

 The body impression of a new genus and species of cymothoid isopod (Cymothoidae, Crustacea) preserved in a vertebrate coprolite from the Atlantic Coastal Plain (Miocene, Chesapeake Group) of Virginia, USA
The body impression of a new genus and species of cymothoid isopod (Cymothoidae, Crustacea) preserved in a vertebrate coprolite from the Atlantic Coastal Plain (Miocene, Chesapeake Group) of Virginia, USA
Article number: 27.3.a51
https://doi.org/10.26879/1371
Copyright Society of Vertebrate Paleontology, October 2024
Author biographies
Plain-language and multi-lingual abstracts
PDF version
Submission: 27 January 2024. Acceptance: 17 October 2024.
ABSTRACT
Body impressions of ingested organisms preserved in coprolites are exceedingly rare in the fossil record. Only two previous publications report on the preservation of an impression of the body of an organism in a coprolite. Herein, the impression of the dorsal surface of the exoskeleton of a new genus and species of cymothoid isopod, Calverteca osbornei, preserved in a vertebrate coprolite is described. The coprolite probably originated from the Newport News beds of the lower Miocene Calvert Formation of the Chesapeake Group of Virginia, USA. This coprolite is the first report in the fossil record of the impression of the body of a crustacean (Arthropoda) preserved in a coprolite. This is also the first report of an isopod within the Chesapeake Group. The producer of the original fecal mass is unknown, although a fish seems the most likely given that cymothoid isopods parasitize the oral cavities of bony fishes.
Stephen J. Godfrey. Department of Paleontology, Calvert Marine Museum, PO Box 97, Solomons, Maryland, 20688, USA (corresponding author) Stephen.Godfrey@calvertcountymd.gov
and Research Associate, National Museum of Natural History, Smithsonian Institution, Washington, DC, 20560, USA and Research Associate, Natural History Museum of LA County, 700 Exposition Park Dr, Los Angeles, California, 90037, USA
Rodney M. Feldmann. Department of Earth Sciences, Kent State University, Kent, Ohio, 44242, USA
Carrie E. Schweitzer. Department of Earth Sciences, Kent State University at Stark, North Canton, Ohio, 44729, USA. cschweit@kent.edu
Keywords: new genus; new species; coprolite; cymothoid isopod; Newport News beds; Calvert Formation
Final citation: Godfrey, Stephen J., Feldmann, Rodney M., and Schweitzer, Carrie E. 2024. The body impression of a new genus and species of cymothoid isopod (Cymothoidae, Crustacea) preserved in a vertebrate coprolite from the Atlantic Coastal Plain (Miocene, Chesapeake Group) of Virginia, USA. Palaeontologia Electronica, 27(3):a51.
https://doi.org/10.26879/1371
palaeo-electronica.org/content/2024/5363-isopod-impression-in-a-coprolite
Copyright: October 2024 Society of Vertebrate Paleontology.
This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
creativecommons.org/licenses/by/4.0
https://zoobank.org/CA9A87F1-361D-4F99-B9FF-CB47ACAEFCBF
INTRODUCTION
Only two previous reports of the impression of the body of an organism preserved in a coprolite are known (Godfrey et al., 2017; Frandsen, 2022). Godfrey et al. (2017) described the impression of a portion of the carapace of a hatchling turtle preserved in a coprolite, and Frandsen (2022) reported on the body impression of an ammonite in a coprolite. These impressions, along with actual fossilized plants or body parts preserved in coprolites, comprise the most definitive evidences of diets, either accidental or intentional, of extinct organisms.
 For much of the Miocene (circa 22-8 Ma), the Chesapeake Bay region in Maryland and Virginia was intermittently inundated by the Atlantic Ocean (Figure 1). Some of the continental siliciclastic sediments that were laid down within this arm of the Atlantic Ocean (i.e., the Salisbury Embayment) are now exposed as sea bluffs along a relatively short portion of the western shore of Chesapeake Bay (i.e., Calvert Cliffs) and along some of the tributaries flowing into the bay. These sediments abound with marine fossils, including the tests of planktonic organisms, shelled invertebrates, shark teeth, teleost and marine mammal bones, and many other kinds of fossils numbering over 650 taxa (Godfrey, 2018, 2023; Vogt et al., 2018). In addition to innumerable body fossils, these Miocene sediments preserve trace fossils including burrows made by invertebrates (Thalassinoides, Gyrolithes and Ophiomorpha; Kidwell et al., 2015), shark bite marks, usually on cetacean bone (Godfrey et al., 2018; Godfrey and Lowry, 2021), and coprolites from both vertebrates and invertebrates (Wetmore, 1943; Godfrey and Smith, 2010; Kent, 2018; Weems, 2018; Godfrey and Collareta, 2022; Godfrey et al., 2022).
