, 2012).
The astylospongid lithistid sponge found is also unexpected, as this and other sphaerocladines of the suborder Tricranocladina, were previously thought to have originated and diversified in the Late Ordovician and Silurian. The finding of this sphaeroclone demosponge in Lower–Middle Ordovician reefs represents the oldest record of the group and predates their later diversification, which was primarily in different environments such as siliciclastic or mixed siliclastic-carbonate facies. Silurian and Late Ordovician sphaeocladines are mainly found in Eastern Laurentia, Baltica and New South Wales (Rigby and Webby 1988; Carrera and Rigby, 1999; 2004; Finks and Rigby 2004c).
CONCLUSIONS
Three previously undocumented poriferan components of Middle Ordovician microbial-sponge reef mounds have been recovered from the San Juan Formation of western Argentina. The new species, Niquivilispongia asteria gen. et sp. nov. is an astraeospongiid heteractinid with complex and robust skeleton, that predates the complexity found in Late Ordovician and Silurian forms representing the oldest record of the family.
A reticulosan (? Mattaspongia sp.) and a sphaeroclone demosponge have also been recovered, both of which are unusual reef components at this time and also represent the oldest records of both groups. This shows that a consortium of sponges was evolving to adapt to reef environments during the earlier part of the Ordovician, including some that only appear as rare components of the ecosystem.
ACKNOWLEDGEMENTS
We acknowledge support from CONICET (PIP 2021-2023 DD787 Cod F81643) to MGC. We are grateful to the editors of the journal and two anonymous reviewers for their helpful comments. This is a contribution for the IGCP 735 “Rocks and the Rise of the Ordovician life: Filling knowledge gaps in the Early Paleozoic Biodiversification”.
REFERENCES
Adachi, N., Ezaki, Y., Liu, J., and Cao, J. 2009. Early Ordovician reef construction in Anhui Province, South China: A geobiological transition from microbial- to metazoan-dominant reefs. Sedimentary Geology, 220:1–11.
https://doi.org/10.1016/j.sedgeo.2009.05.012
Albanesi, G., Carrera, M.G., Cañas, F., and Saltzman, M. 2006. Definition of a global boundary stratotype section and point for the base of the Middle Ordovician Series: The Niquivil section, Precordillera of San Juan, Argentina. Episodes, 29:1–15.
https://doi.org/10.18814/epiiugs/2006/v29i1/001
Benedetto, J.L. 2002. The Ordovician brachiopod faunas of Argentina: Chronology and biostratigraphic succession, p. 87–106. In Aceñolaza, F.G. (ed.), Aspects of the Ordovician System in Argentina. INSUGEO, Serie Correlación Geológica 16, San Miguel de Tucumán.
Beresi M.S. and Heredia, S.E. 2000. Sponge spicule assemblages from the Middle Ordovician of Ponón Trehué, southern Mendoza, Argentina. Revista Española de Paleontología 15:37–48.
https://doi.org/10.7203/sjp.15.1.22011
Botting, J.P. 2005. Exceptionally well-preserved Middle Ordovician sponges from the Llandegley Rocks Lagerstätte, Wales. Palaeontology, 48:577–617.
https://doi.org/10.1111/j.1475-4983.2005.00470.x
Botting, J.P. 2021. Hexactins in the ‘protomonaxonid’ sponge Choiaella and proposal of Ascospongiae (class nov.) as a formal replacement for the Protomonaxonida. Bulletin of Geosciences, 96:265–277.
https://doi.org/10.3140/bull.geosci.1823
Botting, J.P. and Butterfield, N.J. 2005. Reconstructing early sponge relationships by using the Burgess Shale fossil Eiffelia globosa, Walcott. Proceedings of the National Academy of Sciences of the United States of America, 102:1554–1559.
https://doi.org/10.1073/pnas.0405867102
Botting, J.P. and Muir, L.A. 2018. Early sponge evolution: a review and phylogenetic framework. Palaeoworld, 27:1–29.
https://doi.org/10.1016/j.palwor.2017.07.001
Bowerbank, J.S. 1864. A Monograph of the British Spongiadae. Vol. I, Ray Society, London.
