

 A new giraffid Bramiscus micros nov. gen. nov. sp. (Ruminantia, Giraffidae) from the Miocene of northern Pakistan
A new giraffid Bramiscus micros nov. gen. nov. sp. (Ruminantia, Giraffidae) from the Miocene of northern Pakistan
Article number: 27.2.a29
https://doi.org/10.26879/1243
Copyright Society of Vertebrate Paleontology, June 2024
Author biographies
Plain-language and multi-lingual abstracts
PDF version
Submission: 14 September 2022. Acceptance: 23 May 2024.
ABSTRACT
We report new cranial, dental, and postcranial remains of a new early giraffid from the Middle Miocene of Pakistan: Bramiscus micros nov. gen. nov. sp. (Ruminantia, Giraffidae). The material comes from the fossil site Dhok Bun Amir Khatoon (Chinji Formation 14-11.4 Ma, Lower Siwaliks), located in the Chakwal district (Punjab, Pakistan). Additional remains were identified from the Hasvard Siwaliks Collection. The new findings show fused anterior ossicones, as in Bramatherium. The metatarsals possess a strong medial ridge and a weak lateral ridge, a feature encountered only in Bramatherium perimense. The size of our taxon is smaller than Bramatherium, hence the name. Bramiscus micros nov. gen. nov. sp. coexisted at the same time with other giraffids, such as Progiraffa exigua, Giraffokeryx punjabiensis, Giraffa priscilla, Giraffa punjabiensis, and other still unnamed taxa.
María Ríos. Departamento de Ciências da Terra, NOVA School of Science and Technology, Universidade Nova de Lisboa, GeoBioTec, 2829-516 Caparica, Portugal. maria.rios.iba@fct.unl.pt
Sayyed Ghyour Abbas. Department of Zoology, University of Sialkot, Daska Rd, near Victoria Palace, Beerh, Sialkot, Punjab, Pakistan. ghyour.kazmi@gmail.com; and Dr. Abu Bakr Fossil Display & Research Centre. Department of Zoology, University of the Punjab, Quaid-e-Azam Campus, Lahore, Punjab, Pakistan, 54590.
Muhammad Akbar Khan. Dr. Abu Bakr Fossil Display & Research Centre. Department of Zoology, University of the Punjab, Quaid-e-Azam Campus, Lahore, Punjab, Pakistan, 54590. akbar.zool.@pu.edu.pk
Nikos Solounias. Department of Anatomy, New York Institute of Technology College of Osteopathic Medicine, 8000 Northern Boulevard, Old Westbury, New York 11568, USA. mdanowit@nyit.edu and Department of Paleontology, American Museum of Natural History, Central Park West at 79th Street, New York, New York City 10024, USA.
Keywords: new species, new genus, Vertebrate; Mammalia; Giraffidae; Chinji; Miocene; Siwaliks
https://zoobank.org/74F8228C-796D-4939-86A5-87E7F530D8AA
Final citation: Ríos, María, Abbas, Sayyed Ghyour, Khan, Muhammad Akbar, and Solounias, Nikos. 2024. A new giraffid Bramiscus micros nov. gen. nov. sp. (Ruminantia, Giraffidae) from the Miocene of northern Pakistan. Palaeontologia Electronica, 27(2):a29.
https://doi.org/10.26879/1243
palaeo-electronica.org/content/2024/5228-new-giraffid-genus
Copyright: June 2024 Society of Vertebrate Paleontology.
This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
creativecommons.org/licenses/by/4.0
INTRODUCTION
Early Miocene Giraffidae are less diverse and abundant than during the later Miocene, with most of the evidence coming from the Chinji Formation of Pakistan (Ullah et al., 2006; Khan et al., 2009, 2013; Aftab et al., 2013; Mahmood et al., 2015). There are also giraffid fossils from Kalodirr, Loperot, and Moruorot Hill at the Lothidok Formation of West Turkana (Kenya), as well as Rusinga Island at the Hiwegi Formation (Kenya), and from Gebel Zelten (Libya) (Churcher, 1970; Hamilton, 1973, 1978; Grossman and Solounias, 2014). The only known Early Miocene genera are “Progiraffa” and Canthumeryx. By the Middle Miocene, there are several additional species (e.g., Giraffokeryx, Giraffa) and an adaptive radiation had occurred (Solounias, 2007), with several more giraffid taxa appearing in the Chinji Formation (Pakistan), in Fort Ternan (Kenya), as well as in Chios, Greece (Paraskevaidis, 1940; de Bonis et al., 1997) and more occurrences from Africa and Middle East (Harris et al., 2010). In the upper Chinji and younger Siwaliks there is an increase in species diversity with the appearance of large sivatherine giraffids Bramatherium, Libytherium, and Sivatherium (Aftab et al., 2016; Ríos et al., 2019, 2022a). Research on these occurrences is presented in an extensive number of studies, such as Lydekker (1883), Pilgrim (1911), Paraskevaidis (1940), Churcher (1970), Hamilton (1973, 1978), de Bonis et al. (1997), Barry et al. (2005), Geraads and Aslan (2003), Solounias (2007), and Aftab et al. (2016).
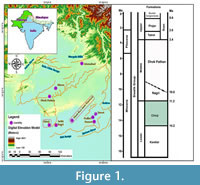 The present article deals with the description of a new genus and species from the Middle Miocene of the Chinji Formation. The holotype is a frontal bearing two anterior ossicones. It comes from the site of Dhok Bun Amir Khatoon, which contains the well exposed Chinji Formation deposits and is in the Chakwal district (Punjab, Pakistan) (Figure 1). The site has yielded two frontals and isolated teeth, as well as posterior ossicones. Additional metapodials were collected during the Harvard-GSP expeditions. The age of the fossil site is 14-11.4 Ma, while the metatarsal Y-GSP-14999 is of later age (9.286-9.265 Ma).
The present article deals with the description of a new genus and species from the Middle Miocene of the Chinji Formation. The holotype is a frontal bearing two anterior ossicones. It comes from the site of Dhok Bun Amir Khatoon, which contains the well exposed Chinji Formation deposits and is in the Chakwal district (Punjab, Pakistan) (Figure 1). The site has yielded two frontals and isolated teeth, as well as posterior ossicones. Additional metapodials were collected during the Harvard-GSP expeditions. The age of the fossil site is 14-11.4 Ma, while the metatarsal Y-GSP-14999 is of later age (9.286-9.265 Ma).
Geological Setting
The Siwalik Group contains a rich vertebrate fossil succession (Lydekker, 1883; Colbert, 1935; Barry et al., 2002; Nanda, 2002; Khan et al., 2009, 2010; Aftab et al., 2016) and is stratigraphically divided into the Lower, Middle, and Upper Siwalik subgroups (Barry et al., 2002). The Lower Siwalik Subgroup (Kamlial and Chinji formations) corresponds with the Astaracian and Vallesian mammall ages (18-11.4 Ma) (Barry et al., 2013).
The Dhok Bun Amir Khatoon outcrops represent fluvial deposits of the Chinji Formation (Lower Siwaliks). They are situated 52.48 km East from the Chinji stratotype (Chakwal district, Pakistan, Figure 1) (Aftab et al., 2016). The geographical coordinates of the site are Lat. 32°47'N, Long. 72°55'E. The outcrops comprise shales, silt/mudstones, and sandstones deposited in a fluvial environment, mainly filled by unweathered igneous minerals. The shale and silts/mudstones are brick red, a characteristic of the Chinji Formation at the type section. The silts/mudstones comprise about 70% of the outcrop, while the sandstone only represents about 30%. The sandstone is dull to bright gray, and mostly present as grit. At certain places bright or brick red colored conglomerates and breccia are interbedded between the silt/mud layers. This lithology corresponds to the Chinji type section. In the Chinji Rest House area, conglomerates are thicker than the well-exposed sediments at Dhok Bun Amir Khatoon. Based on lithological and biostratigraphic data (see Johnson et al., 1985; Cheema et al., 1997; Cheema, 2003; Khan et al., 2008, 2009, 2013, 2017), the fossil bearing section of the site has been dated as Middle Miocene-early Late Miocene. The biostratigraphic range of the included faunal assemblage suggests an age between 14 and 11.4 Ma (Cheema, 2003; Khan et al., 2008, 2011, 2013, 2017; Barry et al., 2013).
This paper also includes a unique metatarsal described from the later Dhok Pathan Formation (Upper Miocene). The type section of the Dhok Pathan Formation is present around the Dhok Pathan Rest House, near the Dhok Pathan village at the bank of Soan River, Chakwal, Pakistan (Pilgrim, 1911; Pilbeam et al., 1977, 1980). Like the Chinji stratotype, Dhok Pathan stratotype is also over 10 km long (Pilgrim, 1911). This formation is characterized by the cyclic deposition of the varied colored sandstone and light-yellow clay and between these two strata, conglomerate is sandwiched or lenses out (Pilgrim, 1911; Barry et al., 2002, 2013). The Dhok Pathan Formation determined age is 9.8 to ca. 3.5 Ma (Barry et al., 2013).
Institutional/Technical Abbreviations
AMNH: American Museum of Natural History (New York, USA)
GSI-B: Geological Survey of India - Brown Collection
GSP-H: Geological Survey of Pakistan - Howard University Project
GSP-Y, Y-GSP: Geological Survey of Pakistan - Harvard University Project
GSP-S: Geological Survey of Pakistan - Sind collection
PUPC: Punjab University Palaeontological Collection, Lahore, Pakistan
PU: Punjab University, Lahore, Pakistan
MATERIALS AND METHODS
Materials
The material comprises 13 specimens including the frontals with ossicones, isolated ossicones, isolated upper and lower premolars and molars, and a mandibular fragment. Y-GSP material is of known age (Johnson et al., 1985, 1988; McRae, 1990; Badgley et al., 1998).
