

 Proboscidea from Miocene strata of the Texas Coastal Plain with a reappraisal of the biochronology of the Fleming Group
Proboscidea from Miocene strata of the Texas Coastal Plain with a reappraisal of the biochronology of the Fleming Group
Article number: 27.3.a55
https://doi.org/10.26879/1404
Copyright Society of Vertebrate Paleontology, November 2024
Author biography
Plain-language and multi-lingual abstracts
PDF version
Submission: 13 May 2024. Acceptance: 29 October 2024.
ABSTRACT
The earliest confident identification of fossil proboscideans from Texas occurs in the Early Barstovian Lagarto Formation of the Texas Coastal Plain. Three local faunas contain at least two different species of the genus Gomphotherium. One of these, identified as Gomphotherium cf. Gomphotherium inopinatum, is similar to a species of Gomphotherium known from Hemingfordian equivalent faunas in China. The other species is identified as Gomphotherium sp. with similarities to both Gomphotherium subtapiroideum and Gomphotherium productum. The Belts Creek, Noble Farm, and Brenham Local Faunas have been incorrectly interpreted as Late Barstovian faunas by previous authors in part because of the hypothesis of a “Proboscidean Datum.” A single fragment of a proboscidean vertebra is known from a locality that may be Late Hemingfordian in age, although both the age and identification of that specimen are uncertain. Additional gomphotheres are known from Late Barstovian and Early Clarendonian faunas in the Texas Coastal Plain, but mammutids are absent. Late Barstovian gomphotheres from the upper part of the Lagarto Formation are small and share characteristics with Gomphotherium angustidens and Gomphotherium productum. The Early Clarendonian Lapara Creek Fauna includes cf. Gomphotherium sp. as well as Blancotherium buckneri. The arrival of gomphotheres along the southern coast of North America correlates with low stands of sea level during the Miocene Climate Optimum. Virtual-wear occlusal patterns derived from CT scans of unworn gomphothere molars proved useful for description and identification.
Steven R. May, Jackson School Museum of Earth History, The University of Texas at Austin, Austin, Texas 78758, USA. srmay@utexas.edu
Keywords: Proboscidea; gomphothere; Miocene; Texas
Final citation: May, Steven R. 2024. Proboscidea from Miocene strata of the Texas Coastal Plain with a reappraisal of the biochronology of the Fleming Group. Palaeontologia Electronica, 27(3):a55.
https://doi.org/10.26879/1404
palaeo-electronica.org/content/2024/5377-texas-proboscidea
Copyright: November 2024 Society of Vertebrate Paleontology.
This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
creativecommons.org/licenses/by/4.0
INTRODUCTION
Proboscideans are an immigrant group to North America that first appeared there in the fossil record during the Miocene. The group had its origins in northern Africa during the Paleocene (Gheerbrant, 2009, Sanders, 2024) and evolved into a diverse order during the Eocene-Miocene in Africa and Eurasia (Gheerbrant and Tassy, 2009). During the Miocene, a major geographic dispersal of proboscideans included the arrival of the mammutid Zygolophodon and the gomphothere Gomphotherium in North America. The notion of a biochronologic “datum” associated with the entrance of proboscideans into North America has been considered by several authors (Woodburne and Swisher, 1995; Tedford et al., 1987, Tedford et al., 2004; Prothero et al., 2008a; Prothero and Dold, 2008, Pagnac, 2009). The appearance of proboscideans in North America faunas has been used to define and/or characterize the Late Barstovian North American Land Mammal Age (Tedford et al., 1987, Tedford et al., 2004). However, in their own discussions, Tedford et al. (2004) noted complications with this model including Late Hemingfordian occurrences of Zygolophodon and Early Barstovian occurrences of Zygolophodon and Gomphotherium. The actual fossil record is complex and the concept of a ‘Proboscidean Datum’ has led to incorrect age assignments for some Miocene vertebrate faunas in Texas. In his review of mammalian biostratigraphy and biochronology from the Barstow Formation, Pagnac (2009) argued for abandoning the use of proboscideans as defining taxa for Ba1 and Ba2 divisions of the Barstovian.
The earliest record of proboscideans in North America is commonly cited as a partial cheek tooth from the Late Hemingfordian Massacre Lake Local Fauna in northwestern Nevada (Morea, 1981; Prothero et al., 2008a). This specimen was assigned to Zygolophodon based on the zygodont molar morphology. Implying some uncertainty associated with the Massacre Lake specimen, Tedford et al. (2004) stated that the Early Barstovian (Ba1) could be characterized by the “definite” occurrence of Zygolophodon. They referred to a number of Ba1 occurrences of proboscideans including the California Coast Ranges where they listed Zygolophodon and possibly Gomphotherium from the North Coalinga and Sharktooth Hill Local Faunas. Early Barstovian occurrences of Zygolophodon from Oregon, Nevada, Montana, Texas Coastal Plain (Burkeville), Florida, and Mexico have been documented leading Tedford et al. (2004) to conclude “it may be useful to include Zygolophodon as part of the characterizing fauna of the early Barstovian.” The fossil record of mammutids in North America includes numerous Early Barstovian occurrences most of which have been recently assigned to Zygolophodon proavus (Von Koenigswald et al., 2023). Early Barstovian faunas from the interior of North America lack proboscideans, although gomphotheriids are known in this region from Late Barstovian faunas. This observation led Tedford et al. (2004) to conclude “The beginning of the late Barstovian (Ba2) is defined by the first appearance of gomphotheriid Proboscidea.” Although this statement was only supported by data from the interior of North America, it has been applied more broadly leading to several erroneous age interpretations.
There are numerous published reviews of the systematics and phylogeny of gomphotheres. Most of those concluded that the family Gomphotheriidae (attributed to Hay, 1922) is a paraphyletic group (Tassy, 1996b; Shoshani, 1996; Lambert, 1996; Wang et al., 2017). In their review of Gomphotherium from North America, Lambert and Shoshani (1998) recognized Gomphotherium angustidens, Gomphotherium productum, and a group referred to as Gomphotherium sp. that included 14 named species of uncertain validity. Based on the Eurasian record, Wang et al. (2017) presented a phylogenetic analysis of the genus Gomphotherium and recognized three groups: “ Gomphotherium annectens”, “Gomphotherium angustidens”, and “derived Gomphotherium.” The gomphotheres are a challenging group in part because of their conservative evolution but widespread distribution in space and time. They are a group in need of revision, an endeavor that is beyond the scope of this paper, but a fact that tempers the systematic paleontology discussed below.
Materials and Methods
Abbreviations. AMNH, American Museum of Natural History; BEG, Bureau of Economic Geology, The University of Texas at Austin; F:AM, Frick Collection, American Museum of Natural History; TAM, Texas A&M Museum; TxVP, Texas Vertebrate Paleontology Collections, The University of Texas at Austin; UCMP, Museum of Paleontology University of California, Berkeley; NALMA, North American Land Mammal “Age”; L.F., Local Fauna; AP, anteroposterior; I incisor; M, upper molar; m, lower molar; MCH, crown height measured along the mesostyle; P, upper premolar; p, lower premolar; DP, deciduous upper premolar; Dp, deciduous lower premolar; L, left; R, right; mm, millimeters. Specimens and archival material at The University of Texas at Austin are curated in the Texas Vertebrate Paleontology Collections. Locality numbers are formatted TMM ##### and specimen numbers include the locality number, followed by a dash and the catalog number, i.e., TMM #####-####. Records of Mark Francis, Curtis Hesse, John Wilson, and the Statewide Paleontologic-Mineralogic Survey are housed in the Texas Vertebrate Paleontology Archives on the J.J. Pickle Research Campus.
Anatomical Terminology. Dental terminology used here is based in part on Tassy (1996a) and Sanders and Miller (2002). Cranial terminology is based on Tassy (2013). Lophs or lophids on the cheek teeth of gomphotheres are generally separated into two halves by a median sulcus. Pretrite refers to the more worn half of a loph or lophid while postrite refers to the less worn half. Pretrite is labial in lower cheek teeth and lingual in uppers. Trefoil refers to the ‘three-leaf clover’ pattern associated with wear of the main cusp of each loph(id) and associated anterior and posterior cones.
Faunal Terminology. The terms local fauna and fauna are used following the definitions discussed by Woodburne (2004). Locality, local fauna, and fauna names have been complicated historically by varied use in publications, archival records, and specimen cards. Local fauna names used here are generally like those used by Tedford et al. (2004) for the Texas Coastal Plain. In his summary of fossil vertebrates from the Texas Coastal Plain, Hesse (1942) used the term “fauna” to refer to a collection of specimens from one, or a few, geographically and stratigraphically restricted localities. The terminology of Wilson (1956), in which he attempted to use faunal names as geographically extensive biostratigraphic units or biozones, is recognized, but not applied in this paper. For example, the term Cold Spring Local Fauna used here (Cold Spring fauna of Hesse, 1942) includes only the specimens from three localities just north of Coldspring, TX, whereas Quinn (1955) and Wilson (1956) used the term ‘Cold Spring Fauna’ to include numerous local faunas from across 200 miles of outcrop of the Fleming Group. Wilson (1956) argued explicitly that the “local fauna” name should be considered as the “type” fauna for the more widespread series of correlative local faunas, all of which comprised the fauna. Application of interpreted biozones, that utilize the same name as a type of local fauna, but that are referred to as faunas, has led to confusion in the literature associated with the Miocene biostratigraphy and biochronology of the Texas Coastal Plain.
Following Tedford et al. (2004), North American Land Mammal ‘Ages’ (NALMA) include the Early and Late Hemingfordian, referred to as He1 and He2 respectively, and the Early and Late Barstovian, referred to as Ba1 and Ba2, respectively. The Clarendonian NALMA includes Cl1, Cl2, and Cl3 divisions. Numerical calibration of these ages and their divisions follows the time scale of Raffi et al. (2020). At present, there are no independent radiometric dates, nor magnetostratigraphic correlations, associated with fossil vertebrate localities from the Fleming Group strata of the Texas Coastal Plain. Ages are based on stratigraphy, biochronological interpretations, and correlations with equivalent marine facies.
Preparation. Fossils were originally prepared prior to this study at both the University of Texas and Texas A&M using undocumented methods and materials. However, at least some of the techniques likely followed the Handbook of Recommended Procedures (McAnulty, 1939), as outlined by Brown (2013). Prior to this study, preparators at the University of Texas partially dismantled the 1939 TAM museum mount of a gomphothere partial skull and dentary (TMM 10201-3046).
Photography. Specimens were photographed using an iPhone 11 and processed using Adobe Photoshop (v. 20).
Computed Tomography (CT). All CT imaging was performed at The University of Texas at Austin High-Resolution X-ray CT Facility using a North Star Imaging micro-computed tomography scanner with a Comet Microfocal source and a Perkin Elmer detector. TMM 40775-46 was scanned at kV = 220; voxel size = 82.5 μm. TMM 40775-47 was scanned at kV = 220; voxel size = 82.4 μm. Scans are archived at www.morphosource.org. The CT data were visualized and analyzed using 3D Slicer.
STRATIGRAPHY OF THE FLEMING GROUP IN TEXAS
 The Fleming Group has a long and convoluted history in the literature of the Miocene stratigraphy of the Texas Coastal Plain. Wilson (1956) reviewed the Stratigraphic terminology, and vertebrate biostratigraphy was reviewed by Wilson (1956) in which he used the names Oakville Formation, containing the “Garvin Gully Fauna”, and Fleming Formation, containing the younger “Burkeville” and “Cold Spring Faunas”. However, Plummer (1932) and Renick (1936) had previously elevated the Fleming to group status composed of the underlying Oakville Formation and the overlying Lagarto Formation. Recent authors (Galloway et al., 1986; Young et al., 2010) consider the Oakville and Lagarto Formations to be part of the Fleming Group. That is the stratigraphic nomenclature used by May and Brown (2023) and in this paper (Figure 1).
The Fleming Group has a long and convoluted history in the literature of the Miocene stratigraphy of the Texas Coastal Plain. Wilson (1956) reviewed the Stratigraphic terminology, and vertebrate biostratigraphy was reviewed by Wilson (1956) in which he used the names Oakville Formation, containing the “Garvin Gully Fauna”, and Fleming Formation, containing the younger “Burkeville” and “Cold Spring Faunas”. However, Plummer (1932) and Renick (1936) had previously elevated the Fleming to group status composed of the underlying Oakville Formation and the overlying Lagarto Formation. Recent authors (Galloway et al., 1986; Young et al., 2010) consider the Oakville and Lagarto Formations to be part of the Fleming Group. That is the stratigraphic nomenclature used by May and Brown (2023) and in this paper (Figure 1).
The Fleming Group includes several fluvial systems in up-dip areas that transition to deltaic and marginal marine systems down-dip and to fully marine offshore (Galloway et al., 1986; Morton et al., 1988). Measured thicknesses from outcrop include 90-215 m in the Oakville Formation and 215-430 m in the Lagarto Formation (Galloway et al., 1986). Incorporation of subsurface data indicates that the Oakville Formation is a sandy progradational sequence above a maximum transgression associated with the Anahuac Shale. The Lagarto Formation is muddy with an aggradational to retrogradational architecture (Galloway et al., 1986).
Detailed stratigraphic correlation between localities in the fluvial-dominated outcrops of the Oakville and Lagarto Formations has not been possible. The relative stratigraphic positions of local faunas can be reasonably well established based on location with respect to lower and upper lithostratigraphic boundaries. Geologic map patterns and regional bedding attitudes provide a consistent stratigraphic framework for biostratigraphic analysis. Except for the Burkeville locality, all local faunas are contained within the “Moulton/Point Blank Depositional System” of Galloway et al. (1986). They interpreted this system as having been deposited in small, sinuous streams that resulted in thin channel-fill sandstones and associated calcareous floodplain mudstones.
Based on correlation of outcrop and subsurface, both planktonic and benthonic foraminifera constrain the age of the Oakville Formation to be Aquitanian through middle Burdigalian, with the Lagarto Formation being middle Burdigalian through Langhian (Galloway et al., 1986). According to the time scale of Raffi et al. (2020), this correlation suggests that the Oakville Formation would be largely Arikareean (Ar3-Ar4) through Early Hemingfordian (He1) in age and the Lagarto Formation would be Late Hemingfordian and Barstovian (Figure 1). Although generally consistent with ages interpreted from the fossil vertebrates, Late Barstovian (Ba2) faunas from the Lagarto Formation suggest that the upper part of the Fleming Group extends into the early Serrivalian. The Garvin Gully L.F., from the Oakville Formation, was interpreted as a late He1 fauna by Prothero and Manning (1987) based on the presence of the rhinoceros Menoceras barbouri, while the age of this local fauna was depicted by Tedford et al. (2004) straddling the He1-He2 boundary.
The apparent paucity of Late Hemingfordian faunas in the Texas Coastal Plain is interesting given the number of Ba1 local faunas in the lower to middle part of the Lagarto Formation. There is relatively little stratigraphic separation between the He1 Garvin Gully L.F. and the Ba1 Noble Farm L.F. and, farther east, the Ba1 Moscow L.F. is very near the mapped contact between the Lagarto and underlying Catahoula Formations. The first occurrences of Monosaulax, Aelurodon, Calippus, Hipparion, and the last occurrence of Amphicyon were used to characterize Ba1 faunas by Tedford et al. (2004). All of these taxa are known from Ba1 faunas from the Texas Coastal Plain. Until the record of small mammals, and especially rodents, can be improved from strata of the Fleming Group, or until tuffaceous units can be identified and dated, uncertainty will remain about the He2-Ba1 boundary in this region.
The Geologic Map of Texas (Barnes et al., 1992), and a depth-structure map on the top of the Frio-Anahuac Formations that defines the base of the Fleming Group (Swanson and Karlsen, 2009), have been used to develop a stratigraphic framework for local faunas of the Texas Coastal Plain. The Oakville Formation is recognized on the state map west of the Brazos River valley. However, Wilson (1957) stated that it could easily be recognized east of Navasota in the Garvin Gully area. The base of the Fleming Group has been difficult to precisely define in outcrop (Albright, 1998), but recognizing this uncertainty as well as thickness variability, a geologic map of the Miocene strata emphasizing the projected base and top of key stratigraphic units has been developed. Because the topography is relatively flat-lying and the Fleming Group strata dip gently to the south-southeast, the stratigraphic position of specific localities can be interpreted from map patterns. All localities with Barstovian faunas from the Lagarto Formation discussed in this paper, except for Burkeville, are from the “Moulton/Point Blank streamplain” depositional system of Galloway et al. (1986). Deeply incised channels with thick channel-fill deposits that could complicate the stratigraphic model described in this paper have not been described from the “Moulton/Point Blank streamplain” depositional system.
