

 The first fossil bird from the Miocene swamps of Gračanica, Bosnia and Herzegovina: A novel and very unique duck
The first fossil bird from the Miocene swamps of Gračanica, Bosnia and Herzegovina: A novel and very unique duck
Article number: 28.1.a14
https://doi.org/10.26879/1512
Copyright Paleontological Society, March 2025
Author biographies
Plain-language and multi-lingual abstracts
PDF version
Appendix
Submission: 7 November 2024. Acceptance: 12 February 2025.
ABSTRACT
The authors describe a nearly complete skeleton of a new genus and species of duck from the Middle Miocene, preserved on a slab. The skull, mandible, sternum and pelvis are also preserved, making this specimen even more unique. This is the first fossil duck with documented gastroliths. The gastroliths appear as a cluster of several dozen polished stones in a place that anatomically corresponds to the approximate location of the gizzard in ducks. This is also the first fossil bird from the Gračanica site, which has long been known for numerous early Middle Miocene fossils, ranging from large mammals and mollusks, and more recently also for fish, amphibians and reptiles, as well as arthropods, insects, sponges, protists, macroflora and plant pollen. The specimen displays a mosaic of primitive and advanced characters that are not exclusive to one tribe of ducks, therefore, apart from excluding Anatini, its systematic affiliation cannot be precisely indicated. However, we have sufficient reason to believe that at least in the ecological sense it was a diving duck.
Zbigniew M. Bochenski. Institute of Systematics and Evolution of Animals, Polish Academy of Sciences, Slawkowska 17, 31-016 Krakow, Poland. (corresponding author). bochenski@isez.pan.krakow.pl
https://orcid.org/0000-0002-9405-3933
Johannes Happ. Reptilienzoo Happ, Villacherstrasse 237, 9020 Klagenfurt, Austria. johanneshapp@yahoo.de
https://orcid.org/0009-0007-3206-4315
Grzegorz Salwa. Regional Subcarpathian John Paul II Hospital in Krosno, Department of Neurology, Korczyńska 57, 38-400 Krosno, Poland. grzegorz.tomasz.salwa@gmail.com
https://orcid.org/0000-0003-0589-4675
Teresa Tomek. Institute of Systematics and Evolution of Animals, Polish Academy of Sciences, Slawkowska 17, 31-016 Krakow, Poland. toteresa45@gmail.com
https://orcid.org/0000-0002-8278-3031
Keywords: new species; new genus; Miocene; Anatidae; gastroliths; Gračanica
Final citation: Bochenski, Zbigniew M., Happ, Johannes, Salwa, Grzegorz, and Tomek, Teresa. 2025. The first fossil bird from the Miocene swamps of Gračanica, Bosnia and Herzegovina: A novel and very unique duck. Palaeontologia Electronica, 28(1):a14.
https://doi.org/10.26879/1512
palaeo-electronica.org/content/2025/5476-first-miocene-duck-from-bosnia-and-herzegovina
Copyright: March 2025 Paleontological Society,
This is an open access article distributed under the terms of Attribution-NonCommercial-ShareAlike 4.0 International (CC BY-NC-SA 4.0), which permits users to copy and redistribute the material in any medium or format, provided it is not used for commercial purposes and the original author and source are credited, with indications if any changes are made.
creativecommons.org/licenses/by-nc-sa/4.0/
https://zoobank.org/E098A11E-E715-4A26-AF85-BF2B3E4DB3FA
INTRODUCTION
Gračanica is still an active opencast lignite mine, the exploitation of which began in 1938. The site has long been known for early Middle Miocene fossils of large mammals and mollusks, more recently also for fish, amphibians, reptiles, as well as arthropods, insects, sponges, protists, macroflora and plant pollen (Göhlich and Mandic, 2020). It is a type locality for many taxa from different systematic groups. So far, representatives of almost all vertebrate classes have been found in Gračanica - the exception being birds. The authors describe the first almost complete fossil bird from this locality: a new genus and species of a medium-sized duck preserved on a slab.
The oldest representatives of the crown group Anatidae are the genera Mionetta and Uyrekura from the early Oligocene of Kazakhstan (Zelenkov, 2024). From the late Oligocene, the genera Mionetta and Pinpanetta are known from France and Australia (Mayr, 2022) and Aminornis and Teleornis from Argentina (Tambussi et al., 2023). Miocene anatids include several dozen species worldwide (Mlíkovský, 2002; Zelenkov, 2020, 2023b). Sites rich in diverse anseriform fauna are known from Europe (Mlíkovský, 2002), Asia (Gorobets, 2013; Zelenkov, 2011, 2012a, 2012b, 2020, 2023b; Zelenkov and Kurochkin, 2012), Australia and New Zealand (Worthy et al., 2007, 2022; Worthy, 2009). Two species, Mionetta blanchardi from Europe and Manuherikia lacustrina from New Zealand, are exceptional because they are known from very numerous specimens (Cheneval, 1987; Worthy et al., 2007). However, most fossil duck species are represented by scarce material, sometimes even only single bones (e.g., Boev, 2007, 2020; Gál et al., 2000; Göhlich, 2009; Kessler and Venczel, 2009). More complete specimens with associated skeletal elements are rare (Bochenski et al., 2023). All this makes comparing different species of Miocene ducks very difficult (Mayr et al., 2022). It is even more difficult to determine the relationships between Anatidae (Worthy, 2009; Worthy and Lee, 2008; Worthy et al., 2022; Zelenkov et al., 2018). It is not surprising that this problem has not yet been fully resolved (Mayr et al., 2022; Worthy et al., 2022; Zelenkov, 2020). The new and almost complete specimen described in this paper is one of the few preserved on slabs, so there is no doubt that all the bones belong to the same individual. It makes a small but important contribution to expanding our knowledge of Miocene ducks.
MATERIAL AND METHODS
The osteological terminology follows Baumel and Witmer (1993) and the nomenclature and classification of modern taxa follow Dickinson and Remsen (2013). Measurements are in millimeters.
The specimen consists of fossilized bones and their impressions on a slab. The slab was thin and brittle, so immediately after the specimen was found it was reinforced with epoxy resin, which adhered to parts of some of the bones. This reinforcement allowed the specimen to be preserved in its entirety, but prevented later, more precise dissection of individual skeletal elements by one of us (GS). The exception is the os carpi ulnare, which was cut out of the specimen and is now a three-dimensional object. The specimen was examined by computed tomography performed using a Siemens EMOTION 6 device. The scan layer thickness was 0.625 mm, which formed the basis for 3D reconstruction of the specimen.
The osteological collection of the Institute of Systematics and Evolution of Animals, Polish Academy of Sciences (ISEA PAS) made the following Anatidae specimens available for comparisons: Dendrocygninae: Dendrocygna arborea, D. autumnalis, D. bicolor, D. eytoni, D. viduata. Oxyurinae: Oxyura jamaicensis. Anserinae: Anser albifrons, A. anser, A. caerulescens, A. canagicus, A. erythropus, A. fabalis, A. indicus, Branta bernicla, B. canadensis, B. leucopsis, B. ruficollis, B. sandvicensis, Cereopsis novaehollandiae, Coscoroba coscoroba, Cygnus atratus, C. columbianus, C. cygnus, C. melanocoryphus, C. olor. Anatinae: Mergini: Bucephala clangula, B. islandica, Clangula hyemalis, Lophodytes cucullatus, Melanitta fusca, M. nigra, Mergellus albellus, Mergus merganser, M. serrator, Somateria mollissima, S. spectabilis. Tadornini: Alopochen aegyptiaca, Chloephaga picta, Tadorna ferruginea, T. tadorna, T. variegata. Aythyini: Aythya ferina, A. fuligula, A. marila, A. nyroca, Netta rufina. Anatini: Amazonetta brasiliensis, Anas acuta, A. bahamensis, A. capensis, A. crecca, A. formosa, A. gibberifrons, A. platyrhynchos, A. poecilorhyncha, A. rubipres, A. superciliosa, Malacorhynchus membranaceus, Mareca strepera, M. falcate, M. penelope, M. sibilatrix, Spatula clypeata, S. cyanoptera, S. discors, S. querquedula, S. rhynchotis. Genera incertae sedis: Aix galericulata, A. sponsa, Biziura lobata, Callonetta leucophrys, Chenonetta jubata, Nettapus auritus, N. coromandelianus, N. pulchellus, Stictonetta naevosa.