Repository
The specimens are stored at the Dr. Abu Bakr Fossil Display and Research Centre, University of the Punjab, Lahore, Pakistan (PUPC IDs) and Harvard Peabody Museum, USA (Y-GSP IDs). The material of the new species is also represented in the Harvard-GSP project (Y-GSP) collections and belongs to the Geological Survey of Pakistan. While the final deposition of the fossils is uncertain, they currently are all at Harvard Peabody Museum in the Department of Human Evolutionary Biology’s Paleoanthropology Lab.
Methods
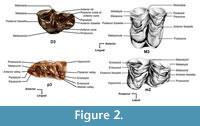 Measurements and terminology. We took all measurements with digital calipers. We follow the measurements and terminology proposed for ruminants by Hamilton (1973) and by Bärmann and Rössner (2011) (Figure 2). The measurements used to compare the material were collected during visits to the American Museum of Natural History (New York, USA), the Institut Català de Paleontologia-Miquel Crusafont (Barcelona, Spain), the Museo Nacional de Ciencias Naturales-CSIC (Madrid, Spain), the Musèum National d’Histoire Naturelle (Paris, France), and the Natural History Museum (London, UK). The rest of the morphological and biometrical information in this paper was collected from several publications (Gaudry, 1861; Rodler and Weithofer, 1890; Pilgrim, 1911; de Mecquenem, 1924; Bohlin, 1926; Dietrich, 1937, 1942; Singer and Boné, 1960; Churcher, 1970, 1978; Hendey, 1970; Hamilton, 1973, 1978; Μelentis, 1974; Heintz et al., 1981; Brunet and Heintz, 1983; Harris, 1976a, b, 1991; Kostopoulos et al., 1996; de Bonis et al., 1997; Geraads, 1994, 2009; Kostopoulos, 2009; Robinson, 2011).
Measurements and terminology. We took all measurements with digital calipers. We follow the measurements and terminology proposed for ruminants by Hamilton (1973) and by Bärmann and Rössner (2011) (Figure 2). The measurements used to compare the material were collected during visits to the American Museum of Natural History (New York, USA), the Institut Català de Paleontologia-Miquel Crusafont (Barcelona, Spain), the Museo Nacional de Ciencias Naturales-CSIC (Madrid, Spain), the Musèum National d’Histoire Naturelle (Paris, France), and the Natural History Museum (London, UK). The rest of the morphological and biometrical information in this paper was collected from several publications (Gaudry, 1861; Rodler and Weithofer, 1890; Pilgrim, 1911; de Mecquenem, 1924; Bohlin, 1926; Dietrich, 1937, 1942; Singer and Boné, 1960; Churcher, 1970, 1978; Hendey, 1970; Hamilton, 1973, 1978; Μelentis, 1974; Heintz et al., 1981; Brunet and Heintz, 1983; Harris, 1976a, b, 1991; Kostopoulos et al., 1996; de Bonis et al., 1997; Geraads, 1994, 2009; Kostopoulos, 2009; Robinson, 2011).
Maximum parsimony analysis. We used the character list from Ríos et al. (2017) and included the Bramiscus micros nov. gen. nov. sp. data into their matrix. The data matrices including the new data are presented as supporting information (Appendix 1). We compiled the data matrices using Mesquite 3.04 (Windows version). We performed a Maximum Parsimony analysis to check the phylogenetic position of Bramiscus micros nov. gen. nov. sp. within the Giraffidae. The resulting dataset consists of 111 characters (cranial, dental, and postcranial), and 32 taxa. We used TNT v1.5-beta software (Goloboff et al., 2008) to analyze the dataset. All characters are non-additive and unweighted. We analyzed the trees using a Traditional Search method (heuristic algorithm) with Tree Bisection Reconnection (TBR) and 1000 replicates (holding the 10 most parsimonious trees for each replicate). We used the Bremer included in the script (BREMER.RUN) to assess branch support.
SYSTEMATIC PALAEONTOLOGY
Superfamily GIRAFFOIDEA Gray, 1821
Family GIRAFFIDAE Gray, 1821
Genus BRAMISCUS nov. gen.
zoobank.org/ECF5AE35-4F43-491F-8C9C-117AD2DAADCA
BRAMISCUS MICROS nov. sp.
zoobank.org/EBEC061A-707C-402E-A3C1-43EB04486CA5
 Diagnosis. Small-sized giraffid with probably two pairs of ossicones. The ossicones of the anterior pair are fused at their base (Bramatherium -like). The posterior pair is straight and possibly directed laterally. All ossicones have an oval cross-section at the base and are vaguely circular at the apex. The ossicone surface is covered with longitudinal ridges of medium depth. The ridges or grooves are deeper and larger distally on the apical part and laterally than dorsally on the proximal part and medially. The dentition is brachydont and shows a rugose enamel. The upper molars have a postmetaconule fold and a labial cingulum, as well as a developed entostyle. The lower p2 has a crest on the anterior valley. The lower p3 has a well-developed paraconid and parastylid. The metaconid is narrower than the entoconid. The lower molars have a slightly rounded protoconid and hypoconid. Labially, they have rugose enamel and an anterolabial cingulum. A Palaeomeryx fold of the lower molars is absent (sensu Sánchez et al., 2011). The lower molars have a high L/W index of ~2. The metatarsal has a characteristic large medial ridge and a low lateral ridge.
Diagnosis. Small-sized giraffid with probably two pairs of ossicones. The ossicones of the anterior pair are fused at their base (Bramatherium -like). The posterior pair is straight and possibly directed laterally. All ossicones have an oval cross-section at the base and are vaguely circular at the apex. The ossicone surface is covered with longitudinal ridges of medium depth. The ridges or grooves are deeper and larger distally on the apical part and laterally than dorsally on the proximal part and medially. The dentition is brachydont and shows a rugose enamel. The upper molars have a postmetaconule fold and a labial cingulum, as well as a developed entostyle. The lower p2 has a crest on the anterior valley. The lower p3 has a well-developed paraconid and parastylid. The metaconid is narrower than the entoconid. The lower molars have a slightly rounded protoconid and hypoconid. Labially, they have rugose enamel and an anterolabial cingulum. A Palaeomeryx fold of the lower molars is absent (sensu Sánchez et al., 2011). The lower molars have a high L/W index of ~2. The metatarsal has a characteristic large medial ridge and a low lateral ridge.
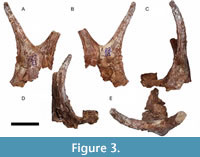
Holotype. PUPC 22/01, fragment of frontal bone bearing a pair of ossicones with their fused bases (Figure 3).
Paratype. PUPC 13/375, pair of ossicones with their fused bases (Figure 4).
Etymology. Bramiscus refers to the morphological similarities between the ossicones of Bramiscus and Bramatherium, and means small Bramatherium: -iscus means very small in Greek. Micros also means small.
Geographic distribution and age. Chinji Formation (14 Ma, Astaracian) to Dhok Pathan Formation (9.265 Ma, Vallesian), Lower to Middle Siwaliks, Punjab, Pakistan.
Anatomical Description
 Ossicones. PUPC 22/01 is a fragment of an ossicone-bearing frontal bone fragment (Figure 3). The right ossicone is complete while the left is broken in the middle. Both ossicones are fused at the base like in the paratype PUPC 13/375 (Figure 4). The maximum preserved length of the ossicone fused bases is 127.90 mm and that of width is 92.95 mm. The fused bases form a convex structure anteriorly and mostly flat but a bit concave posteriorly. The ossicones are tilting laterally, hence they form a V-shape in rostral view. Their fused bases form an angle of approximately 110º with each other. They both have deep grooves running over the surface, but the lateral groove is extremely deep. The ossicones are straight with a slight twist that can be seen from the running grooves as well. As the ossicones move in opposite directions from their base to their apex, and away from the sagittal plane of the frontal, they create an outward orientation. Small bumps are present over the surface of the ossicones, especially the right one. The cross section at the free distal base is oval, gradually becoming circular towards the apex, and the anteroposterior and transversal diameters (DAP and DT) of the ossicones also decrease towards the apex. There is no hint of a suture line, probably indicating that the individual was fully adult. Small bumps on the ossicones indicate that they belong to an older individual as it has been commonly observed in many giraffid genera including Palaeomeryx, Schansitherium, Decennatheirum, and Giraffa, where the number of bumps increase with age, and are only found in adult individuals (Ríos and Solounias, 2019, Ríos et al., 2022b).
Ossicones. PUPC 22/01 is a fragment of an ossicone-bearing frontal bone fragment (Figure 3). The right ossicone is complete while the left is broken in the middle. Both ossicones are fused at the base like in the paratype PUPC 13/375 (Figure 4). The maximum preserved length of the ossicone fused bases is 127.90 mm and that of width is 92.95 mm. The fused bases form a convex structure anteriorly and mostly flat but a bit concave posteriorly. The ossicones are tilting laterally, hence they form a V-shape in rostral view. Their fused bases form an angle of approximately 110º with each other. They both have deep grooves running over the surface, but the lateral groove is extremely deep. The ossicones are straight with a slight twist that can be seen from the running grooves as well. As the ossicones move in opposite directions from their base to their apex, and away from the sagittal plane of the frontal, they create an outward orientation. Small bumps are present over the surface of the ossicones, especially the right one. The cross section at the free distal base is oval, gradually becoming circular towards the apex, and the anteroposterior and transversal diameters (DAP and DT) of the ossicones also decrease towards the apex. There is no hint of a suture line, probably indicating that the individual was fully adult. Small bumps on the ossicones indicate that they belong to an older individual as it has been commonly observed in many giraffid genera including Palaeomeryx, Schansitherium, Decennatheirum, and Giraffa, where the number of bumps increase with age, and are only found in adult individuals (Ríos and Solounias, 2019, Ríos et al., 2022b).