For much of the Miocene (circa 22-8 Ma), the Chesapeake Bay region in Maryland and Virginia was intermittently inundated by the Atlantic Ocean (Figure 1). Some of the continental siliciclastic sediments that were laid down within this arm of the Atlantic Ocean (i.e., the Salisbury Embayment) are now exposed as sea bluffs along a relatively short portion of the western shore of Chesapeake Bay (i.e., Calvert Cliffs) and along some of the tributaries flowing into the bay. These sediments abound with marine fossils, including the tests of planktonic organisms, shelled invertebrates, shark teeth, teleost and marine mammal bones, and many other kinds of fossils numbering over 650 taxa (Godfrey, 2018, 2023; Vogt et al., 2018). In addition to innumerable body fossils, these Miocene sediments preserve trace fossils including burrows made by invertebrates (Thalassinoides, Gyrolithes and Ophiomorpha; Kidwell et al., 2015), shark bite marks, usually on cetacean bone (Godfrey et al., 2018; Godfrey and Lowry, 2021), and coprolites from both vertebrates and invertebrates (Wetmore, 1943; Godfrey and Smith, 2010; Kent, 2018; Weems, 2018; Godfrey and Collareta, 2022; Godfrey et al., 2022).
These coprolites include microcoprolites (Godfrey et al., 2022), burrowed/scavenged coprolites (Godfrey and Collareta, 2022), and uncommon finds such as fish-bitten coprolites and those preserving tooth impressions (Godfrey and Smith, 2010; Lindholm et al., 2023).
In this report we describe another exceptionally rare kind of coprolite, one that preserves an impression of much of the body of an ingested organism, in this case, a cymothoid isopod (Crustacea) (Figure 2). This remarkable coprolite boasts a number of firsts: the first preserving the body impression of an arthropod, the first preserving the body impression of an isopod, sufficiently preserved to describe and establish the name of a new genus and species, and the first record of an isopod from the Miocene Chesapeake Group. The present paper aims to 1) enhance our knowledge of the diverse coprofauna of the Atlantic Coastal Plain, 2) show that the exoskeleton of an isopod can pass essentially intact through the digestive tract of another organism, and 3) report on a new genus and species of parasitic isopod from these shallow marine deposits.
MATERIALS AND METHODS
Abbreviations
CMM-V-: Calvert Marine Museum vertebrate paleontology collection, Solomons, Maryland, U.S.A.; ICZN: International Commission on Zoological Nomenclature; Ma: Mega-annum; PCS: Popes Creek Sand.
Specimen and Image Preparation
A latex cast was made of the natural mold of the isopod (Figure 2 and Figure 3). Mold release (i.e., Polytek® Plo-Ease 2500 Release Agent) was sprayed on the coprolite so that once the molding compound (latex) cured, the flexible cast could be more easily removed from the natural mold of the isopod without damaging the impression. Before the latex was applied to the specimen, it was diluted with water to the consistency of milk so that it would penetrate more easily into all the crevices in the natural mold. The diluted latex was applied using a standard artist’s airbrush. Natural latex is cream-colored, a difficult tone on which to see anatomical details. Therefore, the dilute latex was pigmented with black acrylic paint.
A single application of dilute latex dries to a very thin and delicate yet flexible layer. Therefore, multiple layers (approximately 10) of dilute latex were applied using an airbrush. Successive latex layers consisted of more viscous latex applied with a tiny paint brush to build up the thickness of the latex cast whereby making it easier to demold because of its thickness and resulting tear-resistance. The final two layers consisted of undiluted latex. The resulting flexible, high-resolution cast was whitened with sublimed ammonium chloride (see below) and photographed (Figure 3).
In its natural state, CMM-V-11695 is dark in color, which makes it difficult to see and interpret the anatomical details preserved in the impression of the isopod. Therefore, to improve contrast and visibility, the coprolite was lightly dusted with sublimed ammonium chloride (Figure 2), a whitening technique described by Cooper (1935) and Feldmann (1989). Both specimens were then photographed: the coprolite with a Nikon Coolpix P510 camera on black velvet under fluorescent light, and the latex cast using a Leica Z6APO macroscope with a Planepo 0.5X lens and Spotflex digital camera.
The ammonium chloride was removed by holding the specimen under running water (Shelburne and Thompson, 2016). Prior to whitening any fossil with ammonium chloride, it is important to determine whether the specimen is stable enough to withstand a fresh water rinse because without that, there would be the possibility of a residue of hydrochloric acid (HCl) left on the specimen if it was not washed thoroughly (although we are not sure if the possibility of an acidic residue has ever been tested).