The fossil specimen was compared both with the original descriptions of Miocene ducks previously described and with later publications containing descriptions of the features and/or their illustrations. These additional publications, if any, are listed here in square brackets after the taxonomic authorities of the species: Dunstanetta Worthy, Tennyson, Jones, McNamara, and Douglas, 2007 [Worthy, 2009; Worthy and Lee, 2008; Worthy et al., 2008, 2022]; Manuherikia Worthy, Tennyson, Jones, McNamara, and Douglas, 2007 [Worthy, 2009; Worthy and Lee, 2008; Worthy et al., 2008, 2022]; Mionetta blanchardi (Milne-Edwards, 1863) [Canterbury Museum’s account on www.sketchfab.com; Cheneval, 1983; Howard, 1964; Livezey and Martin, 1988; Milne-Edwards, 1867-1868; Worthy and Lee, 2008; Zelenkov, 2023c, 2024]; Mionetta consobrina (Milne-Edwards, 1867-1868) [Lambrecht, 1933; Howard, 1964; Cheneval, 1983]; Mionetta defossa Zelenkov, 2023c [Lambrecht, 1933; Švec, 1981; Cheneval, 1983; Göhlich, 2002]; Caerulonettion natator (Milne-Edwards, 1867-1868) [Lambrecht, 1933; Švec, 1981; Cheneval, 1983; Göhlich, 2002; Zelenkov, 2023c]; Pinpanetta Worthy, 2009 [Worthy et al., 2022]; Aythya denesi (Kessler, 2013) [Zelenkov 2016]; Aythya molesta (Kurochkin, 1985) [Zelenkov, 2012a, 2016]; Ayhya shihuibas Hou, 1985 [Stidham, 2015]; Miotadorna sanctibathansi Worthy, Tennyson, Jones, McNamara, and Douglas, 2007 [Worthy and Lee, 2008]; Anas kurochkini Zelenkov et Panteleyev, 2015 [Zelenkov, unpublished photo]; Matanas enrighti Worthy, Tennyson, Jones, McNamara and Douglas, 2007; Mioquerquedula minutissima Zelenkov and Kurochkin, 2012 [Zelenkov, 2023a]; Mioquerquedula soporata (Kurochkin, 1976) [Zelenkov, 2023a; Zelenkov and Kurochkin, 2012]; Mioquerquedula palaeotagaica Zelenkov, 2023b; ? Mioquerquedula sp. [Zelenkov, 2017: PIN no. 5073/142]; Bambolinietta lignitifila (Portis, 1884) [Mayr and Pavia, 2014]; Chenoanas asiatica Zelenkov, Stidham, Martynovich, Volkova, Li, Qui, 2018; Chenoanas deserta Zelenkov, 2012b [Zelenkov et al., 2018]; Chenoanas sansaniensis (Milne-Edwards, 1867-1868) [Göhlich, 2009; Zelenkov et al., 2018]; Aix praeclara Zelenkov and Kurochkin, 2012; Nogusunna conflictoides Zelenkov, 2011 [Stidham and Zelenkov, 2017]; Protomelanitta gracilis Zelenkov, 2011 [Stidham and Zelenkov, 2017; Zelenkov, 2023a]; Protomelanitta bakeri Stidham and Zelenkov, 2017; Protomelanitta velox (Milne-Edwards, 1867-1868) [Švec, 1981; Zelenkov, 2023a; Zelenkov and Kurochkin, 2012]; Tagayanetta palaeobaikalensis Zelenkov, 2023b; Selenonetta lacustrina Zelenkov, 2023b; Sharganetta mongolica Zelenkov, 2011.
The following ratios between skeletal elements were calculated: the brachial index (humerus length/ulna length), carpometacarpus length/phalanx proximalis digiti majoris length, humerus length/carpometacarpus length, and carpometacarpus length/coracoid medial length.
SYSTEMATIC PALAEONTOLOGY
Class AVES Linnaeus, 1758
Order ANSERIFORMES Wagler, 1831
Family ANATIDAE Leach, 1819
Genus Gracanicanetta gen. nov.
zoobank.org/B466431A-DAD9-4C5C-A309-10BCC3F8A98B
Type species. Gracanicanetta happi gen. et sp. nov.
Etymology. After Gračanica where the specimen was found, added to the Greek “netta” meaning “duck”.
Taxonomic remarks. The new species resembles Anatidae in overall morphology of all skeletal elements, including long processus retroarticularis of the mandibula, foramina neurovascularia on the beak and mandibula, coracoid with deeply concave and rounded cotyla scapularis, laterally protruding facies articularis humeralis, humerus with capital shaft ridge and carpometacarpus with a notch in the caudal rim of the dorsal portion of the trochlea carpalis.
Differential diagnosis. Gracanicanetta happi gen. et sp. nov. is distinguished from:
* Lavanttalornis hassleri Bochenski, Happ, Salwa, Tomek, 2023, from the late Middle Miocene of southern Austria, in: coracoid with straight medial edge of shaft that does not incline medially, humerus with distinct notch of incisura capitis in proximal outline of the bone, carpometacarpus with short synostosis metacarpalis distalis, and a relatively small value of the ratio of the length of carpometacarpus to the length of phalanx proximalis digiti majoris;
* Mionetta blanchardi (Milne-Edwards, 1863), from the late Oligocene to Middle Miocene of Europe, in: skull with beak significantly shorter than the braincase, and elongated preorbital region, clavicula very robust and thick, humerus with distinct notch of incisura capitis in proximal outline of the bone; carpometacarpus with short synostosis metacarpalis distalis;
* Caerulonettion natator (Milne-Edwards, 1867-1868), from the Early Miocene of France and the Czech Republic, and early Middle Miocene of Germany, in: coracoid with processus acrocoracoideus hardly protruding beyond the medial margin of shaft, and humerus with distinct notch of incisura capitis in proximal outline of the bone;
* Mionetta defossa Zelenkov, 2023c, from the Early Miocene of France and the Aral Sea region, Kazakhstan, Miotadorna sanctibathansi Worthy, Tennyson, Jones, McNamara, and Douglas, 2007, from the early Middle Miocene of New Zealand, and Aythya denesi (Kessler, 2013) from the Late Miocene of Hungary, in: humerus with distinct notch of incisura capitis in proximal outline of the bone;
* Manuherikia Worthy, Tennyson, Jones, McNamara, and Douglas, 2007 from the early middle Miocene of New Zealand, in: coracoid with the plane through the depth of the acrocoracoid subperpendicular to the plane of extremitas sternalis;
* Dunstanetta Worthy, Tennyson, Jones, McNamara, and Douglas, 2007, from the early middle Miocene of New Zealand, in: coracoid with processus acrocoracoideus hardly protruding beyond the medial margin of shaft, and facies articularis clavicularis without a distinct notch in its caudal margin;
* Pinpanetta Worthy, 2009, from the Late Oligocene/Early Miocene of Australia, in coracoid with processus acrocoracoideus hardly protruding beyond the medial margin of shaft, and humerus with distinct notch of incisura capitis in proximal outline of the bone;
* Sharganetta mongolica Zelenkov, 2011 from the Middle Miocene of Mongolia, in: humerus with distal margin of the caput humeri in caudal view almost straight, and the caput humeri undercut by the fossa pneumotricipitalis dorsalis;
* Nogusunna conflictoides Zelenkov, 2011 from the middle Miocene of Mongolia, in: humerus with a weak capital shaft ridge oriented towards the tuberculum dorsale;
* Protomelanitta gracilis Zelenkov, 2011, from the middle Miocene of Mongolia, in: coracoid with processus acrocoracoideus hardly protruding beyond the medial margin of shaft, and humerus with the caput humeri somewhat wider proximodistally, and tuberculum ventrale directed caudo-cranially;
* Protomelanitta bakeri Stidham and Zelenkov, 2017 from the Middle Miocene of the USA, in: humerus with the caput humeri somewhat wider proximodistally and tuberculum dorsale is oriented more caudally;