Faunal composition and stratigraphic location have been used to reevaluate the chronostratigraphic distribution of Miocene local faunas from the Texas Coastal Plain (Figure 1). Assemblages of fossil vertebrates from the Oakville Formation, or the lower part of the Fleming Group have been interpreted as Arikareean and Hemingfordian faunas (Albright, 1998, Tedford et al 2004, etc.). Assemblages from the upper part of the Fleming Group (upper Lagarto Formation) have been interpreted as Late Barstovian (Ba2) faunas (Tedford et al., 2004). Assemblages from the lower and middle Lagarto Formation have been interpreted as Early Barstovian (Ba1) faunas (Tedford et al., 2004; May and Brown, 2023). The model shown in Figure 1 is generally consistent with Tedford et al. (2004) but differs in certain details. It also differs from previous age interpretations by several other authors that were often based on either the notion of a ‘Proboscidean datum’, or on incorrect application of the biozone model promoted by Quinn (1955) and Wilson (1956), as described previously. Specifically, the Belts Creek L.F. and the Noble Farm L.F. are here shown to be Ba1 faunas, rather than Ba2 as suggested by Prothero and Manning (1987), Hulbert (1988), and Wang et al. (1999). Fossils from TMM 40418 (Brenham L.F.) are interpreted to represent a Ba1 fauna. Usage of the name “Sam Houston L.F.” is dropped as TMM 31191 is merely a specific locality within the restricted area that has produced the Cold Spring L.F. The Sam Houston locality is part of the Ba2 Cold Spring L.F. rather than a younger local fauna as depicted in Tedford et al. (2004).
VERTEBRATE BIOCHRONOLOGY
Hemingfordian Faunas
The Garvin Gully L.F. is an Early Hemingfordian fauna that includes carnivores, equids, rhinoceroses, and artiodactyls, but no proboscideans (Patton, 1969; Forsten, 1975; Prothero and Manning, 1987, Tedford et al., 2004). Most of the specimens from Garvin Gully were collected by Mark Francis and his associates at Texas A&M from the late 1800s through the early 1920s and are now curated at the University of Texas at Austin Vertebrate Paleontology Collections. The Garvin Gully L.F. includes both the Garvin Farm (TMM 31084) and the Hidalgo Bluff (TMM 40067) localities. A re-examination of all Garvin Gully material at TxVP confirmed that there is no evidence of proboscideans. Similarly, searches of the AMNH and UCMP databases for specimens from Garvin Gully yielded no proboscidean material.
The composition of the Aiken Hill L.F. in Walker County was interpreted by Albright (1998) as an He1 fauna at least partially correlative with the Garvin Gully L.F. Prosynthetoceras texanus was identified by Patton and Taylor (1971) from Aiken Hill and considered “medial Hemingfordian.” That material was originally collected by Claude Riley for TAM and was included in a short description of TAM 46 by Hesse (1942). He noted that locality as being “very low in the Fleming section.” No proboscidean material is known from Aiken Hill.
Early Barstovian Faunas
Early Barstovian (Ba1) local faunas from the lower and middle Lagarto Formation are distinct from both the Hemingfordian Garvin Gully L.F. of the Oakville Formation and from the Ba2 local faunas of the upper Lagarto Formation. The Ba1 local faunas include Noble Farm, Brenham, Belts Creek, Push Creek, Trinity River, Point Blank, Moscow, and Burkeville (Figure 1). The Noble Farm, Brenham, and Belts Creek Local Faunas contain gomphotheriids. These are the earliest confident identifications of proboscideans in the Texas Coastal Plain.
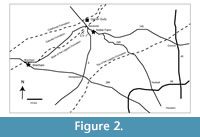 Noble Farm Local Fauna. The Noble Farm L.F. (Hay, 1924) was collected from approximately 7 miles southeast of the Hemingfordian Garvin Gully L.F. in the lower part of the Lagarto Formation (Hesse, 1942) (Figure 2).
Noble Farm Local Fauna. The Noble Farm L.F. (Hay, 1924) was collected from approximately 7 miles southeast of the Hemingfordian Garvin Gully L.F. in the lower part of the Lagarto Formation (Hesse, 1942) (Figure 2).
The stratigraphic position of the Noble Farm L.F. can clearly be assessed from geologic map patterns as being in the lower part of the Lagarto Formation (Figure 2). The composition of the Noble Farm L.F. is consistent with Ba1 faunas, elements of which are reviewed here. Two new species of the carnivore Aelurodon (Aelurodon francisi and Aelurodon simulans), and a new species of Merychippus (Merychippus francisi), along with the proboscidean material he assigned to Gomphotherium cimarronis were reported by Hay (1924). Aelurodon simulans was considered to be synonymous with Aelurodon francisi by Van der Hoof and Gregory (1940) who concluded that Aelurodon francisi from Grimes Prairie “is probably from the same horizon as the Noble Farm fauna from which A. simulans was obtained”. The Aelurodon francisi holotype was dug up from a well at the farm of J. Niscavit in Grimes Prairie, 12 miles east of Navasota (TMM 40140). Although no detailed stratigraphic correlation could have been made, specimens from the Niscavit Farm locality were likely from the lower to middle Lagarto Formation as also concluded by Wilson (1960). This implies that the holotype of Aelurodon francisi is from a Ba1 fauna. Aelurodon francisi and Aelurodon simulans, from the Noble Farm L.F., were interpreted by Wang et al. (1999) to be junior synonyms of Tomarctus brevirostris. The range of Tomarctus brevirostris was stated by Wang et al. (1999) as Ba1 for Colorado, Nebraska, New Mexico, and California; early Ba2 in Colorado (Pawnee Creek Formation); and Ba2 in Texas. They considered the Noble Farm occurrence to be Ba2 in age, however, acknowledging the Ba1 age for the Noble Farm L.F. makes Tomarctus brevirostris a dominant Ba1 taxon with possible extension into the early Ba2 of Colorado.
Merychippus francisi was named by Hay (1924) based on a few isolated teeth from Noble Farm that were later interpreted by Quinn (1955) as Calippus francisi. He also assigned additional material from Barstovian localities to that taxon. The Noble Farm fauna was interpreted by Quinn (1955) to be best correlated with the ‘Cold Spring Fauna’ (Ba2) rather than the Burkeville beds (Ba1), because it included a camelid and Gomphotherium. Recognizing that the stratigraphic position of the Noble Farm locality would be more consistent with a Ba1 age, Quinn (1955) attributed the apparent dilemma to “some structural deformation of the strata.” This represents an early application of the ‘Proboscidean Datum’ hypothesis. The camelids Procamelus and Rakomylus are now considered as characterizing taxa for Ba1 faunas (Tedford et al., 2004) and Gomphotherium is now clearly documented from Ba1 faunas in North America. The need to invoke unobserved structural deformation to explain model-driven biochronology is not necessary. Furthermore, the specimen identified by Hay (1924) as Procamelus leptognathus from the Noble Farm L.F. (TMM 40775-2394) is a partial right dentary with two moderately worn cheek teeth. Based on notes associated with this specimen, Thomas Patton identified it as Prosynthetoceras, a taxon known from Ba1 and Ba2 faunas in the Texas Coastal Plain (Patton, 1969). The dentary of TMM 40775-2394 is shallower dorso-ventrally (16.7 mm below m1) than those of Prosynthetoceras francisi from the Cold Spring L.F. (TMM 31219-213 22.2 mm, TMM 31219-214 27.7 mm). The molar lengths are similar to Prosynthetoceras francisi and larger than Prosynthetoceras texanus from Garvin Gully. The first molar of TMM 40775-2394 is narrower than in Prosynthetoceras francisi from the Cold Spring L.F. and the parastylid and entostylid are not as well developed. Prosynthetoceras texanus from Burkeville was intermediate in size and morphology between Prosynthetoceras texanus from the Garvin Gully L.F. and Prosynthetoceras francisi from the Cold Spring L.F. (Patton, 1969). The limited amount of material, and especially the lack of cranial material, precludes confident identification of the specimen from Noble Farm.
The equid fauna from Noble Farm includes Calippus proplacidus, Calippus circulus and Protohippus perditus primarily based on isolated cheek teeth (Hulbert, 1988). Calippus circulus is only known from a few isolated teeth from Texas Coastal Plain localities limiting its biochronologic utility. Merychippus francisi (Hay, 1924) and Calippus francisi (Quinn, 1955) were included within Calippus proplacidus by Hulbert (1988), a taxon that he identified from both the Noble Farm and Cold Spring Local Faunas. He considered Noble Farm to be Late Barstovian because it contained Gomphotherium and cited “Tedford et al. in press” for his statement that “its age is no older than late Barstovian.” The Tedford et al. “in press” paper cited by Hulbert (1988) was published as Tedford et al. (1987). In that paper, they listed Gomphotherium as a defining taxon for the Late Barstovian. Later, Tedford et al. (2004) recognized several Ba1 occurrences of Gomphotherium. It is now clear that the earlier hypothesis of a Late Barstovian ‘Proboscidean Datum’ resulted in previous incorrect age assignments for the Noble Farm L.F.
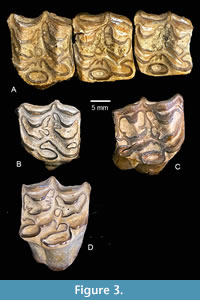 I reexamined the specimens assigned to Calippus proplacidus from Noble Farm and note consistent differences relative to the sample of this taxon from the Cold Spring L.F. (Figure 3). Plications on the prefossette are generally simpler, as is the pli caballin. The hypoconal groove was closed to an isolated lake earlier in wear. Although the sample size is limited, Calippus proplacidus from the Noble Farm L.F. is smaller than Calippus proplacidus from the Cold Spring L.F. and the protocone length is shorter. The small sample size renders the significance of these differences unclear. However, they generally reflect a more primitive morphotype in the specimens from Noble Farm. A few specimens from the Ba1 “Burkeville Fauna” were identified by Forsten (1975) as Calippus proplacidus (C. francisi), and Hulbert (1988) noted that they were more primitive than the material from Cold Spring. As an example, he listed TMM 40070-26 as an M1 with an MCH “of only 32.4 mm.” TMM 40070 (Summer’s Ranch) is a probable Ba1 locality in Washington County, TX from the lower part of the Fleming Group. The limited collection includes primarily isolated equid teeth, although TMM 40070-33 is a small, waterworn fragment of a possible proboscidean tooth. The material from Summers Ranch and Noble Farm suggests that Calippus proplacidus is known from both Ba1 and Ba2 faunas of the Texas Coastal Plain with the Ba1 forms reflecting a more primitive morphotype.
I reexamined the specimens assigned to Calippus proplacidus from Noble Farm and note consistent differences relative to the sample of this taxon from the Cold Spring L.F. (Figure 3). Plications on the prefossette are generally simpler, as is the pli caballin. The hypoconal groove was closed to an isolated lake earlier in wear. Although the sample size is limited, Calippus proplacidus from the Noble Farm L.F. is smaller than Calippus proplacidus from the Cold Spring L.F. and the protocone length is shorter. The small sample size renders the significance of these differences unclear. However, they generally reflect a more primitive morphotype in the specimens from Noble Farm. A few specimens from the Ba1 “Burkeville Fauna” were identified by Forsten (1975) as Calippus proplacidus (C. francisi), and Hulbert (1988) noted that they were more primitive than the material from Cold Spring. As an example, he listed TMM 40070-26 as an M1 with an MCH “of only 32.4 mm.” TMM 40070 (Summer’s Ranch) is a probable Ba1 locality in Washington County, TX from the lower part of the Fleming Group. The limited collection includes primarily isolated equid teeth, although TMM 40070-33 is a small, waterworn fragment of a possible proboscidean tooth. The material from Summers Ranch and Noble Farm suggests that Calippus proplacidus is known from both Ba1 and Ba2 faunas of the Texas Coastal Plain with the Ba1 forms reflecting a more primitive morphotype.
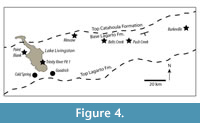 Cormohipparion goorsi was described from the Ba1 Trinity River Pit 1 L.F. in San Jacinto County (MacFadden and Skinner, 1981). Trinity River is a Ba1 local fauna from the middle part of the Lagarto Formation that Tedford et al. (2004) considered to be slightly younger than the Point Blank L.F. and slightly older than the Burkeville L.F. MacFadden (1984) Cormohipparion goorsi has also been reported from the Point Blank L.F. (TMM 31242), but MacFadden (1984) mistakenly considered Point Blank to be correlative with the Cold Spring L.F. The Point Blank L.F. is from quite low in the Lagarto Formation and is older than both the Trinity River Pit 1 L.F. and the Burkeville L.F. (Figure 1 and Figure 4) (Tedford et al., 1987; Prothero and Manning, 1987; Tedford et al., 2004; May and Brown, 2023). Cormohipparion cf. Cormohipparion goorsi is also present in the Noble Farm L.F. (TMM 40775-2544, Figure 4), as well as the Belts Creek L.F. consistent with the Ba1 age interpretation.
Cormohipparion goorsi was described from the Ba1 Trinity River Pit 1 L.F. in San Jacinto County (MacFadden and Skinner, 1981). Trinity River is a Ba1 local fauna from the middle part of the Lagarto Formation that Tedford et al. (2004) considered to be slightly younger than the Point Blank L.F. and slightly older than the Burkeville L.F. MacFadden (1984) Cormohipparion goorsi has also been reported from the Point Blank L.F. (TMM 31242), but MacFadden (1984) mistakenly considered Point Blank to be correlative with the Cold Spring L.F. The Point Blank L.F. is from quite low in the Lagarto Formation and is older than both the Trinity River Pit 1 L.F. and the Burkeville L.F. (Figure 1 and Figure 4) (Tedford et al., 1987; Prothero and Manning, 1987; Tedford et al., 2004; May and Brown, 2023). Cormohipparion cf. Cormohipparion goorsi is also present in the Noble Farm L.F. (TMM 40775-2544, Figure 4), as well as the Belts Creek L.F. consistent with the Ba1 age interpretation.
Protohippus perditus was identified by Hulbert (1988) from the Noble Farm L.F. based on a few isolated check teeth and one partial mandible (“TAM 3031-3033”, TAM 3035”). TAM 3035, now labeled TMM 38-3035, is from the Cold Spring L.F. TAM 3034 listed by Hulbert (1988) includes two specimens that are now labeled TMM 40775-3034 and TMM 40775-3385. TMM 40775-3385 is an unworn LM1 with dimensions of 20.4 mm x 17.5 mm, an MCH of 35.6 mm, a well-developed parastyle, and little cement. TMM 40775-3034 is a worn R M1/M2 with dimensions of 19.5 mm x 20.3 mm with a well-developed parastyle and little cement. These attributes are characteristic of Protohippus vetus, which is the more primitive morphotype of Protohippus and consistent with interpretation of Noble Farm as a Ba1 fauna. The R p3 or p4 (TMM 40775-3052), from Noble Farm noted by Hulbert (1988), is worn and could be either Protohippus vetus or Protohippus perditus. It appears that cheek teeth assignable to both Protohippus vetus and Protohippus perditus are present in the Ba1 Noble Farm L.F., although only Protohippus perditus is present in the Ba2 Cold Spring L.F.
Both Early and Late Barstovian faunas of the Texas Coastal Plain include as many as four rhinoceros’ taxa, including two smaller forms (Peraceras hessei, Teleoceras meridianum) and two larger forms (Aphelops megalodus, Teleoceras medicornutum) (Prothero and Manning, 1987). Peraceras hessei, Teleoceras meridianum, and Teleoceras medicornutum were identified from Noble Farm by Prothero and Manning (1987), but they included that fauna with the Late Barstovian “Cold Spring Fauna.” The presence of these three rhinoceros’ taxa is more consistent with Early Barstovian faunas. The only Late Barstovian occurrence of Teleoceras medicornutum in the Texas Coastal Plain according to Prothero and Manning (1987), was from Noble Farm. Recognizing that Noble Farm is a Ba1 fauna restricts Teleoceras medicornutum to Ba1 faunas in this region. The stratigraphic position within the Lagarto Formation and faunal composition indicate that the Noble Farm L.F. should be interpreted as a Ba1 fauna.
Brenham Local Fauna. TMM 40418 is a locality from a roadcut in the lower Lagarto Formation near Brenham, Texas, where a partial dentary was identified by Wilson (1960) as Amphicyon longiramus (Figure 2). At that time, Wilson (1960) concluded that TMM 40418 was equivalent to the ‘Cold Spring Fauna’ that he interpreted to be Hemingfordian in age. This was at least partly influenced by identifications of equid teeth from the locality including Protohippus and Calippus. Amphicyon longiramus is known from the Garvin Gully L.F. (Wilson, 1960), the Thomas Farm L.F. in Florida (Olsen, 1958), and the Pollack Farm L.F. in Delaware (Emry and Eshelman, 1998), all of which are Hemingfordian faunas. Differences in dental morphology were noted between the specimen from Brenham and the type material from Thomas Farm (Wilson, 1960). Amphicyon is known from He1 through Ba1 faunas in North America (Hunt, 1998; Tedford et al., 2004). Additional taxa from TMM 44018 include Prosynthetoceras and Cormohipparion sp., both of which are known from other Ba1 faunas in the Texas Coastal Plain. Stratigraphic position and faunal composition indicate that TMM 44018 is a Ba1 fauna.