PUPC 13/375 bears two ossicones (Figure 4) that are fused at their base. The fusion forms a long plate of bone. From a proximal view the portion of the ossicones bases has an irregular surface. It is divided into three areas with two lateral triangular concave areas and a deeper middle section. The estimated preserved length of the ossicones is 175 mm from the base of the skull. The ossicones form an angle with the skull fragment of approximately 90º (practically straight angle). The ossicones are straight. The section at the ossicone base is oval (Figure 4H), with the long axis of the oval oriented approximately 34º outwards from the sagittal plane of the fragment. The section becomes slightly semicircular towards the apex of the ossicone, with a flattened anteromedial side. The ossicones narrow gradually towards the apex. The surface of the ossicone is rugose, ornamented with longitudinal grooves that run along all the surface of the ossicone, with the lateral groove being also deeper than the rest.
Although a complete cranium has not been found, there is a high possibility that the isolated ossicones found at the site belong to the same taxon as they share a similar proportions and morphology. The isolated ossicones found at Dhok Bun Amir Khatoon (Figure 4G) resemble the morphology of the posterior ossicones of Bramatherium, and as the anterior ossicones also resemble Bramatherium, we conclude that there is a high probability that these specimens represent the broken-off posterior ossicones belonging to the frontals found at the same site. However, it is important to note that there is no direct evidence to support this claim. While the new taxon may have similar anterior ossicones, it cannot be assumed that it had the exact same posterior ossicone morphology. However, being similar in size, the presumed posterior ossicones are also described here. They are straight and contain compact bone as in Bramatherium. The surface is smooth. and they contain two canals separated internally by a thin plate of bone. One canal is bigger than the other. The ossicones terminate in a small knob.
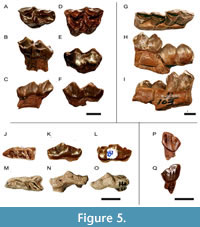 Upper dentition. The available specimens are all isolated teeth. The DP3 (Figure 5A-C) is bilobed, slightly molarized, and longer than wide. In occlusal view the anterior lobe is longer and semicircular, and the posterior lobe is shorter, wider, and triangular in shape. The enamel is smooth and shiny lingually and more rugose labially. The parastyle, mesostyle, and metastyle are thick basally but narrow towards the apices. Labially, the stylids are strong. The paracone and metacone are strongly developed. Lingually, strong cingulum is present at the base of the protocone whereas a weak cingulum is present posteriorly. The posterior fossette is triangular. A shallow median valley is present. The postprotocrista reaches the premetaconulecrista. The premetaconulecrista is prominent.
Upper dentition. The available specimens are all isolated teeth. The DP3 (Figure 5A-C) is bilobed, slightly molarized, and longer than wide. In occlusal view the anterior lobe is longer and semicircular, and the posterior lobe is shorter, wider, and triangular in shape. The enamel is smooth and shiny lingually and more rugose labially. The parastyle, mesostyle, and metastyle are thick basally but narrow towards the apices. Labially, the stylids are strong. The paracone and metacone are strongly developed. Lingually, strong cingulum is present at the base of the protocone whereas a weak cingulum is present posteriorly. The posterior fossette is triangular. A shallow median valley is present. The postprotocrista reaches the premetaconulecrista. The premetaconulecrista is prominent.
The M1 (Figure 5D-F) has a labial wall with stronger parastyle and mesostyle and a slightly weaker metastyle. The paracone and metacone are very strong. Lingually, the protocone and metaconule are V-shaped, with the posterior lobe slightly buccolingually wider than the anterior one. The anterior fossette is semicircular whereas the posterior fossette is crescent shaped. The transverse valley represents a weak entostyle. The preprotocrista is more elongated than the postprotocrista. The enamel is rugose and there is a lingual cingulum.
Lower dentition. The available specimens consist of a canine, isolated premolars, and a mandible fragment with well-preserved m1 and m2. The canine (Figure 5P-Q) has a pointy larger anterior lobe while the smaller posterior lobe is rounded and represents 45% of the total width of the tooth.
The p2, is a simple tooth, elongated and with a slightly pointed protoconid. The metaconid and the entoconid are separated. The anterior lingual valley is wider than the posterior one and there is a crest in it (Figure 5J-L).
The p3 (Figure 5M-O) shows a well-developed paraconid and parastylid. The paraconid is well distinguished from the parastylid and compressed laterally. The metaconid is posteriorly oriented and is separated from the entoconid, which is also posteriorly projected. The postparacristid is wider than the extremely thin preparacristid and they are oriented lingually. The metaconid is strong and coniform. The hypoconid is prominent.
The lower molars (Figure 5J-L) are brachydont with sharp and pointed conids. They have a lingual wall with prominent entostylid, mesostylid, and metastylid. The stylids are broader at the base and gradually narrow towards the tip. Labially, there is a short and robust ectostylid. The anteroposterior cingulum is present. The Palaeomeryx -fold is absent (Sánchez et al., 2010). The fossettes are crescent shaped and only slightly pointed.
 Metatarsal. Y-GSP-14999 is a well-preserved right proximal metatarsal (Figure 6). In proximal view there is an asymmetry between the lateral and the medial profiles. The kidney-shaped articulation facet for the ectomesocuneiform is similar in size to the semicircular navicular-cuboid facet. The entocuneiform facet is relatively large and subtriangular, and it does not contact the ectomesocuneiform facet. The synovial fossa is oval and deep. The medial epicondyle is large with a large plantar head, the distal aspect of which continues onto the medial ridge. The lateral epicondyle is split into a dorsal and plantar head by a deep longitudinal groove on the lateral shaft. The plantar head is continuous distally with the weak lateral ridge. Both heads of the lateral and medial epicondyles are oriented longitudinally, however the dorsal heads slightly flare outward. The medial ridge is high and rounded and the lateral ridge is sharp and thinner. The central trough is intermediate in depth, and it flattens distally. The most distinctive feature is the presence of an acute asymmetry in the medial and lateral diaphysis plantar ridges. Y-GSP-1196 shares the same morphology, with a high and wide medial ridge while the lateral is thinner and lower.
Metatarsal. Y-GSP-14999 is a well-preserved right proximal metatarsal (Figure 6). In proximal view there is an asymmetry between the lateral and the medial profiles. The kidney-shaped articulation facet for the ectomesocuneiform is similar in size to the semicircular navicular-cuboid facet. The entocuneiform facet is relatively large and subtriangular, and it does not contact the ectomesocuneiform facet. The synovial fossa is oval and deep. The medial epicondyle is large with a large plantar head, the distal aspect of which continues onto the medial ridge. The lateral epicondyle is split into a dorsal and plantar head by a deep longitudinal groove on the lateral shaft. The plantar head is continuous distally with the weak lateral ridge. Both heads of the lateral and medial epicondyles are oriented longitudinally, however the dorsal heads slightly flare outward. The medial ridge is high and rounded and the lateral ridge is sharp and thinner. The central trough is intermediate in depth, and it flattens distally. The most distinctive feature is the presence of an acute asymmetry in the medial and lateral diaphysis plantar ridges. Y-GSP-1196 shares the same morphology, with a high and wide medial ridge while the lateral is thinner and lower.
RESULTS
The morphology and proportions of the specimens from Dhok Bun Amir Khatoon (Chinji) analyzed in this article appear to indicate that they belong to a new Middle Miocene giraffid taxon: Bramiscus micros nov. gen. nov. sp.
Ossicones
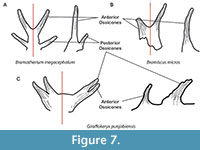 Orientation and Curvature. The ossicones of PUPC 13/375 are only slightly, and moderately in PUPC 22/01, posterolaterally oriented, while the other early taxa as Giraffokeryx, Canthumeryx, Injanatherium, and Georgiomeryx have ossicones that are oriented more laterally, with a much higher divergence angle (Ríos et al., 2017). The ossicones of PUPC 13/375 and PUPC 22/01 show barely any curvature towards the apex, while this curvature is marked in Giraffokeryx (Colbert, 1933), and Decennatherium asiaticum (Ríos et al., 2019). Also, the ossicones of Giraffokeryx are directed laterally and not vertically as in Bramiscus. The long transverse axis of the ossicone is oriented approximately 34º outwards from the sagittal plane in Bramiscus while in Giraffokeryx the same angle is 52º (see Colbert, 1933) (Figure 7). Interestingly, the orientation and space between the ossicones of Bramiscus is like that found in Bramatherium megacephalum.