Specimen Repository
The specimen described herein is stored in the vertebrate paleontology collection as CMM-V-11695 in the Calvert Marine Museum, Solomons, Maryland, USA.
RESULTS
Locality and Geologic Setting
CMM-V-11695 was found and collected by Jason E. Osborne while scuba diving along the bed of the Pamunkey River in Virginia, USA. GPS coordinates for the location where the coprolite was found are recorded in the database of the Calvert Marine Museum and will be made available to qualified researchers.
During the Miocene, a large depositional basin (the Salisbury Embayment) occupied what is now a small portion of eastern Virginia, much of Maryland, and all of Delaware (Ward, 1992). It was located between the South New Jersey arch to the north and the Norfolk arch to the south. Within that basin, the accumulated siliciclastic sediments have been subdivided into five Miocene-age formations: the Old Church (mostly an Oligocene-age Formation), Calvert, Choptank, St. Marys, and Eastover, in ascending stratigraphic order. These five Miocene stratal packages belong to the northern portion of the geographically wide-ranging Chesapeake Group. Although these formations preserve multiple transgressive-regressive cycles, in their totality, they evidence a gradual retreat over a 14-million-year period of the Atlantic Ocean eastward across the coastal plain (Kidwell at al., 2015; Vogt et al., 2018).
Although the isopod-bearing coprolite, CMM-V-11695, was not found in situ, and neither were there any adhering sediments that could have been analyzed for stratigraphic index microfossils, there is no reason to think that the coprolite was not locally derived from either the Old Church Formation or the Newport News beds of the Calvert Formation. In the area where the coprolite was found, the Newport News beds lie unconformably over the Old Church Formation (Edwards, et al., 2005). The Old Church Formation is exposed in small pockets along the bottom of the river (Jason Osborne, pers. comm., 2024). Edwards et al. (2005) and Weems (2018) confirmed the presence of the Newport News beds of the Calvert Formation and the Old Church Formation based on other specimens found at the site. An analysis of the dinoflagellates found in neighboring sediments confirm the base of the river to be Newport News beds. In addition, the coprolite was found among many other lower Calvert Formation fossils (Jason Osborne, pers. comm., 2024). Edwards et al. (2005) confirmed the Newport News beds to be basal Calvert Formation, early Burdigalian. Approximately 1.5 km downriver from the site of discovery, sediments of the upper Miocene St Marys and Eastover formations are exposed. Given the location of the coprolite upriver from these other younger Chesapeake Group formations, it would not have been possible for the coprolite to have been derived from them. At the point along the Pamunkey River where the coprolite was found, there are only Newport News beds above the Old Church Formation.
The flora and fauna indicate deposition of the Newport News beds in marine nearshore to shallow-shelf water depths, in an outer neritic (to possibly middle neritic) environment during a paleoclimactic period that was somewhat warmer than the present climate in Virginia, USA (Edwards et al., 2005).
Description of the Coprolite
The dark reddish-brown colored coprolite is 73 mm long and 22 mm at its widest point (Figure 2). One end of CMM-V-11695 is bluntly pointed, the other, close to the location of the impression of the isopod, appears naturally truncated. On this same end, a small portion of the coprolite is covered with linear striations, suggesting the activity of mouth parts or feeding appendages of an unknown organism. The surface of the coprolite is variously textured, clearly seen when coated with sublimed ammonium chloride (Figure 2). There are no observable inclusions within CMM-V-11695.
From its size, this coprolite (CMM-V-11695) is regarded as vertebrate in origin. Within the Calvert Formation, there are no known invertebrates that could have produced such a relatively large fecal mass. The vertebrate fauna from the same formation is large and diverse, including a large suite of chondrichthians; bony fishes; turtles; crocodilians; mostly pelagic birds; marine mammals including seals, sea cows, odontocetes, and mysticetes, and very rarely, the isolated remains of large land mammals (Godfrey, 2018, 2023). It is possible to rule out a chondrichthyan origin of the coprolite because their feces are scrolled or spiraled, the result of the corkscrew-shaped terminal portion of the intestine (i.e., their spiral valve). Avian feces usually do not occur as such large and compact masses, although what the fecal mass of the large and pelagic super-soarer Pelagornis was like is unknown. Coprolites attributed to seals and sea cows are also not yet known and characterized. It is unlikely that the coprolite originated from a terrestrial mammal (but not impossible) given the marine sediments from within which the coprolite was recovered.
