

 New primates from the Eocene of Saskatchewan, Canada: Revision of the primates from the Cypress Hills Formation with description of new taxa
New primates from the Eocene of Saskatchewan, Canada: Revision of the primates from the Cypress Hills Formation with description of new taxa
Article number: 26.2.20
https://doi.org/10.26879/1246
Copyright Society for Vertebrate Paleontology, June 2023
Author biographies
Plain-language and multi-lingual abstracts
PDF version
Submission: 5 October 2022. Acceptance: 3 June 2023.
ABSTRACT
True primates in the Canadian fossil record are known from only the Eocene epoch of southwestern Saskatchewan. They were originally described in the 1980s from two sets of localities: the Lac Pelletier localities (Duchesnean) and the Swift Current Creek (SCC) locality (Uintan). Additional fieldwork and sample sorting were carried out over the last decade. The present study includes a description of new primate dental material from the Swift Current Creek (SCC) locality. We name a new genus and two new species and revise the known primates for both the Lac Pelletier and SCC faunas. The Cypress Hills region of Saskatchewan likely represented a haven for small primates in the Eocene and may have supported an endemic primate fauna.
Jonathan M.G. Perry. Department of Physical Therapy Education, College of Health Sciences–Northwest, Western University of Health Sciences, 2665 S. Santiam Hwy., Lebanon, Oregon, 97355, U.S.A. jperry@westernu.edu
Alex R. Dutchak. Department of Geoscience, University of Calgary, Calgary, Alberta, T2N 1N4, Canada. alexander.dutchak@ucalgary.ca
Jessica M. Theodor. Department of Biological Sciences, University of Calgary, Calgary, Alberta, T2N 1N4, Canada. jtheodor@ucalgary.ca
Keywords: new genus; new species; primate; Eocene; Saskatchewan; omomyid
Final citation: Perry, Jonathan M.G., Dutchak, Alex R., and Theodor, Jessica M. 2023. New primates from the Eocene of Saskatchewan, Canada: Revision of the primates from the Cypress Hills Formation with description of new taxa. Palaeontologia Electronica, 26(2):a20.
https://doi.org/10.26879/1246
palaeo-electronica.org/content/2023/3867-new-eocene-primates-of-saskatchewan
Copyright: June 2023 Society of Vertebrate Paleontology.
This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
creativecommons.org/licenses/by/4.0
https://zoobank.org/15109906-D416-47EE-88A4-752EF23AA436
INTRODUCTION
Context for This Study
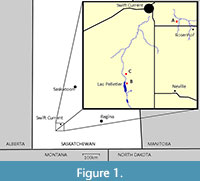 The only true primates (“euprimates”) known from the entire fossil record of Canada comprise a modest sample of omomyid teeth from two areas in southwestern Saskatchewan (Figure 1). They are all from the middle Eocene. They were all described by John E. Storer three decades ago, from a series of roadcuts and hillsides around man-made lakes (Lac Pelletier and Lizard Lake) and a single hillside 50 km to the northeast, near Rosenhof (Storer, 1984, 1990). The first set of localities are considered Duchesnean and the second is Uintan, all part of the Cypress Hills Formation. The primates from the Duchesnean localities were described as belonging to Trogolemur leonardi, Macrotarsius cf. Macrotarsius montanus, and Omomys sp. The primates from the Uintan locality were described as belonging to two unnamed species designated “Omomyidae sp. 1” and “Omomyidae sp. 2”.
The only true primates (“euprimates”) known from the entire fossil record of Canada comprise a modest sample of omomyid teeth from two areas in southwestern Saskatchewan (Figure 1). They are all from the middle Eocene. They were all described by John E. Storer three decades ago, from a series of roadcuts and hillsides around man-made lakes (Lac Pelletier and Lizard Lake) and a single hillside 50 km to the northeast, near Rosenhof (Storer, 1984, 1990). The first set of localities are considered Duchesnean and the second is Uintan, all part of the Cypress Hills Formation. The primates from the Duchesnean localities were described as belonging to Trogolemur leonardi, Macrotarsius cf. Macrotarsius montanus, and Omomys sp. The primates from the Uintan locality were described as belonging to two unnamed species designated “Omomyidae sp. 1” and “Omomyidae sp. 2”.
Although this sample is small and represents two narrow windows in time, it is very important. Whereas there is a rich record of plesiadapiforms from Canada (e.g., Scott and Fox, 2005; Fox, 2011; Boyer, et al., 2012; Chester, et al., 2015; Scott, et al., 2016; Scott, et al., 2023), the Saskatchewan sample represents the only record of fossil euprimates from Canada. These localities are, therefore, the northern tip of the historical distribution of nonhuman true (Miller, et al., 2023) primates in the Americas. The ecological conditions (climate, competitors, predators, food) that permitted such primates to exist in the Eocene of Saskatchewan might also reveal (as they change) the conditions that led to their extinction.
Additional fieldwork, microsample sorting, and comparisons to newly discovered/described relatives have enabled us to contribute new elements to this primate sample and to provide new comparisons to other Eocene primates. The four genera of primates are two trogolemurin omomyids (three species) and two omomyine omomyids (two species).
Institutional Acronyms
AMNH, American Museum of Natural History, New York, New York, U.S.A.; CM, Carnegie Museum of Natural History, Pittsburgh, Pennsylvania, U.S.A.; LACM, Natural History Museum of Los Angeles County, Los Angeles, California, U.S.A.; RSM, Royal Saskatchewan Museum, Regina, Saskatchewan, Canada; UCM, University of Colorado Museum of Natural History, Boulder, Colorado, U.S.A.; UCMP, University of California Museum of Paleontology, Berkeley, California, U.S.A.; UM, University of Michigan Museum of Paleontology, Ann Arbor, Michigan, U.S.A.; USNM, United States National Museum of Natural History (Smithsonian), Washington, District of Columbia, U.S.A.; YPM, Yale Peabody Museum of Natural History, New Haven, Connecticut, U.S.A.
History of Fieldwork
Initial fieldwork at the localities was carried out by Loris Russell and colleagues in the 1930s, continuing into the late 1960s (Russell, 1965a, 1965b). Subsequent fieldwork in the 1970s and 1980s was carried out by the Royal Ontario Museum and the Royal Saskatchewan Museum (RSM, led by John Storer) (Storer, 1984). RSM field techniques included an innovation: power dry screening. A motor-driven machine sorted dry matrix into three size fractions, enabling the paleontologists to isolate the size fraction most appropriate for recovering small mammal teeth. Very few specimens in articulation or even mandibles with multiple teeth are present in these faunas. The power dry screening method created a high-yield matrix in the field, increasing the efficiency of micro vertebrate sorting in the lab. Unfortunately, it also lost specimens smaller than 1.5 mm and potentially increased specimen breakage due to the vigorous nature of the sorting mechanism.
We carried out fieldwork in the Cypress Hills Formation from 2014-2017, targeting the Swift Current Creek (SCC) and Lac Pelletier localities. Our initial forays were made possible with the assistance of John Storer who kindly showed the locations of the original sites. Currently, all Lac Pelletier localities are significantly overgrown, though they still yield fossils even upon cursory inspection. The SCC locality was inspected in 2014 and proved to be fossil rich.
After permissions were secured from landowners, the SCC locality was reopened as a quarry in the summer of 2015 with a modest-sized field crew (4-8 people). Another full field season was carried out at the quarry in 2017 when a new locality of potentially the same age was discovered across the creek. The SCC locality was found to have two fossil-rich conglomeritic sandstones superimposed with a small layer of clean, fossil-poor sand in between. We took care to excavate the two fossil horizons separately. We hand-quarried the loose material and collected conglomeritic blocks for later acid-dissolution. Acid preparation took place in JMT’s lab at the University of Calgary and at JMGP’s lab at Johns Hopkins University. We used 10% acetic acid over long periods of time (1-2 weeks). This broke up the weak limestone matrix and enabled identification of larger fossil fragments. Subsequent wet screening (with a 0.5 mm minimum hole diameter) yielded a rich matrix that was then dried and sorted independently in the two labs. In parallel, we acquired from the Royal Saskatchewan Museum the remaining power screen matrix that Storer had collected from the SCC, which is a composite of both conglomeritic layers. We sorted the matrix to identify fossils. Newly discovered specimens were compared to cast collections at Johns Hopkins University and the University of Calgary. The present work describes the primates that were recovered by our team.