* Protomelanitta velox (Milne-Edwards, 1867), from the Middle Miocene of France, in: coracoid with a moderately deep depression in the dorsal part of the sulcus m. supracoracoidei and carpometacarpus with a deep fossa infratrochlearis;
* Chenoanas deserta Zelenkov, 2012b and Chenoanas asiatica Zelenkov, Stidham, Martynovich, Volkova, Li, Qui, 2018, both from the Middle Miocene of eastern Russia and Mongolia, Chenoanas sansaniensis (Milne-Edwards, 1867-1868), from the Early to Middle Miocene of Europe, eastern Russia and Mongolia, and Matanas enrighti Worthy, Tennyson, Jones, McNamara and Douglas, 2007, from the Early-Middle Miocene of New Zealand, in coracoid with processus acrocoracoideus hardly protruding beyond the medial margin of shaft;
* Aix praeclara Zelenkov et Kurochkin, 2012, Mioquerquedula soporata (Kurochkin, 1976), and Mioquerquedula minutissima Zelenkov et Kurochkin, 2012, all from the middle Miocene of Mongolia, in: coracoid with the plane through the depth of the acrocoracoid subperpendicular to the plane of extremitas sternalis, and processus acrocoracoideus hardly protruding beyond the medial margin of shaft;
* Tagayanetta palaeobaikalensis Zelenkov, 2023b, from Early-Middle Miocene of Lake Baikal region, Russia, and Aythya molesta (Kurochkin, 1985), from the Late Miocene of Mongolia, in coracoid with facies articularis clavicularis without a distinct notch in its caudal margin;
* Selenonetta lacustrina Zelenkov, 2023b, from Early-Middle Miocene of Lake Baikal region, Russia, in: humerus with distinct notch of incisura capitis in proximal outline of the bone, and the crista deltopectoralis noticeably longer than the proximal width of the humerus;
* Aythya shihuibas Hou, 1985, from the Late Miocene of China, in: humerus with the tuberculum dorsale elevated above caudal surface of shaft;
* Anas kurochkini Zelenkov et Panteleyev, 2015, from the Late Miocene of northeastern Sea of Azov region, Russia, in: coracoid with the plane through the depth of the acrocoracoid subperpendicular to the plane of extremitas sternalis, and facies articularis clavicularis without a distinct notch in its caudal margin;
* all extant ducks by a unique combination of characters: wide beak significantly shorter than the braincase, elongated preorbital (lacrimal) region of the skull, relatively wide sternum, coracoid with straight medial edge of shaft that does not incline medially, with the plane through the depth of the acrocoracoid subperpendicular to the plane of extremitas sternalis, and facies articularis clavicularis without a distinct notch in its caudal margin, humerus with distinct notch of incisura capitis in proximal outline of the bone, a weak capital shaft ridge oriented towards the tuberculum dorsale, and tuberculum dorsale elevated above caudal surface of shaft, carpometacarpus with short synostosis metacarpalis distalis, and a relatively small value of the ratio of the length of carpometacarpus to the length of phalanx proximalis digiti majoris, os carpi ulnare with almost flat distal surface.
Gracanicanetta happi sp. nov.
Figure 1, Figure 2A, Figure 3A, Figure 4, Figure 5A, B, D, Figure 6A, B
zoobank.org/5D64C46B-10EF-4B80-BFCB-746ED09AA113
Etymology. The species is named after Helga and Friedrich Happ, herpetologists and donors of the fossil, who dedicated their lives to the conservation of reptiles and amphibians of Carinthia, southern Austria.
Holotype. ISEA AF/GRA1 (Figure 1, Figure 2A, Figure 3A, Figure 4, Figure 5, Figure 6A, B), partly articulated, nearly complete skeleton on a slab from which os carpi ulnare has been removed and is visible as a 3D object; housed at the Institute of Systematics and Evolution of Animals, Polish Academy of Sciences, Krakow, Poland.
 Type locality and horizon. Gračanica open-cast lignite mine, still utilized commercially, situated in the Bugojno basin, between Gornji Vakuf and Donji Vakuf, central Bosnia and Herzegovina; GPS coordinates using WGS84 datum: 43.997662° N, 17.518516° E (Mandic et al., 2016). The age of the Gračanica mine deposits is estimated to be Early-Middle Miocene, late Langhian, 16-13.5 Ma, however, the dating is still considered to be contentious (Göhlich and Mandic, 2020). The specimen was found by miners during their working routine in the lower half of the 40 m thick deposits, rich in lignites (Göhlich and Mandic, 2020, fig. 1c).
Type locality and horizon. Gračanica open-cast lignite mine, still utilized commercially, situated in the Bugojno basin, between Gornji Vakuf and Donji Vakuf, central Bosnia and Herzegovina; GPS coordinates using WGS84 datum: 43.997662° N, 17.518516° E (Mandic et al., 2016). The age of the Gračanica mine deposits is estimated to be Early-Middle Miocene, late Langhian, 16-13.5 Ma, however, the dating is still considered to be contentious (Göhlich and Mandic, 2020). The specimen was found by miners during their working routine in the lower half of the 40 m thick deposits, rich in lignites (Göhlich and Mandic, 2020, fig. 1c).
Diagnosis. As for the genus.
Measurements. Measurements (in mm) including total length (TL) and others: coracoid, medial length ca 36.7; scapula TL, 46.2; humerus TL, 68.0, proximal width, 15.9, distal width, 10.5; ulna TL, 59.7; carpometacarpus TL, 39.1, proximal width, 9.6; phalanx proximalis digiti majoris TL, 17.6; femur TL, ca 36.4.
Description and Comparison
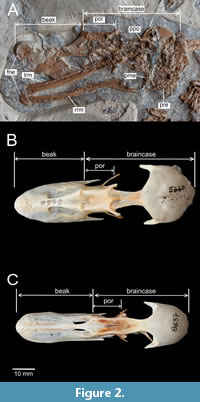 Skull and mandible. The skull is visible in ventrolateral view (Figure 1, Figure 2A). As in some extant Mergini (e.g., Bucephala, Clangula ) the beak is wide and significantly shorter than the braincase. Other extant Mergini (e.g., Melanitta), Tadornini (e.g., Tadorna), Aythyini (Aythya and Netta), Dendrocygninae (Dendrocygna) and Oxyurinae (Oxyura) also have wide beaks, but the length of their beaks is similar to the length of the braincase. The beaks of extant Anatini are usually slightly narrower but like in the Miocene Mionetta blanchardi and Lavanttalorins hassleri, their length is also similar to the length of the braincase. Completely different beaks (extremely narrow and very long) are observed in extant Mergus and Mergellus. As in derived Anatinae (i.e., Anatini, Aythyini, Mergini) and Tadornini, the preorbital (lacrimal) region of the skull is elongated; in Mionetta and Mioquerquedula, and more primitive extant Anatidae such as Dendrocygninae, Thalassornis, Biziura), this region is shortened. As in extant anseriforms, the foramina neurovascularia are visible at the end of the beak and mandible. As in extant Anatidae, a long and rostrally projecting processus postorbitalis can be distinguished within the crushed braincase. The articular part of the mandible bears the processus medialis and the long processus retroarticularis which is typical of all anatids.