Cañas, F.L. 1999. Facies and sequences of the late Cambrian–early Ordovician carbonates of the Argentine Precordillera: a stratigraphic comparison with Laurentian platforms, p. 43–62. In Ramos, V.A. and Keppie, J.D. (eds.), Laurentia-Gondwana Connections before Pangea.
https://doi.org/10.1130/0-8137-2336-1.43
Cañas, F.L. and Carrera, M.G. 1993. Early Ordovician microbial-sponge-receptaculitid bioherms of the Precordillera, Western Argentina. Facies, 29:169–178.
https://doi.org/10.1007/BF02536927
Cañas, F.L. and Carrera, M.G. 2003. Precordilleran reefs, p. 131–153. In Benedetto, J.L. (ed.), Ordovician Fossils of Argentina. Secretaría de Ciencia y Técnica, Universidad Nacional de Córdoba, Córdoba.
Cañas, F.L. and Carrera, M.G. 2023. Microbial origin of the Ordovician stromatoporoid-like organism Zondarella from the Argentine Precordillera and the post-Cambrian persistence of stromatolite microbialite reefs. Palaeogeography, Palaeoclimatology, Palaeoecology, 627:111752.
https://doi.org/10.1016/j.palaeo.2023.111752
Cañas, F.L. and Keller, M. 1993. Arrecifes y montículos arrecifales en la Formación San Juan (Precordillera Sanjuanina, Argentina): los arrecifes más antiguos de Sudamérica. Boletin Real Sociedad Española Historia Natural, Sección Geologia. 88, 127–136.
Carrera, M.G. 1991. Los géneros Selenoides Owen y Calathium Billings (Receptaculitaceae) en el Ordovícico de la Precordillera de San Juan, Argentina. Ameghiniana, 28:375–380.
Carrera, M.G. 1994. An Ordovician sponge fauna from San Juan Formation, Precordillera basin, western Argentina. Neues Jahrbuch für Geologie und Paläontologie (Abhandlungen), 191:201–220.
Carrera, M.G. 2001 Análisis de la distribución y composición de las biofacies de la Formación San Juan (Ordovícico temprano), Precordillera Argentina. Ameghiniana, 38:169–184.
Carrera, M.G. and Botting, J.P. 2008. Evolutionary history of Cambrian spiculate sponges: implication for the Cambrian evolutionary fauna. Palaios, 23:124–138.
https://doi.org/10.2110/palo.2006.p06-089r
Carrera, M.G. and Maletz, J. 2014. Ordovician sponge spicules from Spitsbergen, Nevada and Newfoundland: new evidence for hexactinellid and demosponge early diversification. Journal of Systematic Palaeontology, 12:961–981
https://doi.org/10.1080/14772019.2013.846946
Carrera, M.G. and Rigby, J.K. 1999. Biogeography of the Ordovician sponges. Journal of Paleontology, 73:26–37.
https://doi.org/10.1017/S0022336000027517
Carrera, M.G. and Rigby, J.K. 2004. Sponges, p. 102–111. In Webby, B.D., Droser, M., Paris F., and Percival, I. (eds.), The Great Ordovician Biodiversification Event. Columbia University Press.
Carrera, M.G., Astini, R.A., and Gomez, F. 2017. The lowermost Ordovician Tabulate-like coralomorph from the Precordillera of Western Argentina: main component of a reef-framework consortium. Journal of Paleontology, 91:73–85.
https://doi.org/10.1017/jpa.2016.145
Choh, S.-J., Hong, J., Sun, N., Kwon, S.-W., Park, T.-Y., Woo, J., Kwon, Y.K., Lee, D.-C., and Lee, D.-J. 2013. Early Ordovician reefs from the Taebaek Group, Korea: constituents, types, and geological implications. Geoscience Journal, 17:139–149.
https://doi.org/10.1007/s12303-013-0024-0
Church, S.B. 1974. Lower Ordovician patch reefs in western Utah. Brigham Young University Geology Studies, 21:41–62.
Conway, K.W., Krautter, M., Barrie, J.V., and Neuweiler, M. 2001. Hexactinellid sponge reefs on the Canadian continental shelf: A unique “living fossil”. Geoscience Canada 28:71–78.
de Laubenfels, M.W. 1955. Porifera, p. E21–E112. In Moore, R.C. (ed.), Treatise on Inver-tebrate Paleontology, Part E. Geological Society of America and University of Kansas Press, Lawrence.