Belts Creek Local Fauna. The Belts Creek locality is from the lower part of the Lagarto Formation southwest of Colmesneil, Texas (Figure 4). Specimens from this locality were originally collected by the property owners and by J.A. Wilson in 1963. Although the base of the Fleming Group can be difficult to identify in the field (see Albright, 1998), the approximate stratigraphic position of the Belts Creek L.F. is clear based on map patterns and bedding attitude. The Belts Creek L.F. is at least as old as the Ba1 Burkeville and Trinity River Pit 1 Local Faunas and has been interpreted as a Ba1 fauna based on stratigraphic position and faunal composition. The castorid Anchitheriomys buceei is present in the Belts Creek L.F. as well as several other Ba1 local faunas from the Lagarto Formation (May and Brown, 2023).
The Belts Creek locality was incorrectly included within the ‘Cold Spring Fauna’ by Prothero and Manning (1987). They considered it to be a Late Barstovian fauna, with no discussion other than a statement that the “mammalian stratigraphy of the Texas Gulf Coastal Plain has been extensively discussed by Hesse (1942), Quinn (1955), Wilson (1956), Patton (1969) and Patton and Taylor (1971, 1973)”. (Note: in an apparent typographical error, they list Belts Creek as TMM 40662 in their Table 1. It is TMM 40622). The rhinoceros Peraceras hessei from the Belts Creek L.F. (TMM 40622-1) is known from both Ba1 and Ba2 faunas of the Texas Coastal Plain (Prothero and Manning, 1987). TMM 40622-3 is a partial lower cheek tooth of a much larger rhinoceros and represents either Aphelops or Teleoceras. Both Ba1 and Ba2 faunas of the Texas Coastal Plain include as many as four rhino taxa, including two smaller forms (Peraceras hessei, Teleoceras meridianus) and two larger forms (Aphelops megalodus, Teleoceras medicornutum; Prothero and Manning, 1987). Therefore, the rhinoceros material from Belts Creek is consistent with both Ba1 and Ba2 faunas in the region.
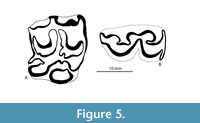 The Belts Creek L.F. includes an equid represented by a few isolated cheek teeth. TMM 40622-23 is a worn LP/M with L x W dimensions of 16.5 mm x 18.9 mm (Figure 5). The protocone is isolated with dimensions of 6.6 mm x 4.2 mm. The occlusal pattern compares favorably with Cormohipparion goorsi (MacFadden and Skinner, 1981). TMM 40622-22 is an unworn, broken RP/M with the protocone and hypocone missing. The tooth exhibits significant cement with maximum preserved dimensions of 19.9 mm x 19.0 mm. Dimensions near the base of the enamel (16.8 mm x 19.0 mm) are like TMM 40622-23. While the size and cement are consistent with Cormohipparion goorsi, the unworn mesostyle crown height (31.7 mm) is significantly taller than that of Cormohipparion goorsi reported previously.
The Belts Creek L.F. includes an equid represented by a few isolated cheek teeth. TMM 40622-23 is a worn LP/M with L x W dimensions of 16.5 mm x 18.9 mm (Figure 5). The protocone is isolated with dimensions of 6.6 mm x 4.2 mm. The occlusal pattern compares favorably with Cormohipparion goorsi (MacFadden and Skinner, 1981). TMM 40622-22 is an unworn, broken RP/M with the protocone and hypocone missing. The tooth exhibits significant cement with maximum preserved dimensions of 19.9 mm x 19.0 mm. Dimensions near the base of the enamel (16.8 mm x 19.0 mm) are like TMM 40622-23. While the size and cement are consistent with Cormohipparion goorsi, the unworn mesostyle crown height (31.7 mm) is significantly taller than that of Cormohipparion goorsi reported previously.
TMM 40622-19 is a moderately worn, Rp/m with dimensions of 18.2 mm x 11.0 mm. The ectoflexid extends laterally almost to the linguaflexid and the protostylid is well developed. This tooth compares favorably with the m1 of Cormohipparion goorsi figured by MacFadden and Skinner (1981) from the Ba1 Trinity River Pit I (F:AM 73948).
 Burkeville Local Fauna. Several authors previously interpreted the Burkeville L.F. as Ba1 fauna based on stratigraphic position within the Fleming Group and faunal composition (Tedford et al., 2004, May and Brown, 2023). The fauna was proposed as a possible Ba1 proboscidean occurrence by Tedford et al. (2004) based on fragments of teeth originally identified as ?Desmostylus by Stenzel, et al. (1944) that were later referred to either Proboscidea or indeterminate by Reinhart (1976). Further analysis of TMM 31160-2888 suggests that this fragment likely represents a rhinoceros, a relatively common group in the Burkeville L.F. There is a clear fold in the enamel fragment (Figure 6A, B) that is similar to morphologies observed in other rhinoceros’ teeth from this locality. While some fragments have relatively thick enamel, others have thin enamel like artiodactyl cheek teeth (Figure 6C). The specimens are too fragmentary to permit confident identification at the generic level and probably represent multiple taxa, but certainly not Desmostylus or Proboscidea.
Burkeville Local Fauna. Several authors previously interpreted the Burkeville L.F. as Ba1 fauna based on stratigraphic position within the Fleming Group and faunal composition (Tedford et al., 2004, May and Brown, 2023). The fauna was proposed as a possible Ba1 proboscidean occurrence by Tedford et al. (2004) based on fragments of teeth originally identified as ?Desmostylus by Stenzel, et al. (1944) that were later referred to either Proboscidea or indeterminate by Reinhart (1976). Further analysis of TMM 31160-2888 suggests that this fragment likely represents a rhinoceros, a relatively common group in the Burkeville L.F. There is a clear fold in the enamel fragment (Figure 6A, B) that is similar to morphologies observed in other rhinoceros’ teeth from this locality. While some fragments have relatively thick enamel, others have thin enamel like artiodactyl cheek teeth (Figure 6C). The specimens are too fragmentary to permit confident identification at the generic level and probably represent multiple taxa, but certainly not Desmostylus or Proboscidea.
Late Barstovian Faunas
Fossil proboscideans are also known from Ba2 local faunas from the upper part of the Lagarto Formation in the Texas Coastal Plain. Multiple specimens, from at least three different local faunas, are curated at the Texas Vertebrate Paleontology Collections. A number of these specimens were referred to in faunal lists as Trilophodon sp. by Hesse (1942) and to Gomphotherium by Tobien (1973). As discussed below, many of the previous interpretations require revision.
 Cold Spring Local Fauna. The Cold Spring Local Fauna from north of Cold Spring, Texas, is from the upper Lagarto Formation. This became the “type fauna” for the “regional biozone” named the ‘Cold Spring Fauna’ of Quinn (1955) and Wilson (1956). The Cold Spring Fauna was interpreted by Tedford et al. (2004) as consisting of the Cold Spring, Sam Houston, and Goodrich Local Faunas in decreasing age, and clearly separated from the Ba1 faunas of the lower Fleming Group. Consideration of the Cold Spring L.F. and Sam Houston L.F. as separate was the artificial construct of a forest boundary line that was cut through what was originally a single TAM locality (TAM 38, Hesse, 1942). The Sam Houston L.F. (TMM 31191) includes all of the material collected from within the Sam Houston National Forest. TMM 31219 refers specifically to the Donahoe Farm quarry within the larger TAM 38 locality area (Figure 7). Specimens collected from TAM 38, but not specifically assigned to either TMM 31191 or TMM 31219, are now assigned to TMM 38. All specimens from TMM 31191 (Sam Houston locality), TMM 31219 (Donahoe Farm locality) and TMM 38 are included in the Cold Spring L.F.
Cold Spring Local Fauna. The Cold Spring Local Fauna from north of Cold Spring, Texas, is from the upper Lagarto Formation. This became the “type fauna” for the “regional biozone” named the ‘Cold Spring Fauna’ of Quinn (1955) and Wilson (1956). The Cold Spring Fauna was interpreted by Tedford et al. (2004) as consisting of the Cold Spring, Sam Houston, and Goodrich Local Faunas in decreasing age, and clearly separated from the Ba1 faunas of the lower Fleming Group. Consideration of the Cold Spring L.F. and Sam Houston L.F. as separate was the artificial construct of a forest boundary line that was cut through what was originally a single TAM locality (TAM 38, Hesse, 1942). The Sam Houston L.F. (TMM 31191) includes all of the material collected from within the Sam Houston National Forest. TMM 31219 refers specifically to the Donahoe Farm quarry within the larger TAM 38 locality area (Figure 7). Specimens collected from TAM 38, but not specifically assigned to either TMM 31191 or TMM 31219, are now assigned to TMM 38. All specimens from TMM 31191 (Sam Houston locality), TMM 31219 (Donahoe Farm locality) and TMM 38 are included in the Cold Spring L.F.
Chappell Hill Local Fauna. A small assemblage of fossil vertebrates is known from a locality in Washington County near Chappell Hill, Texas. This locality is from the upper part of the Lagarto Formation and includes specimens assignable to Rhinocerotidae, Camelidae, and Equidae as well as a partial gomphothere skull (TMM 31272-42). A few isolated teeth compare well with Calippus proplacidus and Protohippus perditus from the Cold Spring L.F. Material from this locality was considered part of the ‘Cold Spring Fauna’ by Quinn (1955). Hulbert (1988) considered Calippus proplacidus and Protohippus perditus from the Chappell Hill “Site” to be Late Barstovian. Prothero and Manning (1987) identified a calcaneum (TMM 31272-24) from the Chappell Hill locality as Peraceras hessei and assigned it to the ‘Cold Spring fauna’. TMM 31272-42 was included by Tobien (1973) in his discussion of the small-sized Gomphotherium sp. from the ‘Cold Spring fauna.’ He cited Wilson (1962, fig. 2) as evidence for a Late Hemingfordian age, but failed to include Wilson (1962) in his reference citations. The reference most likely refers to a guidebook published for Field Excursion No. 10 by the Houston Geological Society (Wilson, 1962). Fig. 2, from that guidebook, is a correlation table in which Wilson showed the Fleming Group as late Arikareean and Hemingfordian in age and the Cold Spring Fauna as Late Hemingfordian. Although clearly superseded by more recent publications, that reference is most likely to what Tobien was referring to. Based on stratigraphic position and faunal composition, the Chappell Hill L.F. is a Ba2 fauna.
DISTRIBUTION OF FOSSIL PROBOSCIDEA IN THE TEXAS COASTAL PLAIN
The distribution of fossil proboscidea from the Texas Coastal Plain is summarized in Table 1.
Hemingfordian Proboscidea
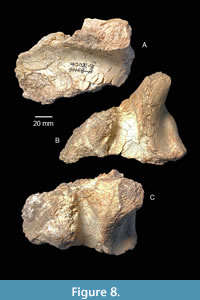 A survey of the collections at TxVP resulted in a single proboscidean specimen from TMM 40081 that J.A. Wilson apparently considered to be from the Oakville Formation and part of the “Garvin Gully Fauna.” TMM 40081-14 (renumbered from TMM 40081-3) was originally assigned the specimen number TMM 31158-1. Archival records at TxVP suggest that Wilson revisited this locality and collected additional material after which time all specimens were reassigned to TMM 40081. Based on the Geologic Map of Texas (Barnes et al., 1992), the upper boundary of the Oakville Formation is located just north of Brenham, Texas, near TMM 40081. Unfortunately, the small collection from TMM 40081 otherwise only includes undiagnostic, fragmentary specimens of equids and artiodactyls. TMM 40081-14 is a fragment of a thoracic vertebra of a large proboscidean (Figure 8). It may represent the earliest record of a proboscidean from the Miocene of the Texas Coastal Plain. Unfortunately, uncertainty about stratigraphic location and taxonomic identification limit the significance of this specimen. Based on size alone, it appears to be larger than would be expected for the small gomphotheres, that are described below, from Ba1 faunas in this region.
A survey of the collections at TxVP resulted in a single proboscidean specimen from TMM 40081 that J.A. Wilson apparently considered to be from the Oakville Formation and part of the “Garvin Gully Fauna.” TMM 40081-14 (renumbered from TMM 40081-3) was originally assigned the specimen number TMM 31158-1. Archival records at TxVP suggest that Wilson revisited this locality and collected additional material after which time all specimens were reassigned to TMM 40081. Based on the Geologic Map of Texas (Barnes et al., 1992), the upper boundary of the Oakville Formation is located just north of Brenham, Texas, near TMM 40081. Unfortunately, the small collection from TMM 40081 otherwise only includes undiagnostic, fragmentary specimens of equids and artiodactyls. TMM 40081-14 is a fragment of a thoracic vertebra of a large proboscidean (Figure 8). It may represent the earliest record of a proboscidean from the Miocene of the Texas Coastal Plain. Unfortunately, uncertainty about stratigraphic location and taxonomic identification limit the significance of this specimen. Based on size alone, it appears to be larger than would be expected for the small gomphotheres, that are described below, from Ba1 faunas in this region.
Early Barstovian Proboscidea
Noble Farm Local Fauna. A gomphothere from the Noble Farm L.F. was described by Hay (1925) based on a collection of “teeth and tusks” that was sent to him by Dr. Mark Francis at Texas A&M. They had been collected in 1915 at the farm of Edward Noble which was located 5 miles southeast of Navasota, Texas. No specimen numbers were included with Hay’s description; however, the figures can be used to identify specimens now curated at the Texas Vertebrate Paleontology Collections (Table 2).
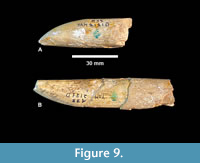 As described by Hay (1925), the proboscidean from Noble Farm includes down-curved upper tusks with an enamel band and no spiral. The tusks are slightly oval in cross-section. He hypothesized that at least four individuals were represented by the tusk fragments. At least one was a young individual represented by two lower tusks (TMM 40775-48.1 and 48.2) that are interpreted as deciduous and exhibit longitudinal striations with no spiral (Figure 9). Although broken, there is no evidence of a growth cone as seen in the ever-growing permanent tusks (lower and upper). Neither is there evidence of an enamel band, although both exhibit a wear facet along the distal end. The larger of the two juvenile lower tusks is ~90 mm long and both are slightly oval in cross-section with dimensions of 19.5 mm x 15.3 and 20.2 mm x 15.5 mm.
As described by Hay (1925), the proboscidean from Noble Farm includes down-curved upper tusks with an enamel band and no spiral. The tusks are slightly oval in cross-section. He hypothesized that at least four individuals were represented by the tusk fragments. At least one was a young individual represented by two lower tusks (TMM 40775-48.1 and 48.2) that are interpreted as deciduous and exhibit longitudinal striations with no spiral (Figure 9). Although broken, there is no evidence of a growth cone as seen in the ever-growing permanent tusks (lower and upper). Neither is there evidence of an enamel band, although both exhibit a wear facet along the distal end. The larger of the two juvenile lower tusks is ~90 mm long and both are slightly oval in cross-section with dimensions of 19.5 mm x 15.3 and 20.2 mm x 15.5 mm.
 TMM 40775-59 is a partial left upper tusk 488 mm long with a proximal cross-section of 61 mm x 50 mm and a distal cross-section of 48 mm x 41 mm (Figure 10A, B). The tusk is down-curved with an enamel band on the labial side. The enamel band is 31.5 mm wide at the distal end of the specimen and 25 mm wide at the proximal end, where it interfingers with cementum. Although the most anterior portion of the tusk is missing, a large wear facet is present on the ventro-lingual side of the tusk. That wear facet is sub-oval in shape and extends to the ventral edge of the enamel band. A relatively sharp cutting edge is preserved at the boundary between the wear facet and the enamel band. The preserved wear facet is 196 mm long and 36 mm wide (Figure 10A).
TMM 40775-59 is a partial left upper tusk 488 mm long with a proximal cross-section of 61 mm x 50 mm and a distal cross-section of 48 mm x 41 mm (Figure 10A, B). The tusk is down-curved with an enamel band on the labial side. The enamel band is 31.5 mm wide at the distal end of the specimen and 25 mm wide at the proximal end, where it interfingers with cementum. Although the most anterior portion of the tusk is missing, a large wear facet is present on the ventro-lingual side of the tusk. That wear facet is sub-oval in shape and extends to the ventral edge of the enamel band. A relatively sharp cutting edge is preserved at the boundary between the wear facet and the enamel band. The preserved wear facet is 196 mm long and 36 mm wide (Figure 10A).
TMM 40775-2121 includes a nearly complete right upper tusk and part of the right premaxillary bone (Figure 10C, D). The distal part of this tusk (figured by Hay, 1925) was previously removed and incorporated into a 1939 TAM museum mount of a partial gomphothere skull from the Ba2 Cold Spring L.F. It was recently removed from that mount and returned to the material from Noble Farm. When reconstructed, the length of the tusk is ~517 mm. It is down-curved with a labial enamel band that is ~28 mm wide and exhibits inter-fingering with cementum at the proximal end. The distal tip of the tusk is rounded from wear and the enamel band shows clear signs of having been broken into a saw-blade pattern during the lifetime of the gomphothere (Figure 10B). The broken edge of the enamel band experienced post-damage wear pre-mortem. Whether this damage was the result of fighting, feeding, or some other behavior is unknown.