The occurrence of a cymothoid isopod, which is typically associated with fishes, suggests that the coprolite was produced by a fish or that the isopod, or isopod-parasitized fish, was ingested as a food source by another vertebrate organism, such as a turtle (Hunt and Lucas, 2012; Bajdek et al., 2019), a crocodile (Weems, 2018), or a mammal.
The size and shape of CMM-V-11695 is congruent with coprolites produced by turtles (Hunt and Lucas, 2012; Bajcek et al., 2019). The coprolite could also be crocodilian in origin. Crocodilian coprolites are generally sausage shaped and circular in cross section, with few structures visible on their outer surface except for occasional striations or traces from coprophagous organisms (Souto, 2010; Milàn, 2012, Milàn et al., 2018; Godfrey et al., 2022; Godfrey and Collareta, 2022; Lindholm et al., 2023). CMM-V-11695 is consistent in size and shape to those of crocodilian coprolites collected elsewhere (Häntzschel et al., 1968; Sawyer, 1981, 1998; Hunt and Lucas, 2010; Milàn, 2012).
SYSTEMATIC PALEONTOLOGY
Order ISOPODA Latreille, 1816
Suborder CYMOTHOIDA Leach, 1814
Remarks. Placement within Cymothoida follows the phylogenetic analysis of Wägele (1989) (WoRMS Editorial Board, 2023) based upon possession of five free pleonites in which the first is similar to the others and pleopods that are ovoid and rounded distally (Brandt and Poore, 2003). Other characters of the suborder are not visible in the fossil specimen.
Superfamily CYMOTHOOIDEA Leach, 1814
Family CYMOTHOIDAE Leach, 1814
Remarks. Cymothoidae has been subject to numerous analyses confirming its monophyletic status (Trilles, 1999; Brandt and Poore, 2003; Smit et al., 2014). Brandt and Poore (2003) recognized the family and referred to Trilles (1999) for a diagnosis. Cymothoidae includes a speciose assemblage of primarily marine taxa characterized by an outline becoming widest at about pereon 5; coxal plates 5-7 visible; pleonite one similar to others and large, ovoid pleopods (Brandt and Poore, 2003). Members of the family are characterized by often having a large size (1-6 cm), and all are parasitic on fish. Cymothoidae is one of the largest families of isopods, and they have been described as parasites on nearly every marine fish family as well as some freshwater groups (Smit et al., 2014).
Isopods have a modest fossil record (Schram and Koenemann, 2021). Because their classification at the family and genus level is so dependent on morphology that does not frequently fossilize, comparison of fossil and extant isopods is precarious (Poore and Bruce, 2012). The fossil remains described here as they are preserved do not provide sufficient morphological detail to assign the fossil to a genus within the Cymothoidae with certainty. Generic characters used to classify isopods include details of the head, mouthparts, and pereopods, none of which is preserved in the specimen (Brandt and Poore, 2003; Poore and Bruce, 2012). Furthermore, assigning fossil forms to extant genera is severely limited because in many cases preservable hard parts are also not present (Poore and Bruce, 2012).
The new fossil is similar to species of Livoneca Leach, 1818; Nerocila Leach, 1818; and Aegathoa Dana, 1853, all of which are known from coastal Maryland today (marylandbiodiversity.com; OBIS). Each of these taxa are characterized by an overall ovate pereion and pleonites much narrower than pereionites. The fossil differs somewhat in narrowing strongly anteriorly beginning at pereonite five, unlike in these extant genera wherein the pereon maintains a broadly convex margin along its entire length. Thus, because assignment of fossil isopods to extant taxa is limited by preservation, and because the specimen differs from the extant, geographically proximal forms, we elect to name a new genus to accommodate the new material.
Genus Calverteca gen. nov.
zoobank.org/A6694AAB-B375-4290-8E31-F54D6E63EC39
Type species. Calverteca osbornei new species (based on a body impression preserved in CMM-V-11695), by original designation.
Etymology. The genus name is derived from the Calvert Formation, from which it was collected, and the ending -eca, as in some of the genera in the family. The gender is feminine.
Diagnosis. Small size, about 17 mm long and ca. 10 mm wide at pereonite 6. Cephalon missing, apparently positioned in advance of pereonite 1 and not enveloped by it. Pereonites increase in width from pereonite 1 to 6 and narrowing to pereonite 7. Pereonite 1 longest; length of pereonites 2-7 increasing posteriorly. Coxal plates exposed laterally on pereonites 5-7. Pleon markedly narrower than pereon. Pleonites 1-5 equally long and decreasing in width posteriorly. Pleotelson broad anteriorly and broken and missing posteriorly. Pleopods long, blade-like.