Del Mouro, L., Lerosey-Aubril, R., Botting, J., Coleman, R., Gaines, R.R., Skabelund, J., Weaver, J.C., and Ortega-Hernández, J. 2024. A new sponge from the Marjum Formation of Utah documents the Cambrian origin of the hexactinellid body plan. Royal Society Open Science, 11 (231845).
Finks, R.M. 2003. Palaeozoic Hexactinellida: morphology and phylogeny, p.135–154. In Finks, R.M., Reid, R.E.H., and Rigby, J.K. (eds.), Treatise on Invertebrate Paleontology, Part E (Revised), Volume 2, Geological Society of America and the University of Kansas Press, Lawrence.
Finks, R.M. and Rigby, J.K. 2004a. Heteractinida, p. 557–583. In Kaesler, R.L. (ed.), Treatise on Invertebrate Paleontology, Part E (Revised), Volume 3. Geological Society of America and the University of Kansas Press, Lawrence.
Finks, R.M. and Rigby, J.K. 2004b. Paleozoic hexactinellid sponges, p. 320–448. In Kaesler, R.L. (ed.), Treatise on Invertebrate Paleontology, Part E (Revised), Volume 3. Geological Society of America and the University of Kansas Press, Lawrence.
Finks, R.M. and Rigby, J.K. 2004c. Paleozoic demosponges, p. 9–171. In Kaesler, R.L. (ed.), Treatise on Invertebrate Paleontology, Part E, Porifera (revised), Volume 3. Geological Society of America and University of Kansas, Boulder, Colorado and Lawrence, Kansas.
Hall, J. and Clarke, J.M. 1899. A memoir on the Paleozoic reticulated sponges constituting the family Dictyospongidae. Memoir of the New York State Museum 2 (for 1898): 1–350
Jarochowska, E. and Munnecke, A. 2014. The Paleozoic problematica Wetheredella and Allonema are two aspects of the same organism. Facies, 60:651–662.
https://doi.org/10.1007/s10347-014-0399-z
Jones, W.C. 1970. The composition, development, form and orientation of calcareous sponge spicules. Symposia of the Zoological Society of London, 25:91–123.
Keller, M. 1999. Argentine Precordillera. Sedimentary and Plate Tectonic History of a Laurentian Crustal Fragment in South America. GSA Special Paper 341, Boulder.
Keller, M. and Flügel, E. 1996. Early Ordovician reefs from Argentina: stromatoporoid vs. stromatolite origin. Facies, 34:177–192.
https://doi.org/10.1007/BF02546163
Keupp, H. and Schweigert, G. 2012. Neochoiaella n. gen. (Demospongeae, Choiaellidae) a second poriferan Lazarus taxon from the Solnhofen Plattenkalk (Upper Jurassic, southern Germany). Paläontologische Zeitschrift, 86:269–274.
https://doi.org/10.1007/s12542-012-0134-4
Kling, S.A. and Reif, W.-E. 1969. The Paleozoic history of amphidisc and hemidisc sponges: new evidence from the Carboniferous of Uruguay. Journal of Paleontology, 43:1429–1434.
Koenig, C.D.E. 1825. Icones Fossilium Sectiles. Centuria Prima. G. B. Sowerby, London.
Kozur, H.W., Mostler, H., and Repetski, J.E. 2008. A new heteractinellid calcareous sponge from the lowermost Ordovician of Nevada and a discussion of the suborder Heteractinellidae. Geo.Alp, 5:53–67.
Krautter, M. 2002. Fossil Hexactinellida: an overview. In Hooper, J.N.A. and van Soest, R.W.M. (eds.), Systema Porifera. A Guide to the Classification of Sponges. Kluwer Academics/Plenum Press, New York, pp. 1211–1223
Lee, J.-H., Chen, J., and Chough, S.K. 2015. The middle–late Cambrian reef transition and related geological events: A review and new view. Earth Science Review, 145:66–84.
https://doi.org/10.1016/j.earscirev.2015.03.002
Lee, J.-H. and Riding, R. 2018. Marine oxygenation, lithistid sponges, and the early history of Paleozoic skeletal reefs. Earth Science Review, 181:98–121.
https://doi.org/10.1016/j.earscirev.2018.04.003
Lehnert, O. and Keller, M. 1993. Posición estratigráfica de los arrecifes arenigianos en la Precordillera Argentina. Document Laboratoire Géologique Lyon, 125:263–275.