Skull and mandible. The skull is visible in ventrolateral view (Figure 1, Figure 2A). As in some extant Mergini (e.g., Bucephala, Clangula ) the beak is wide and significantly shorter than the braincase. Other extant Mergini (e.g., Melanitta), Tadornini (e.g., Tadorna), Aythyini (Aythya and Netta), Dendrocygninae (Dendrocygna) and Oxyurinae (Oxyura) also have wide beaks, but the length of their beaks is similar to the length of the braincase. The beaks of extant Anatini are usually slightly narrower but like in the Miocene Mionetta blanchardi and Lavanttalorins hassleri, their length is also similar to the length of the braincase. Completely different beaks (extremely narrow and very long) are observed in extant Mergus and Mergellus. As in derived Anatinae (i.e., Anatini, Aythyini, Mergini) and Tadornini, the preorbital (lacrimal) region of the skull is elongated; in Mionetta and Mioquerquedula, and more primitive extant Anatidae such as Dendrocygninae, Thalassornis, Biziura), this region is shortened. As in extant anseriforms, the foramina neurovascularia are visible at the end of the beak and mandible. As in extant Anatidae, a long and rostrally projecting processus postorbitalis can be distinguished within the crushed braincase. The articular part of the mandible bears the processus medialis and the long processus retroarticularis which is typical of all anatids.
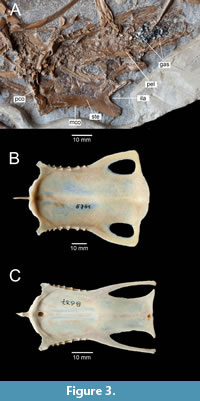 Sternum. The sternum is visible in dorsal view (Figure 1, Figure 3A). Although the right part of the sternum is partially obscured by the pelvis, the sternum was wide in relation to its length. Similar sternum proportions are observed in extant Mergini (except Mergus and Mergellus) and Aythyini. Ducks belonging to Anatini, Tadornini, Oxyurini and Dendrocygnini have a more elongated sternum. As in most extant anatids but unlike Oxyura, the pila coracoidea forms a thickened ridge (Worthy and Lee, 2008: character 34). The remaining details of the rostrum sterni are not visible because they are covered by crushed vertebrae.
Sternum. The sternum is visible in dorsal view (Figure 1, Figure 3A). Although the right part of the sternum is partially obscured by the pelvis, the sternum was wide in relation to its length. Similar sternum proportions are observed in extant Mergini (except Mergus and Mergellus) and Aythyini. Ducks belonging to Anatini, Tadornini, Oxyurini and Dendrocygnini have a more elongated sternum. As in most extant anatids but unlike Oxyura, the pila coracoidea forms a thickened ridge (Worthy and Lee, 2008: character 34). The remaining details of the rostrum sterni are not visible because they are covered by crushed vertebrae.
Coracoid. Coracoids are visible in dorsal view; the right one is complete but partially obscured by other bones, and the left one shows only a fragment of the shaft (Figure 1, Figure 4). In length, the coracoid is similar to that of extant Spatula querquedula but its shaft is more massive and wider. As in Chenoanas, the medial edge of the shaft is straight and does not deviate medially; in Lavanttalornis, the omal section of the shaft inclines strongly medially. As in Mergini and part of Aythyini but unlike Mioquerquedula minutissima, Manuherikia lacustrina, Aix praeclara, Anas kurochkini, and extant Anatitni and Tadornini, the plane through the depth of the acrocoracoid is subperpendicular to the plane of extremitas sternalis (Worthy and Lee, 2008: character 45). The processus acrocoracoideus projects only slightly beyond the medial edge of the shaft, which distinguishes it from Chenoanas, Dunstanetta, Pinpanetta, Caerulonettion natator, Matanas enrighi, Aix praeclara, Mioquerquedula minutissima, Mioquerquedula soporata, Mioquerquedula palaeotagaica, Protomelanitta gracilis, and 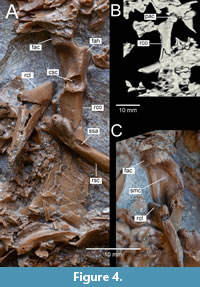 many extant taxa including Anatini and Aix. Unlike Dunstanetta, Aythya molesta, Tagayanetta palaeobaikalensis, Anas kurochkini and extant Anas s.l., facies articularis clavicularis does not bear a distinct notch in its caudal margin although its surface is slightly concave. There is a moderately deep depression in the dorsal part of the sulcus m. supracoracoidei, which differs from Protomelanitta velox, where this sulcus is without a pronounced concavity and pits. As in all anatids, the margin of the facies articularis humeralis projects laterally, and the cotyla scapularis is large, rounded and deeply concave.
many extant taxa including Anatini and Aix. Unlike Dunstanetta, Aythya molesta, Tagayanetta palaeobaikalensis, Anas kurochkini and extant Anas s.l., facies articularis clavicularis does not bear a distinct notch in its caudal margin although its surface is slightly concave. There is a moderately deep depression in the dorsal part of the sulcus m. supracoracoidei, which differs from Protomelanitta velox, where this sulcus is without a pronounced concavity and pits. As in all anatids, the margin of the facies articularis humeralis projects laterally, and the cotyla scapularis is large, rounded and deeply concave.
Scapula. The right scapula is visible in ventrolateral view (Figure 1, Figure 4). This element has a typical duck-like appearance with an extended acromion and a moderately prominent tuberculum coracoideum. In the ventral edge, just below the articular part, there is a “shelf-shaped” attachment with a longitudinal depression. A similar structure is found in extant Clangula hyemalis. In Aythya, Melanitta and Bucephala there is a flattening but without a depression, while in Anas the flattening, if present, is very small.
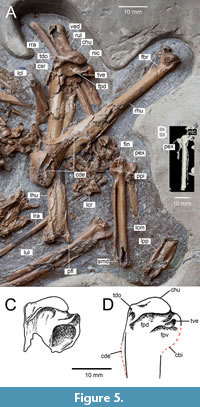 Clavicula. Omal fragments of both claviculae have been preserved; the right one is visible in lateral view, and the left in medial view (Figure 1, Figure 4, Figure 5). Unlike Mionetta blanchardi and all extant Anatidae, the clavicula is very robust and thick also lateromedially. The thick facies articularis acrocoracoidea is well developed and moderately long, somewhat similar to that in Melanitta ; in many extant Anatidae it is longer and rather narrow (e.g. Anatini, Aythyini) or poorly developed (e.g., Dendrocygninae, Tadornini, Somateria ).
Clavicula. Omal fragments of both claviculae have been preserved; the right one is visible in lateral view, and the left in medial view (Figure 1, Figure 4, Figure 5). Unlike Mionetta blanchardi and all extant Anatidae, the clavicula is very robust and thick also lateromedially. The thick facies articularis acrocoracoidea is well developed and moderately long, somewhat similar to that in Melanitta ; in many extant Anatidae it is longer and rather narrow (e.g. Anatini, Aythyini) or poorly developed (e.g., Dendrocygninae, Tadornini, Somateria ).