TROGOLEMURINS FROM SASKATCHEWAN
Previous Work
The previously known specimens of Trogolemur from Saskatchewan consist of nine teeth from Lac Pelletier, three of which are complete upper molars and two of which are fragmentary upper molars. These were not originally recognized as Trogolemur but were described as a new species of plesiadapiform, Phenacolemur leonardi (see Storer, 1990, figure 1a and 1b); Storer also referred five lower teeth from the Lac Pelletier sample to Trogolemur sp. (see Storer, 1990), but one of these, RSM P1899.1001, is actually an omomyine (see below). Later he referred all of the teeth described as Phenacolemur leonardi to Trogolemur (e.g., Storer and Tokaryk, 1992; Storer, 1996). Emry (1990) described the first upper molars of Trogolemur. He referred three complete upper molars (as well as some fragmentary upper molars and lower dentition) from Elderberry Canyon, Nevada, to the then only species of Trogolemur, T. myodes. Trogolemur myodes was originally described from a mandibular specimen by Matthew (1909). Additional specimens of T. myodes include a small collection of upper molars from the Bridgerian of the Green River Basin, curated at the University of Michigan (Gunnell, 1995). These include the only known associated uppers, UM 31201, a partial right maxilla with M2 and M3. Beard and colleagues described two additional species of Trogolemur, T. amplior and T. fragilis, but these are known from lower teeth alone (Beard, et al., 1992). Gunnell (1995) compared Trogolemur myodes UM 31201 to an additional upper molar attributed to the same species (UM31557, a suspected M1), and established some criteria for distinguishing M1 from M2 in Trogolemur. These include–for the M1–a longer buccal margin, a more mesiodistally oriented postprotocingulum, and a narrower hypocone shelf, all giving the tooth a more squared-off look.
Emry (1990) compared the upper molars of Trogolemur from Elderberry Canyon to the complete upper molar row of Strigorhysis, an omomyid of some similar morphology. From this comparison, Emry identified USNM 417391 as an M3 (Emry, 1990, fig. 6G), and USNM 417396 and 417390 as M2s (Emry, 1990, figs. 6H and 6I). Storer (1990) identified one of the complete upper molars in the Lac Pelletier sample as an M1 (RSM P1899.1007), one as an M2 (RSM P1899.1016), and one as an M3 (RSM P1899.1014). Based on Gunnell’s criteria and comparison with other omomyids, especially Strigorhysis, we reevaluate these identifications to tooth locus. In general, omomyid M2s are broader (buccolingual dimension) than M1 and M3; this makes M2 more rectangular and M1 and M3 squarer. Often the protocone lobe is very large in M2, more so than in the other upper molars. In most omomyids, the M3 is easily distinguished from the other molars by the reduction of the distal margin, with the metacone and hypocone being small. By these criteria, Emry’s identifications are correct, except that USNM 417396 is probably an M1, not an M2. The protocone lobe of this tooth is smaller than in USNM 417390 (an undoubted M2), and the postprotocingulum runs almost directly distally from the protocone. Furthermore, USNM 417396 is much narrower than USNM 417390 (2.3 mm versus 2.6 mm), though not shorter (both 1.6 mm).
We think Storer (1990) correctly identified RSM P1899.1014 as an M3; however, the M2 (RSM P1899.1016) is probably also an M3. This is based on the much smaller metacone than paracone and the apparent reduction of the distal margin of the tooth. The hypocone shelf is slightly larger in RSM P1899.1016 compared to the same in RSM P1899.1014, and the tooth is slightly larger overall, but this is consistent with intraspecific variation–often greater in the third molar than in other molar loci (e.g., Gingerich and Winkler, 1979). RSM P1899.1007, identified as an M1, is more likely an M2 based on comparison to Gunnell’s (1995) sample from the Green River Basin. This tooth is quite broad and has an expanded protocone lobe, greatly resembling the M2 in UM 31201 and also the M2 in USNM 417390. It does not closely resemble USNM 417396 (here identified as M1): RSM P1899.1007 has a much larger protocone lobe than USNM 417396 and the base of its paracone projects more buccally than that of its metacone. These differences are more consistent with RSM P1899.1007 being an M2 than an M1. Thus, no complete M1 for Trogolemur is yet known from Saskatchewan. It is likely, however, that RSM P1899.1005 and RSM P1899.1006, known only from the lingual portion of the tooth are correctly identified as M1 based on size (see Table 1).
The lower teeth in the Storer Lac Pelletier collection include RSM P1899.1004, a fragmentary right m1 or m2, and RSM P1899.1002, a complete left m1 or m2. By comparison to specimens of Trogolemur myodes with associated lower molars (AMNH 12599, Matthew, 1909, fig. 5; USNM 417356, Emry, 1990, fig. 6A-C; and USNM 417355, Emry, 1990, fig. 6D-F), RSM P1899.1002 appears to be an m1. In the m1 of Trogolemur, the mesial terminus of the cristid obliqua is more lingual than it is in many omomyids; this causes the cristid obliqua to meet the base of the metaconid rather than the protocristid or the base of the protoconid. This configuration (seen in some other omomyoids, for example Teilhardina) leaves a small trough (hypoflexid) mesial to the cristid obliqua but distal to the protoconid and it makes for a discontinuous or stepped postvallid. The cristid obliqua in the m2 terminates more buccally and meets the protocristid rather than the metaconid. RSM P1899.1002 has the m1 configuration of the cristid obliqua. RSM P1899.1003, identified as a right m1 or m2, also has these features, so might very well be an m1. The m1 hypoconulid is slightly less well-developed than the m2 hypoconulid in AMNH 12599; in USNM 417355 and USNM 417356, the m1 hypoconulid is markedly less well-developed than in the m2. RSM P1899.1002 has a well-developed hyoconulid, but clearly hypoconulid development is variable in the m1 of Trogolemur. Also, both RSM teeth, as well as the fragmentary RSM P1899.1004, have a prominent paraconid, consistent with them being first molars.
Comparing the Trogolemur leonardi m1 sample to that of T. myodes yields a few differences. Most notable is the greater degree of distal offset in the metaconid in the former. This offset is great enough that a deep transverse trough exists distal to the protoconid and paraconid, but mesial to the metaconid. Presumably the hypocone slides along this trough during mastication; the mesiodistal distance between the tip of the protocone and the tip of the hypocone is about the same as the distance between this trough and the deepest part of the talonid basin, at about a millimeter. There is a slight trough in some m1s of T. myodes (e.g., AMNH 12599), but it is less developed than in T. leonardi–especially RSM P1899.1002. Trogolemur leonardi has a much slighter buccal cingulid on the talonid than in T. myodes; whereas both species have approximately the same degree of development for the buccal cingulid on the trigonid. Otherwise, the lower m1s of T. leonardi are very similar to those of T. myodes.
New Discoveries–Trogolemur
New teeth of Trogolemur were discovered in matrix originally collected and power screened by John Storer and colleagues from the Uintan Swift Current Creek locality, Saskatchewan, Canada. These teeth were recovered via sorting using a stereo microscope. The first new Trogolemur tooth, RSM P3450.1466, appears to be a left M1. The second, RSM P3450.1467, is a left m3. The third, RSM P3450.1468, is a right m3.
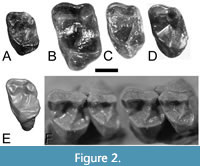 The new Trogolemur upper tooth, RSM P3450.1466, is small compared to the conspecifics in the Lac Pelletier sample (see Figure 2 and Table 1). The tooth is somewhat worn, and slightly broken on the distal side of the metacone. Apical wear has exposed dentine lacunae on the metacone and paracone. Regardless, morphological differences between it and the M1s of Trogolemur myodes are apparent. The clearest of these is a lack of buccal notch on the contour of the tooth, in contrast to USNM 417396 and UM 31557. Instead, the buccal margin in the new tooth is relatively flat. The new tooth is the only upper tooth of Trogolemur from the Uintan SCC locality, and all the other Trogolemur upper teeth from Saskatchewan are from the Duchesnean Lac Pelletier localities. The latter are approximately 30% larger in linear dimensions.
The new Trogolemur upper tooth, RSM P3450.1466, is small compared to the conspecifics in the Lac Pelletier sample (see Figure 2 and Table 1). The tooth is somewhat worn, and slightly broken on the distal side of the metacone. Apical wear has exposed dentine lacunae on the metacone and paracone. Regardless, morphological differences between it and the M1s of Trogolemur myodes are apparent. The clearest of these is a lack of buccal notch on the contour of the tooth, in contrast to USNM 417396 and UM 31557. Instead, the buccal margin in the new tooth is relatively flat. The new tooth is the only upper tooth of Trogolemur from the Uintan SCC locality, and all the other Trogolemur upper teeth from Saskatchewan are from the Duchesnean Lac Pelletier localities. The latter are approximately 30% larger in linear dimensions.
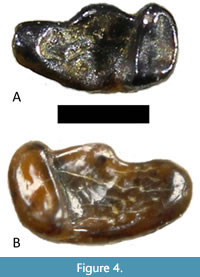
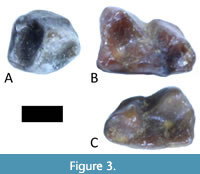 Storer’s (1990) sample of Trogolemur teeth from Lac Pelletier includes several lower teeth as well (Figure 3); however, none of these is an m3. Therefore, we compare the new m3s RSM P3450.1467 and RSM P3450.1468 to the m3 present in Trogolemur myodes AMNH 12599, UM 99703, USNM 417355, USNM 417356, YPM 13523, and UCM 67883. The new teeth are similar in size to each other and represent similar wear stages, the main difference being that the hypoconulid lobe is slightly narrowed in RSM P3450.1467 and this tooth is slightly smaller overall (Figure 4). The new teeth are moderately worn, with the cusp tips worn nearly flat and with some pitting in the talonid and trigonid basins. In less worn specimens, crenulations in the basins are pronounced, but in the new teeth, the high points of the crenulations are nearly obliterated. Except for UCM 67883 (Kelly and Murphey, 2016), the T. myodes sample is uniform in m3 morphology, and the new teeth are somewhat different from these four in the following ways. They are smaller and narrower overall, and the hypoconulid lobe is narrower and more squared off. In the new specimens, the paraconid sits buccal to the metaconid, whereas, in all other specimens, these two cusps are aligned on the lingual side of the tooth (or nearly so).