Leinfelder, R.R., Werner, W., Nose, M., Schmid, D.U., Krautter, M., Laternser, R., Takacs, M., and Hartmann, D. 1996, Paleoecology, growth parameters and dynamics of coral, sponge and microbolite reefs from the Late Jurassic. Göttinger Arbeiten zur Geologie und Paläontologie, 2:227–248.
Li, Q.J., Li, Y., Wang, J.P., and Kiessling, W. 2015. Early Ordovician lithistid sponge-Calathium reefs on the Yangtze Platform and their paleoceanographic implications. Palaeogeography, Palaeoclimatology, Palaeoecology, 425:84–96.
https://doi.org/10.1016/j.palaeo.2015.02.034
Mango, M.J. and Albanesi, G.L. 2020. Conodont biostratigraphy and correlation of the San Juan Formatio n at the Cerro La Silla section, middle Tremadocian - lower Dapingian, Central Precordillera, Argentina. Andean Geology, 47:577–598.
https://doi.org/10.5027/andgeoV47n3-3271
Mehl, D. and Reitner, J. 1996. Observations on Astraeospongium meniscum (Roemer, 1848) from the Silurian of western Tennessee: constructional morphology and palaeobiology of the Astraeospongiidae (Calcarea, Heteractinellidae). Berliner Geowissenschaften Abhandlungen, 18:243–255.
Mehl, D. and Lehnert, O. 1997. Cambro-Ordovician sponge spicule assemblages in the Ordovician of the Argentine Precordillera and paleoenvironmental ties. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, 204:221–246.
https://doi.org/10.1127/njgpa/204/1997/221
Miller, S.A. 1889. Class Porifera, p. 152–167. North American Geology and Paleontology. Western Methodist Book Concern, Cincinnati, Ohio.
Muir, L.A. and Botting, J.P. 2015. An outline of the distribution and diversity of Porifera in the Ordovician Builth Inlier (Wales, UK). Palaeoworld, 24:176–190.
https://doi.org/10.1016/j.palwor.2014.11.003
Nose, M., Vodrážka, R., Fernández, L.P., and Méndez-Bedia, I. 2013. First record of chambered hexactinellid sponges from the Palaeozoic. Acta Palaeontologica Polonica, 59:985–996.
https://doi.org/10.4202/app.2012.0112
Palmer, T. J. and Fürsich, F.T. 1981. Ecology of sponge reefs from the Upper Bathonian (Middle Jurassic) of Normandy. Palaeontology, 24:1–23.
Pisera A. 1997. Upper Jurassic siliceous sponges from the Swabian Alb: taxonomy and paleoecology. Palaeontologica Polonica, 57:1–216.
Pisera, A. 2004. What can we learn about siliceous sponges from palaeontology. Bollettino dei Musei e degli Istituti biologici dell'Università di Genova, 68:55–69.
Pisera, A. and Bodzioch, A. 1991. Middle Triassic lyssacinosan sponges from the Upper Silesia (southern Poland) and the history of hexactinosan and lychniscosan sponges. Acta Geologica Polonica, 41:193–207.
Rauff, H. 1895. Palaeospongiologie: Palaeontographica, 41:223–271
Reimann, I.G. 1945. New Middle Devonian octactinellids. Paleontological Contributions. Bulletin of the Buffalo Society of Natural Sciences, 19:16–21.
Rhebergen, F. 2007. Revision of the species Astraeospongium patina Roemer, 1861, based on Ordovician bedrock specimens from Estonia. GFF 129,17–22.
https://doi.org/10.1080/11035890701291017
Rigby, J.K. 1970. Two new Upper Devonian Hexactinellid sponges from Alberta. Journal of Paleontology, 44:7–16.
Rigby, J.K. 1986. Sponges of the Burgess Shale (Middle Cambrian) British Columbia. Palaeontographica Canadiana 2:1–105.