Orientation and Curvature. The ossicones of PUPC 13/375 are only slightly, and moderately in PUPC 22/01, posterolaterally oriented, while the other early taxa as Giraffokeryx, Canthumeryx, Injanatherium, and Georgiomeryx have ossicones that are oriented more laterally, with a much higher divergence angle (Ríos et al., 2017). The ossicones of PUPC 13/375 and PUPC 22/01 show barely any curvature towards the apex, while this curvature is marked in Giraffokeryx (Colbert, 1933), and Decennatherium asiaticum (Ríos et al., 2019). Also, the ossicones of Giraffokeryx are directed laterally and not vertically as in Bramiscus. The long transverse axis of the ossicone is oriented approximately 34º outwards from the sagittal plane in Bramiscus while in Giraffokeryx the same angle is 52º (see Colbert, 1933) (Figure 7). Interestingly, the orientation and space between the ossicones of Bramiscus is like that found in Bramatherium megacephalum.
Ornamentation. The surface of the ossicone is rugose, but not as rugose as in Giraffokeryx, and Decennatherium, and not as smooth as in Samotherium (faint ridges) or Palaeotragus (no ridges). These ridges in PUPC 13/375 and PUPC 22/01 are deeper and larger on the distal and lateral sides as in Bramatherium.
Size. The estimated length of the anterior ossicones from the base of the skull is small, being similar in length to some Giraffa specimens, although both show significant morphological differences, e.g., Giraffa demonstrates greater girth than Bramiscus (Solounias, 2007). They are similar in size to the anterior ossicones of Giraffokeryx. The dimensions of the ossicones of PUPC 13/375 and PUPC 22/01 also exclude most giraffids that bear larger ossicones as Decennatherium, Birgerbohlinia, Bramatherium, and Sivatherium as this reaches lengths of 5x the length of Bramiscus (Ríos et al., 2017). They are also very different in size and morphology when compared to the bovid horns with marked keels found in the same locality.
Morphology of the ossicone bases. The bases of the ossicones in PUPC 13/375 and PUPC 22/01 are fused forming a structure that connects the ossicones with the frontal. This creates a space between the skull and the flaring part of the ossicones. Such a condition is found in two giraffid genera where the bases of the ossicones are fused, Bramatherium, and Giraffokeryx; one palaeomerycid, Prolibytherium; and three bovid genera, Tsaidamotherium, Plesiaddax, and Urmiatherium (Falconer, 1845; Bettington, 1846; Rodler, 1889; Colbert, 1933; Bohlin, 1935; Lewis, 1939; Kostopoulos, 2009; Jafarzadeh et al., 2012; Shi et al., 2016). However, only Bramatherium has a large structure formed by fused bases, whereas in Giraffokeryx only a small portion is formed by the fusion of the ossicone bases and the space between the ossicones and frontal is very small (Figure 7). Prolibytherium also shows the fused ossicone bases, but these bases are very close, even merging, with the frontal bone. The bovid genera also show a partial fusion of horn core bases, but it is not as great as Bramatherium or Bramiscus.
To summarize, the anterior ossicones of Bramiscus narrow gradually towards the apex and have the same “V” shape as the ossicones of Bramatherium megacephalum (e.g., YPM-13881) and Bramatherium perimense. The ossicones are fused at their base similarly to the figured anterior ossicones of B. megacephalum (Lewis, 1939: plate 2, YPM-13881), and B. perimense (AMNH-27016; Falconer, 1845; Bettington, 1846: plate 1 MRCS 1436 and NMHUK 20009 cast; Colbert, 1935, fig. 174, AMNH 19771; Lydekker, 1876, plate 7, fig. 13, AMNH-27016). The possible posterior ossicones of Bramiscus are straight as the posterior ossicones of Bramatherium perimense AMNH 19688. The posterior ossicones of B. megacephalum also share with Bramiscus the presence of two internal canals.
Dentition
Differences with primitive Giraffomorpha. Our specimens also differ from other primitive Giraffomorpha such as Propalaeoryx, Climacoceras, Prolibytherium and Nyanzameryx by the presence of a more prominent and well-separated metastylid in the p3 (Sánchez et al., 2010). Climacoceras has narrower upper molars and in Nyanzameryx and Prolibytherium the lingual conids are compressed and the lingual wall of the hypoconulid is incomplete in the m3 (Barry et al., 2005).
Due to the high morphological stability in the Giraffidae (Hamilton, 1978; Solounias, 2007) the dentition of Bramiscus micros nov. gen. nov. sp. differs little in morphology and size to the dentition of other early giraffids, such as “Progiraffa exigua”, Canthumeryx sirtensis, or Giraffokeryx punjabiensis, as they sport small, simple teeth. Despite this, there are some morphological differences, especially regarding the p3, which is one of the most diagnostic teeth in giraffids (Hamilton, 1978; Ríos et al., 2016a, 2017).
p3. Regarding the p3 Length/Width index (L/W index), the remains from Dhok Bun Amir Khatoon show average values of 2.06 (N=3), with a minimum value of 1.86 and a maximum of 2.37. These are very close to the average values of most of the other Early and Middle Miocene giraffids “Progiraffa exigua”, Canthumeryx sirtensis, and Georgiomeryx georgalasi and higher than other early giraffids such as Injanatherium arabicum, which never reaches a L/W value higher than 1.74. Crown giraffids from the late Miocene and the Plio-Pleistocene show lower L/W index values (Ríos et al., 2017).
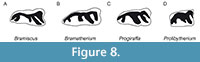 Morphologically, the p3 PUPC 14/143 is like “Progiraffa exigua” specimen S 412 from Locality S 2 at the Zinda Pir Dome (upper unit of the Chitarwata Formation and the overlying Vihowa Formation, 20 to 17 Ma, Pakistan, Barry et al., 2005: fig. 9:1; Lindsay et al., 2005), although in S 412, the metaconid, entoconid, and posterior stylid are more parallel. The “Progiraffa exigua” specimen Y 31797 (Barry et al., 2005: fig. 9:2) from the Locality Y 747 (base of the Potwar sequence, ca. 18.3, Pakistan) is smaller and does not have a developed posterolabial lobe. However, is important to recall that the basicranium ascribed to “Progiraffa exigua” by Barry et al. (2005), was later used to establish the new taxon Prolibytherium fusus (Danowitz et al., 2016) so the teeth taken as “Progiraffa” by Barry et al. (2005) may belong to Prolibytherium fusus as well, although this needs further analysis. The premolars ascribed by Aftab et al. (2016) to “Progiraffa exigua” are also questionable, and they come from the same formation as the cranial appendages of Bramiscus. The original “Progiraffa exigua” material is older, as it comes from the earlier Kamlial Formation (ca 18.3-14.2 Ma). Canthumeryx sirtensis has a less developed entoconid on the p3, and Giraffokeryx punjabiensis has a p3 with a more developed metaconid that projects posteriorly and an entoconid that reaches the lingual side of the tooth (Figure 8, Hamilton, 1978). Georgiomeryx georgalasi from the early Middle Miocene of Thymiana (Greece) (de Bonis et al., 1997). This species shares similar size and proportions with Bramiscus specimens, particularly PUPC 11/141. However, it can be distinguished by the absence of the metaconid and a less parallel entoconid and entostylid.
Morphologically, the p3 PUPC 14/143 is like “Progiraffa exigua” specimen S 412 from Locality S 2 at the Zinda Pir Dome (upper unit of the Chitarwata Formation and the overlying Vihowa Formation, 20 to 17 Ma, Pakistan, Barry et al., 2005: fig. 9:1; Lindsay et al., 2005), although in S 412, the metaconid, entoconid, and posterior stylid are more parallel. The “Progiraffa exigua” specimen Y 31797 (Barry et al., 2005: fig. 9:2) from the Locality Y 747 (base of the Potwar sequence, ca. 18.3, Pakistan) is smaller and does not have a developed posterolabial lobe. However, is important to recall that the basicranium ascribed to “Progiraffa exigua” by Barry et al. (2005), was later used to establish the new taxon Prolibytherium fusus (Danowitz et al., 2016) so the teeth taken as “Progiraffa” by Barry et al. (2005) may belong to Prolibytherium fusus as well, although this needs further analysis. The premolars ascribed by Aftab et al. (2016) to “Progiraffa exigua” are also questionable, and they come from the same formation as the cranial appendages of Bramiscus. The original “Progiraffa exigua” material is older, as it comes from the earlier Kamlial Formation (ca 18.3-14.2 Ma). Canthumeryx sirtensis has a less developed entoconid on the p3, and Giraffokeryx punjabiensis has a p3 with a more developed metaconid that projects posteriorly and an entoconid that reaches the lingual side of the tooth (Figure 8, Hamilton, 1978). Georgiomeryx georgalasi from the early Middle Miocene of Thymiana (Greece) (de Bonis et al., 1997). This species shares similar size and proportions with Bramiscus specimens, particularly PUPC 11/141. However, it can be distinguished by the absence of the metaconid and a less parallel entoconid and entostylid.
The p3 of Bramatherium is much larger and has a lower L/W index but retains an almost identical morphology otherwise.
Lower molars. Concerning the lower molars, they are simple and small, like those of “Progiraffa exigua” (Barry et al., 2005: fig. 10:1-5), but in P. exigua the m1, m2, and m3 anterior and posterior lobes are labially more pointed and with a more acute V-shape (Figure 5). They are also smaller in size and differ in their L/W Index as “P. exigua” has a much lower index than Bramiscus specimen PUPC 11/102 (m1: 1.66 vs 2.04; m2: 1.51 vs. 2.00; Pilgrim, 1911; Barry et al., 2005).
The lower molars of Canthumeryx sirtensis, Georgiomeryx georgalasi, Injanatherium arabicum, Giraffokeryx primaevus, and Giraffokeryx punjabiensis and the late Miocene Palaeotragus rouenii, have a similar size but are slightly larger and have lower L/W values than the new specimens from Dhok Bun Amir Khatoon. The other late Miocene and Plio-Pleistocene giraffids all have much larger lengths.