Storer’s (1990) sample of Trogolemur teeth from Lac Pelletier includes several lower teeth as well (Figure 3); however, none of these is an m3. Therefore, we compare the new m3s RSM P3450.1467 and RSM P3450.1468 to the m3 present in Trogolemur myodes AMNH 12599, UM 99703, USNM 417355, USNM 417356, YPM 13523, and UCM 67883. The new teeth are similar in size to each other and represent similar wear stages, the main difference being that the hypoconulid lobe is slightly narrowed in RSM P3450.1467 and this tooth is slightly smaller overall (Figure 4). The new teeth are moderately worn, with the cusp tips worn nearly flat and with some pitting in the talonid and trigonid basins. In less worn specimens, crenulations in the basins are pronounced, but in the new teeth, the high points of the crenulations are nearly obliterated. Except for UCM 67883 (Kelly and Murphey, 2016), the T. myodes sample is uniform in m3 morphology, and the new teeth are somewhat different from these four in the following ways. They are smaller and narrower overall, and the hypoconulid lobe is narrower and more squared off. In the new specimens, the paraconid sits buccal to the metaconid, whereas, in all other specimens, these two cusps are aligned on the lingual side of the tooth (or nearly so).
 Based on the aforementioned morphological differences between RSM P3450.1466 and Trogolemur myodes, and on the basis of major size differences between RSM P3450.1466 and T. leonardi (e.g., RSM P1899.1005 and RSM P1899.1006), we consider the new tooth to belong to a new species of Trogolemur. The new m3s from SCC are similar in size and morphology to those of T. myodes and are too small to belong to T. leonardi (compare with RSM P1899.1002 and RSM P1899.1003). Given that the new teeth all come from the same Uintan locality and horizon, it is likely that they belong to a new species, distinct from T. leonardi from the Duchesnean Lac Pelletier.
Based on the aforementioned morphological differences between RSM P3450.1466 and Trogolemur myodes, and on the basis of major size differences between RSM P3450.1466 and T. leonardi (e.g., RSM P1899.1005 and RSM P1899.1006), we consider the new tooth to belong to a new species of Trogolemur. The new m3s from SCC are similar in size and morphology to those of T. myodes and are too small to belong to T. leonardi (compare with RSM P1899.1002 and RSM P1899.1003). Given that the new teeth all come from the same Uintan locality and horizon, it is likely that they belong to a new species, distinct from T. leonardi from the Duchesnean Lac Pelletier.
The exceptional m3, UCM 67883, from southwestern Wyoming, is very similar to the new ones described here. Although Kelly and Murphey (2016) described their m3 as being indistinguishable from the prior Trogolemur myodes sample, it has a much more tapered hypconulid lobe than is true of the others (see especially the type specimen of Trogolemur myodes, AMNH 12599, and those from Nevada, USNM 417355 and USNM 417356). The Saskatchewan specimens share this greater degree of tapering. Given variability in third molars, this might not have phylogenetic significance. However, UCM 67883 might belong in the new species and not in Trogolemur myodes.
SYSTEMATIC PALEONTOLOGY
Order Primates Linnaeus, 1758
Superfamily Omomyoidea Trouessart, 1879
Family Omomyidae Trouessart, 1879
Subfamily Anaptomorphinae Cope, 1883
Tribe Trogolemurini Szalay, 1976
Genus Trogolemur Matthew, 1909
Trogolemur leonardi (Storer, 1990)
Type. Right upper second molar, RSM P1899.1007.
Horizon and type locality. Channel sands and gravels of the mid-to-late Eocene age along the east side of Lac Pelletier, southwestern Saskatchewan, Canada (Storer, 1983, 1987, 1996).
Revised diagnosis. Teeth 30-40% larger in linear dimensions than Trogolemur myodes; upper molars broader relative to length, more distinct and larger protocone lobe, smaller metaconule, more distinct lingual cingulum, more reduced metacone on M3; lower molars less exodaenodont with less pronounced buccal cingulum; m3 protoconid and metaconid closer to each other; m3 hypoconulid lobe narrower. Differs from Trogolemur amplior in being significantly smaller; notch buccal to the entoconid and distinct break in cristid obliqua on m1/m2. Differs from Trogolemur fragilis in being significantly larger; m3 protoconid and metaconid closer to each other.
Hypodigm. LM1, RSM P1899.1005; RM1, RSM P1899.1006; LM3, RSM P1899.1014; RM3, RSM P1899.1016; Lm1 or m2, RSM P1899.1002; Rm1 or m2, RSM P1899.1003; Rm1 or m2, RSM P1899.1004; Rm1 or m2, RSM P1900.51.
Trogolemur storeri n. sp.
zoobank.org/99CE821F-1FDA-4FC0-A7CE-AC1B6A84971F
Type. Left upper first molar discovered by K. A. Prufrock in matrix collected by John Storer and crew, RSM P3450.1466 (fig. 2A, 2E).
Horizon and type locality. Interbedded conglomerate and coarse sandstone of Uintan middle Eocene age, Swift Current Creek locality, southwestern Saskatchewan, Canada (Russell, 1965a, 1965b, 1972; Storer, 1978, 1996).
Diagnosis. Approximately 30% smaller in linear dimensions than Trogolemur leonardi; buccal notch much shallower; greater development of lingual cingulum. Differs from Trogolemur myodes in M1 being wider relative to length, more pronounced lingual cingulum, much shallower buccal notch. Differs from Trogolemur amplior in being considerably smaller. Differs from Trogolemur myodes in m3 having much more taper distally, with long and narrow hypoconulid lobe. Differs from Trogolemur fragilis in having a less distinct paraconid on m3 that sits closer to the metaconid, slightly longer and narrower hypoconulid lobe.
Etymology. ‘storeri’ named for John E. Storer who described the first primates from the Swift Current Creek locality and in honor of his extensive contributions to Canadian paleontology.
Hypodigm. Lm3, RSM P3450.1467; Rm3, RSM P3450.1468.
Summary of Changes
The following list summarizes suggested changes to the record for the genus Trogolemur.
- Trogolemur myodes USNM 417396 is an M1, not an M2.
- Trogolemur leonardi RSM P1899.1014 and RSM P1899.1016 are M3s, not M2s.
- Trogolemur leonardi RSM P1899.1007 is an M2, not an M1.
- Trogolemur leonardi RSM P1899.1002, RSM P1899.1003, and RSM P1899.1004, all identified as m1 or m2, are likely m1s.
New records of Trogolemur include a left M1 (RSM P3450.1466), a left m3 (RSM P3450.1467), and a right m3 (RSM P3450.1468). These are described as a new species, Trogolemur storeri. With very little morphological difference between Trogolemur storeri and Trogolemur leonardi apart from size, and with little geographic separation, it seems likely that the former is a phyletic antecedent to the latter.
New Discoveries–Walshina
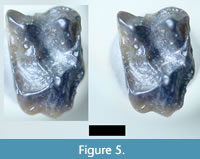 An upper molar resembling Trogolemur leonardi, but approximately 50% larger in linear dimensions, was discovered in the SCC matrix (Figure 5). Slight erosion along the buccal and distal margins has altered the outline of the tooth. Nevertheless, this tooth (RSM P3450.1469) is recognizable as a primate right M1 (or possibly M2). The tooth has a broad basin distal to the protocone; this is distinctive of trogolemurin omomyoids. It greatly resembles CM 15635 (LM1) from the Uintan Badwater Creek, Wyoming. That tooth was originally described as Phenacolemur mcgrewi by Peter Robinson, then allocated to Ignacius by Bown and Rose (Robinson, 1968; Bown and Rose, 1976). Subsequent analysis by López-Torres and colleagues has shifted this material from the Paromomyidae (Plesiadapiformes) to the Trogolemurini (Omomyidae), renaming the relevant material as Walshina mcgrewi (López-Torres, et al., 2018). We provisionally refer the new tooth to Walshina mcgrewi.
An upper molar resembling Trogolemur leonardi, but approximately 50% larger in linear dimensions, was discovered in the SCC matrix (Figure 5). Slight erosion along the buccal and distal margins has altered the outline of the tooth. Nevertheless, this tooth (RSM P3450.1469) is recognizable as a primate right M1 (or possibly M2). The tooth has a broad basin distal to the protocone; this is distinctive of trogolemurin omomyoids. It greatly resembles CM 15635 (LM1) from the Uintan Badwater Creek, Wyoming. That tooth was originally described as Phenacolemur mcgrewi by Peter Robinson, then allocated to Ignacius by Bown and Rose (Robinson, 1968; Bown and Rose, 1976). Subsequent analysis by López-Torres and colleagues has shifted this material from the Paromomyidae (Plesiadapiformes) to the Trogolemurini (Omomyidae), renaming the relevant material as Walshina mcgrewi (López-Torres, et al., 2018). We provisionally refer the new tooth to Walshina mcgrewi.