A linear feature in the premaxilla was interpreted by Hay (1925) as the suture between the premaxilla and maxilla (his fig. 4, page 11), but he apparently did not recognize that TAM 2213 was part of this specimen. Not only is it part of the original specimen, TAM 2213 preserves the termination of the feature suggesting it may be a taphonomic crack associated with desiccation prior to burial (Figure 10D). Additional desiccation cracks, subparallel to this feature, are present. The cross-sectional dimensions of the tusk at the broken anterior end are 60 mm x 45 mm.
 The only specimens of adult lower tusks are TMM 40775-51 and TMM 40775-56 that are here interpreted as being from the same individual. They are associated with a partial symphysis (Figure 11). The anterior cross-section of TMM 40775-51 is sub-oval in shape with dimensions of 47 mm x 32 mm and shows tightly packed laminations with dense crosshatching (Figure 11C). The mandibular tusks are placed closely together with only 5 mm separating them at the distal end of the preserved portion of the symphysis. The cross-sectional shape of the lower tusks differs from the pyriform lower tusks seen in forms like Gomphotherium angustidens and Gomphotherium subtapiroideum. Only the proximal ends of these lower tusks are preserved, so the geometry of the distal ends is unknown. The distal ends of the juvenile tusks (TMM 40775-48.1, TMM 40775-48.2) are sub-oval in cross-section.
The only specimens of adult lower tusks are TMM 40775-51 and TMM 40775-56 that are here interpreted as being from the same individual. They are associated with a partial symphysis (Figure 11). The anterior cross-section of TMM 40775-51 is sub-oval in shape with dimensions of 47 mm x 32 mm and shows tightly packed laminations with dense crosshatching (Figure 11C). The mandibular tusks are placed closely together with only 5 mm separating them at the distal end of the preserved portion of the symphysis. The cross-sectional shape of the lower tusks differs from the pyriform lower tusks seen in forms like Gomphotherium angustidens and Gomphotherium subtapiroideum. Only the proximal ends of these lower tusks are preserved, so the geometry of the distal ends is unknown. The distal ends of the juvenile tusks (TMM 40775-48.1, TMM 40775-48.2) are sub-oval in cross-section.
There are no enamel bands on the lower tusks although a 1-2 mm thick layer of cementum covers the underlying dentin. Shallow longitudinal ridges are present in the dentin. The lower tusks are ever-growing, as evidenced by the growth cones apparent in the pulp cavity at the proximal end. The partial symphysis is approximately 50 mm wide and slightly concave on the dorsal surface (Figure 11A). The fragment of symphysis is not sufficient to directly determine the length or morphology of the mandible, but the size and geometry of the tusks suggests that the gomphothere from Noble Farm was likely a longirostrine gomphothere.
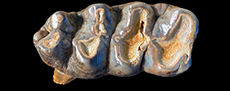
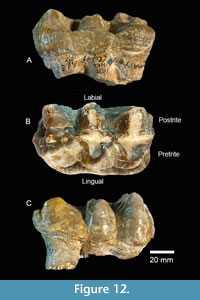 TMM 40775-49 is a worn RM2, 105.1 mm long by 65.7 mm in maximum width, with three lophs, a well-developed lingual cingulum and simple pretrite trefoils (Figure 12). Measured widths at each loph include first: 65.5 mm, second: 65.7 mm, and third: 65.3 mm. The crown height, at the slightly worn third loph, is 43.8 mm. An antero-labial and a posterior cingulum are weakly developed. Pretrite valleys are relatively wide, while postrite valleys are narrow. The median sulcus is weakly developed and minor amounts of cementum are present in transverse valleys.
TMM 40775-49 is a worn RM2, 105.1 mm long by 65.7 mm in maximum width, with three lophs, a well-developed lingual cingulum and simple pretrite trefoils (Figure 12). Measured widths at each loph include first: 65.5 mm, second: 65.7 mm, and third: 65.3 mm. The crown height, at the slightly worn third loph, is 43.8 mm. An antero-labial and a posterior cingulum are weakly developed. Pretrite valleys are relatively wide, while postrite valleys are narrow. The median sulcus is weakly developed and minor amounts of cementum are present in transverse valleys.
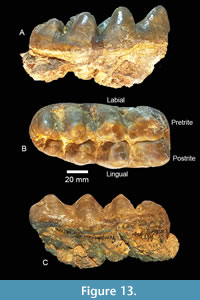 TMM 40775-46 is a Lm3 with three main lophids, a smaller fourth lophid and a small talonid (Figure 13). The tooth is slightly worn (young adult) but would develop simple pretrite trefoils on the three anterior lophids with additional wear. It has a maximum length x width of 136.6 mm x 64.0 mm (at second lophid) with a crown height of 44.7 mm at the second lophid and 42.7 mm at the third. The m3 lacks a well-defined lateral cingulum although a weak antero-labial cingulum is present. The median sulcus is well developed. Transverse valleys are relatively tight and lack secondary lophids. Minor amounts of cementum are present in the talonid and between the third and fourth lophids.
TMM 40775-46 is a Lm3 with three main lophids, a smaller fourth lophid and a small talonid (Figure 13). The tooth is slightly worn (young adult) but would develop simple pretrite trefoils on the three anterior lophids with additional wear. It has a maximum length x width of 136.6 mm x 64.0 mm (at second lophid) with a crown height of 44.7 mm at the second lophid and 42.7 mm at the third. The m3 lacks a well-defined lateral cingulum although a weak antero-labial cingulum is present. The median sulcus is well developed. Transverse valleys are relatively tight and lack secondary lophids. Minor amounts of cementum are present in the talonid and between the third and fourth lophids.
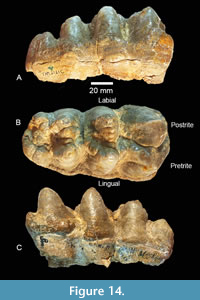 TMM 40775-47 is a RM3 that is slightly worn and exhibits strong lingual and antero-labial cingula (Figure 14). There are three main lophs and a talon consisting of five cones. The main lophs would develop simple pretrite trefoils with additional wear. This tooth is 136.6 mm x 70.4 mm, with a crown height at the second loph of 48.7 mm and 43.9 mm at the third. The median sulcus is relatively shallow, and the transverse valleys are relatively narrow. An accessory cone is present on the postero-labial side of the third loph.
TMM 40775-47 is a RM3 that is slightly worn and exhibits strong lingual and antero-labial cingula (Figure 14). There are three main lophs and a talon consisting of five cones. The main lophs would develop simple pretrite trefoils with additional wear. This tooth is 136.6 mm x 70.4 mm, with a crown height at the second loph of 48.7 mm and 43.9 mm at the third. The median sulcus is relatively shallow, and the transverse valleys are relatively narrow. An accessory cone is present on the postero-labial side of the third loph.
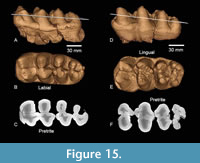 Figure 15 illustrates the enamel pattern from an arbitrary slice (white line in A and D) through the CT volumes for TMM 40775-46 and TMM 40775-47. These images illustrate the simple trefoil pattern characteristic of gomphotheriids that would be observed in these specimens with additional wear (labial in m3 and lingual in M3).
Figure 15 illustrates the enamel pattern from an arbitrary slice (white line in A and D) through the CT volumes for TMM 40775-46 and TMM 40775-47. These images illustrate the simple trefoil pattern characteristic of gomphotheriids that would be observed in these specimens with additional wear (labial in m3 and lingual in M3).
The third molars of the Noble Farm gomphothere are relatively small and plot near the lower end of all Gomphotherium taxa sampled by Wang et al. (2017) and are much smaller than specimens identified as Gomphotherium productum. They are also significantly shorter than measurements compiled by Madden and Storer (1985) for M3 of Gomphotherium obscurum, as well as being smaller and lower crowned than the M3 of Gomphotherium osborni (190 mm x 87 mm) (Madden and Storer, 1985). Similarly sized taxa, based on the data in Wang et al. (2017), include Gomphotherium inopinatum, Gomphotherium angustidens, Gomphotherium mongoliense, Gomphotherium subtapiroideum, and Gomphotherium annectens. M2 (TMM 40775-49) is similar morphologically to Gomphotherium subtapiroideum including the loph and valley geometry emphasized by Wang et al. (2017).
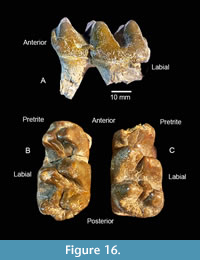 Two small cheek teeth were interpreted by Hay (1925) as L and R Dp3. These teeth are only slightly worn with three lophids and a small posterior heel (Figure 16). Based on the analysis of dental ages within a large sample of Gomphotherium angustidens by Tassy (2013), TMM 40775-50.1 and 50.2 are now interpreted as L and R Dp4. TMM 40775-50.1 is an L Dp4 with maximum length of 59.4 mm, width at the anterior lophid of 27.4 mm, and width at posterior lophid of 35.8 mm. TMM 40775-50.2 is essentially the same size, although the anterior portion is broken. When compared to TMM40418-10, from the Ba1 Brenham L.F., these teeth are consistent with identification as Dp4. Dp3 and Dp4 are replaced by p3 and p4 while m1 is in early occlusion in Gomphotherium angustidens (Tassy, 2013).
Two small cheek teeth were interpreted by Hay (1925) as L and R Dp3. These teeth are only slightly worn with three lophids and a small posterior heel (Figure 16). Based on the analysis of dental ages within a large sample of Gomphotherium angustidens by Tassy (2013), TMM 40775-50.1 and 50.2 are now interpreted as L and R Dp4. TMM 40775-50.1 is an L Dp4 with maximum length of 59.4 mm, width at the anterior lophid of 27.4 mm, and width at posterior lophid of 35.8 mm. TMM 40775-50.2 is essentially the same size, although the anterior portion is broken. When compared to TMM40418-10, from the Ba1 Brenham L.F., these teeth are consistent with identification as Dp4. Dp3 and Dp4 are replaced by p3 and p4 while m1 is in early occlusion in Gomphotherium angustidens (Tassy, 2013).
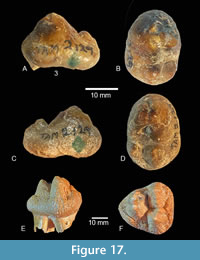 Additional cheek teeth, apparently associated with the Noble Farm gomphothere, include R and L p3 as interpreted by Hay (1925, his plate 3 and figs. 2, 3). These specimens are unworn and include caps with no roots (Figure 17). Their assignment as permanent premolars is uncertain. Length x width measurements for these two teeth are 28.5 mm x 20.1 mm and 26.2 x 18.9 mm. An additional juvenile cheek tooth (TMM 40775-61) contains no TAM number and was not included by Hay in his 1925 paper. The provenance of this specimen is uncertain, but it may represent a premolar (P/p2?).
Additional cheek teeth, apparently associated with the Noble Farm gomphothere, include R and L p3 as interpreted by Hay (1925, his plate 3 and figs. 2, 3). These specimens are unworn and include caps with no roots (Figure 17). Their assignment as permanent premolars is uncertain. Length x width measurements for these two teeth are 28.5 mm x 20.1 mm and 26.2 x 18.9 mm. An additional juvenile cheek tooth (TMM 40775-61) contains no TAM number and was not included by Hay in his 1925 paper. The provenance of this specimen is uncertain, but it may represent a premolar (P/p2?).
The proboscidean from the Noble Farm L.F. belongs to Gomphotheriinae based on the presence of accessory conules on the pretrite half-lophids forming trefoil patterns, a relatively narrow median sulcus, trilophodont M2/m2, incipient tetralophodonty on M3/m3, down-turned upper tusks with enamel bands, and lower tusks without enamel bands. The specimens were identified by Hay (1925) as “Cope’s form” Tetrabelodon serridens cimarronis which he renamed Gomphotherium cimarronis (Cope). A single tooth was assigned by Cope (1893) as Tetrabelodon serridens?, although he stated significant doubt as to that identification because of the small size. In fact, he said that the small size “renders the identification doubtful, unless it indicates a small race of the same species.” “It may pertain to a different species, but for the present I refer it to a small race of the T. serridens, under the name of T. serridens cimarronis.” This cautious attempt to recognize a subspecies (Cope’s “race”) was elevated to species status by Hay (1925). The “type” specimen of Tetrabelodon serridens cimarronis (no specimen number and no explicit holotype designation) was described by Cope (1893) as a left m3 from the “Loup Fork beds north of the South Fork of the Red River.” Hay (1925) disagreed and correctly interpreted the tooth as an M3. The specimen is curated in the Texas Vertebrate Paleontology Collection at the University of Texas, Austin (TMM 40282-12). It is included with several specimens from the Clarendonian Stanton Ranch locality (TMM 40282) near Clarendon, Texas. Gomphotherium cimarronis was interpreted by Tobien (1973) as a junior synonym of Gomphotherium productum.
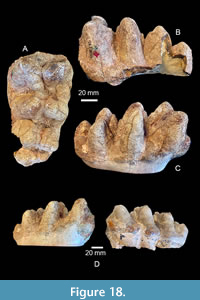 In their review of North American occurrences of Gomphotherium, Lambert and Shoshani (1998) only recognized Gomphotherium angustidens and Gomphotherium productum, but also listed many previously named species under Gomphotherium sp. They include Gomphotherium cimarronis in the later group suggesting uncertain validity for the taxon. Regardless of the validity of the name Gomphotherium cimarronis, the “type” specimen of the taxon from the Stanton Ranch L.F. is clearly different from the gomphothere from Noble Farm (Figure 18). TMM 40282-12 from Stanton Ranch is 76.6 mm wide at the first loph, 70.0 mm wide at the second, and at least 58 mm wide at the third (now partially broken, even though it was shown as complete by Cope (1893)). TMM 40775-47 (RM3) is significantly narrower at the first loph (69.9 mm). The crown height of the “holotype” of Gomphotherium cimarronis is much taller: 65.7 mm at the first loph and 62.7 mm at the second, compared to 43.9 mm and 48.7 mm for the same lophs on the gomphothere from Noble Farm. The talon is also different being composed of only three conules, instead of the five to six seen in the specimen from Noble Farm. The lingual (pretrite) cingulum is also weaker in Gomphotherium cimarronis, although it is partially missing now along the posterior part of the tooth. Although specimens from both the Stanton Ranch and Noble Farm Local Faunas represent a relatively small gomphothere with simple pretrite trefoils, they are not the same taxon. Furthermore, Gomphotherium cimarronis, from the Clarendonian of the Texas panhandle, is considered nomen dubium because the “holotype” is a partial M2 lacking diagnostic characters.
In their review of North American occurrences of Gomphotherium, Lambert and Shoshani (1998) only recognized Gomphotherium angustidens and Gomphotherium productum, but also listed many previously named species under Gomphotherium sp. They include Gomphotherium cimarronis in the later group suggesting uncertain validity for the taxon. Regardless of the validity of the name Gomphotherium cimarronis, the “type” specimen of the taxon from the Stanton Ranch L.F. is clearly different from the gomphothere from Noble Farm (Figure 18). TMM 40282-12 from Stanton Ranch is 76.6 mm wide at the first loph, 70.0 mm wide at the second, and at least 58 mm wide at the third (now partially broken, even though it was shown as complete by Cope (1893)). TMM 40775-47 (RM3) is significantly narrower at the first loph (69.9 mm). The crown height of the “holotype” of Gomphotherium cimarronis is much taller: 65.7 mm at the first loph and 62.7 mm at the second, compared to 43.9 mm and 48.7 mm for the same lophs on the gomphothere from Noble Farm. The talon is also different being composed of only three conules, instead of the five to six seen in the specimen from Noble Farm. The lingual (pretrite) cingulum is also weaker in Gomphotherium cimarronis, although it is partially missing now along the posterior part of the tooth. Although specimens from both the Stanton Ranch and Noble Farm Local Faunas represent a relatively small gomphothere with simple pretrite trefoils, they are not the same taxon. Furthermore, Gomphotherium cimarronis, from the Clarendonian of the Texas panhandle, is considered nomen dubium because the “holotype” is a partial M2 lacking diagnostic characters.
Based on L x W measurements for M2, M3, and m3, the gomphothere from Noble Farm plots near the center of the distribution of sizes compiled by Sanders and Miller (2002) for Gomphotherium angustidens. However, significant sexual dimorphism within M3 length for Gomphotherium angustidens was documented by Tassy (2014), and the gomphothere from Noble Farm plots near the low end of the males. Unlike Gomphotherium angustidens, the upper tusks show no evidence of the spiral architecture described by Tassy (2014), and the lower tusks appear to be straighter and sub-oval in cross-section, rather than pyriform. Differences in tusk morphology, with respect to both Gomphotherium productum and Gomphotherium angustidens were also discussed by Hay (1925). The M3 of Gomphotherium agustidens has four lophs while the gomphothere from Noble Farm has only three main lophs and an incipient fourth loph with a posterior heel. Central conules are also less well developed in the gomphothere from Noble Farm.