Giraffa priscilla, another one of the middle Miocene giraffid found in the area (based on a single tooth), shows more brachydont teeth and differs also in several upper molar characters (e.g., plus upper molars with lower L/W index values, smaller anterior and posterior fossettes and a rounder metaconule and protocone) (Matthew, 1929; Bhatti et al., 2012; Aftab et al., 2013, 2016).
Metatarsals. As mentioned above, the most distinctive feature is the presence of a very particular and acute asymmetry in the medial and lateral diaphysis ridges, only seen so far in Bramatherium (Ríos et al., 2016). Y-GSP-14999 has a high and wide medial ridge while the lateral is thinner and lower. The same is true for Bramatherium perimense AMNH 19688 (Figure 6). However, Bramatherium has longer and less gracile metapodials (Ríos et al., 2016). In Giraffokeryx, the metatarsal (unpublished data) has two ridges that are of equal development, while Giraffa has very elongated metapodials, which exclude them as possible matches for these specimens.
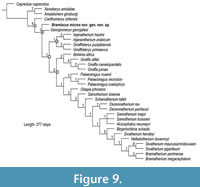 Results of the phylogenetic analysis. Our data matrix (Appendix 1) includes 32 taxa and 111 characters (cranial, dental, mandibular, and postcranial). The Maximum Parsimony (MP) search produced three Most Parsimonious Trees (MPT), which resulted in a consensus tree (Strict, Nielsen) of 277 steps (Figure 9) and recovered a monophyletic Giraffidae (node A). The most basal off-shoot is the stratigraphically older Canthumeryx sirtensis, followed by Bramiscus micros and Georgiomeryx georgalasi (node B). The remaining taxa (node C) are grouped into two clades: a group including Injanatherium hazimi, Injanatherium arabicum, Giraffokeryx punjabiensis, and Giraffokeryx primaevus (node D), and the large group of younger giraffids (node E). Our resulting MPT matches the results of Ríos et al. (2017).
Results of the phylogenetic analysis. Our data matrix (Appendix 1) includes 32 taxa and 111 characters (cranial, dental, mandibular, and postcranial). The Maximum Parsimony (MP) search produced three Most Parsimonious Trees (MPT), which resulted in a consensus tree (Strict, Nielsen) of 277 steps (Figure 9) and recovered a monophyletic Giraffidae (node A). The most basal off-shoot is the stratigraphically older Canthumeryx sirtensis, followed by Bramiscus micros and Georgiomeryx georgalasi (node B). The remaining taxa (node C) are grouped into two clades: a group including Injanatherium hazimi, Injanatherium arabicum, Giraffokeryx punjabiensis, and Giraffokeryx primaevus (node D), and the large group of younger giraffids (node E). Our resulting MPT matches the results of Ríos et al. (2017).
DISCUSSION
The ossicones found from Dhok Bun Amir Khatoon (Pakistan) and Chinji are the first ones recovered with this morphology. The recently identified species is smaller in size, yet it shares similarities with Bramatherium in terms of its cranial appendage’s distinctive V-shaped morphology. Additionally, it exhibits a notable and sharp asymmetry in the medial and lateral diaphysis ridges of its metatarsals, akin to that observed in Bramatherium. Therefore, and despite Bramiscus micros nov. gen. nov. sp. pertaining to a much more basal clade of giraffids, we have named this new genus after Bramatherium because of the morphological similarity of its ossicones to those of the younger and larger Bramatherium species also found in Pakistan. The anterior ossicones are shaped and fused in a similar way. Due to the age of the material, as well as the presence of bumps in the ossicones, a feature related to ontogenetic development in certain giraffids and only found in adult individuals (Ríos et al., 2020) we don’t think that the frontals with ossicones belong to a young Bramatherium.
We acknowledge the similarities with Bramatherium, and that the trichotomy in our cladogram is potentially an issue of missing data. However, the estimated length of the anterior ossicones from the base of the skull is significantly smaller in Bramiscus. Additionally, the p3 of Bramatherium is much larger and has a lower L/W index than Bramiscus, and Bramatherium has longer and less gracile metapodials than Bramiscus, which all together grants a generic difference.
Bramiscus micros nov. gen. nov. sp., had most likely two pairs of ossicones that differ from other Early and Middle Miocene giraffids and giraffomorphs ossicones in size, orientation, and ornamentation. Giraffokeryx punjabiensis shares with PUPC 13/375 and PUPC 22/01, an ossicone surface ornamented with longitudinal ridges and an oval section of posterior pair (Colbert, 1935). PUPC 13/375 are also similar in size to the anterior ossicones of the adult Giraffokeryx punjabiensis skull AMNH-19475, but PUPC 13/375 and PUPC 22/01 ossicones otherwise differ from those of G. punjabiensis in several features, such as the size of the bases, the inclination, and the curvature, as they are less curved and inclined and are not so laterally projected. In addition, PUPC 13/375 and PUPC 22/01 ossicones show less shallow longitudinal grooves (see Ríos et al., 2017, figures 3-4).
The ossicones of PUPC 13/375 and PUPC 22/01 also differ from the ossicones of the other most basal giraffids recovered in our MPT, Canthumeryx sirtensis, Georgiomeryx georgalasi, Injanatherium arabicum, and Injanatherium hazimi (Heintz et al., 1981; Churcher, 1990; de Bonis et al., 1997, and literature within), which have a much more horizontal insertion to the skull.
There are no known “Progiraffa exigua” ossicones yet. In fact, the description of the species P. exigua was based only on non-very diagnostic lower molars from the Upper Nari beds (now the Chitarwata Formation of Dako Nala in the Bugti Hills of Baluchistan, Pakistan), and therefore we suggest it to be considered a nomem nudum or to be used very cautiously (Pilgrim, 1908). The basicranium ascribed to “Progiraffa exigua” by Barry et al. (2005) (see Barry et al., 2005: figure 13), was later used to establish the new taxon Prolibytherium fusus (Danowitz et al., 2016).
The ossicones of PUPC 13/375 and PUPC 22/01 also differ from other early Giraffomorpha as they are much larger and lack the typical characteristics of the ossicones of those taxa. They lack the spikes of the horns of Climacoceratids, the disposition, and the fused and narrow base of Prolibytherium female appendages, and the morphology and number of palaeomerycid protuberances (Hamilton, 1973, 1978; Barry et al., 2005; Solounias, 2007; Sanchez et al., 2010, 2015). The frontal is small and has thin lateral walls as in the material of early giraffids from Gebel Zelten (Africa) and Wadi Moghara (Egypt) (Pickford et al., 2001).
Regarding the teeth attributed to “Progiraffa exigua” only the two lower molars identified by Pilgrim (1908) can definitively be linked to this taxon. However, these molars lack distinct characteristics and closely resemble the lower molars of other early giraffids. Consequently, the giraffid material from the Chinji formation, designated as “P. exigua” by Aftab et al. (2016), raises doubts about its attribution (refer to Aftab et al., 2016: fig. 3J-L, fig. 3M-O). It’s worth noting that the specimens presented by Aftab et al. (2016) originate from the same formation as the cranial appendages of Bramiscus (rP2 PUPC 81/91, Ghungrila; lP3 PUPC 71/70, Rakh Wasnal). Furthermore, the original “P. exigua” material (consisting of two lower molars) is older, originating from the earlier Kamlial Formation (approximately 18.3-14.2 Ma). As a result, any comparisons of the premolars from Dhok Bun Amir Khatoon to “P. exigua” are based on material that is not contemporaneous with the holotype material of “P. exigua.” Thus, in this study, these premolars are more appropriately tentatively assigned to Bramiscus, given that other giraffids from the same time period in the region, such as Giraffokeryx and Giraffa, exhibit distinct morphologies (see Results section).
Finally, the metatarsals Y-GSP-14999 and Y-GSP 1196 strike a high resemblance to those of Bramatherium megacephalum (e.g., AMNH 19688). The combination of a high and wide medial ridge and, especially, a small lateral ridge (within giraffids only Bramatherium has this feature) defines Bramatherium as unique and our species Bramiscus as similar to it in that aspect (Ríos et al., 2016b). So, this unites the two taxa as well as the ossicone morphology. However, it is important to note that Bramatherium comes from later deposits corresponding with the Late Miocene and Bramiscus micros nov. gen. nov. sp. metatarsals are somewhat older (Middle Miocene-Late Miocene Boundary, 11.827- 9.265 Ma). Our phylogenetic analysis suggests that further analysis of these similarities is warranted, as they belong to different offshoots of the giraffid tree based on our current data.
CONCLUSIONS
We describe a new giraffid genus and species: Bramiscus micros nov. gen. nov. sp. based on the ossicones and dentition recovered from the Middle Miocene of Dhok Bun Amir Khatoon (Pakistan, 14-11.4 Ma) and the Chinji stratotype and which are stored at the Dr. Abu Bakr Fossil Display & Research Centre, University of the Punjab (Lahore, Pakistan) and at the Harvard Museum (USA). These represent the earliest example of ossicones that arise from a fused base, like what is observed in the posterior ossicones of the later and much larger Bramatherium megacephalum However, according to the phylogenetic analysis, these two genera are placed in different offshoots of the giraffid tree and further analysis is required. The dental, cranial, and postcranial information provided by this new material helps us understand the early evolution of this enigmatic family and contribute to elucidating the true nature of giraffoid ossicones and provide evidence of epiphyseal ossicones in this taxon.