Given the rarity of primate material at SCC, it is interesting that there are two known species of trogolemurin omomyid at this locality. Thus, diversity of trogolemurins is slightly higher here than in contemporaneous faunas in Wyoming or southern California. Despite the enormous geographic separation, trogolemurins existed simultaneously in all these places. This speaks to the similarity in climate across much of North America in the middle Eocene, and/or to the adaptability of these tiny primates.
OMOMYINES FROM SASKATCHEWAN
Macrotarsius
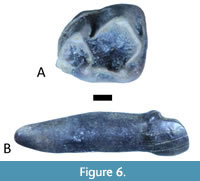 Storer (1990) described two teeth from Lac Pelletier as Macrotarsius cf. M. montanus (see Figure 6). One of these (RSM P1899.1008) he identified as a right lower second premolar. The other (RSM P1899.1009) he identified as a right lower first molar. RSM P1899.1008 is almost certainly not a p2 of Macrotarsius. This tooth is relatively large and robust (crown length = 2.65 mm, width = 2.11 mm, height = 2.16 mm) with a complete, compressed single root (length = 5.26 mm, length of major axis of cross-section = 1.82 mm). The type and only specimen of Macrotarsius montanus (CM 9592) does not have a p2; however, it retains the p2 alveolus. This alveolus is far too small (major axis of cross-section < 1 mm) to accommodate the root of RSM P1899.1008. By contrast, the m1 (RSM P1899.1009) is nearly identical in size to the m1 of CM 9592 (see Table 2). RSM P1899.1008 is much larger than the p2 in Macrotarsius roederi (crown length = 1.85 mm, crown breadth = 1.55 mm, see Kelly, 1990). Also, the p2 in M. roederi has a distinct basal cusp which is lacking in the Saskatchewan tooth. RSM P1899.1008 could be a primate canine, or a primate lower central incisor, or even an anterior tooth of a non-primate mammal. The canine in M. roederi is larger than the p2, but the i1 is unknown (Kelly, 1990).
Storer (1990) described two teeth from Lac Pelletier as Macrotarsius cf. M. montanus (see Figure 6). One of these (RSM P1899.1008) he identified as a right lower second premolar. The other (RSM P1899.1009) he identified as a right lower first molar. RSM P1899.1008 is almost certainly not a p2 of Macrotarsius. This tooth is relatively large and robust (crown length = 2.65 mm, width = 2.11 mm, height = 2.16 mm) with a complete, compressed single root (length = 5.26 mm, length of major axis of cross-section = 1.82 mm). The type and only specimen of Macrotarsius montanus (CM 9592) does not have a p2; however, it retains the p2 alveolus. This alveolus is far too small (major axis of cross-section < 1 mm) to accommodate the root of RSM P1899.1008. By contrast, the m1 (RSM P1899.1009) is nearly identical in size to the m1 of CM 9592 (see Table 2). RSM P1899.1008 is much larger than the p2 in Macrotarsius roederi (crown length = 1.85 mm, crown breadth = 1.55 mm, see Kelly, 1990). Also, the p2 in M. roederi has a distinct basal cusp which is lacking in the Saskatchewan tooth. RSM P1899.1008 could be a primate canine, or a primate lower central incisor, or even an anterior tooth of a non-primate mammal. The canine in M. roederi is larger than the p2, but the i1 is unknown (Kelly, 1990).
The m1 that Storer (1990) described as Macrotarsius cf. M. montanus strongly resembles the m1 of the holotype of M. montanus (CM 9592) and is nearly identical in size (see Table 2). Szalay (1976) suggested that Macrotarsius could be distinguished from similar omomyines (e.g., Ourayia and Hemiacodon) because it possesses a tall crest running distally from the metaconid which sometimes bears cuspules and which raises (or ‘shallows’) the taloned notch. This feature is clearly present in RSM P1899.1009; the crest running distally from the metaconid is tall and the talonid notch is shallow. Although no distinct cuspules are present in the Saskatchewan tooth, this might be because they have been obliterated by wear. Hemiacodon engardae has a smaller m1 (length = 3.84 mm), with more pronounced crenulation, and a much deeper taloned notch (Murphey and Dunn, 2009). In our opinion, the m1 described by Storer (1990) should remain Macrotarsius cf. M. montanus pending the discovery of additional material.
“Omomys”–Sample and Previous Work
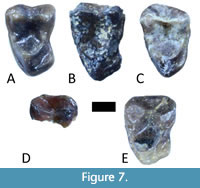 Storer (1990) described a sample of 17 primate teeth from Lac Pelletier as Omomys sp. These include five upper molars (one fragmentary), 10 lower molars (two fragmentary), and two lower premolars (Figure 7 and Figure 8). Storer (1984) also described three molars from Swift Current Creek as “Omomyidae” (genus and species unspecified). These are two right m3s (RSM P1654.343 and RSM P1654.344) described as Omomyidae sp. 1 and a right M3 (RSM P1654.470) described as Omomyidae sp.2 (Figure 9). Comparison with other mammals in the local faunas suggests that the M3 belongs to an artiodactyl.
Storer (1990) described a sample of 17 primate teeth from Lac Pelletier as Omomys sp. These include five upper molars (one fragmentary), 10 lower molars (two fragmentary), and two lower premolars (Figure 7 and Figure 8). Storer (1984) also described three molars from Swift Current Creek as “Omomyidae” (genus and species unspecified). These are two right m3s (RSM P1654.343 and RSM P1654.344) described as Omomyidae sp. 1 and a right M3 (RSM P1654.470) described as Omomyidae sp.2 (Figure 9). Comparison with other mammals in the local faunas suggests that the M3 belongs to an artiodactyl.
 The SCC m3s resemble each other very closely and are very likely from the same species. Comparing the two SCC molars to the Omomys sp. molars from Lac Pelletier is difficult because the samples have no definite overlapping tooth loci. Therefore, we compare the m2s to the m3s by looking at serial changes in morphology in omomyids with m2/m3 associations. The m2s from Lac Pelletier and the m3s from SCC do follow serial trends. The m2s and m3s are similar in breadth, but the m3s are longer, consistent with these proportions on associated molars of other omomyids (e.g., Omomys). The m2s have relatively longer shearing crests and a deeper talonid basin than the m3s. The m3s have blunter and more rounded cusps than the m2s. These serial differences are consistent with the m2s from Lac Pelletier being from the same species as the m3s from SCC. This conclusion is further supported by new discoveries of omomyid upper molars from the SCC matrix (see below); these include teeth that overlap in loci with “ Omomys sp.” previously described from Lac Pelletier.
The SCC m3s resemble each other very closely and are very likely from the same species. Comparing the two SCC molars to the Omomys sp. molars from Lac Pelletier is difficult because the samples have no definite overlapping tooth loci. Therefore, we compare the m2s to the m3s by looking at serial changes in morphology in omomyids with m2/m3 associations. The m2s from Lac Pelletier and the m3s from SCC do follow serial trends. The m2s and m3s are similar in breadth, but the m3s are longer, consistent with these proportions on associated molars of other omomyids (e.g., Omomys). The m2s have relatively longer shearing crests and a deeper talonid basin than the m3s. The m3s have blunter and more rounded cusps than the m2s. These serial differences are consistent with the m2s from Lac Pelletier being from the same species as the m3s from SCC. This conclusion is further supported by new discoveries of omomyid upper molars from the SCC matrix (see below); these include teeth that overlap in loci with “ Omomys sp.” previously described from Lac Pelletier.
 The RSM Lac Pelletier collection includes ten lower molars that Storer (1990) assigned to Omomys sp. These include eight complete tooth crowns and two isolated trigonids. Some of these are identified as m1, some as m2, some as “m1 or m2”. In order to assess the identities of the ambiguously-assigned molars, we made comparisons to omomyids that have m1/m2 associations (e.g., Omomys, Chumashius, Dyseolemur). The main differences between m1 and m2 appear to be that m1 has a narrower talonid, a larger and more mesially-shifted paraconid, and a more buccally-shifted hypoconulid with a notch just lingual to that cusp to accommodate the m2 paraconid.
The RSM Lac Pelletier collection includes ten lower molars that Storer (1990) assigned to Omomys sp. These include eight complete tooth crowns and two isolated trigonids. Some of these are identified as m1, some as m2, some as “m1 or m2”. In order to assess the identities of the ambiguously-assigned molars, we made comparisons to omomyids that have m1/m2 associations (e.g., Omomys, Chumashius, Dyseolemur). The main differences between m1 and m2 appear to be that m1 has a narrower talonid, a larger and more mesially-shifted paraconid, and a more buccally-shifted hypoconulid with a notch just lingual to that cusp to accommodate the m2 paraconid.