The gomphothere from Noble Farm differs from Gomphotherium angustidens, Gomphotherium productum, and from the “holotype” of Gomphotherium cimarronis. The cheek tooth morphology is similar to Gomphotherium subtapiroideum, while the mandibular tusks are similar to Gomphotherium productum. Similarity to Gomphotherium subtapiroideum (Mastodon angustidens subtapiroidea, Schlesinger, 1917) was noted by Hay (1925), but Wang et al. (2020) have described the lower tusks of Gomphotherium subtapiroideum as pyriform, rather than sub oval as seen in the gomphothere from Noble Farm (Figure 11). They interpret pyriform lower tusks to be a plesiomorphic character for elephantimorphs with rounded or oval tusks being a derived morphology. The gomphothere from Noble Farm is a relatively primitive morphotype that is smaller and lower crowned than Gomphotherium productum and differs in dental morphology from Gomphotherium angustidens. It is possible that the gomphothere from Noble Farm is a primitive morphotype of Gomphotherium productum, but it is here referred to as Gomphotherium. sp. Although representing a similarly conservative morphotype, it differs in detail from the gomphothere in the Ba1 Belts Creek L.F. described below.
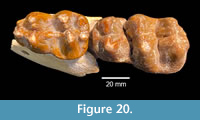
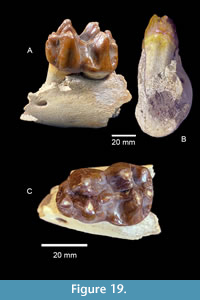 Brenham Local Fauna. TMM 40418-10 is a partial left dentary of a juvenile gomphothere with a deciduous premolar and a matrix-filled alveolus for a deciduous lower tusk (Figure 19B). The tooth is a slightly worn Dp3 with a maximum length of 42.6 mm. There are two primary lophids, an incipient anterior lophid, and a small posterior heel. The tooth is 29.0 mm wide at the second lophid and 23.8 mm wide at the first lophid. The median sulcus is moderately well developed and a small accessory conulid is present in the labial side of the transverse valley between the lophids. A relatively large mental foramen is located on the labial side of the dentary, just anterior to Dp3. A sub-circular alveolus for a deciduous lower tusk is approximately 17 mm in diameter. This specimen compares favorably with Gomphotherium sp. from the Noble Farm L.F., extends the geographic range of the taxon and supports the Ba1 faunal interpretation (Figure 20). TMM 40418-10 was previously misidentified as Dinohyus.
Brenham Local Fauna. TMM 40418-10 is a partial left dentary of a juvenile gomphothere with a deciduous premolar and a matrix-filled alveolus for a deciduous lower tusk (Figure 19B). The tooth is a slightly worn Dp3 with a maximum length of 42.6 mm. There are two primary lophids, an incipient anterior lophid, and a small posterior heel. The tooth is 29.0 mm wide at the second lophid and 23.8 mm wide at the first lophid. The median sulcus is moderately well developed and a small accessory conulid is present in the labial side of the transverse valley between the lophids. A relatively large mental foramen is located on the labial side of the dentary, just anterior to Dp3. A sub-circular alveolus for a deciduous lower tusk is approximately 17 mm in diameter. This specimen compares favorably with Gomphotherium sp. from the Noble Farm L.F., extends the geographic range of the taxon and supports the Ba1 faunal interpretation (Figure 20). TMM 40418-10 was previously misidentified as Dinohyus.
 Belts Creek Local Fauna. A gomphotheriid proboscidean from the Belts Creek L.F. is represented by a single cheek tooth (Figure 21). TMM 40622-25 is a moderately worn Rm3 with an occlusal pattern that is characterized by simple pretrite trefoils, with three main lophids, a smaller fourth lophid and a small posterior conulid. The maximum length of this tooth is 150 mm and the width varies from 69.1 mm at the first lophid, to 76.0 mm at the second, and 76.2 mm at the third lophid. The fourth lophid is 55.6 mm wide, and the posterior heel is a single conulid. A minimum crown height of ~44.3 mm was measured on the lingual side of the slightly worn third lophid. The anterior cingulum is worn nearly flat and accessory conulids are present only on the lingual (postrite) margin between lophids. A weak median sulcus is present and accessory conulids are absent from the transverse valleys. Postrite valleys are relatively broad at their base while pretrite valleys are narrow. There is no lingual cingulum and no cementum between the lophids.
Belts Creek Local Fauna. A gomphotheriid proboscidean from the Belts Creek L.F. is represented by a single cheek tooth (Figure 21). TMM 40622-25 is a moderately worn Rm3 with an occlusal pattern that is characterized by simple pretrite trefoils, with three main lophids, a smaller fourth lophid and a small posterior conulid. The maximum length of this tooth is 150 mm and the width varies from 69.1 mm at the first lophid, to 76.0 mm at the second, and 76.2 mm at the third lophid. The fourth lophid is 55.6 mm wide, and the posterior heel is a single conulid. A minimum crown height of ~44.3 mm was measured on the lingual side of the slightly worn third lophid. The anterior cingulum is worn nearly flat and accessory conulids are present only on the lingual (postrite) margin between lophids. A weak median sulcus is present and accessory conulids are absent from the transverse valleys. Postrite valleys are relatively broad at their base while pretrite valleys are narrow. There is no lingual cingulum and no cementum between the lophids.
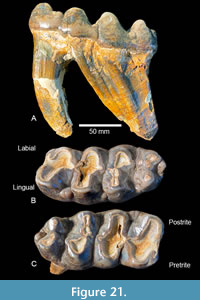 Morphologically, the Belts Creek proboscidean is assignable to the genus Gomphotherium and the bunodont, pretrite trefoil morphology distinguishes TMM 40622-25 from Zygolophodon. However, species-level identification becomes difficult partly because it is a single tooth and partly because of the current state of the taxonomy of the genus. Eurasian species of Gomphotherium were reviewed by Wang et al. (2017), and, based on their compilations, the Gomphotherium from Belts Creek compares well with the small- to moderate-sized taxa with m3 dimensions similar to Gomphotherium inopinatum, Gomphotherium subtapiroideum, Gomphotherium angustidens, Gomphotherium cooperi, and Gomphotherium sylvaticum. In terms of size, crown height and occlusal morphology, TMM 40622-25 appears most similar to Gomphotherium inopinatum figured by Wang (2014; IVPP V 18700). The fourth lophid on TMM 40622-25 is slightly better developed and the posterior conulid appears absent on Gomphotherium inopinatum. TMM 40622-25 also has postrite conulids between the lophids, a character that appears absent in Gomphotherium inopinatum. Gomphotherium inopinatum is known from the Linxia Basin in central China, from the lower part of the Dongxiang Formation. According to Wang (2014), this formation is late Early Miocene in age or approximately equivalent to the Hemingfordian NALMA. Gomphotherium inopinatum was interpreted as a basal species of Gomphotherium and possibly ancestral to Gomphotherium angustidens Wang (2014). The type locality of Gomphotherium inopinatum is from the Turgai region in Kazakhstan and was considered to be Lower Miocene by (Tassy, 1985). In their phylogenetic analysis of Gomphotherium, Wang et al. (2017) recovered Gomphotherium inopinatum as a basal member of the “G. angustidens group” with an age range of approximately 15-17 Ma. The morphological differences between the gomphothere from Belts Creek and Gomphotherium inopinatum suggest a slightly more derived morphotype in the Texas form.
Morphologically, the Belts Creek proboscidean is assignable to the genus Gomphotherium and the bunodont, pretrite trefoil morphology distinguishes TMM 40622-25 from Zygolophodon. However, species-level identification becomes difficult partly because it is a single tooth and partly because of the current state of the taxonomy of the genus. Eurasian species of Gomphotherium were reviewed by Wang et al. (2017), and, based on their compilations, the Gomphotherium from Belts Creek compares well with the small- to moderate-sized taxa with m3 dimensions similar to Gomphotherium inopinatum, Gomphotherium subtapiroideum, Gomphotherium angustidens, Gomphotherium cooperi, and Gomphotherium sylvaticum. In terms of size, crown height and occlusal morphology, TMM 40622-25 appears most similar to Gomphotherium inopinatum figured by Wang (2014; IVPP V 18700). The fourth lophid on TMM 40622-25 is slightly better developed and the posterior conulid appears absent on Gomphotherium inopinatum. TMM 40622-25 also has postrite conulids between the lophids, a character that appears absent in Gomphotherium inopinatum. Gomphotherium inopinatum is known from the Linxia Basin in central China, from the lower part of the Dongxiang Formation. According to Wang (2014), this formation is late Early Miocene in age or approximately equivalent to the Hemingfordian NALMA. Gomphotherium inopinatum was interpreted as a basal species of Gomphotherium and possibly ancestral to Gomphotherium angustidens Wang (2014). The type locality of Gomphotherium inopinatum is from the Turgai region in Kazakhstan and was considered to be Lower Miocene by (Tassy, 1985). In their phylogenetic analysis of Gomphotherium, Wang et al. (2017) recovered Gomphotherium inopinatum as a basal member of the “G. angustidens group” with an age range of approximately 15-17 Ma. The morphological differences between the gomphothere from Belts Creek and Gomphotherium inopinatum suggest a slightly more derived morphotype in the Texas form.
Although known from a single molar, the gomphothere from the Belts Creek L.F. represents a conservative species of Gomphotherium and is herein identified as Gomphotherium cf. Gomphotherium inopinatum. This suggests that a species of Gomphotherium, like Gomphotherium inopinatum, immigrated from Eurasia to North America, probably during the Hemingfordian, arriving in Texas by the Early Barstovian. Gomphotherium cf. Gomphotherium inopinatum from the Belts Creek L.F. is morphologically distinct from the Gomphotherium sp. from Noble Farm described above. The third lower molar is longer (150 mm vs. 136.6 mm), has a weaker median sulcus and a less complex trefoil pattern. The posterior heel is simpler (single conulid), and the postrite valleys are wider at the base with better developed accessory conulids. At least two different species of Gomphotherium are present in Ba1 faunas of the Texas Coastal Plain.
Discussion. There is significant confusion in the literature regarding interpretation of Early Barstovian proboscideans from the Miocene Fleming Group localities of the Texas Coastal Plain. Specimens from the Burkeville L.F. were cited by Tedford et al. (2004) as a possible Ba1 proboscidean occurrence, but Prothero et al. (2006) stated “Only the High Plains and the Texas Gulf Coastal Plain show no evidence of early Barstovian proboscideans,...” Previous authors misinterpreted the ages of the Noble Farm, Belts Creek, and Brenham Local Faunas, but all of these are here shown to be Ba1 based on stratigraphic position and faunal evidence. It is now clear that at least two different species of Gomphotherium are known from Ba1 local faunas in the lower to middle Lagarto Formation. Gomphotherium cf. Gomphotherium inopinatum is known from a single m3 from the Belts Creek L.F. while Gomphotherium sp. is known from multiple specimens from the Noble Farm and Brenham Local Faunas. Gomphotherium sp. from Noble Farm should not be referred to Gomphotherium cimarronis as suggested by Hay (1925). The Ba1 gomphotheres from the Lagarto Formation are all relatively small with primitive dental morphology and probably represent early immigrants from Asia.
Late Barstovian Proboscidea
Fossil proboscidea are known from the Cold Spring (including Sam Houston) and Chappell Hill local faunas from the upper part of the Lagarto Formation.
 Cold Spring Fauna. Trilophodon sp. was included in faunal lists by Hesse (1942) for both the “Sam Houston” and “Cold Spring fauna”. As discussed above, the “Sam Houston fauna” is now considered a locality within the Cold Spring L.F. Material cited by Hesse (1942) in the faunal list for the ‘Cold Spring Fauna’ included “ Trilophodon, fair skull in A&M Museum”. This description undoubtedly refers to TMM 38-3046 that was mounted for display at TAM in 1939. TMM 38-3046 includes fragments of both a partial skull and a partial left dentary, however, the amount of plaster reconstruction limits the ability to provide a useful description (Figure 22, Figure 23). The upper tusk, incorporated into the mount, is TMM 40775-2121 from the Ba1 Noble Farm L.F. and does not belong to the mounted skull. Other than the fragments of tusk associated with TMM 38-3046, there are no known upper tusks from gomphotheres found in any of the Ba2 local faunas of the upper Lagarto Formation. The tooth associated with the partial skull identified as an M3 by Tobien (1973), is largely reconstructed from plaster and has a coating of plaster covering the occlusal surface.
Cold Spring Fauna. Trilophodon sp. was included in faunal lists by Hesse (1942) for both the “Sam Houston” and “Cold Spring fauna”. As discussed above, the “Sam Houston fauna” is now considered a locality within the Cold Spring L.F. Material cited by Hesse (1942) in the faunal list for the ‘Cold Spring Fauna’ included “ Trilophodon, fair skull in A&M Museum”. This description undoubtedly refers to TMM 38-3046 that was mounted for display at TAM in 1939. TMM 38-3046 includes fragments of both a partial skull and a partial left dentary, however, the amount of plaster reconstruction limits the ability to provide a useful description (Figure 22, Figure 23). The upper tusk, incorporated into the mount, is TMM 40775-2121 from the Ba1 Noble Farm L.F. and does not belong to the mounted skull. Other than the fragments of tusk associated with TMM 38-3046, there are no known upper tusks from gomphotheres found in any of the Ba2 local faunas of the upper Lagarto Formation. The tooth associated with the partial skull identified as an M3 by Tobien (1973), is largely reconstructed from plaster and has a coating of plaster covering the occlusal surface.
 The anterior portion of the skull (premaxilla, maxilla, frontals) includes some original bone, although much of the skull is either sculpted from plaster or coated with plaster and paint. The specific dimensions are of limited value, but the length of the “skull”, from the broken edge of the premaxilla to the posterior edge of the cranium, is 55.5 cm. The height, from the ventral side of the zygomatic arch to the dorsal side of the cranium, is 38.5 cm. The height, from the top of the tooth row to the top of the premaxilla, is 14 cm. The length of the zygomatic arch is approximately 32 cm. If the reconstruction honored at least some original bone morphology, this is a small gomphothere and similar in size to TMM 31219-237 and to the skull from the Ba2 locality near Chappell Hill, TX (TMM 31272-42).
The anterior portion of the skull (premaxilla, maxilla, frontals) includes some original bone, although much of the skull is either sculpted from plaster or coated with plaster and paint. The specific dimensions are of limited value, but the length of the “skull”, from the broken edge of the premaxilla to the posterior edge of the cranium, is 55.5 cm. The height, from the ventral side of the zygomatic arch to the dorsal side of the cranium, is 38.5 cm. The height, from the top of the tooth row to the top of the premaxilla, is 14 cm. The length of the zygomatic arch is approximately 32 cm. If the reconstruction honored at least some original bone morphology, this is a small gomphothere and similar in size to TMM 31219-237 and to the skull from the Ba2 locality near Chappell Hill, TX (TMM 31272-42).
 The partial left dentary mounted with TMM 31219-3046 has no visible specimen number. Handwritten notes state “left side of skull and left mandible”, collected “1.3 miles north of the court house, Cold Spring, San Jacinto Co., Texas” in 1934 by Claude Riley. The worn lower m3 incorporated into the dentary only includes a small amount of original tooth (lingual margin) and is mostly modeled in plaster. The occlusal pattern is of no value and the tooth appears to be attached to the dentary with plaster. The lower tusk, associated with the dentary, also appears to be set in plaster at the alveolus with no actual fit and therefore is of uncertain provenance. This tusk is somewhat oval in cross-section (38 mm x 32 mm) at the broken proximal end (Figure 24). The total length of the tusk fragment is ~280 mm and is relatively straight with no enamel band. It is significantly smaller than the lower tusk of Gomphotherium sp. from the Sam Houston locality (TMM 31191-2233) (53 mm x 47 mm cross-section diameters and 416 mm length). The latter appears less compressed in cross-section, but the significance of these differences is uncertain and could easily reflect ontogeny or sexual dimorphism. Neither specimen exhibits growth cones, suggesting the proximal ends are missing. The sub-oval cross-section of the mandibular tusks differs from the dorso-ventrally flattened lower tusks seen in the holotype of Gomphotherium productum from New Mexico (Morgan 2015).
The partial left dentary mounted with TMM 31219-3046 has no visible specimen number. Handwritten notes state “left side of skull and left mandible”, collected “1.3 miles north of the court house, Cold Spring, San Jacinto Co., Texas” in 1934 by Claude Riley. The worn lower m3 incorporated into the dentary only includes a small amount of original tooth (lingual margin) and is mostly modeled in plaster. The occlusal pattern is of no value and the tooth appears to be attached to the dentary with plaster. The lower tusk, associated with the dentary, also appears to be set in plaster at the alveolus with no actual fit and therefore is of uncertain provenance. This tusk is somewhat oval in cross-section (38 mm x 32 mm) at the broken proximal end (Figure 24). The total length of the tusk fragment is ~280 mm and is relatively straight with no enamel band. It is significantly smaller than the lower tusk of Gomphotherium sp. from the Sam Houston locality (TMM 31191-2233) (53 mm x 47 mm cross-section diameters and 416 mm length). The latter appears less compressed in cross-section, but the significance of these differences is uncertain and could easily reflect ontogeny or sexual dimorphism. Neither specimen exhibits growth cones, suggesting the proximal ends are missing. The sub-oval cross-section of the mandibular tusks differs from the dorso-ventrally flattened lower tusks seen in the holotype of Gomphotherium productum from New Mexico (Morgan 2015).