Remarks. This is the first notice of a member of Cymothoidae in the fossil record (Smit et al., 2014). Members of Cirolanidae, in the same infraorder, have a fossil record ranging from the Early Jurassic to Holocene (Poore and Bruce, 2012; Maguire et al., 2018; Schram and Koenemann, 2021). Members of the same superfamily, Cymothooidea, are known from the Late Jurassic to Holocene (Schram and Koenemann, 2021).
Calverteca osbornei gen. et sp. nov.
Figure 2 and Figure 3
zoobank.org/239D9389-C015-42D7-AB74-3E502E0BC355
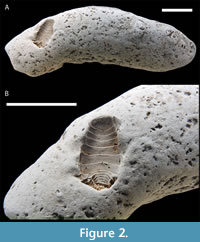 Holotype. The sole specimen, the holotype, of Calverteca osbornei sp. nov., CMM-V-11695, is deposited in the vertebrate paleontology collection in the Calvert Marine Museum, Department of Paleontology, Solomons, Maryland, USA. It was included in the vertebrate paleontology collection because the coprolite was deemed to be vertebrate in origin.
Holotype. The sole specimen, the holotype, of Calverteca osbornei sp. nov., CMM-V-11695, is deposited in the vertebrate paleontology collection in the Calvert Marine Museum, Department of Paleontology, Solomons, Maryland, USA. It was included in the vertebrate paleontology collection because the coprolite was deemed to be vertebrate in origin.
Etymology. Named in honor of Jason E. Osborne who found and donated the type specimen to the Calvert Marine Museum. “Osbornei” was created by adding an “i” to “Osborne” in accordance with Article 31.1.2. of the ICZN.
Locality and stratigraphic position. The specimen was collected on the bed of the Pamunkey River, Virginia, USA in association with sediments of the Newport News beds in the lower Miocene Calvert Formation.
Diagnosis. As for genus.
Decription. Impressions of the cephalon and antennae do not appear to have been preserved and neither are there any impressions of pereopods. However, the seven pereonites and five pleonites are clearly visible. Within the pereon, the first six pereonites increase evenly in width from 3.3 mm for the first pereonite to 5.9 mm for the width of the sixth pereonite. Pereonite 7 decreases to approximately 5.2 mm wide. The midline length of the pereonites ranges from 0.7 mm for the second to 1.1 mm for the seventh (Figure 2). The entire pleon is approximately 2 mm long by 4 mm wide across the first pleonite. Along the midline, the length of each pleonite is about 0.3 mm. The pleonites decrease in width evenly to approximately 1.9 mm for the fifth. The pleopods are also preserved. They are strong and eliptical in outline.
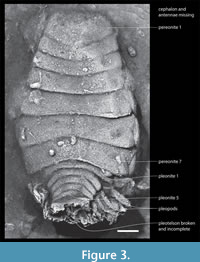 Remarks. The sole specimen is preserved as a mold of the exterior situated transverse to the long axis of the vertebrate coprolite that embraces it. The mold is embedded deeply enough within the coprolite to suggest that the isopod was not well exposed on the surface. Preservation in this fashion, with no matrix material filling the mold, suggests that the isopod was complete both when ingested and voided to be incorporated as it is into the coprolite.
Remarks. The sole specimen is preserved as a mold of the exterior situated transverse to the long axis of the vertebrate coprolite that embraces it. The mold is embedded deeply enough within the coprolite to suggest that the isopod was not well exposed on the surface. Preservation in this fashion, with no matrix material filling the mold, suggests that the isopod was complete both when ingested and voided to be incorporated as it is into the coprolite.
Cymothoids are parasitic isopods (Smit et al., 2014) that attach to the tongue and other soft-tissue regions in fishes. It has been well-established that isopods of this family and body form are parasitic on fish (Smit et al., 2014). A search of the literature did not reveal any reference to cymothoids as parasites on reptiles. Very large isopods, Bathynomus, have been observed to scavenge upon dead crocodiles. Thus, several scenarios may be visualized to account for the small isopod within a fecal mass. 1) The individual enclosed within the coprolite might have been ingested by the reptilian producing the coprolite when the isopod was detached from its original host. 2) This cymothoid may have parasitized a wider variety of hosts in the Miocene than today. However, the fossil is sufficiently similar to modern cymothoids that one can safely say that it was an external parasite of fish. 3) The coprolite may have been produced by a fish, and the isopod was, indeed, a parasite that was dislodged and ingested.