Humerus. The left humerus is visible in caudal view and the right humerus in dorsocranial view (Figure 1, Figure 5). Although the proximal part of the shaft is partially crushed, a weak capital shaft ridge oriented to the tuberculum dorsale is visible, as in Protomelanitta. This ridge orientation is also seen in fossil Mionetta, Manuherikia, and Dunstanetta and in extant Oxyura and Mergini, except that the ridge is prominent in these taxa (Worthy and Lee, 2008: character 51). In other taxa including fossil Nogusunna and Miotadorna and extant Dendrocygninae and Tadornini, the prominent capital shaft ridge is oriented between tubercule dorsale and caput humeri. Miocene Aythya shihuibas and extant Anatini have virtually no capital shaft ridge. Of all Miocene taxa, the caput humeri is most similar to that of Protomelanitta. It is true that it is wider proximodistally than in Protomelanitta gracilis and P. bakeri but still relatively narrow, distinctly narrower than, for example, in Caerulonettion natator. The distal margin of the caput humeri in caudal view is almost straight as in Protomelanitta ; in Sharganetta and especially in Caerulonettion the distal margin is convex. Similar to, among others, Protomelanitta gracilis, P. bakeri and Nogosunna conflictoides, the fossa pneumotricipitalis dorsalis is narrower than the fossa pneumotricipitalis ventralis (Worthy and Lee, 2008: character 53) and undercuts the caput humeri; in Sharganetta and extant Dendrocygninae, Tadornini, Aythyini and Anatini, the fossa does not undercut the head (Worthy and Lee, 2009: character 134). Although the apex of the tuberculum ventrale is missing, it seems that, like in Manuherikia lacustrina, Lavanttalornis hassleri and extant Oxyurinae, Mergini, Aythyini and Anatini, it was directed caudo-cranially (Worthy and Lee, 2008: character 57) because the caudal border of fossa pneumotricipitalis ventralis is not concave as in Protomelanitta gracilis but straight or even slightly convex. In Protomelanitta gracilis, Mionetta blanchardi, Miotadorna sanctibathansi, Chenoanas, Sharganetta and extant Dendrocygninae and Tadornini, the tuberculum ventrale is directed proximally. As in most extant ducks including Oxyurinae, Mergini, Aythyini and Anatini, the proximal profile of the humerus is interrupted by a distinct notch created by incisura capitis (Worthy and Lee, 2008: character 59). A similarly distinct notch is also found in many fossil taxa such as Sharganetta mongolica, Nogusunna conflictoides, Protomelanitta gracilis, P. bakeri, Aythya molesta, A. shihuibas, Manuherikia and Dunstanetta. A shallow notch was reported for Mionetta blanchardi, Mionetta defossa, Carulonettion natator, Miotadorna sanctibathansi, Aythya denesi and Lavanttalornis hassleri whereas in Selenonetta and Pinpanetta as well as extant Dendrocygninae and Tadornini it is almost missing. The tuberculum dorsale is similar to that of Protomelanitta in that it is subtriangular, has only a slightly extended distal margin, and is somewhat elevated above the caudal surface of shaft - the latter not as much as the modern Dendrocygninae but more than the Miocene Aythya shihuibas, modern Aythyini, Mergini, or Anatini which are defined as “coplanar” (Worthy and Lee, 2008: character 56). Unlike in Protomelanitta bakeri but similar to P.garacilis, the tuberculum dorsale is oriented more caudally than dorsally. Proximal fragments of the crista deltopectoralis are visible on both humeri, and the right humerus also shows remnants of the connection between the crista deltopectoralis and the humeral shaft, which allows for the assessment of the length of the crista. Unlike Selenonetta, the crista deltopectoralis is noticeably longer than the proximal width of the humerus. Other details of the proximal humerus, including fossa pneumotricipitalis ventralis and crista bicipitalis, are not visible. The processus flexorius extends distally roughly as far as the condylus dorsalis (Worthy and Lee, 2008: character 63). As in many extant Mergini, the fossa brachialis is large, wide and has a distinct ventral margin; in extant Anatini, the fossa is usually narrower and has a smaller surface area.
Ulna. The left ulna is visible in ventral view, and the distal part of the right bone is visible in dorsal view (Figure 1, Figure 5). The ulna is noticeably shorter than the humerus. The proximal part is too poorly preserved for meaningful comparisons. As in some Mergini (e.g., Melanitta), the ventral edge of the condylus dorsalis is somewhat elongated and merges quite gently with the shaft; in Anatini, the edge is semicircular and joins more abruptly with the shaft.
Carpometacarpus. The left carpometacarpus is visible in ventral view, the right one is missing (Figure 1, Figure 5). Processus extensorius is less prominent cranially than in extant Tadornini; as in the similarly sized extant Aythya nyroca, it is relatively wide proximodistally and its proximal edge is oriented mainly cranially, whereas in many extant Anatini it is directed proximocranially. Contrary to e.g., Mioquerquedula palaeotagaica and Protomelanitta velox, the fossa infratrochlearis is deep, and another fossa located distocaudally to the processus pisiformis is also very pronouned. The caudal rim of the dorsal portion of the trochlea carpalis has a wide notch, which can be seen on the CT scan. The synostosis metacarpalis distalis is shorter than the width of the carpometacarpus measured at the distal end of the spatium intermetacarpale (Worthy and Lee, 2008: character 84). Short synostosis is present in some extant Mergini (Bucephala, Clangula, Melanitta) and Tadornini (Tadorna). A long (i.e., different) synostosis is present in Miocene Lavanttalornis hassleri, Mionetta blanchardi, Manuherikia lacustrina and in many extant taxa, including Dendrocygninae, Aythyini and Anatini.
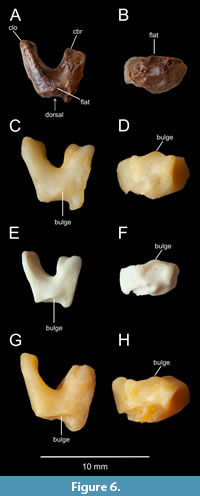 Os carpi ulnare. The left os carpi ulnare was originally visible in proximal view but has been carved from the slab and is now a three-dimensional object (Figure 1, Figure 6). Its distal surface is different from all extant ducks (this element has not yet been described in Miocene ducks): In dorsal view, the distal edge is almost flat, while in extant Anatidae there is a bulge of varying shape.
Os carpi ulnare. The left os carpi ulnare was originally visible in proximal view but has been carved from the slab and is now a three-dimensional object (Figure 1, Figure 6). Its distal surface is different from all extant ducks (this element has not yet been described in Miocene ducks): In dorsal view, the distal edge is almost flat, while in extant Anatidae there is a bulge of varying shape.
Pelvis. The damaged pelvis, which appears rather narrow, is visible in dorsal view against the background of the sternum (Figure 1, Figure 3A). The right femur is in articulation with it. There is a cluster of several dozen smooth gastroliths in the postacetabular region; some of them are whitish and others are black. The cluster measures approximately 18 x 10 mm. The dimensions of the exposed stones range approximately from 0.5 to 2.5 mm.
Femur. The crushed but almost complete right femur, in articulation with the pelvis, is visible in caudolateral view (Figure 1). In the proximal part, the outline of the trochanter has been preserved; it protrudes above the caput femoris. Contrary to all extant Anatidae, the shaft is very thick for its length; it does not seem that this thickness results exclusively from crushing because both edges (cranial and caudal) are smooth. Similarly stout bones, albeit of the wing, have been observed in Bambolinetta ; leg elements are not known for this fossil. The shaft is moderately bent caudally in the distal third (Worthy and Lee, 2008: character 93). Members of extant Anas have straight shafts, whereas Aythni and Mergini usually have shafts that are bent to varying degrees. The distal part does not allow a meaningful description.