The Lac Pelletier sample includes two correctly-identified m1s: RSM P1899.1019 and.1020 (both left). RSM P1899.1019 has a narrow talonid, a large and mesially-shifted paraconid, and a buccally-shifted hypoconulid with a notch lingual to it. RSM P1899.1020 has a narrow talonid, but the area surrounding and including the hypoconulid is broken away. There are two teeth identified as m1 (RSM P1899.1022, right; RSM P1899.1024, left) which are probably from that locus based on overall morphology. RSM P1899.1022 has a large paraconid, although it is not very mesially shifted; its hypoconulid is very buccally-shifted by the notch lingual to it is slight. RSM P1899.1024 has these same features, but its paraconid is more mesially shifted than is the case in RSM P1899.1022.
RSM P1899.1021, identified as a left m2, appears to be an m1 based on the mesial shift of its paraconid and the substantial notch lingual to the hypoconulid; however, the hypoconulid is not very buccally shifted nor is the trigonid much narrower than the talonid. A trigonid identified as a right m1 or m2 (RSM P1899.1026) appears to be that of an m2.
RSM P1899.1023 is a correctly-identified left m2; RSM P1899.1025 and RSM P1899.1027 are correctly-identified right m2s. All three possess a trigonid that is roughly equal to the talonid in breadth, a small and more distally situated paraconid, a hypoconulid lying at about the buccolingual mid-point of the tooth, and a lack of notch lingual to the hypoconulid. RSM P1899.1027 is very worn, but nevertheless possesses these m2 features.
The trigonid that Storer (1990) identified as a right m2 (RSM P1899.1028) could be an m3; it compares favorably with the m3s from SCC (especially RSM P1654.344), but there is no way to be sure without the talonid.
The upper molars of Omomys sp. from Lac Pelletier consist of a left molar described by Storer (1990) as an M1 (RSM P1899.1010), two left molars described by Storer (1990) as M2s (RSM P1899.1011 and RSM P1899.1012), a left molar described as an M3 (RSM P1899.1013), and the buccal part of a right M1 or M2 bearing the paracone and metacone (RSM P1899.1015). In omomyines (e.g., Omomys carteri YPM 11854), M1 typically has the following: a more convex mesial margin, a longer buccal side, a more pronounced buccal notch, a more extensive base of the paracone and parastylar shelf, a more concave distal margin, and a more extensive hypocone. Usually the M2 is broader, with an expanded protocone lobe. In these respects, RSM P1899.1010 and RSM P1899.1012 are more like an M2, whereas RSM P1899.1011 is more like an M1. RSM P1899.1010 is very worn and therefore difficult to assign with any confidence to either locus; nevertheless, the outline of the tooth demonstrates very little development of either a buccal or distal notch, suggesting it is an M2, contrary to the original identification (Storer, 1990). RSM P1899.1012 has a lot of apical wear but shows the same, reinforcing its original identification as an M2. RSM P1899.1011 has very little wear and is clearly more like an M1, with a distinct buccal notch and distal notch, as well as a more convex mesial margin; it also has a more well developed hypocone than either RSM P1899.1010 or RSM P1899.1012.
RSM P1899.1013 has essentially no hypocone (and but a small hypocone shelf), a small metacone, and a buccal shelf that slopes inward distally; these are all hallmarks of an M3. RSM P1899.1015 has a buccal notch that is very shallow, intermediate in depth between RSM P1899.1011 and RSM P1899.1012; this makes it impossible to discern whether this tooth fragment is an M1 or an M2.
“Omomys”–New Discoveries
We discovered two new upper molars of omomyids in the SCC unsorted matrix that was originally collected by John Storer. They consist of a left M1 (RSM P3450.1470) and a right M3 (RSM P3450.1471). They are similar in several respects to RSM P1899.1011 and RSM P1899.1013, respectively, as follows. When compared to the previously described M1 (RSM P1899.1011), the new M1 (RSM P3450.1470) has similar hypocone development and a similarly sized distal notch, but the paracone is shifted quite far distally and the length of the entire parastylar region is less. There is a continuous lingual cingulum in RSM P3450.1470, but it is discontinuous in RSM P1899.1011. The new M3, RSM P3450.1471, is very similar to RSM P1899.1013, but with a slightly shorter trigon basin. Also, these M3s differ in their degree of lingual cingulum development (see below). The new tooth is broken away buccal to the paracone and metacone, so no comparison of that region is possible.
Sufficient differences exist between the omomyid sample from Saskatchewan and known species of omomyids to warrant the description of a new genus and species for it. Comparisons to several other omomyid species are made below.
SYSTEMATIC PALEONTOLOGY
Order Primates Linnaeus, 1758
Superfamily Omomyoidea Trouessart, 1879
Family Omomyidae Trouessart, 1879
Subfamily Omomyinae Trouessart, 1879
Tribe Omomyini Trouessart, 1879
Saskomomys, new genus
zoobank.org/3CD70AC7-2ECE-406B-BB8E-E480D644F963
Type species. Saskomomys lindsayorum, new species. Figure 10. Table 2 and Table 3.
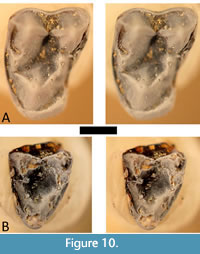 Diagnosis. Differs from Omomys in being smaller in occlusal area of all compared teeth and in the following: p4 with indistinct metaconid, crest emitting from protoconid buccally oriented, lingually-seated paraconid, and short talonid that rises quite dorsally at its distal end; m1 and m2 narrower; m3 broader, with more rounded cusps, with paraconid located centrally; M1 less well-developed hypocone shelf; M2 more convex mesial margin and deeper notch at distal margin; M3 less triangular (i.e., more extensive hypocone shelf), relatively less extensive expansion of the base of the protocone lingually, lingual cingulum continuous or nearly so. Differs from Chumashius in having no distinct metaconid on p4, crest running from protoconid runs buccally rather than distobuccally, less lingually projecting paraconid; m1 and m2 narrower, less well-developed buccal cingulid; m3 hypoconulid lobe more isolated (i.e., constricted at its mesial end), trigonid longer at lingual side and more rounded mesially; M2 narrower (especially at protocone), greater extent of hypocone, more distinct conules, more distinct buccal cingulum. Differs from Dyseolemur in having less basal inflation and crenulation on the lower molars and in lacking a metastylid and the fovea separating the metastylid from the metaconid; m3 has more isolated hypoconulid lobe; M1 and M2 have smaller hypocone, conules, and buccal cingulum, with less-distinct crests emitting from the conules lingually; M3 has less-distinct buccal cingulum and is more triangular in outline (narrower at protocone than at buccal cusps). Differs from Ekwiiyemakius in having a shorter and broader p3 and m3; more constriction mesial to the hypoconulid lobe on m3; M1 buccal notch is more pronounced (deeper and narrower); M1 and M2 trigon basin and hypocone shelf are broader; M3 is longer (mesiodistal dimension).
Diagnosis. Differs from Omomys in being smaller in occlusal area of all compared teeth and in the following: p4 with indistinct metaconid, crest emitting from protoconid buccally oriented, lingually-seated paraconid, and short talonid that rises quite dorsally at its distal end; m1 and m2 narrower; m3 broader, with more rounded cusps, with paraconid located centrally; M1 less well-developed hypocone shelf; M2 more convex mesial margin and deeper notch at distal margin; M3 less triangular (i.e., more extensive hypocone shelf), relatively less extensive expansion of the base of the protocone lingually, lingual cingulum continuous or nearly so. Differs from Chumashius in having no distinct metaconid on p4, crest running from protoconid runs buccally rather than distobuccally, less lingually projecting paraconid; m1 and m2 narrower, less well-developed buccal cingulid; m3 hypoconulid lobe more isolated (i.e., constricted at its mesial end), trigonid longer at lingual side and more rounded mesially; M2 narrower (especially at protocone), greater extent of hypocone, more distinct conules, more distinct buccal cingulum. Differs from Dyseolemur in having less basal inflation and crenulation on the lower molars and in lacking a metastylid and the fovea separating the metastylid from the metaconid; m3 has more isolated hypoconulid lobe; M1 and M2 have smaller hypocone, conules, and buccal cingulum, with less-distinct crests emitting from the conules lingually; M3 has less-distinct buccal cingulum and is more triangular in outline (narrower at protocone than at buccal cusps). Differs from Ekwiiyemakius in having a shorter and broader p3 and m3; more constriction mesial to the hypoconulid lobe on m3; M1 buccal notch is more pronounced (deeper and narrower); M1 and M2 trigon basin and hypocone shelf are broader; M3 is longer (mesiodistal dimension).
Etymology. Prefix ‘Sask–’ in reference to the distribution of the taxon in the province of Saskatchewan. Suffix ‘–omomys’ (Greek, “shoulder-mouse”) refers to omomyoid primates generally.
Saskomomys lindsayorum n. sp.
zoobank.org/4E722957-0808-4D79-9DAB-172C135C186B
Type. Right lower third molar collected by John Storer and crew, RSM P1654.343 (Figure 9A).