Rigby, J.K. and Nitecki, M.H. 1975. An unusually well preserved heteractinid sponge from the Pennsylvanian of Illinois and a possible classification and evolutionary scheme for the Heteractinida. Journal of Paleontology, 49:329–339.
Rigby, J.K. and Lenz, A.C. 1978. A new Silurian astylospongid sponge from Baillie-Hamilton Island, Canadian Arctic Archipelago. Canadian Journal of Earth Sciences, 15:157–162
Rigby, J. K. and Webby, B.D. 1988. Late Ordovician sponges from the Malongulli Formation of central New South Wales, Australia. Palaeontographica Americana, 56:1–147.
Rigby, J.K. and Collins, D. 2004. Sponges of the Middle Cambrian Burgess and Stephen Shale Formations, British Columbia. Royal Ontario Museum Contributions in Science, 1:1–164.
Rigby, J.K. 1991. Evolution of Paleozoic Heteractinid Calcareous Sponges and Demosponges-Patterns and Records, p. 83–101. In Reitner, J. and Keupp, H. (eds.), Fossil and Recent Sponges, Springer-Verlag.
Rigby, J.K., Nitecki, M., Zhu, Z., Liu, B., and Jiang, Y. 1995. Lower Ordovician reefs of Hubei, China, and the western United States, p. 423–428. In Cooper, J.D., Droser, M.L., and Finney, S., (eds.), Ordovician Odyssey: Seventh International Symposium on the Ordovician System, SEPM Pacific Section, Las Vegas.
Rigby, J.K., Pisera, A., Wrzoèek, T., and Racki, G. 2001. Upper Devonian sponges from the Holy Cross mountains, central Poland. Palaeontology. 44:447–488.
https://doi.org/10.1111/1475-4983.00187
Rowland, S.M. and Gangloff, R.A. 1988. Structure and paleoecology of Lower Cambrian reefs. Palaios, 3:111–135.
Rowland, S.M. and Shapiro, R.S. 2002. Reef patterns and environmental influences in the Cambrian and earliest Ordovician, p. 95–128. In Kiessling, W., Flügel, E., and Golonka, J. (eds.), Phanerozoic Reef Patterns. SEPM Spec. Pub. 72, Tulsa.
Senowbary-Daryan, B. and Amirhassankhani, F. 2012. Chambered hexactinellid sponges from Upper Triassic (Norian-Rhaetian?) reefs of Nayband Formation in central Iran. Rivista Italiana di Paleontologia e Stratigrafia, 118:247–260.
https://doi.org/10.13130/2039-4942/6004
Tomás, S., Aurell, M., Badenas, B., Borge, M., Duaso, M., and Mutti, M. 2019. Architecture and paleoenvironment of mid–Jurassic microbial–siliceous sponge mounds, northeastern Spain. Journal of Sedimentary Research, 89:110–134.
https://doi.org/10.2110/jsr.2019.5
Toomey, D.F. and Nitecki, M.H. 1979. Organic buildups in the Lower Ordovician (Canadian) of Texas and Oklahoma. Fieldiana, Geology, 2:1–181
Vacelet, J., Boury-Esnault, N., and Harmelin, J.G. 1994. Hexactinellid cave, a unique deep-sea habitat in the scuba zone. Deep Sea Research Part I: Oceanographic Research Papers, 41:965–973.
Webby, B.D. 2002. Patterns of Ordovician reef development, p. 129–179. In Kiessling, W., Flügel, E., and Golonka, J. (eds.), Phanerozoic Reef Patterns. SEPM Special Publication. 72, Tulsa.
Werner, W., Leinfelder, R.R., Fürsich, F.T., and Krautter, M. 1994. Comparative palaeoecology of marly coralline sponge-bearing reefal associations from the Kimmeridgian (Upper Jurassic) of Portugal and Southwestern Germany. Courier Forschungsinstitut Senckenberg, 172:381–397.
Zittel, 1877 (1876-1880). Handbuch der Palaeozoologie, v. 1, Protozoa, Coelenterata, Echinodermata, und Molluscoidea. München: Druck und Verlag von R. Oldenburg.