Ratios between skeletal elements. The brachial index (humerus length/ulna length) for Gracanicanetta happi is 1.1 and is the same as for Lavanttalornis hassleri and many extant species from all Anatinae groups (Appendix 1). Slightly lower values (approximately 1.0) are observed in all Dendrocygninae and parts of Tadornini, and slightly higher values (1.2) are observed in representatives of various groups. The length ratio of carpometacarpus / phalanx proximalis digiti majoris is more diversified in Anatidae. This ratio for Gracanicanetta happi is 2.2. No extant species examined had such a low value of this ratio; the closest value (2.3) occurred in some Dendrocygna, Oxyura and Mergini. Higher values (2.4 - 2.7) were observed in various taxonomic groups. Lavanttalornis hassleri also showed a higher ratio (2.4). The ratio humerus length / carpometacarpus length for Gracanicanetta happi is 1.7. This value was higher (1.8 - 2.0) in all examined representatives of Dendrocygninae (5 species), Oxyurinae (1 species) and Aythyini (5 species) (Appendix 1). The vast majority of Anatini (15 out of 20) showed slightly lower values of this ratio (1.6), but the rest had the same ratio (1.7). A similar range of ratio was in Tadornini (1.6 - 1.7). For Mergini, the values of the ratio ranged from 1.6 to 1.8, with more than half of the species (6 out of 10) having the same value as for Gracanicanetta (1.7). The carpometacarpus length / coracoid medial length ratio for Gracanicanetta is 1.1 (Appendix 1). The ranges encompassing this ratio value were found in Mergini (09 - 1.2), Aythyini (0.9 - 1.1) and Anatini (1.1 - 1.2). All Dendrocygninae and Tadornini had larger ratio values (1.2 - 1.4 and 1.3 - 1.4, respectively), and Oxyurini had a distinctly smaller value (0.9).
DISCUSSION
Most Miocene species have been described as new genera (Mayr et al., 2022); extant anatid genera emerged at the end of the Miocene (Zelenkov, 2020). Many Miocene anatids show a mosaic of features typical of various modern taxa (Zelenkov, 2012b) and Gracanicanetta happi is no exception, as has been shown in the differential diagnosis. In Gracanicanetta happi, the most important phylogenetic features are preserved on the coracoid and humerus. The fact that the plane of the acrocoracoid is inclined with respect to the plane of the extremitas sternalis, and that the processus acrocoracoideus does not protrude far medially excludes Anatini and suggests a diving duck in the ecological sense of the word (Zelenkov, 2011, 2023a). Also, a narrow pelvis suggests diving ability (Raikow, 1970; Worthy and Lee, 2008). A relatively wide sternum does not exclude diving because some extant diving species have a wide sternum and a narrow pelvis (Raikow, 1971). The exclusion of Anatini is also supported by several other features on various skeletal elements (see description above). Many features seem to agree with Mergini but none of them is exclusive to this tribe; they also occur in other groups of anatids. In studying Protomelanitta velox Zelenkov (2023a) noted that modern Mergini (diving ducks in the ecological sense) tend to have a shorter carpometacarpus than Anatini (dabbling ducks). Our preliminary analysis of the two ratios (humerus/carpometacarpus and carpometacarpus/coracoid) to some extent confirms this observation. However, the matter seems to be somewhat more complicated, mainly because of the large variation among Mergini. Mergus and Mergellus have the relatively shortest carpometacarpi, also Clangula and Lophodytes differ from Melanitta and Somateria, whose two ratios may be the same as in some Anatini. These two ratios seem to differentiate Dendrocygninae and Oxyurinae, as well as Aythyini and Tadornini from other taxonomic groups of ducks. However, the values found for Gracanicanetta are within the range of both Mergini (diving ducks in the ecological sense) and Anatini (dabbling ducks), making it difficult to draw firm conclusions based on this criterion. There is quite a large diversity of taxa within the Mergini tribe. The extant genera Mergus and Mergellus have anatomical adaptations for piscivory (e.g., narrow long beaks, narrow sternae), and they can certainly be ruled out. In terms of the appearance of its beak and sternum, Gracanicanetta happi is more like the modern genera Bucephala or Clangula, but it is likely that these characteristics reflect lifestyle rather than phylogenetic relationship.
Gracanicanetta happi was probably diving for food. However, it is difficult to clearly indicate its more precise systematic affiliation. The preserved skeletal elements, on the one hand, show a mosaic of features characteristic of various groups of ducks, and on the other hand, features specific to Gracanicanetta happi including a shelf-shaped attachment on the scapula, robust clavicula, flat distal surface of os carpi ulnare, and long (compared to carpometacarpus) phalanx proximalis digiti majoris. This set of features indicates a new species, and the age of the sediment (Middle Miocene) rather excludes an extant genus (Zelenkov, 2020). In Gracanicanetta happi, the humerus, which is one of the most important carriers of the phylogenetic signal (Zelenkov, 2012b and references therein), displays both primitive (e.g., elevated tuberculum dorsale) and progressive (e.g., caudo-cranially oriented tuberculum ventrale) characters. The advanced morphology of various humeral characters was acquired independently in at least two anatid lineages (Anatinae and Oxyurinae), suggesting parallel evolution (Zelenkov, 2012b). The situation is further complicated by the fact that each of the diving duck tribes (Oxyurini, Mergini and Aythyini) underwent convergent evolution (Stidham and Zelenkov, 2017). In some respects, Gracanicanetta is like Protomelanitta. However, differences between the two genera in the tuberculum ventrale (one of the diagnostic features of the genus Protomelanitta) indicate that the new specimen belongs to a separate genus, which is nevertheless closely related to Protomelanitta. Stidham and Zelenkov (2017) conclude that the placement of Protomelanitta within a specific clade of diving ducks is not clearly indicated. A similar problem is with the placement of Gracanicanetta. Due to the mosaic of characters, it may be a primitive representative of any clade of diving ducks.
Gastroliths are found in many different species of birds, not only in those that feed on plant food (Norman and Brown, 1985; Panichev and Seryodkin, 2022), but they are also present in the stomachs of modern ducks (Norman and Brown, 1985; Panichev and Seryodkin, 2022). The cluster of several dozen small, polished stones in Gracanicanetta happi is in a place that anatomically corresponds to the approximate location of the gizzard in ducks, which confirms that they are indeed gastroliths. This makes Gracanicanetta happi the first fossil duck with documented gastroliths. This is probably because most fossils are loose bones, and only a few specimens are preserved as reasonably complete remains on slabs where gastroliths are likely to be recognized. It is most often assumed that gastroliths form a gastric mill in gizzard, but they may also play a role in mineral supplementation or be ingested accidentally (Wings, 2007). Thus, the presence of gastroliths suggests herbivory but as mentioned above, this is not a strict correlation and in the case of fossil species, caution in interpretation is recommended (O’Connor et al., 2018). Future palynological studies of these gastroliths may shed light on the composition of plants consumed by Gracanicanetta happi.
The absence of gnawing marks on the one hand and the preservation of the skeleton, mainly in articulation on the other suggest a rapid covering of the body in an anoxic calm environment, without major underwater turbulence that could have stimulated its dismemberment and loss of skeletal elements (Bienkowska-Wasiluk, 2010; Vasilyan, 2020). This may suggest that the fossil was deposited in a waterier part of the swamp, which allowed for rapid submergence. However, it may also suggest a multitude of suspended particles that quickly and densely covered the submerged body. The lignite layers from Gracanica, where the fossil was discovered, also indicate a large amount of wood and therefore a peat-forming process, which would explain the abundance of suspended particles and the rapid and dense covering that led to an anoxic environment.
ACKNOWLEDGEMENTS
We are grateful to D. Sobieraj (Poland) for processing the photographs and making the drawings, and to N. Zelenkov (Russia) and another anonymous referee whose reviews improved the manuscript. We sincerely thank L. Kwaśniewski, director of the Regional Subcarpathian John Paul II Hospital in Krosno for providing the computed tomography scan for our research. We are also indebted to M. Gródek-Bargieł, head of the Department of Imaging Diagnostics, and her team B. Muszyńska and A. Pelczarska for performing the computed tomography scan, technical development and digital processing of the specimen images.