Horizon and type locality. Interbedded conglomerate and coarse sandstone of Uintan middle Eocene age, Swift Current Creek locality, southwestern Saskatchewan, Canada (Russell, 1965a, 1965b, 1972; Storer, 1978, 1996).
Diagnosis. As for the genus.
Etymology. ‘lindsayorum’ named for the Lindsay family of Saskatchewan (Brendan and Nadine) who graciously welcomed us to carry out fieldwork on their land where the fossiliferous beds of the Swift Current Creek site are located.
Hypodigm. All material previously described as “Omomyidae, sp. 1” from the Uintan Swift Current Creek locality: Rm3, P1654.343; Rm3, P1654.344. New material from the SCC locality: LM1, RSM P3450.1470; and RM3, RSM P3450.1471. All material previously described as “ Omomys sp.” from the Duchesnean Lac Pelletier localities: LM1, RSM P1899.1011 (originally identified as M2); LM2, RSM P1899.1010 (originally identified as M1); LM2, RSM P1899.1012; RM1 or M2, RSM P1899.1015; LM3, RSM P1899.1013; Lp3, RSM P1899.1017; Lp4, RSM P1899.1001; Rp4, RSM P1899.1018; Lm1, RSM P1899.1019; Lm1, RSM P1899.1020; Lm1, RSM P1899.1021 (originally identified as m2); Lm1, RSM P1899.1024; Rm1, RSM P1899.1022; Lm2, RSM P1899.1023; Rm2, RSM P1899.1025; Rm2 trigonid, RSM P1899.1026 (originally identified as m1 or m2); Rm2 trigonid, RSM P1899.1027; Rm2 trigonid, RSM P1899.1028.
Discussion
The material described by Storer (1984, 1990) as “Omomyidae sp. 1” from Swift Current Creek and the material he described as “ Omomys sp.” from Lac Pelletier differ in several respects from any known omomyid and are probably from the same genus. The new M1 (RSM P3450.1470) from SCC can be compared with the M1 from Lac Pelletier (RSM P1899.1011). In most respects, these teeth are very similar. In RSM P3450.1470, the mesial margin is more curved and the hypocone is slightly smaller than in RSM P1899.1011. Also, the lingual cingulum is continuous across the base of the protocone in the SCC tooth, and its buccal cingulum is more pronounced. Furthermore, the length and breadth of the tooth are about 17% greater than those of the Lac Pelletier tooth (see Table 3). With only two teeth to compare, and with only slight differences, it seems unreasonable to conclude that they are different species, despite the temporal separation.
The new M3 (RSM P3450.1471) from SCC can be compared with the M3 from Lac Pelletier (RSM P1899.1013). These teeth are similar in size; the SCC tooth is broken at the buccal end, so the total breadth is an underestimate. The teeth are very similar morphologically in the preserved parts, differing only in the lesser development of a hypocone shelf in the SCC tooth.
Based on overall similarity between the upper molars, the “ Omomys sp.” sample from Lac Pelletier (Storer, 1990), the two m3s from Swift Current Creek described as “Omomyidae sp. 1” (Storer, 1984), and the new teeth herein (RSM P3450.1470 and RSM P3450.1471) are all described here as part of a single new species, Saskomomys lindsayorum. Future discoveries might well provoke taxonomic splitting of Duchesnean sample from the Uintan sample, but this would require additional teeth of overlapping loci to assess quantitative variation.
The Saskatchewan material most closely resembles omomyids of the tribe Omomyini, namely Omomys, Chumashius, and Ekwiiyemakius (e.g., Tornow, 2008; Atwater and Kirk, 2018), in the following: lack of crenulation in molars, relatively small conules, broad rounded basins, relatively blunt crests, paraconid distinct and shifted lingually, small and buccolingually restricted hypoconulid lobe on m3, simple p4. Saskomomys is more similar to Omomys in having subequally sized upper molars, whereas in Chumashius the M3 is much smaller than M2. Like Omomys and Chumashius, Saskomomys has more basal inflation of the cusps generally than does Steinius. The upper molars of Steinius are mesiodistally narrow relative to those of Saskomomys, and the M3 is much smaller than the M2 in Steinius. In terms of upper molar size proportions (M1:M2:M3), Saskomomys is most like Omomys among Omomyini (Tornow, 2008). The upper molars of Saskomomys are similar in overall size and proportion to those of Ekwiiyemakius (see Atwater and Kirk, 2018); in the former, the trigon basin is broader, as is the hypocone shelf (projecting less from the rest of the tooth), the buccal notch is deeper in M1, and the M3 is longer.
Storer (1984) suggested that the m3s from SCC resemble Chumashius in size and occlusal pattern. We agree that the m3s do closely resemble those of Chumashius balchi (e.g., LACM 1392). However, differences include a much smaller and more separate hypoconulid lobe on the Saskatchewan specimens as well as a longer trigonid lingually–the trigonid in the m3s from Saskatchewan has a very rounded mesial margin. The SCC molars differ in many ways from the m3 of Omomys carteri (e.g., YPM 11805 and 13209); foremost, the Saskatchewan teeth are more rounded (and broad) overall and Omomys carteri is 15-30% larger (linear measurements). Furthermore, the paraconid is centrally located in the SCC m3s, whereas it is lingually located in Omomys. The SCC m3s resemble some members of the tribe Washakiini (e.g., Dyseolemur pacificus, LACM 1395) even more than they do Chumashius, although differences include a more restricted hypoconulid lobe in the SCC teeth, a less blade-like postmetaconid cristid (without the distinct fovea seen in Dyseolemur), less crenulation overall (although the apparent crenulation is influenced by wear) and less basal inflation of the cusps. As in both Chumashius and Dyseolemur, the SCC molars have a small fovea distobuccal to the posthypoconid cristid. Compared to Ekwiiyemakius, the m3 in Saskomomys is broad relative to its length, and there is a more pronounced constriction between the hypoconulid lobe and the rest of the tooth.
The m1 and m2 of the Lac Pelletier Omomys sp. sample greatly resemble those tooth loci in Omomys carteri. The former are slightly narrower than the latter (talonid breadth:tooth length of 0.80 versus 0.86–see Table 2). Any small differences between these species are swamped by intraspecific variation. This is consistent with omomyids generally in that these tooth loci are highly conservative within broad taxa. These teeth are different from those in Chumashius by having a less well-developed buccal cingulid and by being slightly narrower overall (talonid breadth:tooth length for Chumashius is 0.87). The m1 and m2 in Saskomomys are very similar to those of Ekwiiyemakius with the only clear differences being overall blunter crests and lesser development of a buccal cingulum in the former. Dyseolemur is quite different in the m1 and m2, having much more basal inflation and having a small fovea that separates the metaconid from a cuspule lying distolingual to it. The m2 of Utahia is broader and the talonid, especially, is quite short; the tooth is more crenulated and the cristid obliqua dips quite far ventrally before ascending to meet the trigonid. In general, all other omomyids are quite different from the Lac Pelletier m1 and m2 and comparisons to them are not worth listing here (Tornow, 2008).
Premolars are more diagnostic for omomyids generally (Gunnell and Rose, 2002; Gunnell, et al., 2008; Morse, et al., 2019). In the Lac Pelletier sample, there is one right p4 (RSM P1899.1018), one left p4 (RSM P1899.1001), and one left p3 (RSM P1899.1017). Note that RSM P1899.1001 was identified by Storer (1990) as a Trogolemur left fourth premolar. However, this tooth is 1.54 mm long, much closer to the 1.60 mm long RSM P1899.1018 than to the p4 of T. myodes (1.34 mm). It is more likely that RSM P1899.1001 belongs with the Omomys sp. sample and it is treated as such here. The right p4 is very worn, so the following is based on the relatively unworn left p4. The p4 more greatly resembles that of Dyseolemur than it does that of Omomys or that of Chumashius in lacking a distinct metaconid and in having a relatively short talonid that rises dorsally toward its distal aspect. In place of a metaconid, the Lac Pelletier p4 has a shelf that runs lingually from the preprotocristid. It does have a distinct paraconid that projects lingually, though not to the degree seen in Chumashius. The crest running down from the protoconid is very buccally oriented, much more so than in Omomys or Chumashius and more like the condition in Dyseolemur. The p3 conforms in most respects to this morphology; unfortunately, it is somewhat worn. It is shorter and broader than that in Ekwiiyemakius. Thus, the omomyine from Saskatchewan combines a unique m3 morphology with a lower premolar morphology somewhat like Dyseolemur and a m1/m2 morphology somewhat like Omomys carteri.
The upper molar sample from Saskatchewan is small and, as with the lower molar sample, unassociated. The two teeth identified as third upper molars–one from Lac Pelletier and the other from SCC–are very certainly M3s and resemble each other to a great degree. They differ from each other in that the preprotocone crista and postprotocone crista are more linear (less curved) in the SCC tooth, accompanied by the fact that the lingual side of the SCC tooth is shorter and the hypocone crest is slighter. The two M2 teeth differ from each other in similar ways: the one from SCC has straight crests on either side of the protocone whereas the Lac Pelletier tooth has more curved crests. The hypocone crest or shelf is also slighter on the SCC tooth. Furthermore, the SCC M2 has a more “bent” look to it, with a more convex mesial margin and a more concave distal margin. These differences, albeit small, might constitute a species-level difference. However, with such a small sample of overlapping teeth, we cannot substantiate such a claim.