Bramiscus micros nov. gen. nov. sp. coexisted in the same area with “Progiraffa exigua”, Giraffokeryx punjabiensis, and Giraffa priscilla. Finally, our material represents the earliest record of giraffids with true ossicones in the Indian subcontinent.
ACKNOWLEDGMENTS
We especially thank J. Barry for his numerous interactions and his help with so many problems over the years. We also thank J. Barry for putting up with us working at his desk where the fossils are. We also thank D. Pilbeam and the Department of Human and Evolutionary Biology and the Peabody Museum of Harvard and the Departments of Mammalogy and Palaeontology of the AMNH for access to specimens.
MR and NS would also like to thank P. Perez, S. Fraile and E. Cantero (MNCN-CSIC, Madrid, Spain), L. Celia and D. DeMiguel (ICP, Barcelona, Spain), P. Montoya (MGUV, Valencia, Spain), J. Galkin, J. Meng, and E. Westwig (AMNH; New York, USA), J. Hooker, P. Brewer, and S. Pappa. Brewer and S. Pappa (NHM, London, UK), S. Sen, C. Argot, and S. Peigne (MNHN, Paris, France) and Ursula B. Göhlich (NHM, Vienna, Austria) and their respective home institutions for access to their giraffe fossil collections.
Funding was provided by NS and the Department of Anatomy at NYIT-COM. We thank our Anatomy Department, librarian Mahnaz Tehrani, and Matthew Mihlbachler. MR also thanks the Stimulus of Scientific Employment, Individual Support - 2018 Call grant from the Fundação para a Ciência e a Tecnologia (Portugal, CEECIND/02199/2018) and GeoBioTec, and the FPI predoctoral grant (2012-2016) (Spanish government MINECO http://www.mineco.gob.es/ codes: BES-2012- 052589, EEBB-I-14-07947, EEBB-I-15-09349, EEBB-I-16-10617) and the EEBB-FPI fellowship programme 2013, 2014 and 2015. MR also thanks the SYNTHESIS+ programme.
We would also like to extend our sincere thanks to the Handling Editor, M. Pole, for his exceptional support and guidance throughout the publication process.
REFERENCES
Aftab, K., Ahmad, Z., Khan, M.A., and Akhtar, M. 2013. New remains of Giraffa priscilla from Parrhewala Chinji Formation, Northern Pakistan. Biologia, 59:233-238.
Aftab, K., Khan, M.A., Ahmad, Z., and Akhtar, M. 2016. Progiraffa (Artiodactyla: Ruminantia: Giraffidae) from the Lower Siwalik Subgroup (Miocene) of Pakistan. Alcheringa, 40(2):207-215.
https://doi.org/10.1080/03115518.2016.1109295
Bärmann, E.V. and Rössner, G.E. 2011. Dental nomenclature in Ruminantia: Towards a standard terminological framework. Mammalian Biology-Zeitschrift für Säugetierkunde, 76(6):762-768.
https://doi.org/10.1016/j.mambio.2011.07.002
Barry, J.C., Morgan, M., Flynn, L., Pilbeam, D., Behrensmeyer, A.K., Raza, S., Khan, I., Badgely, C., Hicks, J., and Kelley, J. 2002. Faunal and Environmental change in the Late Miocene Siwaliks of Northern Pakistan. Paleobiology, 28:1c72.
https://doi.org/10.1666/0094-8373(2002)28[1:faecit]2.0.co;2
Barry, J.C., Cote, S., Maclatchy, L., Lindsay E.H., Kityo, R., and Rajpar, A.R. 2005. Oligocene and Early Miocene Ruminants (Mammalia, Artiodactyla) from Pakistan and Uganda. Palaeontologia Electronica, 8:1-29.
https://palaeo-electronica.org/2005_1/barry22/issue1_05.htm
Barry, J.C., Behrensmeyer, A.K., Badgley, C.E., Flynn, L.J., Peltonen, H., Cheema, I., Pilbeam, D., Lindsay, E.H., Raza, S.M., and Rajpar, A.R. 2013. The Neogene Siwaliks of the Potwar Plateau, Pakistan, p. 373-399. In Wang, X. F., Flynn, L.J., and Fortelius, M. (eds.), Fossil Mammals of Asia: Neogene Biostratigraphy and Chronology. Columbia University Press, New York, 744 pp.
https://doi.org/10.7312/columbia/9780231150125.003.0015
Basu, P.K. 2004. Siwalik mammals of the Jammu Sub-Himalaya, India: an appraisal of their diversity and habitats. Quaternary International, 117(1):105-118.
https://doi.org/10.1016/s1040-6182(03)00120-4
Bettington, A. 1846. Memorandum on certain fossils, more particularly a new Ruminant found at the island of Perim, in the Gulf of Cambay. of the Royal Asiatic Society of Great Britain and Ireland, 8:340-348:417-419.
https://doi.org/10.1017/s0035869x0014290x
Bhatti, Z.H., Khan, M.A., Akhtar, M., Khan, A.M., Ghaffar, A., Iqbal, M., and Ikram, T. 2012. Giraffokeryx (Artiodactyla: Mammalia) remains from the Lower Siwaliks of Pakistan. Pakistan of Zoology, 44:1623-1631.
Bohlin, B. 1926. Die Familie Giraffidae mit besonderer Berücksichtigung der fossilen Formen aus China. Palaeontologia Sinica, Series C4 (I):1-170.
Bohlin, B. 1935. Tsaidamotherium hedini, ng, n. sp. Ein Einhörniger Ovibovine, aus den Tertiären Ablagerungen aus der Gegend des Tossun nor, Tsaidam. Geografiska Annaler, 66-74.
https://doi.org/10.2307/519848
Brunet, M. and Heintz, E. 1983. Interprétation paléoécologique et relations biogéographiques de la faune de vertébrés du Miocene Supérieur D'Injana, Irak. Palaeogeography, Palaeoclimatology, Palaeoecology, 44:283-293.
https://doi.org/10.1016/0031-0182(83)90107-4
Cantalapiedra, J.L., Fernández, M.H., Azanza, B., and Morales, J. 2015. Congruent phylogenetic and fossil signatures of mammalian diversification dynamics driven by Tertiary abiotic change. Evolution, 69:2941-2953.
https://doi.org/10.1111/evo.12787
Cheema, I.U. 2003. Phylogeny and Evolution of Neogene Murine Rodents from the Potwar Plateau of Pakistan and Azad Kashmir with Special Emphasis on Zoogeographic Diversification and Stratigraphic Implications. PhD thesis (unpublished), University of the Punjab, Lahore, Pakistan, pp. 1-176.
Cheema, L., Rajpar, A., and Raza, S.M., 1997. Preliminary analysis of the vertebrate fauna from the Siwaliks of Chakwal District; Kallar-Kahar-Dhok Tahlian area, Potwar Plateau, Pakistan. The Geological Bulletin of The Punjab University, (31-32):161-168.
Churcher, C.S. 1970. Two new upper Miocene giraffids from Fort Ternan, Kenya, East Africa: Palaeotragus primaevus n. sp. and Samotherium africanum n. sp. Fossil vertebrates of Africa, 2:1-106.
https://doi.org/10.2307/2799123
Churcher, C.S. 1978. Chapter 25. Giraffidae, p. 509-535. In Maglio V.J. and Cooke H.B. S., (eds.), Evolution of African Mammals. Harvard University Press.
https://doi.org/10.1126/science.204.4394.755
Churcher C. 1990. Cranial Appendages of Giraffoidea, p. 180-194. In Bubenik G. and Bubenik A. (eds.), Horns, Pronghorns, and Antlers. Springer, New York.
https://doi.org/10.1007/978-1-4613-8966-8_5
Colbert, E.H. 1933. A skull and mandible of Giraffokeryx punjabiensis Pilgrim. American Museum novitates. 632:1-14.
Colbert, E.H. 1935. Siwalik mammals in the American Museum of Natural History. Transactions of the American Philosophical Society Nature Series, 26:1-401.
https://doi.org/10.2307/1005467
Colbert, E.H. 1936. Palaeotragus in the Tung Gur formation of Mongolia. American Museum Novitates, 874:1-18.
Danowitz, M., Domalski, R., and Solounias, N. 2016. A new species of Prolibytherium (Ruminantia, Mammalia) from Pakistan, and the functional implications of an atypical atlanto-occipital morphology. of Mammalian Evolution, 23: 201-207.
de Bonis L., Koufos G.D., and Sen S. 1997. A giraffid from the middle Miocene of the island of Chios, Greece. Palaeontology, 40:121-133.
Dietrich, W.O. 1937. Pleistozäne giraffiden und bovinen aus Oldoway, Deutsch-Ostafrika. Wissenschaftliche Ergebnisse der Oldoway-Expedition 1913, 4:106-110.
Dietrich, W.O. 1942. Ältestquartäre säugetiere aus der südlichen Serengeti, Deutsch-Ostafrika. Palaeontographica, XCIV(A): 43-192.