REFERENCES
Baumel, J.J. and Witmer, L.M. 1993. Osteologia, p. 45–132. In Baumel, J.J., King, A.S., Breazile, J.E., Evans, H.E., and Vanden Berge, J.C. (eds.), Handbook of avian anatomy: Nomina anatomica avium. Publications of the Nuttall Ornithological Club, No 23. Nuttal Ornithological Club, Cambridge, Massachusetts.
Bienkowska-Wasiluk, M. 2010. Taphonomy of Oligocene teleost fishes from the Outer Carpathians of Poland. Acta Geologica Polonica, 60:479–533.
Bochenski, Z.M., Happ, J., Salwa, G., and Tomek, T. 2023. An intriguing new species of dabbling duck (Aves: Anseriformes) from the middle Miocene of Austria. Palaeontologia Electronica, 26(3):a52.
https://doi.org/10.26879/1334
Boev, Z. 2007. Neogene avifaunas of Bulgaria (a brief review), p. 26–35. In Bakardjieva, N., Chankova, St., Krastanov, B., and Gateva, Sv. (eds.). Evolution and Ecology-2007. Union of the Scientists of Bulgaria. 3rd National Seminar. Proceedings, Sofia.
Boev, Z. 2020. A little dabbling duck (Anatini Vigors, 1825 - Anseriformes Wagler, 1831) from the Late Miocene of Kremikovtsi (Bulgaria). Bulletin of the Natural History Museum - Plovdiv, 5:25–31.
Canterbury Museum. 2020. Mionetta blanchardi humerus, 3D model available from Sketchfab at
https://skfb.ly/6VUYs. Access 21 October 2024.
Cheneval, J. 1983. Les Anatidae (Aves, Anseriformes) du Gisement aquitanien de Saint-Gerand-Le-Puy (Allier, France), p.525–538. In Buffetaut, E., Mazin, J.M., and Salmon, E. (eds.), Actes du symposium paléontologique G. Cuvier. Montbeliard.
Cheneval, J. 1987. Les Anatidae (Aves, Anseriformes) du Miocène de France. Révision systématique et évolution. Travaux et Documents des Laboratoires de Géologie de Lyon, 99:137–157.
Dickinson, E.C. and Remsen, Jr. J.V. 2013. The Howard and Moore complete checklist of the birds of the world, Volume 1, Aves Press. Eastbourne, UK.
Gál, E., Hír, J., Kessler, E., Kókay, J., and Márton, V. 2000. Middle Miocene fossils from the section of the road at the Rákóczi Chapel, Mátraszőlős. II. Locality Mátraszőlős 2. Folia Historico Naturalia Musei Matraensis, 24:39–75. (In Hungarian with English summary)
Göhlich, U.B. 2002. The Avifauna of the Miocene Fossil-Lagerstätte Sandelzhausen (Bavaria, Southern Germany). Zitteliana, 22:169–190.
Göhlich, U.B. 2009. The early Vallesian vertebrates of Atzelsdorf (late Miocene, Austria). Annalen des Naturhistorischen Museums in Wien, 111A:509–514.
Göhlich, U. B. and Mandic, O. 2020. Introduction to the special issue “The drowning swamp of Gračanica (Bosnia-Herzegovina)–a diversity hotspot from the middle Miocene in the Bugojno Basin”. Palaeobiodiversity and Palaeoenvironments, 100(2):281–293.
https://doi.org/10.1007/s12549-020-00437-0
Gorobets, L.V. 2013. Remains of Miocene birds from the Olkhon Island in the collections of the National Science and Natural History Museum of the National Academy of Sciences of Ukraine, p. 68–73. In Kalyakin, M.V., Zelenkov, N.V., and Gavrilov, V.M. (eds.), Problemy evolyutsii ptits: sistematika, morfologiya, ekologiya i povedenie (Avian evolution: systematics, morphology, ecology, and behavior). KMK Scientific Press Ltd., Moscow. (In Russian)
Hou, L.H. 1985. Upper Miocene birds from Lufeng, Yunnan. Acta Anthropologica Sinica, 4:118–126.
Howard, H. 1964. Fossil Anseriformes, p. 233–326. In Delacour, J. (ed.), The Waterfowl of the World, volume 4, Country Life Limited, London.
Kessler, E. 2013. A Kárpát medence madárvilágának őslénytani kézikönyv [Paleontological handbook of birdlife in the Carpathian Basin]. Könyvmühel, Miskolc.
Kessler, E. and Venczel, M. 2009. Bird remains from the Middle Miocene of Subpiatră. Nymphaea. Folia naturae Bihariae, 36:27–36.
Kurochkin, E.N. 1976. New data on Pliocene birds of Western Mongolia. Transactions of the Joint Soviet-Mongolian Palaeontological Expedition, 3:51–67. (In Russian)
Kurochkin, E.N. 1985. Birds of Central Asia in the Pliocene. Transactions of the Joint Soviet-Mongolian Palaeontological Expedition, 26:1-119. (In Russian)
Lambrecht, K. 1933. Handbuch der Palaeornithologie. Borntraeger, Berlin.
Leach, W.E. 1819. Eleventh room, p. 63–68. In British Museum. Synopsis of the contents of the British Museum (15th ed.). Richard & Arthur Taylor.
Linnaeus, C. 1758. Systema Naturae per Regna Tria Naturae (10th ed.), volume 1: Regnum Animale. Salvii, L. Holmiae.
Livezey, B.C. and Martin, L.D. 1988. The Systematic Position of the Miocene Anatid Anas [?] blanchardi Milne-Edwards. Journal of Vertebrate Paleontology, 8:196–211.
https://doi.org/10.1080/02724634.1988.10011698
Mandic, O., Göhlich, U.B., Krijgsman, W., De Leeuw, A., and Hrvatović, H. 2016. Lake Bugojno - three lacustrine cycles, p. 31–35. In Mandic, O., Sant, K., Andrić, N., Horvatinčić, N., Ilijanić, N., Miko, S., Markić, N., Novosel, A., Matenco, L., and Hrvatović, H., (eds.), Dinarides lakes and basins–From 18 Ma to Present. Field trip guide-book. Lake-basin evolution. RCMNS Interim Colloquium 2016 and Croatian Geological Society Limnogeology Workshop, Zagreb.
Mayr, G. 2022. Paleogene fossil birds. Springer, Berlin.
Mayr, G. and Pavia, M. 2014. On the true affinities of Chenornis graculoides Portis, 1884, and Anas lignitifila Portis, 1884–an albatross and an unusual duck from the Miocene of Italy. Journal of Vertebrate Paleontology, 34:914–923.
https://doi.org/10.1080/02724634.2013.821076
Mayr, G., Lechner, T., and Böhme, M. 2022. Nearly complete leg of an unusual, shelduck-sized anseriform bird from the earliest late Miocene hominid locality Hammerschmiede (Germany). Historical Biology, 4:465–474.
https://doi.org/10.1080/08912963.2022.2045285
Milne-Edwards, A. 1863. Mémoire sur la distribution géologique des oiseaux fossiles et description de quelques espècies nouvelles. Annales des Sciences Naturelles, 4:133–176.
Milne-Edwards, A. 1867-1868. Recherches anatomiques et paléontologiques pour servir à l’histoire des oiseaux fossiles de la France. Éditions Masson, Paris.
Mlíkovský, J. 2002. Cenozoic Birds of the World, volume 1. Europe: Ninox Press, Praha.
Norman, F. I., and Brown, R. S. 1985. Gizzard grit in some Australian waterfowl. Wildfowl, 36(36):77–80.