There are over 10,000 known extant species of isopods that inhabit terrestrial, freshwater, and marine environments (Poore and Bruce, 2012; Schram and Koenemann, 2021). Most members are less than 2 cm (0.8 in) in length. For being such a successful cosmopolitan and speciose group of crustaceans, isopods have a poor and very incomplete fossil record (Schram, 1970; Maguire et al., 2018; Schram and Koenemann, 2021). Although they first appeared in the fossil record during the Carboniferous (Schram, 1970), remarkably few extinct species have been described (Wieder and Feldmann, 1992; Schram and Koenemann, 2021). Maguire et al. (2018) attributed this not to their numerical paucity through time, but rather, likely due to the physical and chemical composition of their thin and lightly sclerotized cuticle. Furthermore, their cuticle would have rapidly disintegrated following their destructive biphasic molting behavior (Tait, 1917). These attributes decrease the chances of fossilization.
CONCLUSIONS
Body impressions of ingested organisms preserved in coprolites are exceedingly rare in the fossil record. Here, we report on the impression of the dorsal surface of the body of a cymothoid isopod preserved in a vertebrate coprolite. The coprolite originated from the lower Miocene Calvert Formation of the Chesapeake Group of Virginia, USA. This is the first report in the fossil record of the impression of the body of a crustacean (Calverteca osbornei gen. et. sp. nov.) preserved in a coprolite. This is also the first report of an isopod within the Chesapeake Group. It is unclear what kind of vertebrate ingested the isopod and produced the original fecal mass, although it was almost certainly produced by a fish.
DECLARATIONS
Availability of Data and Materials
The specimen described herein and GPS coordinates will be made available through the Calvert Marine Museum to qualified researchers.
Funding
This article was made possible by funding from the citizens of Calvert County, Maryland, USA, the County Board of Calvert County Commissioners, Maryland, USA, and the Clarissa and Lincoln Dryden Endowment for Paleontology at the Calvert Marine Museum.
Authors’ Contributions
SJG anchored this project, made the pigmented latex cast of the natural impression of the isopod, contributed to and edited the entire text, prepared Figure 1 and Figure 2 and formatted and added labels to Figure 3.
RMF described and remarked on the occurrence of the isopod specimen, edited the entire text, and photographed the cast shown in Figure 3.
CES edited and expanded upon the isopod description, edited the entire text, and expanded the references.
ACKNOWLEDGMENTS
The discovery and recovery of this remarkable coprolite on the bed of the Pamunky River, would not have come about without the black-water scuba expertice of J.E. Osborne. He then generously donated the specimen to the Calvert Marine Museum. Dr. G.D.F. (Buz) Wilson, Saugatuck Natural History Laboratory, Saugatuck, Michigan, provided invaluable information on the identity of the isopod specimen for R.M.F. We also gratefully acknowledge R.E. Weems for help in the placement of the Newport News beds within the context of the Chesapeake Group. The very helpful comments by the two anonymous reviewers improved the final draft; thank you! We would also like to acknowledge M. Hyžný, the Executive Editor of Palaeontologia Electronica, for helping to shepherd this manuscript through the review process. Very sadly, author Rodney M. Feldmann passed away during the preparation of this publication.
Competing Interests
The authors declare that they have no conflicts of interest.
REFERENCES
Bajdek, P., Szczygielski, T., Kapuścińska, A., and Sulej, T. 2019. Bromalites from a turtle-dominated fossil assemblage from the Triassic of Poland. Palaeogeography, Palaeoclimatology, Palaeoecology, 520:214–228.
https://doi.org/10.1016/j.palaeo.2019.02.002
Brandt, A. and Poore, G.C.B. 2003. Higher classification of the flabelliferan and related Isopoda based on a reappraisal of relationships. Invertebrate Systematics, 17:893–923.
https://doi.org/10.1071/IS02032
Cooper, C.L. 1935. Ammonium chloride sublimate apparatus. Journal of Paleontology, 9(4):357–359.
Dana, J.D. (1853-1855). Crustacea. Part II. In: United States Exploring Expedition. During the years 1838, 1839, 1840, 1841, 1842. Under the command of Charles Wilkes. U. S. N., C. Sherman Printer, Philadelphia. 14:691–1618.
Edwards, L.E., Barron, J.A., Bukry, D., Bybell, L.M., Cronin, T.M., Poag, C.W., Weems, R.E., and Wingard, G.L. 2005. Paleontology of the Upper Eocene to Quaternary Postimpact Section in the USGS-NASA Langley Core, Hampton, Virginia. In Horton Jr., J.W., Powars, D.S., and Gohn, G.S. (eds.), Studies of the Chesapeake Bay Impact Structure–The USGS-NASA Langley Corehole, Hampton, Virginia, and Related Coreholes and Geophysical Surveys, Professional Paper 1688, Chapter H, pp. H1-H71, Reston, VA: United States Geological Survey.
https://doi.org/10.3133/pp1688H
Feldmann, R.M. 1989. Whitening fossils for photographic purposes. In Feldmann, R.M. (ed.), Paleotechniques. The Paleontological Society Special Publication, 4:342–346.
https://doi.org/10.1017/S2475262200005323
Frandsen, G. 2022. Ammonite impression in coprolite. The Ecphora, 37(2):3.