Falconer, H. 1845. Description of some fossil remains of Dinotherium, Giraffe, and other Mammalia, from the Gulf of Cambay, western coast of India, chiefly from the collection presented by Captain Fulljames, of the Bombay Engineers, to the Museum of the Geological Society. Quarterly of the Geological Society, 1:356-372. https://doi.org/10.1144/gsl.jgs.1845.001.01.80
Fennessy, J., Bidon, T., Reuss, F., Kumar, V., Elkan, P., Nilsson, M.A., Vamberger, M., Fritz, U., and Janke, A. 2016. Multi-locus analyses reveal four giraffe species instead of one. Current Biology, 26(18):2543-2549.
https://doi.org/10.1016/j.cub.2016.07.036
Gaudry, A. 1861. Animaux Fossiles de L’Attique Résultats des fouilles exécutées en Grèce sous les auspices de l’Académie des Sciences. Comptes rendus hebdomadaires des séances de l’Académie des Sciences, 51:802-804.
Gaudry, A., Fischer, P., and Tournouer, R. 1873. Etude sur les vertébrés, p. 5-171. In Savy, F. (ed.), Animaux fossiles du Mont Léberon (Vaucluse). Librairie de la Societe Géologique de France.
Gentry, A.W. 1994. The Miocene differentiation of Old World Pecora (Mammalia). Historical Biology, 7:115-158.
https://doi.org/10.1080/10292389409380449
Gentry, A.W., Rossner, G.E., and Heizmann, E.P.J. 1999. Ruminantia, p. 225-258. In Rossner, G.E. and Heissig, K. (eds.), The Miocene Land Mammals of Europe. Dr. Friedrich Pfeil Verlag, Munchen, Germany.
https://doi.org/10.5860/choice.37-3363
Geraads, D. 1994. Les gisements de mammifères du Miocène supérieur de Kemiklitepe, Turquie: 8. Giraffidae. Bulletin du Muséum national d’Histoire naturelle, 4è sér. C, 16:159-172.
Geraads, D. 2009. Giraffidae (Mammalia) de la fin du Néogène de la République de Macédoine (ARYM). Geodiversitas, 31:893-908.
https://doi.org/10.5252/g2009n4a893
Geraads, D. and Aslan, F. 2003. Giraffidae from the middle Miocene hominoid locality of Çandır (Turkey). Courier Forschungsinstitut Senckenberg, 240:201-209.
Geraads, D., Reed, K., and Bobe, R. 2013. Pliocene Giraffidae (Mammalia) from the Hadar Formation of Hadar and Ledi-Geraru, Lower Awash, Ethiopia. of Vertebrate Paleontology, 33(2):470-481.
https://doi.org/10.1080/02724634.2013.723657
Ginsburg, L., Morales, J., and Soria, D. 2001. Les Ruminantia (Artiodactyla, Mammalia) du miocène des Bugti (Balouchistan, Pakistan). Estudios Geologicos, 57:155-170.
Giribet, G. 2007. Efficient tree searches with available algorithms. Evolutionary Bioinformatics, 3: 117693430700300014.
https://doi.org/10.1177/117693430700300014
Goloboff, P.A., Farris, J.S., and Nixon, K.C. 2008. TNT, a free program for phylogenetic analysis. Cladistics, 24(5):774-786.
https://doi.org/10.1111/j.1096-0031.2008.00217.x
Gray, J.E. 1821. On the natural arrangement of vertebrate animals. London Medical Repository, 15:297-310.
Grossman, A. and Solounias, N. 2014. New fossils of Giraffoidea (Mammalia: Artiodactyla) from the Lothidok Formation (Kalodirr Member, early Miocene, West Turkana, Kenya) contribute to our understanding of early giraffoid diversity. Zitteliana, B32:63-70.
https://epub.ub.uni-muenchen.de/22387/1/zitteliana_2014_b32_06.pdf
Hamilton, W.R. 1973. The lower Miocene ruminants of Gebel Zelten, Libya. Bulletin of the British Museum (Natural History) London, Geology, 21:75-150.
Hamilton, W.R. 1978. Fossil giraffes from the Miocene of Africa and a revision of the Giraffoidea. Philosophical Transactions of the Royal Society of London, B. Biological Sciences, 283:165-229.
https://doi.org/10.1098/rstb.1978.0019
Harris, J. 1976a. Giraffidae from the East Rudolf succession, p. 264-267. In Coppens, Y., Howell, F.C., Isaac, G.L., and Leakey, R.E. (eds.), Earliest Man and Environments in the Lake Rudolf Basin; Chicago University Press, Chicago.
https://doi.org/10.1016/s0016-6995(76)80030-7
Harris, J.M. 1976b. Pliocene Giraffoidea (Mammalia, Artiodactyla) from the Cape Province. Annals of the South African Museum, 69:325-353.
Harris, J.M. 1991. Koobi Fora Research Project, Vol. 3: The Fossil Ungulates: Geology, Fossil Artiodactyls, and Paleoenvironments. Clarendon Press, Oxford, 383 pp.
https://doi.org/10.1016/0016-6995(92)80016-7
Harris, J.M., Solounias, N., and Geraads, D. 2010. Chapter 39. Giraffoidea, p. 805-820. In Werdelin L., Sanders, W.J. (eds.), Cenozoic Mammals of Africa. University of California Press.
https://doi.org/10.1525/california/9780520257214.001.0001
Heintz, E., Brunet, M., and Sen, S. 1981. Un nouveau giraffide du Miocene superieur d’Irak: Injanatherium hazimi n.g., n. sp. Comptes Rendus de l'Académie des Sciences Serie II Sciences de la Vie, 292:423-426.
Hendey, Q. 1970. A review of the geology and palaeontology of the Plio-Pleistocene deposits at Langebaanweg, Cape Province. Annals of the South African Museum, 56:75-117.
Jafarzadeh, R., Kostopoulos, D.S., and Daneshian, J. 2012. Skull reconstruction and ecology of Urmiatherium polaki (Bovidae, Mammalia) from the upper Miocene deposits of Maragheh, Iran. Paläontologische Zeitschrift, 86(1):103-111.
https://doi.org/10.1007/s12542-011-0123-z
Johnson, N.M., Stix, J., Tauxe, L., Cerveny, P.F., and Tahirkheli, A.K. 1985. Paleomagnetic chronology, fluvial processes, and tectonic implications of the Siwalik deposits near Chinji village, Pakistan. of Geology, 93:27-40.
https://doi.org/10.1086/628917
Khan, M.A., Akhtar, M., Ghaffar, A., Iqbal, M., Khan, A.M., and Farooq, U. 2008. Early ruminants from Dhok Bin Mir Khatoon (Chakwal, Punjab, Pakistan): Systematics, Biostratigraphy and Paleoecology. Pakistan of Zoology, 40:457-463.
Khan, M.A., Malik, M., Khan, A.M., Iqbal, M., and Akhtar, M. 2009. Mammalian remains in the Chinji type locality of the Chinji Formation: A new collection. of Animal and Plant Sciences, 19:224-229.
Khan, M.A., Butt, S.S., Khan, A.M., and Akhtar, M. 2010. A new collection of Giraffokeryx punjabiensis (Giraffidae, Ruminantia, Artiodactyla) from the Lehri Outcrops, Jhelum, Northern Pakistan. Pakistan of Science, 62:120-123.
Khan, M.A., Akhtar, M., Roohi, G., Iqbal, M., and Samiullah, K. 2011. Sivaceros gradiens Pilgrim 1937 (Mammalia, Bovidae, Boselaphini) from the Lower Siwaliks of Dhok Bun Amir Khatoon, Chakwal, Pakistan: Systematics and Biostratigraphy. Turkish of Zoology, 34:281-286.
https://doi.org/10.3906/zoo-0812-18
Khan, M.A., Akhtar, M., and Ikram, T. 2012. True ungulates from the Nagri type locality (Late Miocene), northern Pakistan. of Animal and Plant Sciences Supplementary Series, 22:1-59.
Khan, M.A., Akhtar, M., Babar, M.A., Abbas, S.G., Siddiq, M.K., Nawaz, M.K., Mubashar, M., Yaqoob, H., Nazir, S., Farheen, S., Draz, O., Shahbaz, S., Sultana, N., Zahra, N., and Noreen, S. 2013. Some New Remains of Middle Miocene Mammals from the Chinji Formation, northern Pakistan. Pakistan of Zoology, Supplementary Series, 13:1-55.
Khan, M.A., Akhtar, M., and Irum, A. 2014. Bramatherium from the Middle Siwaliks of Hasnot, northern Pakistan. Turkish of Earth Sciences, 23:308-320.
https://doi.org/10.3906/yer-1112-11
Khan, M.A., Abbas, S.G., Babar, M.A., Kiran, S., Riaz, A., and Akhtar, M. 2017. Dorcatherium (Mammalia: Tragulidae) from Lower Siwaliks of Dhok Bun Amir Khatoon, Punjab, Pakistan. Pakistan of Zoology, 49(3):883-888. https://doi.org/10.17582/.pjz/2017.49.3.883.888
Köhler, M. 1993. Skeleton and Habitat of fossil and recent Ruminants. Munchner geowiss. Abh.(A), 25, 1-88.
Kostopoulos, D.S. 2009. The Late Miocene Mammal Faunas of the Mytilinii Basin, Samos Island, Greece: New Collection. 13. Giraffidae. Beiträge zur Paläontologie, 31:345-389.
Kostopoulos, D.S., Koliadimou, K.K., and Koufos, G.D. 1996. The giraffids (Mammalia, Artiodactyla) from the late Miocene mammalian localities of Nikiti (Macedonia, Greece). Palaeontographica Abteilung A, 239:61-88.
https://doi.org/10.1127/pala/239/1996/61
Lewis, G.E., 1939. A new Bramatherium skull. American of Science, 237(4):275-280.