The presence of cement between lophs on the M3 of TMM 38-3046 was noted as a distinctive and important character by Tobien (1973). However, the occlusal surface of this heavily worn and partially reconstructed tooth was coated in plaster for mounting in 1939. It is not possible to observe cement as described by Tobien (1973). The description of TMM 38-3046 in Tobien (1973) is misleading and the species of Gomphotherium represented by this specimen is unclear. A significant re-preparation effort might reveal characters that could be used for more confident identification. Based on the actual bone observable at present, all that can be said is that the specimen represents a small gomphothere, possibly with peg-like lower tusks.
Tobien (1973) incorrectly stated that “Hay’s specimens” (referring to “Gomphotherium cimarronis” from Noble Farm) “belong to the Coldspring fauna.” As described elsewhere in this paper, the Noble Farm L.F. is clearly from the lower part of the Lagarto Formation and is a Ba1 fauna while the Cold Spring L.F. is from the upper part of the Lagarto Formation and is a Ba2 fauna. Perhaps influenced by the statement in Tobien (1973), MacFadden et al. (2015) also stated that Gomphotherium cimarronis (Hay 1925) was “from the Cold Spring Fauna of the Texas Gulf Coastal Plain.”
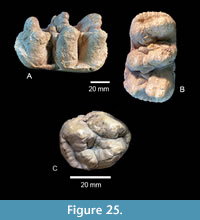 Additional specimens of proboscideans are known from the Cold Spring L.F. TMM 31219-44 is an unworn Lm1 with a maximum length of 83.0 mm and a maximum width of 49.8 mm (Figure 25A, B). There are three lophids, a posterior heel, and both anterior and pretrite cingula. This tooth would develop pretrite trefoils with wear. There is no cement on this tooth. TMM 31219-177 is an unworn premolar with a maximum length of 35.4 mm and width of 29.8 mm (Figure 25C). The tooth displays two primary lophids with a small anterior, posterior, and pretrite cingula. The morphology of this tooth compares favorably with other permanent premolars recognized in some gomphotheres.
Additional specimens of proboscideans are known from the Cold Spring L.F. TMM 31219-44 is an unworn Lm1 with a maximum length of 83.0 mm and a maximum width of 49.8 mm (Figure 25A, B). There are three lophids, a posterior heel, and both anterior and pretrite cingula. This tooth would develop pretrite trefoils with wear. There is no cement on this tooth. TMM 31219-177 is an unworn premolar with a maximum length of 35.4 mm and width of 29.8 mm (Figure 25C). The tooth displays two primary lophids with a small anterior, posterior, and pretrite cingula. The morphology of this tooth compares favorably with other permanent premolars recognized in some gomphotheres.
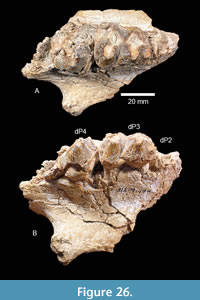 TMM 31219-144 is a partial left maxilla of a juvenile Gomphotherium sp. from the Cold Spring L.F. (Figure 26). The specimen includes partial roots of DP2, a nearly complete DP3, and a partial DP4. The DP3 is 37.6 mm in length and 31.6 mm in maximum width with a subtle pretrite trefoil pattern.
TMM 31219-144 is a partial left maxilla of a juvenile Gomphotherium sp. from the Cold Spring L.F. (Figure 26). The specimen includes partial roots of DP2, a nearly complete DP3, and a partial DP4. The DP3 is 37.6 mm in length and 31.6 mm in maximum width with a subtle pretrite trefoil pattern.
TMM 31219-237 (incorrectly cited by Tobien [1973, p. 218] as 21219-237) was described by Tobien as a partial skull with “deeply worn right M3, min length 116 mm.” This partial skull from the Cold Spring L.F. represents a mature individual (Tassy [2013] age class XXIII). The worn RM3 has enamel only around the posterior loph and heel. Alveoli are present for RM2 and for LM2-M3. The ventral portion of the skull is relatively complete but was locally restored with plaster. The palate and basicranium are well preserved, as are the occipital condyles (Figure 27).
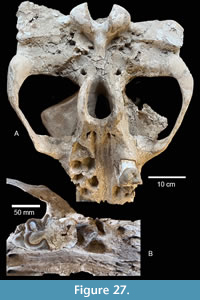 Dimensions, following Tassy (2013), are shown in Table 3. Both the pattern of variation and the absolute values of TMM 31219-237 are similar to SEP185 that Tassy (2013) measured for age class XXI of Gomphotherium angustidens. The palatine foramina are large and elongated antero-posteriorly. The occipital condyles are relatively rounded with a maximum width of 16.2 cm with an oval-shaped foramen magnum. The dorsal portion of the skull is missing, as are the tusks. The width of the skull, as well as the length and width of the basicranium, are significantly larger in Gomphotherium sp. from the Cold Spring L.F. (TMM 31219-237). The single worn M3 is of little use for identification. This specimen supports the observation that the gomphotheres from the Ba2 faunas in the upper Lagarto Formation were generally similar in size to Gomphotherium angustidens, but with a broader skull. The gomphothere from the Cold Spring L.F. was significantly smaller than the Early Clarendonian cf. Gomphotherium. sp. from the Lapara Creek Fauna (May, 2019).
Dimensions, following Tassy (2013), are shown in Table 3. Both the pattern of variation and the absolute values of TMM 31219-237 are similar to SEP185 that Tassy (2013) measured for age class XXI of Gomphotherium angustidens. The palatine foramina are large and elongated antero-posteriorly. The occipital condyles are relatively rounded with a maximum width of 16.2 cm with an oval-shaped foramen magnum. The dorsal portion of the skull is missing, as are the tusks. The width of the skull, as well as the length and width of the basicranium, are significantly larger in Gomphotherium sp. from the Cold Spring L.F. (TMM 31219-237). The single worn M3 is of little use for identification. This specimen supports the observation that the gomphotheres from the Ba2 faunas in the upper Lagarto Formation were generally similar in size to Gomphotherium angustidens, but with a broader skull. The gomphothere from the Cold Spring L.F. was significantly smaller than the Early Clarendonian cf. Gomphotherium. sp. from the Lapara Creek Fauna (May, 2019).
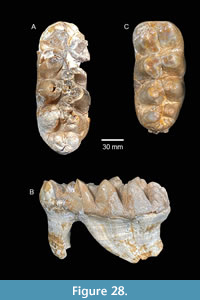 Additional specimens of Gomphotherium sp. are known from the Sam Houston locality within the Cold Spring L.F. (TMM 31191). TMM31191-1 is an Rm3 with four main lophids exhibiting pretrite trefoils. A well-developed posterior heel includes three main conulids and a small posterior conulid (Figure 28). There is no lingual cingulum, and the median sulcus is poorly developed. A lingual conulid is present between both the first and second lophids. The m3 is 152 mm in length. The first and second lophids are partially broken, but the minimum width at the first lophid is 65 mm. The widths at the third and fourth lophids are 65.7 mm and 60.4 mm, respectively. The crown height, on the lingual margin of the third and fourth lophids, is 42.7 mm and 38.6 mm, respectively. TMM 31191-1 is significantly larger than Gomphotherium sp. from Noble Farm (TMM 40775-46) and the fourth lophid, the posterior heel and accessory conulids are all better developed (Figure 28). These characters are all similar to Gomphotherium angustidens and TMM 31191-1 clearly represents a larger and more derived species of Gomphotherium in the Ba2 Cold Spring L.F. compared to the Ba1 Noble Farm L.F. Although similar in size and crown height, it differs from the holotype of Gomphotherium productum from Late Barstovian faunas of New Mexico, in having four lophids with a well-developed posterior heel.
Additional specimens of Gomphotherium sp. are known from the Sam Houston locality within the Cold Spring L.F. (TMM 31191). TMM31191-1 is an Rm3 with four main lophids exhibiting pretrite trefoils. A well-developed posterior heel includes three main conulids and a small posterior conulid (Figure 28). There is no lingual cingulum, and the median sulcus is poorly developed. A lingual conulid is present between both the first and second lophids. The m3 is 152 mm in length. The first and second lophids are partially broken, but the minimum width at the first lophid is 65 mm. The widths at the third and fourth lophids are 65.7 mm and 60.4 mm, respectively. The crown height, on the lingual margin of the third and fourth lophids, is 42.7 mm and 38.6 mm, respectively. TMM 31191-1 is significantly larger than Gomphotherium sp. from Noble Farm (TMM 40775-46) and the fourth lophid, the posterior heel and accessory conulids are all better developed (Figure 28). These characters are all similar to Gomphotherium angustidens and TMM 31191-1 clearly represents a larger and more derived species of Gomphotherium in the Ba2 Cold Spring L.F. compared to the Ba1 Noble Farm L.F. Although similar in size and crown height, it differs from the holotype of Gomphotherium productum from Late Barstovian faunas of New Mexico, in having four lophids with a well-developed posterior heel.
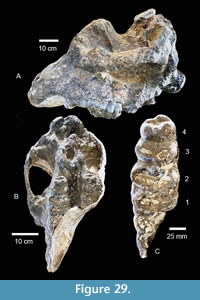 Chappell Hill Local Fauna. TMM 31272-42 is another partial skull of a relatively small gomphothere with both left and right M3 (Figure 29). This skull was collected by J. Quinn in 1953 from a roadcut on Hwy 290 near Chappell Hill, Texas, and is from the upper Lagarto Formation.
Chappell Hill Local Fauna. TMM 31272-42 is another partial skull of a relatively small gomphothere with both left and right M3 (Figure 29). This skull was collected by J. Quinn in 1953 from a roadcut on Hwy 290 near Chappell Hill, Texas, and is from the upper Lagarto Formation.
The left zygomatic arch on TMM 31272-42 is partially preserved, with some plaster restoration. The top of the skull is crushed, and the left anterior portion is missing. The left side of the skull is imbedded in a plaster cradle. The length of the preserved skull, from the occipital condyles to the anterior part of the premaxilla, is 68.5 cm and the length of the zygomatic arch is approximately 35 cm. The height of the preserved skull, from the frontals to the dorsal edge of the tooth row, is 34.5 cm. The width of the occipital condyles is approximately 160 mm, although they are not well preserved. The posterior portion of the skull is crushed and not well preserved. The worn LM3 exhibits simple pretrite trefoils, a lingual cingulum, is 147 mm in length, with widths of 75 mm at the first loph, 71 mm at the second, 64 mm at the third and 46 mm at the fourth. The fourth loph is relatively well developed with a small posterior heel. There is a small amount of cement in the lingual transverse valley between the second and third lophs. No cement is obvious on the RM3. The LM2 is extremely worn with only partial second and third lophs preserved. The LM3 is significantly longer than Gomphotherium sp. from the Noble Farm L.F. with a better developed fourth loph. It is not possible to measure crown height due to the worn nature of the tooth.
Trilophodon sp. was included by Hesse (1942) in his faunal list for the “Goodrich fauna” (TMM 31183) located south of Livingston in Polk County, Texas, and considered by Tedford et al. (2004) to be a late Ba2 fauna. The specimen was apparently collected approximately 0.8 km northeast of the main locality from near the top of the exposed Fleming Group (Hesse, 1942). Both localities are referred to as Smith Ranch in the TxVP collections, but the locality that included a proboscidean, according to Hesse (1942), is now labeled TMM 31200. Although 13 specimens are known from TMM 31200 (Hesse, 1942 noted 12), none are proboscidean.
Discussion. Ba2 local faunas, from the upper Lagarto Formation, include at least one additional species of Gomphotherium that shares characteristics with Gomphotherium angustidens and Gomphotherium productum. The small gomphotheres from Texas with “peg-like lower tusks” were interpreted by Tobien (1973) to be similar to Gomphotherium angustidens but chose to synonymize all of the “Texas material” as Gomphotherium productum. Whereas Gomphotherium productum has down-curved upper tusks with a straight enamel band, those of Gomphotherium angustidens have a spiral architecture (Tassy, 2014). The lack of upper tusks from the Ba2 gomphothere from the upper Lagarto Formation precludes comparison of this distinguishing character. Acknowledging both the limited number of specimens, and the need for a complete taxonomic revision of the Gomphotheriidae, the group of small gomphotheres from Ba2 faunas of the Texas Coastal Plain are here identified as Gomphotherium sp. Whether or not this clade of gomphotheres was derived from the more primitive Ba1 taxa in this region, or represents a separate immigration event from Eurasia, is unclear given the limited amount of material presently available.
Two proboscideans were described from the Early Clarendonian Lapara Creek Fauna in the Goliad Formation that overlies the Fleming Group (May, 2019). The Early Clarendonian Ten Mile Waterhole Creek and Bridge Ranch Local Faunas both contain material that May (2019) assigned to cf. Gomphotherium sp. The specimens include partial upper tusks with enamel bands, isolated cheek teeth, and a segment of a pyriform lower tusk. The cheek teeth include three third molars that range in size from 150-170 mm x 66-75 mm with four main lophs and a posterior heel (May, 2019). These teeth are generally longer and more hypsodont than Gomphotherium sp. from the Ba2 faunas of the upper Lagarto Formation described above. A second molar (TMM 30936-331) exhibits a relatively zygodont morphology (see fig. 70 of May, 2019). Although material is limited, the lower tusk and cheek teeth of cf. Gomphotherium sp. from Ten Mile Waterhole Creek and Bridge Ranch, are more derived than the Ba2 gomphotheres from the Lagarto Formation. This proboscidean is the only Miocene gomphotheriid from the Texas Coastal Plain with pyriform lower tusks. Gomphotherium is absent in the Cl2 Farrish Ranch and Buckner Ranch Local Faunas of the Goliad Formation where it is replaced by Blancotherium buckneri (May, 2019). Blancotherium buckneri is a longirostrine gomphothere that is especially abundant at the Buckner Ranch site TMM 30896. A large amount of both cranial and post-cranial material from this locality makes Blancotherium buckneri the best known gomphotheriid from the Miocene of the Texas Coastal Plain. Numerous additional field jackets, clearly representing a large proboscidean, are curated at TxVP awaiting preparation and almost certainly represent additional specimens of Blancotherium buckneri.
Blancotherium buckneri exhibits clear gomphotheriid affinity with respect to cheek tooth morphology, hyoid morphology, mandibular morphology including a long, shoe-horn shaped symphysis, and lower tusks (May, 2019). Unlike most other gomphotheres, Blancotherium buckneri has large, upwardly curving upper tusks that lack enamel bands. Although several mandibles from the Buckner Ranch locality possess alveoli for lower tusks, no lower tusks have been found within dentaries. Two, relatively small, isolated lower tusks with enamel bands were described by May (2019), but it appears that lower tusks were commonly shed from the mandible prior to maturity. Blancotherium represents a stage of longirostrine gomphotheriid evolution associated with the loss of lower tusks. This corresponds to the development of much larger, upward-curving, upper tusks that lacked enamel bands and show no consistent wear facets as seen in proboscideans from the Barstovian faunas. Ontogenetic loss of lower tusks was also described by Mothé et al. (2016) for the gomphotheriid Cuvieronius.
The phylogenetic relationship between Blancotherium and Gomphotherium is uncertain, although Blancotherium is clearly a more derived gomphotheriid with large, up-turned, upper tusks and apparent ontogenetic loss of lower tusks. Blancotherium is significantly different from all older gomphotheres of the Texas Coastal Plain and appears to be an immigrant that replaced Gomphotherium sp. during the Early Clarendonian. Blancotherium is currently only known from the Lapara Creek Fauna.
All Miocene and Pliocene proboscideans from the Texas panhandle have been referred to Rhynchotherium, Amebelodon, or Stegomastodon (e.g., Hemphillian: Higgins, Box T Local Faunas, Amebelodon (Schultz, 1990), Coffee Ranch, Rhynchotherium (?) sp. (Schultz, 1990); Blancan: Cita Canyon Local Fauna, Stegomastodon mirificus (Savage, 1955), Mt. Blanco Rhynchotherium praecursor, Stegomastodon mirificus (Dalquest, 1975), Red Corral S. mirificus (Schultz, 2016).
MIOCENE PROBOSCIDEAN RECORD FROM NORTH AMERICA
The oldest record of Proboscidea from North America is a partial tooth assigned to Zygolophodon by Morea (1981) from the Late Hemingfordian Massacre Lake L.F. in northern Nevada. The fossil record of Mammutidae in North America was recently reviewed by Von Koenigswald et al. (2023). Those authors assigned all Barstovian mammutid specimens to Zygolophodon proavus and reviewed the documented occurrences from seven different localities. The Ba1 faunas from the Deep River Formation, Montana; Virgin Valley, Nevada; Pawnee Creek, Colorado; and North Coalinga, California all contain proboscideans assigned to Zygolophodon proavus (Von Koenigswald et al., 2023). Zygolophodon was also reported from the Ba1 Shark Tooth Hill L.F. in California (Tedford et al., 2004; Prothero et al., 2008b).