Godfrey, S.J. (ed.) 2018. The Geology and Vertebrate Paleontology of Calvert Cliffs. Smithsonian Contributions to Paleobiology, Washington, 100.
https://lccn.loc.gov/2017059091
Godfrey, S.J. (ed.) 2023. The Geology and Vertebrate Paleontology of Calvert Cliffs - Volume 2 Turtles and Toothed Whales. Smithsonian Contributions to Paleobiology, Washington, 107.
https://doi.org/10.5479/si.23847438
Godfrey, S.J. and Smith, J. 2010. Shark-bitten vertebrate coprolites from the Miocene of Maryland. Naturwissenschaften, 97(5):461–467.
https://doi.org/10.1007/s00114-010-0659-x
Godfrey, S.J. and Lowry, A.J. 2021. The ichnospecies Linichnus bromleyi on a Miocene baleen whale radius preserving multiple shark bite-shake traces suggests scavenging. Carnets Geol, Madrid 21(17):391–398.
https://doi.org/10.2110/carnets.2021.2117
Godfrey, S.J. and Collareta, A. 2022. A new ichnotaxonomic name for burrows in vertebrate coprolites from the Miocene Chesapeake Group of Maryland, U.S.A. Swiss Journal of Palaeontology, 141:9.
https://doi.org/10.1186/s13358-022-00250-6
Godfrey, S.J., Weems, R.E., and Palmer, B. 2017. Turtle shell impression in a coprolite from South Carolina, USA, Ichnos, 26:20–27.
https://doi.org/10.1080/10420940.2017.1386662
Godfrey, S.J., Collareta, A., and Nance, J.R. 2022. Coprolites from Calvert Cliffs: Miocene fecal pellets and burrowed crocodilian droppings from the Chesapeake Group of Maryland, USA. Rivista Italiana di Paleontologia e Stratigrafia, 128(1):69–79.
https://doi.org/10.54103/2039-4942/17064
Godfrey, S. J., Ellwood, M., Groff, S., and Verdin, M.S. 2018. Carcharocles-bitten odontocete caudal vertebrae from the Coastal Eastern United States. Acta Palaeontologica Polonica, 63(3):463–468.
https://doi.org/10.4202/app.00
Häntzschel, W., El-Baz, F., and Amstutz, G.C. 1968. Coprolites: An annotated bibliography. Geological Society of America Memoir, 108:1–132.
Hunt, A.P. and Lucas, S.G. 2010. Crocodylian coprolites and the identification of the producers of coprolites. New Mexico Museum of Natural History and Science Bulletin, 51:219–226.
Hunt, A.P. and Lucas, S.G. 2012. Classification of vertebrate coprolites and related trace fossils. New Mexico Museum of Natural History and Science Bulletin, 57:137–146.
Kent, B.W. 2018. The cartilaginous fishes (chimaeras, sharks and rays) of Calvert Cliffs, Maryland, USA. Smithsonian Contributions to Paleobiology, 100:45–160.
https://lccn.loc.gov/2017059091
Kidwell, S.M., Powars, D.S., Edwards, L.E., and Vogt, P.R. 2015. Miocene stratigraphy and paleoenvironments of the Calvert Cliffs, Maryland. The Geological Society of America Field Guide, 40:231–279.
https://doi.org/10.1130/2015.0040(08)
Latreille, P.A. 1816. Les crustacés, les arachnides, et les insectes. In [G. L. C. F. D.] Cuvier, F. (ed.), Le Règne Animal, Distribué d’après son Organisation, pour Servrir de Base a l’Histoire Naturelle des Animaux et d’Introduction a l’Anatomie Comparée. Volume 3: i-xxix+1-653. Paris: Deterville [published 2 December 1816 fide Roux, 1976].