Lindsay, E.H., Flynn, L.J., Cheema, I.U., Barry, J.C., Downing, K., Rajpar, A.R., and Raza, S.M. 2005. Will Downs and the Zinda Pir Dome, Palaeontologia Electronica, 8(1):19A:18p.
https://palaeo-electronica.org/2005_1/lindsay19/issue1_05.htm
Lydekker, R. 1876. Molar teeth and other remains of Mammalia. Palaeontologia Indica, 10:19-87.
Lydekker, R. 1883. Synopsis of the Fossil Vertebrata of India. Records of the Geological Survey of India, 16:61-93.
Maddison, W.P. and Maddison, D.R. 2016. Mesquite: a modular system for evolutionary analysis. Version 3.04.
http://mesquiteproject.org
Mahmood, K., Khan, M.A., Siddiq, M.K., Babar, M.A., and Akhtar, M. 2015. New Fossils of Giraffokeryx (Mammalia: Cetartiodactyla: Giraffidae) from Chinji Formation, Pakistan. Pakistan of Zoology, 47(2):367-375.
Matthew, W.D. 1929. Critical observations upon Siwalik mammals (exclusive of Proboscidea). Bulletin of American Museum of Natural History, 56:437-560.
Mecquenem, de R. 1924. Contribution a l’étude des fossiles de Maragha. Annales de Paléontologie, 13:135-160; 14:1-36.
Μelentis, Ι.Κ. 1974. Helladotherium duvernoyi Gaudry, 1860 aus Pikermi (Griechenland). Scientific Epetiris: issued by the Faculty of Natural and Mathematical Sciences (AUTH),14:65-73.
Mitchell, G. and Skinner, J.D. 2003. On the origin, evolution and phylogeny of giraffes Giraffa camelopardalis. Transactions of the Royal Society of South Africa, 58(1):1-23.
https://doi.org/10.1080/00359190309519935
Nanda, A.C. 2002. Upper Siwalik mammalian faunas of India and associated events. of Asian Earth Sciences, 21:47-58.
https://doi.org/10.1016/s1367-9120(02)00013-5
Paraskevaidis, I. 1940. Eine obermiocine Fauna von Chios. Neues Jahrbuch fiir Mineralogie, 83:363-442.
Pickford, M., Attia, Y.S., and El Ghany, M.S. 2001. Discovery of Prolibytherium magnieri Arambourg, 1961 (Artiodactyla, Climacoceratidae) in Egypt. Geodiversitas, 23(4):647-652.
Pilgrim, G.E. 1908. The Tertiary and Post-Tertiary freshwater deposits of Baluchistan and Sind with notices of new vertebrates. Records of the Geological Survey of India, 2:139-166.
Pilgrim, G.E. 1910. Notices of new Mammalian genera and species from the tertiaries of India-Calcutta. Records of the Geological Survey of India, 40:63-71.
Pilgrim, G.E. 1911. The fossil Giraffidae of India. Geological Survey of India Memoirs, Palaeontologia Indica, New Series, 4(1):1-29.
Raza, S.M. and Meyer, G.E. 1984. Early Miocene geology and palaeontology of the Bugti Hills, Pakistan. Geological Survey of Pakistan Memoirs, 11:43-63.
Ríos, M., Sánchez, I.M., and Morales. J. 2016a. Comparative anatomy, phylogeny, and systematics of the Miocene giraffid Decennatherium pachecoi Crusafont, 1952 (Mammalia, Ruminantia, Pecora): state of the art. of Vertebrate Paleontology: e1187624
https://doi.org/10.1080/02724634.2016.1187624
Ríos, M., Danowitz, M., and Solounias, S. 2016b. First comprehensive morphological analysis on the metapodials of Giraffidae. Palaeontologia Electronica, 19(3):1-39.
https://doi.org/10.26879/653
Ríos M, Sánchez I.M., and Morales J. 2017. A new giraffid (Mammalia, Ruminantia, Pecora) from the late Miocene of Spain, and the evolution of the sivathere-samothere lineage. Plos One, 12(11):e0185378.
https://doi.org/10.1371/.pone.0185378
Ríos, M., Danowitz, M., and Solounias, N., 2019. First identification of Decennatherium Crusafont, 1952 (Mammalia, Ruminantia, Pecora) in the Siwaliks of Pakistan. Geobios, 57:97-110.
https://doi.org/10.1016/j.geobios.2019.10.007
Ríos, M. and Morales, J. 2019. A new skull of Decennatherium rex Ríos, Sánchez and Morales, 2017 from Batallones-4 (upper Vallesian, MN10, Madrid, Spain). Palaeontologia Electronica, 22.2.vpc_1 1-6.
https://doi.org/10.26879/965palaeo-electronica.org/content/vpc_1
Ríos, M., Abbas, S.G., Khan, M.A., and Solounias, N. 2022a. Distinction of Sivatherium from Libytherium and a new species of Libytherium (Giraffidae, Ruminantia, Mammalia) from the Siwaliks of Pakistan (Miocene). Geobios, 74: 67-76.
https://doi.org/10.1016/j.geobios.2022.06.002
Ríos, M., Cantero, E., Estraviz-Lopez, D., Solounias, N., and Morales, J. 2022b. Anterior ossicone variability in Decennatherium rex Ríos, et al., 2017 (Late Miocene, Iberian Peninsula). Earth and Environmental Science Transactions of the Royal Society of Edinburgh, 114:1-7.
https://doi.org/10.1017/S1755691022000184
Robinson, C.A. 2011. Chapter 14. Giraffidae, p. 339-362. In Harrison, T. (ed.) Paleontology and Geology of Laetoli: Human Evolution in Context: Volume 2: Fossil Hominins and the Associated Fauna. Springer Science and Business Media, Dordrecht.
https://doi.org/10.1007/978-90-481-9962-4_14
Rodler, A. 1889. Über Urmiatherium polaki ng, n. sp., einen neuen Sivatheriiden aus dem Knochenfelde von Maragha. Denkschriften der Kaiserlichen Akademie der Wissenschaften, Mathematisch-Naturwissenschaftliche Klasse, 56:315-322.
Rodler, A., and Weithofer, C.A. 1890. Die Wiederkäuer der Fauna von Maragha. Denkschrift Akademische Wissenschaft, Wien, 57:753-572.
Rössner, G.E. 2010. Systematics and palaeoecology of Ruminantia (Artiodactyla, Mammalia) from the Miocene of Sandelzhausen (southern Germany, northern Alpine Foreland basin). Paläontologische Zeitschrift, 84(1):123--162.
https://doi.org/10.1007/s12542-010-0052-2
Sánchez, I.M., Domingo, M.S., and Morales, J. 2010. The genus Hispanomeryx (Mammalia, Ruminantia, Moschidae) and its bearing on musk deer phylogeny and systematics. Palaeontology, 53:1023-1047.
https://doi.org/10.1111/j.1475-4983.2010.00992.x
Sánchez, I., Cantalapiedra, J., Rios, M., Quiralte, V., and Morales, J. 2015. Systematics and Evolution of the Miocene Three-Horned Palaeomerycid Ruminants (Mammalia, Cetartiodactyla). Plos One, 10(12):e0143034.
https://doi.org/10.1371/.pone.0143034
Scott, R.S., Kappelman, J., and Kelley, J. 1999. The paleoenvironment of Sivapithecus parvada. of Human Evolution, 36(3):245-274.
https://doi.org/10.1006/jhev.1998.0269
Shi, Q.Q., Wang, S.Q., Chen, S.K., and Li, Y.K. 2016. The first discovery of Urmiatherium (Bovidae, Artiodactyla) from Liushu Formation, Linxia Basin. Vertebrata PalAsiatica, 54(4):319-331.
https://doi.org/2012CB821906
Singer, R. and Boné, E. 1960. Modern giraffes and the fossil giraffids of Africa. Annals of the South Africa Museum, 45:375-548.
Solounias, N. 2007. Family Giraffidae, p. 257-277. In Prothero, D.R. and Foss, S.E. (eds.), The Evolution of Artiodactyls. The Johns Hopkins University Press, Baltimore.
Thomas, H. 1983. Les Bovidae (Artiodactyla, Mammalia) du Miocene moyen de la Formation Hofuf (Province du Hasa, Arabie Saoudite). Paleovertebrata, 13(5):157-206.
Ullah, K., Arif, A.M., and Shah, M.T. 2006. Petrography of sandstones from the Kamlial and Chinji formations, southwestern Kohat plateau, NW Pakistan: implications for source lithology and paleoclimate. of Himalayan Earth Sciences, 39:1-13.
https://doi.org/10.3906/yer-1410-21
Vasishat, R.N., Kaul, S., and Chopra. S.R. 1979. Additional fossil suid material from the Lower Siwalik of Ramnagar, J and K State, India. In Proceedings of Colloquium on Paleontological studies in southern region. Geological Survey of India, 45:219-225.
Welcomme, J.L., Antoine, P.O., Duranthon, F., Mein, P., and Ginsburg, L., 1997. New discoveries of Miocene vertebrates in the Dera Bugti syncline (Balochistan, Pakistan). Comptes Rendus de l’Academie des Sciences, Serie IIA- Sciences de la terre des planètes, 325:531-536.
Welcomme, J.L., Benammi, M., Crochet, J.Y., Marivaux, L., Metais, G., Antoine, P.O., and Baloch, I.S. 2001. Himalayan Forelands: palaeontological evidence for Oligocene detrital deposits in the Bugti Hills (Balochistan, Pakistan). Geological Magazine, 138:397-405.
https://doi.org/10.1017/s0016756801005428