A possible gomphothere was figured by Bode (1935) from the Ba1 North Coalinga locality in California. A single P4 shown in that publication was identified as a gomphothere by Tedford et al. (2004) and by Von Koenigswald et al. (2023). All known proboscidean fossils from the Barstow Formation in California were reviewed by Lofgren et al. (2012). Body fossils from the B2 upper member include both Zygolophodon and Gomphotherium, based on molar morphology. Although Zygolophodon proavus is represented by multiple isolated teeth, as well as a partial skull and dentary, Gomphotherium is represented by only two isolated teeth. Zygolophodon occurs near the base of the upper member while the first stratigraphic occurrence of Gomphotherium is slightly higher in the section (Pagnac, 2009). Given the limited number of specimens, the significance of this difference is unclear, but may suggest that mammutids and gomphotheres were not part of the same fauna at Barstow. Fossil trackways from the Calico Mountains, assigned to proboscidea, could be as old as Late Hemingfordian (Reynolds and Woodburne, 2001).
Zygolophodon and Gomphotherium were purported to co-occur in Ba1 faunas of the Mascall Formation in Oregon by Prothero et al. (2006). However, in his review of the Mascall fauna, Downs (1956) did not list any proboscidean occurrences. More recently, Maguire et al. (2018) reported fragmentary tooth material and a partial molar from the Mascall Formation that they suggested was comparable to Zygolophodon but left the identification as Proboscidea indet. They cited Fremd et al. (1994) and Prothero et al. (2006) as evidence for Gomphotherium from the Mascall Formation but stated they could find no evidence for this genus. Zygolophodon and Gomphotherium singulatum [sic] were included in faunal lists for the Mascall Formation by Fremd et al. (1994). This was presumably a mistaken spelling of Gomphotherium cingulatum that was described by Downs (1952). The specimen, designated as the holotype for Gomphotherium cingulatum, was recovered “about twenty-five years ago” according to Downs (1952). He described having been shown the original locality as “sparsely exposed beds in a stream channel surrounded by low, brush-covered hills.” In his description, Downs (1952) listed the fauna as “Indeterminate. Possibly Mascall” and stated that it was not possible to assign the specimen of Gomphotherium cingulatum to an established mammalian fauna because of the absence of associated material and the distance from known faunas. Tedford et al. (1987) explicitly commented on the lack of proboscidea material from the Mascall Fauna. Those authors also commented that the Rattlesnake Formation unconformably overlies the Mascall Formation and that early collectors commonly mixed specimens from the Hemphillian with the Barstovian units. An occurrence of this taxon from the Late Clarendonian Black Hawk Ranch fauna in California was noted by Tobien (1973). He further interpreted Gomphotherium cingulatum to be a junior synonym of Gomphotherium productum.
Mammutids are unknown, and gomphotheres only appear in Ba2 faunas in New Mexico (Morgan, 2015). Although originally described under numerous taxonomic names, most of the gomphothere specimens from New Mexico are now assigned to Gomphotherium productum following Tobien (1973). However, Heckert et al. (2000) concluded that the extreme degree of lumping, proposed by Tobien (1973) for species of Gomphotherium from New Mexico, was inconsistent with the range of variation observed.
Gomphotherium sp. has been reported from several faunas in Mexico, including the El Gramal Local Fauna from the Nejapa area, Oaxaca (Ferrusquia-Villafranca, 1990). The Mexican faunas with Gomphotherium in the Nejapa area were interpreted as late Ba1 faunas by Tedford et al. (2004). Fossil mammals from the Ixtapa Formation in Chiapas also include Gomphotherium sp. (Ferrusquia-Villafranca, 1992, 1996). Association with tuffs dated at 15.02 +/- 0.35 and 16.02 +/- 0.53 Ma indicate an Early Barstovian, or possibly Late Hemingfordian, age according to the Raffi et al. (2020) time scale. Gomphotherium sp. from the Ixtapa fauna is likely as old as the North Coalinga L.F. (California) and possibly as old as Massacre Lake (Nevada).
Although generally not common, fossil proboscideans are known from multiple Ba1 localities in North and Central America. The recognition of Gomphotherium from Ba1 localities in the Texas Coastal Plain is consistent with this pattern. The absence of the mammutid Zygolophodon in these faunas is possibly an artifact of small sample size, although faunal provinciality, perhaps associated with environmental partitioning, may also have been a factor. Zygolophodon is also absent from Barstovian faunas from New Mexico and only a single worn tooth of uncertain, but possible mammutid affinity, is known from the Late Barstovian of Florida (Morgan, 2015; Hulbert, 2001).
SUMMARY
A single partial vertebra may record the earliest known proboscidean in the Texas Coastal Plain. The record becomes more robust with gomphotheres present in three Ba1 local faunas in the lower to middle Lagarto Formation. Two different species of Gomphotherium are represented and one of these is similar to a taxon known from China in Hemingfordian-equivalent faunas. At least one additional gomphothere appears in the Ba2 local faunas of the upper Lagarto Formation with characters similar to Gomphotherium angustidens and Gomphotherium productum. A more derived form of Gomphotherium is known from the Cl1 local faunas of the Goliad Formation (Lapara Creek Fauna) and is identified as cf. Gomphotherium sp. with pyriform lower tusks. In Cl2 localities of the Lapara Creek Fauna, Gomphotherium is replaced by Blancotherium buckneri. There are no mammutids known from Barstovian faunas in Texas.
The diversity of gomphotheres in the Texas Coastal Plain seems to change little during the Barstovian and Clarendonian, with only one or two taxa present at any given time. This is in stark contrast to groups like equids that undergo a dramatic increase in diversity during this same time interval. The Ba1 Belts Creek L.F. contains a gomphothere similar to Gomphotherium inopinatum from China. Certain characters suggest the specimen from Belts Creek is slightly more derived than the Chinese material consistent with the Ba1 interpretation. The Ba1 Noble Farm and Brenham Local Faunas contain a gomphothere that shares characters with Gomphotherium subtapiroideum and Gomphotherium productum. The previously applied name of Gomphotherium cimarronis is considered nomen dubium and should not be used for the gomphothere from Noble Farm. Late Barstovian gomphotheres from the Cold Spring, Goodrich, and Chappell Hill Local Faunas share characters with Gomphotherium angustidens and Gomphotherium productum but appear distinct in detail from those taxa. The lack of upper tusk specimens from the Ba2 gomphothere hinders specific assignment. A gomphothere assigned to cf. Gomphotherium sp., from Cl1 local faunas, is of uncertain specific affinity and is replaced by Blancotherium buckneri in Cl2 local faunas of the Lapara Creek Fauna (May, 2019). Measured by the number of specimens, Blancotherium buckneri is the best known gomphothere from the Miocene of the Texas Coastal Plain. Additional material of this taxon is currently being analyzed.
The concept of a North American ‘Proboscidean Datum’ has been widely discussed in the literature (Tedford et al., 1987, 2004; Woodburne and Swisher, 1995; Prothero et al., 2008a; Prothero and Dold, 2008; Pagnac, 2009). Historically, the first temporal occurrence of proboscideans in North America was an “immigration event” in the Late Barstovian (Tedford et al., 1987). Later, Woodburne and Swisher (1995) discussed the ‘Proboscidean Datum’ and recognized one dispersal event in the Late Hemingfordian followed by a “major lag in proboscidean dispersal” until gomphotheres appear in Ba2 faunas in the interior of North America. They stated that the ‘Proboscidean Datum’ was “subject to the vagaries of biofacies and should be used with extreme caution.” The Massacre Lake L.F. was cited by them as the earliest known record of proboscideans in North America referencing Morea (1981) for a Late Hemingfordian age and for a radiometric calibration “at about 16.2 Ma.” Reversed magnetic polarity was reported by Prothero et al. (2008a) for the stratigraphic interval from which the vertebrate fossils had been collected. They interpreted this as Chron C5Cr which is 17.157-16.637 Ma based on Gradstein et al. (2020). Given the lack of proboscideans from all other He2 faunas in North America, the relatively interior location of Massacre Lake, and the limited nature of the material from that locality, the record should be treated with caution. The earliest confident occurrences of proboscideans from the West Coast, the Gulf Coast, and the Rocky Mountain regions of North America, are otherwise in Early Barstovian faunas.
Proboscideans are immigrants to the western hemisphere, first arriving in the Miocene, and spreading across North, Central, and South America (Alberdi et al., 2011). It is difficult to refute the hypothesis that Zygolophodon and Gomphotherium arrived at the same time in North America because they both appear with certainty in Ba1 faunas. However, there are few faunas of that age in North America that include both taxa (perhaps only North Coalinga, California, and that record includes only isolated teeth). Whereas Early Barstovian occurrences of Zygolophodon are primarily found along the West Coast and Rocky Mountain areas of North America, Gomphotherium appears to be more common in the Southwest, Gulf Coast, and Mexico. This may suggest some form of provinciality, even though these taxa presumably both immigrated from Eurasia through Beringia at approximately the same time. Alternatively, apparent provinciality may be the result of a limited fossil record. The lack of proboscideans from the well-known Ba1 faunas of the Great Plains remains an enigmatic aspect of this dispersal pattern.
Low stands of global sea level were found by Alberdi et al. (2011) to be an important component of transcontinental proboscidean dispersal. The most significant low-stand sea-level events during the Miocene Climate Optimum were the Mi2 at 16.0 Ma (~40 m), the Mi3a at 14.8 Ma (~30 m), and the Mi3 at 13.8 Ma (~50 m) (Miller et al., 2020). The Mi2 event correlates temporally with the base of the Early Barstovian and the first appearance of gomphotheres and other immigrant taxa in the Texas Coastal Plain. The Mi3a is near the base of the Late Barstovian where more advanced gomphotheres, similar to Gomphotherium angustidens, are first seen in the upper Lagarto Formation. Although most paleogeographic maps show a Beringian land connection between Asia and North America during the mid-Miocene, this connection was likely modulated by sea level variation of +/- 50 m, as indicated by the sea level curve of Miller et al. (2020).
Proboscideans from Early Barstovian faunas in North America, including both mammutids and gomphotheres, have long been known. Nevertheless, the concept of a Late Barstovian ‘Proboscidean Datum’ has persisted in literature. Summarizing their review of the hypothesis, Prothero et al. (2008a) concluded “Thus, the validity of the Proboscidean Datum has become less well supported as new data emerge.” The demonstrably diachronous nature of first stratigraphic occurrences in North America led Pagnac (2009) to suggest that proboscidea (both Zygolophodon and Gomphotherium) should be abandoned as defining taxa for both Ba1 and Ba2 faunas. It now seems clear that the previous hypothesis of a ‘Proboscidean Datum’ should not be exclusively used to interpret ages of vertebrate faunas in North America. The concept of a ‘Proboscidean Datum’ has misled age interpretations for Barstovian faunas in the Texas Coastal Plain and continues to influence interpretations elsewhere.
The revised biochronology of proboscidean occurrences from Barstovian strata of the Texas Coastal Plain discussed in this paper improves our understanding of global biogeography and dispersal patterns for these mammals. Further improvements will require collection of additional fossils, improved geochronology, and taxonomic revision of the Gomphotheriidae. Faunal provinciality may be reflected in the pattern of Miocene proboscidean occurrences with potential implications for both temporal correlation of faunas and interpretations of ecological partitioning. Taxonomic nomenclature, stratigraphic terminology, and temporal correlations in the Miocene strata of the Texas Coastal Plain are topics with complicated histories and careful attention to detail is required to progress our understanding of faunal dynamics.
ACKNOWLEDGMENTS
C. Sagebiel and M. Brown provided valuable assistance with the Texas Vertebrate Paleontology Collections. D. Wagner organized gomphothere material from the Noble Farm Local Fauna. J. Maisano, and D. Edey provided essential assistance at The University of Texas at Austin High-Resolution X-ray CT Facility. Reviews by C. Bell, M. May, D. Pagnac, and M. Pasenko significantly improved the manuscript.
REFERENCES
Albright, L.B. 1998. The Arikareean Land Mammal Age in Texas and Florida: southern extension of Great Plains faunas and Gulf Coastal Plain endemism. Geological Society of America Special Paper, 325:167-183.
https://doi.org/10.1130/0-8137-2325-6.167
Barnes, V.E., Hartmann, B.M., and Scranton, D.F. 1992. Geologic Map of Texas. The University of Texas at Austin, Bureau of Economic Geology.
Bode, F.D. 1935. The fauna of the Merychippus Zone, North Coalinga district, California. Carnegie Institution of Washington Publication. Contributions to Palaeontology. 453:66-96.
Brown, M.A. 2013. The development of “modern” laboratory methods: a century of progress. Earth and Environmental Science Transactions of the Royal Society of Edinburgh.
https://doi.org/10.1017/S1755691013000352
Cope, E.D. 1893. A preliminary report of the vertebrate paleontology of the Llano Estacado. Fourth Annual Report of the Geological Survey of Texas, 1892. 137 pp.
Dalquest, W.D. 1975. Vertebrate fossils from the Blanco Local Fauna of Texas. Occasional Papers The Museum Texas Tech University, 30:1-52. https://doi.org/10.5962/bhl.title.142927
Downs, T. 1952. A new mastodont from the Miocene of Oregon with remarks on Gomphotherium. University of California Publications in Geological Sciences, 29:1-20.
Downs, T. 1956. The Mascall fauna from the Miocene of Oregon. University of California Publications in Geological Sciences, 31:199-354.
Dreussen, A. 1924. Geology of the Coastal Plain of Texas west of Brazos River. U.S. Geological Survey Professional Paper, 126. 139 pp.
https://doi.org/10.3133/pp126
Emry, R.E. and Eshelman, R.E. 1998. The early Hemingfordian (early Miocene) Pollack Farm Local Fauna: first Tertiary land mammals described from Delaware. In Benson, R.N. (ed.) Geology and paleontology of the lower Miocene Pollack Farm fossil site, Delaware. Delaware Geological Survey Special Publication, 21:153-173.
Ferrusquia-Villafranca, I. 1990. Biostratigraphy of the Mexican continental Miocene: Part I, Introduction and the northwestern and central faunas; Part II, the southeastern (Oaxacan) faunas; Part III, the southeasternmost (Chiapasan) fauna and concluding remarks on the discussed vertebrate record. Universidad Nacional Autonoma de Mexico, Instituto de Geologia, Paleontologia Mexicana, 56:1-149.
Ferrusquia-Villafranca, I. 1992. Contribución al conocimiento del Cenozoico en al Sureste de Mexico y de su relevancia en el entendimiento de la evolutión geológica regional. VII Congresso Geológico Latinoamericano, Salamanca, Spain. 4:40-44.
Ferrusquia-Villafranca, I. 1996. Contribución al conocimiento geológico de Chiapas: El Area Ixtapa-Soyaló. Universidad Nacional Autonoma Mexico, Instituto de Geológia, Bulletin, 109:1-130.
Forsten, A.M. 1975. The fossil horses of the Texas Gulf Coastal Plain: a revision. Texas Memorial Museum Pearce-Sellards Series, 22:1-86.
Fremd, T., Bestland, E.A., and Retallack, G.J. 1994. John Day Basin paleontology field trip guide and road log. 1994 Society of Vertebrate Paleontology Annual Meeting, 80 pp.
Galloway, W.E., Jirik, L.A., Morton, R.A., and DuBar, J.R. 1986. Lower Miocene (Fleming) depositional episode of the Texas Coastal Plain and continental shelf: structural framework, facies, and hydrocarbon resources. Bureau of Economic Geology, Report of Investigations, 150, 50 pp.
https://doi.org/10.23867/ri0150d
Gheerbrant, E. 2009. Paleocene emergence of elephant relatives and the rapid radiation of African ungulates. Proceedings of the National Academy of Sciences, PNAS 106, 26:10717-10721.
https://doi.org/10.1073/pnas.0900251106
Gheerbrant, E. and Tassy, P. 2009. L’origine et l’évolution des éléphants, Comptes Rendus Palevol, 8:281-294.
https://doi.org/10.1016/j.crpv.2008.08.003
Gradstein, F.M., Ogg, J.G., Schmitz, M., and Ogg, G. 2020. The Geologic Time Scale 2020, Elsevier, Oxford.
Hay, O.P. 1922. Further observations on some extinct elephants. Proceedings of the Biological Society of Washington, 35:97-101.
Hay, O.P. 1924. Description of some fossil vertebrates from the Upper Miocene of Texas. Proceedings of the Biological Society of Washington, 37:1-20.
Hay, O.P. 1925. On remains of mastodons found in Texas, Anancus brazosius and Gomphotherium cimarronis. Proceedings of the U.S. National Museum, 66:1-20.
Hesse, C.J. 1942. A preliminary report on the Miocene vertebrate faunas of Southeast Texas. Proceedings and Transactions of The Texas Academy of Science, 1941, 25:157-179.
Hulbert, R.C. 1988. Calippus and Protohippus (Mammalia, Perissodactyla, Equidae) from the Miocene (Barstovian-early Hemphillian) of the Gulf Coastal Plain. Bulletin of the Florida State Museum, Biological Sciences, 32:221-340.
https://doi.org/10.58782/flmnh.avxe6832
Hulbert, R.C. (ed.) 2001. The Fossil Vertebrates of Florida. Florida Paleontological Society. University Press of Florida. 350 pp.