The lingual cingulum on the upper molars appears to have variable degrees of development in different individuals. This is unusual for omomyids generally and the lingual cingulum is sometimes used as a diagnostic character (e.g., Atwater and Kirk, 2018). The new M1 from Swift Current Creek (RSM P3450.1470) has a prominent, continuous lingual cingulum. However, the M1 from Lac Pelletier (RSM P1899.1011) has a clear break in the lingual cingulum at the protocone, and this is a relatively unworn tooth, so is not likely to have been worn away. The new M3 from SCC (RSM P3450.1470) has a continuous lingual cingulum, although the tooth is somewhat eroded. By contrast, the M3 from Lac Pelletier (RSM P1899.1013) has a discontinuous lingual cingulum similar in condition to the M1 from SCC. Of the two complete M2s of Saskomomys (both from Lac Pelletier), one has a clearly continuous lingual cingulum (RSM P1899.1012), whereas the other (RSM P1899.1010) has a discontinuous one. Thus, the temporal difference in the sample does not match the variation in cingulum development. Unless there are many species in this sample, it is likely that this character is variable in this species.
Primate Diversity in the Eocene of Saskatchewan
The Swift Current Creek locality of southwestern Saskatchewan was first described in 1933 (Russell and Wickenden, 1933). Ultimately, Russell (1965) recognized the Uintan age of the site based on faunal similarity to the upper Uinta Formation of Utah. Storer (1984) undertook the most comprehensive study of the fauna and described the depositional setting and paleoecology of the Swift Current Creek locality in great detail. He presented evidence suggesting the fossil assemblage was a rapid accumulation due to seasonal (spring or summer) flooding and the animal remains were from the local fauna. Our recent fieldwork at the locality revealed at least two pulses of flooding represented by two conglomeritic units, but the overall interpretation remains the same.
Storer (1996) described two local faunas from the Duchesnean sediments surrounding Lac Pelletier and exposures nearby to the south. The lower and upper faunas are separated by a thick non-fossil-bearing mudstone and are themselves in fluvial sands and gravels (Storer, 1996). All of the primates described by him are from the lower fauna and most are from a single locality (72J04-0006), a roadcut on the east side of Lac Pelletier at 831 m above sea level (Storer, 1990). Several primate species appear in his faunal list for the upper fauna (Storer, 1996), but to our knowledge these are unpublished. Unfortunately, the roadcut localities of the Lac Pelletier faunas are now covered by thick vegetation in most places, so surface collection is inefficient, and it is difficult to know where, on any given hillside, to obtain matrix for screening. The taxa present in the two local faunas broadly overlap, with the main difference being less diversity in the upper fauna–possibly an artifact of sampling. There is much more difference between the Lac Pelletier faunas and the Swift Current Creek local fauna.
Considering the Uintan and Duchesnean localities together, diversity and abundance of the various mammalian taxa suggested to Storer that the northern Great Plains constitutes a paleogeographic province with high endemism in the middle and late Eocene, bearing some overlap with the Rocky Mountain region and a little less with the coastal province represented in California (Storer, 1984, 1990, 1996). Considering primates specifically, it is remarkable that omomyid primates were present at modest diversity (and abundance) above the 49th parallel in the middle to late Eocene. It is perhaps not surprising, however, given the record of paromomyid plesiadapiforms in the Eocene of Ellesmere Island (West and Dawson, 1977; McKenna, 1980; Miller, et al., 2023). It is quite possible that the Cypress Hills–especially in areas along heavily treed rivers–represented a refuge for otherwise highly tropical mammals like primates, even as climate change began to favor the radiations of other groups of mammals (Prothero, 1985). However, this possibility cannot be evaluated without more high latitude faunas from this time interval in North America.
López-Torres and colleagues suggested that the persistence of paromomyids in the Arctic and their decline after the early Eocene farther south might be due to adaptations favoring colder climates, whereas trogolemurins might have favored warmer climates (López-Torres et al., 2018). This hypothesis is based on a small number of geographic data points, but southwestern Saskatchewan adds one more.
This review of Saskatchewan Eocene primates is timely considering the many omomyoids described since 1990. More recent evidence suggests a rich record of Uintan omomyoids (especially omomyines) at lower latitudes in North America (Williams and Kirk, 2008; Atwater and Kirk, 2018). The present review decreases the published number of omomyine species from the Uintan of Saskatchewan. One of the previously published species is an artiodactyl tooth and the other now constitutes a single Uintan-Duchesnean species, Saskomomys lindsayorum. In addition, the number of Uintan anaptomorphine species is increased to two (Trogolemur storeri and Walshina mcgrewi), highlighting the temporal and geographic persistence of trogolemurins. This primate fauna represents the only known foray of omomyoids into Canada and, as such, highlights the adaptability of these small, nocturnal primates. Future work will focus on reviewing additional groups of mammals from the Cypress Hills Formation, with descriptions of new specimens and species and further consideration of the Cypress Hills as a refugium for mammals.
ACKNOWLEDGMENTS
We thank our field crews from 2014-2017 for their efforts to increase our knowledge of Saskatchewan fossil mammals; these individuals include H. Kristjanson, S. Canington, K. Prufrock, A. Perry, I. Pratt, R. Black, C. Butts, M. Bradley, C. Zurowski, W. Petryshen, T. Cammidge, and E. Chabon. Thanks to the Lindsay family for permission to excavate on their land and for their generous hospitality while we were in the field. Thanks also to the Ungar family for permission to examine the hillsides on their land for additional localities. Thanks to J. Storer for his generosity in sharing his knowledge of these localities and faunas with us; we deeply appreciate the body of work he has generated and the immense task he performed in sorting out Saskatchewan mammals. We thank the many students and colleagues who spent hours looking for (and finding!) mammal teeth under the microscope; these include K. Prufrock, S. Canington, E. St Clair, A. Harper, J. Choi, M. McCann, J. Ferguson, W. Petryshen, R. Popp, Z. Lamoureux, C. Wilson, J. Creighton, T. Henry, S. Wiley, T. Bosch, K. Paradis, S. Yung, E. Tan, S. Schroeder, and S. Welkert, and numerous members of the Alberta Palaeontological Society. We thank K. Rose for providing access to the comparative collection he amassed and for assisting with identifications. We thank E. St Clair, M. Silcox, R. Dunn, and S. Zack for additional assistance in identifications. We thank Royal Saskatchewan Museum staff R. McKellar, T. Tokaryk, E. Bamforth, and W. Long for their assistance with RSM collections. We thank C. Henderson for additional acid preparation work. We thank Johns Hopkins University, the University of Calgary, and an NSERC Discovery Grant (to JMT) for providing funding for the fieldwork. MicroCT scans of some of the teeth were taken at the Duke University Shared Materials Instrumentation Facility; thanks for D. Boyer and J. Gladmann for their assistance; scanning and access to scans were supported by NSF grants BCS-1552848 and BCS-1440742 to D. Boyer. We thank S. López-Torres and an anonymous reviewer for their time and care in making this a significantly better paper.
REFERENCES
Atwater, A.L. and Kirk, E.C. 2018. New middle Eocene omomyines (Primates, Haplorhini) from San Diego County, California. Journal of Human Evolution, 124:7-24.
https://doi.org/10.1016/j.jhevol.2018.04.010
Beard, K.C., Krishtalka, L. and Stucky, R.K. 1992. Revision of the Wind River faunas, Early Eocene of central Wyoming. Part 12. New species of omomyid primates (Mammalia: Primates: Omomyidae) and omomyid taxonomic composition across the Early-Middle Eocene boundary. Annals of Carnegie Museum, 61:39-62.
Bown, T.M. and Rose, K.D. 1976. New Early Tertiary Primates and a reappraisal of some Plesiadapiformes. Folia Primatologica, 26:109-138. https://doi.org/10.1159/000155734
Boyer, D.M., Scott, C.S. and Fox, R.C. 2012. New craniodental material of Pronothodectes gaoi Fox (Mammalia, “Plesiadapiformes”) and relationships among members of Plesiadapidae. American Journal of Physical Anthropology, 147:511-550. https://doi.org/10.1002/ajpa.22003
Chester, S.G.B., Bloch, J.I., Boyer, D.M. and Clemens, W.A. 2015. Oldest known euarchontan tarsals and affinities of Paleocene Purgatorius to primates. Proceedings of the National Academy of Sciences of the United States of America, 112:1487-1492.
https://doi.org/10.1073/pnas.1421707112
Cope, E.D. 1883. On the mutual relations of the bunotherian Mammalia. Proceedings of the Academy of Natural Sciences of Philadelphia, 35:77-83.