O’Connor, J., Wang, X., Sullivan, C., Wang, Y., Zheng, X., Hu, H., Zhang, X., and Zhou, Z. 2018. First report of gastroliths in the Early Cretaceous basal bird Jeholornis. Cretaceous Research, 84:200–208.
https://doi.org/10.1016/j.cretres.2017.10.031
Panichev, A.M., and Seryodkin, I. V. 2022. The mineral composition of gastroliths in the stomachs of Anatidae in Primorsky Region and the importance of silicon minerals in the physiology of birds. Amurian Zoological Journal, 14(3):469–491.
https://doi.org/10.33910/2686-9519-2022-14-3-469-491
Portis, A. 1884. Contribuzioni alla ornitolitologia italiana. Memorie della Reale Accademia delle Scienze di Torino, 36:361–384.
Raikow, R.J. 1970. Evolution of diving adaptations in the stifftail ducks. University of California Press, Berkeley, Los Angeles, London.
Raikow, R.J. 1971. The osteology and taxonomic position of the White-backed Duck, Thalassornis leuconotus. The Wilson Bulletin, 83:270–277.
https://www.jstor.org/stable/4160101
Stidham, T.A. 2015. Re-description and phylogenetic assessment of the Late Miocene ducks Aythya shihuibas and Anas sp. (Aves: Anseriformes) from Lufeng, Yunnan, China. Vertebrata PalAsiatica, 53:335–349.
http://www.vertpala.ac.cn/EN/Y2015/V53/I4/335
Stidham, T.A. and Zelenkov, N.V. 2017. North American-Asian aquatic bird dispersal in the Miocene: evidence from a new species of diving duck (Anseriformes: Anatidae) from North America (Nevada) with affinities to Mongolian taxa. Alcheringa: An Australasian Journal of Palaeontology, 41:222–230.
https://doi.org/10.1080/03115518.2016.1224439
Švec, P. 1981. Lower Miocene Birds from Dolnice (Cheb Basin), Western Bohemia: Part 2, Časopis pro Mineralogii a Geologii, 26:45–56.
Tambussi, C.P., Degrange, F.J., and de Mendoza, R.S. 2023. “The present state of knowledge of the Cenozoic birds of Argentina” by Tonni 1980: four decades after. Publicación Electrónica de la Asociación Paleontológica Argentina, 23(1):255–295.
https://doi.org/10.5710/PEAPA.13.08.2022.418
Vasilyan, D. 2020. Fish, amphibian and reptilian assemblage from the middle Miocene locality Gračanica–Bugojno palaeolake, Bosnia and Herzegovina. Palaeobiodiversity and Palaeoenvironments, 100(2):437-455.
https://doi.org/10.1007/s12549-019-00381-8
Wagler, J.G. 1831. Einige Mitteilungen über Thiere Mexicos. Isis Von Oken 5:510–534.
Wings, O. 2007. A review of gastrolith function with implications for fossil vertebrates and a revised classification. Acta Palaeontologica Polonica, 52(1):1-16.
Worthy, T.H. 2009. Descriptions and phylogenetic relationships of two new genera and four new species of Oligo-Miocene waterfowl (Aves: Anatidae) from Australia. Zoological Journal of the Linnean Society, 156:411–454.
https://doi.org/10.1111/j.1096-3642.2008.00483.x
Worthy, T.H., Tennyson, A.J., Jones, C., McNamara, J.A., and Douglas, B.J. 2007. Miocene waterfowl and other birds from Central Otago, New Zealand. Journal of Systematic Palaeontology, 5:1–39.
https://doi.org/10.1017/S1477201906001957
Worthy, T.H. and Lee, M.S. 2008. Affinities of Miocene waterfowl (Anatidae: Manuherikia, Dunstanetta and Miotadorna ) from the St Bathans Fauna, New Zealand. Palaeontology, 51:677–708.
http://doi.org/10.1111/j.1475-4983.2008.00778.x
Worthy, T.H., Tennyson, A.J., Hand, S.J., and Scofield, R.P. 2008. A new species of the diving duck Manuherikia and evidence for geese (Aves: Anatidae: Anserinae) in the St Bathans Fauna (Early Miocene), New Zealand. Journal of the Royal Society of New Zealand, 38:97–114.
https://doi.org/10.1080/03014220809510549
Worthy, T.H., Scofield, R.P., Salisbury, S.W., Hand, S.J., De Pietri, V.L., Blokland, J.C., and Archer, M. 2022. A new species of Manuherikia (Aves: Anatidae) provides evidence of faunal turnover in the St Bathans fauna, New Zealand. Geobios, 70:87–107.
https://doi.org/10.1016/j.geobios.2021.08.002
Zelenkov, N.V. 2011. Diving ducks from the middle Miocene of western Mongolia. Paleontological Journal, 45:191–199.
https://doi.org/10.1134/S0031030111020195
Zelenkov, N.V. 2012a. Neogene geese and ducks (Aves: Anatidae) from localities of the Great Lakes Depression, western Mongolia. Paleontological Journal, 46:607–619.
https://doi.org/10.1134/S0031030112060123
Zelenkov, N.V. 2012b. A new duck from the Middle Miocene of Mongolia, with comments on Miocene evolution of ducks. Paleontological Journal, 46:520–530.
https://doi.org/10.1134/S0031030112050139
Zelenkov, N.V. 2016. Revision of non-Passeriform birds from Polgárdi (Hungary, Late Miocene): 1. Anseriformes. Paleontological Journal, 50:514–517.
https://doi.org/10.1134/S0031030116050142
Zelenkov, N.V. 2017. Finds of fragmentary bird skeletons in the Middle Miocene of the northern Caucasus. Doklady Biological Sciences, 477:223–226.
https://doi.org/10.1134/S0012496617060072
Zelenkov, N.V. 2020. Cenozoic evolution of Eurasian anatids (Aves: anatidae s.l.). Biology Bulletin Reviews, 10:417–426.
https://doi.org/10.1134/S2079086420050096.
Zelenkov, N.V. 2023a. Small ducks (Aves: Anatidae) from the Early-Middle Miocene of Eurasia. 1. A revision of Anas velox Milne-Edwards, 1868 and Anas soporata Kurochkin, 1976. Paleontological Journal, 57(4):452–462.
https://doi.org/10.1134/S0031030123040159
Zelenkov, N.V. 2023b. Small ducks (Aves: Anatidae) from the Early-Middle Miocene of Eurasia. Part 2. The fauna of Tagay locality (Baikal Region; eastern Siberia). Paleontological Journal, 57(5):560–572.
https://doi.org/10.1134/S003103012305012X
Zelenkov, N. V. 2023c. Small Ducks (Aves: Anatidae) from the Early-Middle Miocene of Eurasia. 3. A Revision of Mionetta natator (Milne-Edwards, 1867). Paleontological Journal, 57(6), 659-670.
https://doi.org/10.1134/S0031030123060114
Zelenkov, N. 2024. A Remarkable Diversity of Waterfowl (Aves: Anseriformes) from the Upper Eocene and Lower Oligocene of Kazakhstan. Journal of Vertebrate Paleontology.
https://doi.org/10.1080/02724634.2024.2374306
Zelenkov, N.V. and Kurochkin, E.N. 2012. Dabbling ducks (Aves: Anatidae) from the middle Miocene of Mongolia. Paleontological Journal, 46:421–429.
https://doi.org/10.1134/S0031030112040132
Zelenkov, N.V. and Panteleyev, A.V. 2015. Three bird taxa (Aves: Anatidae, Phasianidae, Scolopacidae) from the Late Miocene of the Sea of Azov (Southwestern Russia), Paläontologische Zeitschrift, 89:515–527.
https://doi.org/10.1007/s12542-014-0238-0
Zelenkov, N.V., Stidham, T.A., Martynovich, N.V., Volkova, N.V., Li, Q., and Qiu, Z. 2018. The middle Miocene duck Chenoanas (Aves, Anatidae): new species, phylogeny and geographical range. Papers in Palaeontology, 4:309–326.
https://doi.org/10.1002/spp2.1107