Hunt, R.M., Jr. 1998. Amphicyonidae, p. 196-227. In Janis, C., Scott, K., and Jacobs, L. (eds.), Evolution of Tertiary mammals of North America. Volume 1: Terrestrial carnivores, ungulates and ungulate-like mammals. Cambridge University Press, Cambridge, UK.
Lambert, W.D. 1996. The biogeography of the gomphotheriid proboscideans of North America, p. 143-148. In Shoshani, J. and Tassy, P. (eds.), The Proboscidea Evolution and Palaeoecology of Elephants and Their Relatives. Oxford University Press.
Lambert, W.D. and Shoshani, J. 1998. Proboscidea, p. 606-621. In Janis, C.M., Scott, K.M., and Jacobs, L.L., (eds.), Evolution of Tertiary Mammals of North America. Volume 1: Terrestrial carnivores, ungulates and ungulate-like mammals. Cambridge University Press, Cambridge, UK.
Lofgren, D.L., Hess, A., Silver, D., and Liskanich, P. 2012. Review of proboscideans from the middle Miocene Barstow Formation of California. In Reynolds, R.E. (ed.), Searching for the Pliocene: southern exposures. California State University Desert Studies Center, The 2012 Desert Research Symposium, p. 125-134.
MacFadden, B.J. 1984. Systematics and phylogeny of Hipparion, Neohipparion, Nannippus, and Cormohipparion (Mammalia, Equidae) from the Miocene and Pliocene of the New World. Bulletin of the American Museum of Natural History, 179:1-196.
MacFadden, B.J. and Skinner, M.F. 1981. Earliest Holarctic Hipparion, Cormohipparion goorsi n. sp. (Mammalia, Equidae), from the Barstovian (medial Miocene) Texas Gulf Coastal Plain. Journal of Paleontology, 55:619-627.
MacFadden, B.J., Morgan, G.S., Jones, D.S., and Rincon, A.F. 2015. Gomphothere proboscidean (Gomphotherium) from the late Neogene of Panama. Journal of Paleontology, 89:360-365.
https://doi.org/10.1017/jpa.2014.31
Madden, C.T. and Storer, J.E. 1985. The Proboscidea from the middle Miocene Wood Mountain Formation, Saskatchewan. Canadian Journal of Earth Sciences, 22:1345-1350.
https://doi.org/10.1139/e85-138
Maguire, K.C., Samuels, J.X., and Schmitz, M.D. 2018. The fauna and chronostratigraphy of the middle Miocene Mascall type area, John Day Basin, Oregon, USA, PaleoBios, 35:1-51.
https://doi.org/10.5070/p9351037578
May, S.R. 2019. The Lapara Creek Fauna: Early Clarendonian of south Texas, USA. Palaeontologia Electronica 22.1.15A: 1-129.
https://doi.org/10.26879/929
May, S.R. and Brown, M.A. 2023. Anchitheriomys buceei (Rodentia, Castoridae) from the Miocene of Texas and a review of the Miocene beavers from the Texas Coastal Plain, USA. Palaeontologia Electronica, 26(1): a7: 1-27.
https://doi.org/10.26879/1236
McAnulty, W.N. 1939. The Laboratory Unit. In Evans, G. (ed.), Manual of suggested procedures to be employed by the State-Wide Paleontologic-Mineralogic Survey in Texas, 73-91. The University of Texas at Austin, Bureau of Economic Geology.
Miller, K.G., Browning, J.V., Schmelz, W.J., Kopp, R.E., Mountain, G.S., and Wright, J.D. 2020. Cenozoic sea-level and cryospheric evolution from deep-sea geochemical and continental margin records, Sciences Advances, 6, eaaz1346.
Morea, M.F. 1981. The Massacre Lake Local Fauna (Mammalia, Hemingfordian) From Northwestern Washoe County, Nevada. Unpublished PhD Dissertation, University of California, Riverside, USA. 247 pp.
Morgan, G.S. 2015. Oligocene and Miocene vertebrates of New Mexico, p. 159-231. In Lucas, S.G. and Sullivan, R.M. (eds.), Fossil Vertebrates in New Mexico. New Mexico Museum of Natural History and Science Bulletin, 68.
Morton, R.A., Jirik, L.A., and Galloway, W.E. 1988. Middle-Upper Miocene depositional sequences of the Texas Coastal Plain and continental shelf: geologic framework, sedimentary facies, and hydrocarbon plays. The University of Texas at Austin, Bureau of Economic Geology, Report of Investigations, 174, 40 pp.
https://doi.org/10.23867/ri0174d
Mothé, D., Ferretti, M., and Avilla, L. 2016. The dance of tusks: rediscovery of lower incisors in the Pan-American proboscidean Cuvieronius hyodon revises incisor evolution in Elephantimorpha. PLoS ONE, 11(1): e0147009.
https://doi.org/10.1371/journal.pone.0147009
Olsen, S.J. 1958. The fossil carnivore Amphicyon intermedius from the Thomas Farm Miocene, Part I: Skull and dentition. Bulletin of the Museum of Comparative Zoology, 118:157-172.
Pagnac, D. 2009. Revised large mammal biostratigraphy and biochronology of the Barstow Formation (Middle Miocene), California. PaleoBios, 29:48-59.
https://doi.org/10.5070/p9292021802
Patton, T.H. 1969. Miocene and Pliocene artiodactyls, Texas Gulf Coastal Plain. Bulletin Florida State Museum Biological Sciences, 14:115-226.
Patton, T.H. and Taylor, B.E. 1971. The Synthetoceratinae (Mammalia, Tylopoda, Protocertaidae). Bulletin of the American Museum of Natural History, 145:119-218.
Patton, T.H. and Taylor, B.E. 1973. The Protoceratinae (Mammalia, Tylopoda, Protoceratidae) and the systematics of the Protoceratidae. Bulletin of the American Museum of Natural History, 150:347-414.
Plummer, F.B. 1932. Cenozoic Systems in Texas. In Sellards, E.H., Adkins, W.S., and Plummer, F.B. (eds.), The Geology of Texas, Volume 1, Stratigraphy. The University of Texas at Austin, Bureau of Economic Geology Bulletin, 3232:519-818.
Prothero, D.R. and Manning, E.M. 1987. Miocene rhinoceroses from the Texas Gulf Coastal Plain. Journal of Paleontology, 61:388-423.
https://doi.org/10.1017/S0022336000028559
Prothero, D.R., Draus, E., and Foss, S. 2006. Magnetic stratigraphy of the lower portion of the middle Miocene Mascall Formation, central Oregon. PaleoBios 26(1):27-32.
Prothero, D.R. and Dold, P. 2008. Magnetic stratigraphy of the Hemingfordian-Barstovian (lower to middle Miocene) Martin Canyon and Pawnee Creek Formations, northwestern Colorado, and the age of the ‘Proboscidean Datum’ in the High Plains, p. 247-254. In Lucas, S.G., Morgan, G.S., Spielmann, J.A., and Prothero, D.R. (eds.), Neogene Mammals. New Mexico Museum of Natural History and Science Bulletin 44.
Prothero, D.R., Davis, E.B., and Hopkins, S.S, 2008a. Magnetic stratigraphy of the Massacre Lake Beds (Late Hemingfordian, Early Miocene), northwest Nevada, and the age of the ‘Proboscidean Datum’ in North America, p. 239-246. In Lucas, S.G., Morgan, G.S., Spielmann, J.A., and Prothero, D.R. (eds.), Neogene Mammals. New Mexico Museum of Natural History and Science Bulletin 44.
Prothero, D.R., Liter, M.R., Barnes, L.G., Wang, X., Mitchell, E., McLeod, S., Whistler, D.P., Tedford, R.H., and Ray, C. 2008b. Land mammals from the middle Miocene Sharktooth Hill bonebed, Kern County, California, p. 299-314. In Lucas, S.G., Morgan, G.S., Spielmann, J.A., and Prothero, D.R. (eds.), Neogene Mammals. New Mexico Museum of Natural History and Science Bulletin 44.
Quinn, J.H. 1955. Miocene Equidae of the Texas Gulf Coastal Plain. The University of Texas at Austin, Bureau of Economic Geology, Pub. No. 5516:1-102.
Raffi, I., Wade, B.S., and Pälike, H. 2020. The Neogene Period. In Gradstein, F.M., Ogg, J.G.,
Schmitz, M., and Ogg, G. (eds.) The Geologic Time Scale 2020, Elsevier, Oxford.
https://doi.org/10.1016/B978-0-12-824360-2.00029-2
Reinhart, R.H. 1976. Fossil sirenians and desmostylids from Florida and elsewhere. Bulletin of the Florida State Museum Biological Sciences, 20:187-300.
https://doi.org/10.58782/flmnh.bywr8066
Renick, B.C. 1936. The Jackson Group and the Catahoula and Oakville Formations in a part of the Texas Gulf coastal plain. The University of Texas at Austin, Bureau of Economic Geology, Bulletin, 3619, 104 pp.
Reynolds, R.E. and Woodburne, M.O. 2001. Review of the Proboscidean Datum within the Barstow Formation, Mojave Desert, California. Journal of Vertebrate Paleontology, Abstracts, 21:93A.
Sanders, W.J. and Miller, E.R. 2002. New proboscideans from the early Miocene of Wadi Moghara, Egypt. Journal of Vertebrate Paleontology, 22:388-404.
https://doi.org/10.1671/0272-4634(2002)022[0388:npftem]2.0.co;2
Sanders, W.J. 2024. Evolution and fossil record of African Proboscidea. CRC Press, Taylor and Francis Group, LLC. 346 pp. doi:10.1201/b20016 (Note: this volume was received in October 2023)
Savage, D.E. 1955. Nonmarine lower Pliocene sediments in California: A geochronologic-stratigraphic classification. University of California Publications in Geological Sciences, 31:1-26.
Schlesinger, G. 1917. Die Mastodonten des K. K. Naturhistorischen Hofmuseums. Denkschriften des K. K. Naturhistorischen Hofmuseums, Geologisch-paläontologische Reihe, 1:1-231.
Schultz, G.E. 1990. Clarendonian and Hemphillian vertebrate faunas from the Ogallala Formation (late Miocene-early Pliocene) of the Texas Panhandle and adjacent Oklahoma, p. 56-97. In Gustavson, T.C. (ed.), Geologic Framework and Regional Hydrology: Upper Cenozoic Blackwater Draw and Ogallala Formations, Great Plains. The University of Texas at Austin, Bureau of Economic Geology.
Schultz, G.E. 2016. The Red Corral (Proctor Ranch) Local Fauna (Pliocene, Blancan) of Oldham County, Texas. New Mexico Museum of Natural History and Science, 73:1-62.
Shoshani, J. 1996. Para- or monophyly of the gomphotheres and their position within Proboscidea, p. 149-177. In Shoshani, J. and Tassy, P. (eds.), The Proboscidea Evolution and Palaeoecology of Elephants and Their Relatives. Oxford University Press.
Stenzel, H.B., Turner, F.E., and Hesse, C.J. 1944. Brackish and non-marine Miocene in southeastern Texas. Bulletin of the American Association of Petroleum Geologists, 28:977-1011.
https://doi.org/10.1306/3d933696-16b1-11d7-8645000102c1865d
Swanson, S.M. and Karlsen, W. 2009. USGS Assessment of Undiscovered Oil and Gas Resources for the Oligocene Frio and Anahuac Formations, Onshore Gulf of Mexico Basin, USA. Search and Discovery Article #10178, 44 pp.
Tassy, P. 1985. La Place des Mastodontes Miocènes de l’Ancien Monde dans la Phylogénie des Proboscidea (Mammalia): Hypothèses et Conjectures. Thèse Doctoratès Sciences. Paris: UPMC. 861 pp.
Tassy, P. 1996a. Dental homologies and nomenclature in the Proboscidea. In Shoshani, J. and Tassy, P. (eds.), p. 21-25. The Proboscidea Evolution and Palaeoecology of Elephants and Their Relatives. Oxford University Press.
Tassy, P. 1996b. Who is who among the Proboscidea? In Shoshani, J. and Tassy, P. (eds.), p. 39-48. The Proboscidea Evolution and Palaeoecology of Elephants and Their Relatives. Oxford University Press.
Tassy, P. 2013. L’anatomie cranio-mandibulaire de Gomphotherium angustidens (Cuvier,1817) (Proboscidea, Mammalia): données issues du gisement d’En Péjouan (Miocène moyen du Gers, France). Geodiversitas 35:377-445.
https://doi.org/10.5252/g2013n2a6
Tassy, P. 2014. L’odontologie de Gomphotherium angustidens (Cuvier, 1817) (Proboscidea, Mammalia): données issues du gisement d’En Péjouan (Miocène moyen du Gers, France). Geodiversitas 36:35-115.
https://doi.org/10.5252/g2014n1a2
Tedford, R.H., Galusha, T., Skinner, M.F., Taylor, B.E., Fields, R.W., Macdonald, J.R., Rensberger, J.M., Webb, S.D., and Whistler, D.P. 1987. Faunal succession and biochronology of the Arikareean through Hemphillian interval (late Oligocene through earliest Pliocene epochs) in North America, p. 153-210. In Woodburne, M.O. (ed.), Cenozoic Mammals of North America, Geochronology and Biostratigraphy. University of California Press, Berkeley and Los Angeles, California.
Tedford, R.H., Albright, L.B., Barnosky, A.D., Ferrusquia-Villafranca, I., Hunt, R.M., Storer, J.E., Swisher, C.C., Voorhies, M.R., Webb, S.D., and Whistler, D.P. 2004. Mammalian biochronology of the Arikareean through Hemphillian interval (late Oligocene through early Pliocene epochs). p. 169-231. In Woodburne, M.O. (ed.), Late Cretaceous and Cenozoic Mammals of North America: Biostratigraphy and Geochronology. Columbia University Press, New York.
https://doi.org/10.7312/wood13040-008
Tobien, H. 1973. On the evolution of the mastodonts (Probosciden, Mammalia) Part 1: The bunodont trilophodont groups. Notizblatt des Hessichen Landesamtes für Bodenforschung zu Weisbaden, 101:202-276.
Van der Hoof, V.L. and Gregory, J.T. 1940. A review of the genus Aelurodon. University of California Publications in Geological Sciences, 25:143-164.
Von Koenigswald, W., Widga, C., and Göhlich, U.B. 2023. New mammutids (Proboscidea) from the Clarendonian and Hemphillian of Oregon - a survey of Mio-Pliocene mammutids from North America. The Bulletin of the Museum of Natural History of the University of Oregon. Museum Bulletin No. 30, 63 pp.
Wang, X., Tedford, R.H., and Taylor, B.E. 1999. Phylogenetic systematics of the Borophaginae (Carnivora: Canidae). Bulletin of the American Museum of Natural History, 243:1-391.
Wang, S. 2014. Gomphotherium inopinatum, a basal Gomphotherium species from the Linxia Basin, China, and other Chinese members of the genus. Vertebrata PalAsiatica, 52:183-200.
Wang, S., Li, Y., Duangkrayom, J., Yang, X., He, W., and Chen, S. 2017. A new species of Gomphotherium (Proboscidea, Mammalia) from China and the evolution of Gomphotherium in Eurasia. Journal of Vertebrate Paleontology, 37, e1318284, 15 p.
https://doi.org/10.1080/02724634.2017.1318284
Wilson, J.A. 1956. Miocene formations and vertebrate biostratigraphic units, Texas coastal plain. Bulletin of the American Association of Petroleum Geologists, 40:2233-2246.
https://doi.org/10.1306/5ceae571-16bb-11d7-8645000102c1865d
Wilson, J.A. 1957. Early Miocene Enteledonts, Texas Coastal Plain. American Journal of Science, 255:641-649.
https://doi.org/10.2475/ajs.255.9.641
Wilson, J.A. 1960. Miocene carnivores, Texas coastal plain. Journal of Paleontology, 34:983-1000.
Wilson, J.A. 1962. Tertiary formations between Austin and Houston with special emphasis on the Miocene and Pliocene. Geology of the Gulf Coast and Central Texas, and Guidebook of Excursions. Field Excursion No. 10:342-353.
Woodburne, M.O. (ed.) 2004. Late Cretaceous and Cenozoic Mammals of North America: Biostratigraphy and Geochronology. Columbia University Press, New York.
Woodburne, M.O. and Swisher, C.C. 1995. Land mammal high-resolution geochronology, intercontinental overland dispersals, seal level, climate and vicariance, p. 335-364. In Berggren, W.A., Kent, D.V., Aubry, M., and Hardenbol, J. (eds.), Geochronology, time-scales, and global stratigraphic correlation. Society for Sedimentary Geology Special Publications, 54.
Young, S.C., Budge, T., Knox, P.R., Kalbouss, R., Baker, E., Hamlin, S., Galloway, W.E., and Deeds, N. 2010. Hydrostratigraphy of the Gulf Coast Aquifer from the Brazos River to the Rio Grande. Austin, Texas Water Development Board Report, 203 pp.