Emry, R.J. 1990. Mammals of the Bridgerian (middle Eocene) Elderberry Canyon Local Fauna of eastern Nevada. Geological Society of America Special Papers, 243:187-210.
https://doi.org/10.1130/SPE243-p187
Fox, R.C. 2011. An unusual early primate from the Paleocene Paskapoo Formation, Alberta, Canada. Acta Palaeontologica Polonica, 56:1-10. https://doi.org/10.4202/app.2009.0079
Gingerich, P.D. and Winkler, D.A. 1979. Patterns of variation and correlation in the dentition of the Red Fox, Vulpes vulpes. Journal of Mammalogy, 60:691-704.
https://doi.org/10.2307/1380186
Gunnell, G.F. 1995. Omomyid Primates (Tarsiiformes) from the Bridger Formation, Middle Eocene, Southern Green River Basin, Wyoming. Journal of Human Evolution, 28:147-187. https://doi.org/10.1006/jhev.1995.1012
Gunnell, G.F. and Rose, K.D. 2002. Tarsiiformes: Evolutionary history and adaptation, p. 45-82. In Hartwig, W.C. (ed.), The Primate Fossil Record. Cambridge University Press, Cambridge.
Gunnell, G.F., Rose, K.D. and Rasmussen, D.T. 2008. Euprimates, p. 239-261. In Janis, C.M., Gunnell, G.F. and Uhen, M.D. (ed.), Evolution of Tertiary Mammals of North America. Volume 2: Small Mammals, Xenarthrans, and Marine Mammals. Cambridge University Press, Cambridge.
Kelly, T.S. 1990. Biostratigraphy of Uintan and Duchesnean land mammal assemblages from the middle member of the Sespe Formation, Simi Valley, California. Natural History Museum of Los Angeles County Contributions in Science, 419:1-42.
Kelly, T.S. and Murphey, P.C. 2016. Mammals from the earliest Uintan (middle Eocene) Turtle Bluff Member, Bridger Formation, southwestern Wyoming, USA, Part 1: Primates and Rodentia. Palaeontologia Electronica, 19.2.27A:1-55. https://doi.org/10.26879/586
Linnaeus, C. 1758. Systema naturae per regna tria naturae, secundum classes, ordines genera, species cum characteribus, differentris, synonymis, locis. Editis decima, reformata. Laurentii Salvii, Stockholm.
López-Torres, S., Silcox, M.T. and Holroyd, P.A. 2018. New omomyoids (Euprimates, Mammalia) from the late Uintan of southern California, USA, and the question of the extinction of the Paromomyidae (Plesiadapiformes, Primates). Palaeontologia Electronica, 21.3.37A:1-28. https://doi.org/10.26879/756
Matthew, W.D. 1909. The Carnivora and Insectivora of the Bridger Basin, Middle Eocene. Memoirs of the American Museum of Natural History, 9:287-578.
McKenna, M.C. 1980. Eocene paleolatitude, climate, and mammals of Ellesmere island. Palaeogeography, Palaeoclimatology, Palaeoecology, 30:349-362.
https://doi.org/10.1016/0031-0182(80)90065-6
Miller, K., Tietjen, K. and Beard, K.C. 2023. Basal Primatomorpha colonized Ellesmere Island (Arctic Canada) during the hyperthermal conditions of the early Eocene climatic optimum. PLoS ONE, 18:e0280114. https://doi.org/10.1371/journal.pone.0280114
Morse, P.E., Chester, S.G.B., Boyer, D.M., Smith, T., Smith, R., Gigase, P. and Bloch, J.I. 2019. New fossils, systematics, and biogeography of the oldest known crown primate Teilhardina from the earliest Eocene of Asia, Europe, and North America. Journal of Human Evolution, 128:103-131. https://doi.org/10.1016/j.jhevol.2018.08.005
Murphey, P.C. and Dunn, R.H. 2009. Hemiacodon engardae, a new species of omomyid primate from the earliest Uintan Turtle Buff Member of the Bridger Formation, southwestern Wyoming, USA. Journal of Human Evolution, 57:123-130. https://doi.org/10.1016/j.jhevol.2009.05.006
Prothero, D.R. 1985. North American mammalian diversity and Eocene-Oligocene extinctions. Paleobiology, 11:389-405. https://doi.org/10.1017/S0094837300011696
Robinson, P. 1968. The paleontology and geology of the Badwater Creek area, central Wyoming. Part 4. Late Eocene primates from Badwater, Wyoming, with a discussion of material from Utah. Annals of Carnegie Museum, 39:307-326.
Russell, L. S. 1965a. Tertiary mammals of Saskatchewan Part I: The Eocene fauna. Life Sciences Contributions, Royal Ontario Museum, 67:1-33.
Russell, L.S. 1965b. The continental Tertiary of western Canada, p. 41-52. In Vertebrate Paleontology in Alberta, University of Alberta, Edmonton.
Russell, L.S. 1972. Tertiary mammals of Saskatchewan Part II: The Oligocene fauna, non-ungulate orders. Life Sciences Contributions, Royal Ontario Museum, 84:1-97.
Russell, L.S. and Wickenden, R.T.D. 1933. An Upper Eocene vertebrate fauna from Saskatchewan. Transactions of the Royal Society of Canada, Series 3, 27:53-65.
Scott, C.S. and Fox, R.C. 2005. Windows on the evolution of Picrodus (Plesiadapiformes, Primates): Morphology and relationships of a species complex from the Paleocene of Alberta. Journal of Paleontology, 79:635-657.
https://doi.org/10.1666/0022-3360(2005)079[0635:WOTEOP]2.0.CO;2
Scott, C.S., Fox, R.C. and Redman, C.M. 2016. A new species of the basal plesiadapiform Purgatorius (Mammalia, Primates) from the early Paleocene Ravenscrag Formation, Cypress Hills, southwest Saskatchewan, Canada: further taxonomic and dietary diversity in the earliest primates. Canadian Journal of Earth Sciences, 53:343-354. https://doi.org/10.1139/cjes-2015-0238
Scott, C.S., López-Torres, S., Silcox, M.T. and Fox, R.C. 2023. New paromomyids (Mammalia, Primates) from the Paleocene of southwestern Alberta, Canada, and an analysis of paromomyid interrelationships. Journal of Paleontology, 97:477-498.
https://doi.org/10.1017/jpa.2022.103
Storer, J.E. 1978. Tertiary sands and gravels in Saskatchewan and Alberta: correlation of mammalian faunas, p. 595-602. In Stelck, C.E. and Chatterton, B.D.E. (ed.), Western and Arctic Canadian Biostratigraphy. Geological Association of Canada, St. John's, Newfoundland and Labrador.
Storer, J.E. 1983. A new species of the artiodactyl Heptacodon from the Cypress Hills Formation, Lac Pelletier, Saskatchewan. Canadian Journal of Earth Sciences, 20:1344-1347. https://doi.org/10.1139/e83-119
Storer, J.E. 1984. Mammals of the Swift Current Creek Local Fauna (Eocene: Uintan), Saskatchewan. Natural History Contributions, Saskatchewan Museum of Natural History, 7:1-158.
Storer, J.E. 1987. Dental evolution and radiation of Eocene and Early Oligocene Eomydiae (Mammalia, Rodentia) of North America, with new material from the Duchesnean of Saskatchewan. Dakoterra, 3:108-117.
Storer, J.E. 1990. Primates of the Lac Pelletier Lower Fauna (Eocene: Duchesnean) of Saskatchewan. Canadian Journal of Earth Sciences, 27:520-524.
https://doi.org/10.1139/e90-048
Storer, J.E. 1996. Eocene-Oligocene faunas of the Cypress Hills Formation, Saskatchewan, p. 240-261. In Prothero D.R. and Emry R.J. (eds), The Terrestrial Eocene-Oligocene Transition in North America. Cambridge University Press, Cambridge.
Storer, J.E. and Tokaryk, T.T. 1992. Catalogue of type and figured fossils. Natural History Contributions, Saskatchewan Museum of Natural History, 10:1-67.
Szalay, F.S. 1976. Systematics of the Omomyidae (Tarsiiformes, Primates): taxonomy, phylogeny and adaptations. Bulletin of the American Museum of Natural History, 156:157-450.
Tornow, M.A. 2008. Systematic analysis of the Eocene primate family Omomyidae using gnathic and postcranial data. Bulletin of the Peabody Museum of Natural History, 49:43-129.
https://doi.org/10.3374/0079-032X(2008)49[43:SAOTEP]2.0.CO;2
Trouessart, E.L. 1879. Catalogue des mammifères vivants et fossiles. Revue et magasin de zoologie, 7:223-230.
West, R.M. and Dawson, M.R. 1977. Mammals from the Palaeogene of the EurekaSound Formation: Ellesmere Island, Arctic Canada. Geobios, 10:107-124.
https://doi.org/10.1016/S0016-6995(77)80011-9
Williams, B.A. and Kirk, E.C. 2008. New Uintan primates from Texas and their implications for North American patterns of species richness during the Eocene. Journal of Human Evolution, 55:927-941. https://doi.org/10.1016/j.jhevol.2008.07.007