Leach, W.E. 1813-1815. Crustaceology. In Brewster, D. (ed.), The Edinburgh Encyclopaedia. Balfour, Edinburgh, vol. 7(1): 383–384 [1813], 7(2): 385–437, 765–766 [1814], 9(1): pl. CCXXI [1815].
https://www.biodiversitylibrary.org/page/37187640
Leach, W.E. 1818. Cymothoadées, Cymothoadae. In Cuvier, F. (ed.), Dictionnaire des Sciences Naturelles, dans Lequel on Traite Methodiquement des Differens Etres de la Nature, Consideres Soit en Eux-memes, d'apres l'Etat Actuel de nos Connoissances, Soit Relativement a l'Utilite qu'en Peuvent Retirer la Medecine, l'Agriculture, le Commerce et les Artes, 12:338–354.
https://www.biodiversitylibrary.org/page/23002019
Lindholm, A., Godfrey, S.J., L. Ward, W., and Collareta, A. 2023. A gar-bitten vertebrate coprolite from the Neogene of the Atlantic Coastal Plain, USA, and a new ichnospecies of Machichnus Mikuláš et al., 2006. Ichnos, 29:185–194.
https://doi.org/10.1080/10420940.2023.2182300
Maguire, E.P., Feldmann, R.M., Jones, W.T., Schweitzer, C.E., and Casadío, S. 2018. The first fossil isopod from Argentina: a new species of Cirolanidae (Crustacea: Peracarida) from the Miocene of Patagonia. Journal of Crustacean Biology, 38(1):34–44.
https://doi.org/10.1093/jcbiol/rux100
Maryland Biodiversity Project. Cymothoidae. Accessed May 10, 2024.
https://www.marylandbiodiversity.com/checklist/?family=Cymothoidae
Milàn, J. 2012. Crocodylian scatology - a look into morphology, internal architecture, inter- and intraspecific variation and prey remains in extant crocodylian feces. New Mexico Museum of Natural History and Science Bulletin, 57:65–72.
Milàn, J., Rasmussen, E.S., and Dybkjær, K. 2018. A crocodilian coprolite from the lower Oligocene Viborg Formation of Sofienlund Lergrav, Denmark. Bulletin of the Geological Society of Denmark, 66:181–187.
Poore, G.C.B. and Bruce, N.L. 2012. Global diversity of marine isopods (except Asellota and crustacean symbionts). PLoS ONE, 7:1–15.
https://doi.org/10.1371/annotation/3260cd00-89cf-4e08-ab25-07e0be598ab4
Powars, D.S., Edwards, L.E., Kidwell, S.M., and Schindler, J.S. 2015. Cenozoic stratigraphy and structure of the Chesapeake Bay region. In Brezinski, D.K., Halka, J.P., and Ortt, R.A. Jr., (eds.),Tripping from the Fall Line: Field Excursions for the GSA Annual Meeting, Baltimore. Geological Society of America Field Guide, 40:171–229.
https://doi:10.1130/2015.0040(07)
Sawyer, G.T. 1981. A study of crocodilian coprolites from Wannagan Creek Quarry (Paleocene–North Dakota), Ichnofossils II, Minnesota. Scientific Publication of the Science Museum, 5(2):1–29.
Sawyer, G.T. 1998. Coprolites of the Black Mingo Group (Paleocene) of South Carolina. In Sanders, A.E. (ed.), Paleobiology of the Williamsburg Formation (Black Mingo Group; Paleocene) of South Carolina, U.S.A. Transactions of the American Philosophical Society, 88(4):221–228.
https://doi.org/10.2307/1006676
Schram, F.R. 1970. Isopod from the Pennsylvanian of Illinois. Science, 169:854–855.
https://doi.org/10.1126/science.169.3948.854
Schram, F.R., and S. Koenemann. 2021. Evolution and phylogeny of Pancrustacea. Oxford University Press, Oxford, UK.
Shelburne, E.C.H. and Thompson, A.C. 2016. Specimen whitening: An assessment of methods of ammonium chloride smoke removal. Collection Forum, 30(1):63–72.
https://doi.org/10.14351/0831-4985-30.1.63
Smit, N J., Bruce, N.L., and Hadfield, K.A. 2014. Global diversity of fish parasitic isopod crustaceans of the family Cymothoidae. International Journal for Parasitology: Parasites and Wildlife, 3(2):188–197.
https://doi.org/10.1016/j.ijppaw.2014.03.004
Souto, P.R.F. 2010. Crocodylomorph coprolites from the Bauru basin, Upper Cretaceous, Brazil. New Mexico Museum of Natural History and Science Bulletin, 51:201–208.
Tait, J. 1917. Experiments and observations on crustacea. Part II Moulting of isopods. Proceedings of the Royal Society of Edinburgh, 37:59–68.
Trilles, J.-P. 1999. Ordre des isopodes sous-ordre des épicarides (Epicaridea Latreille, 1825). In Forest, J. (ed.), Traité de Zoologie, Anatomie, Systémique, Biologie. Tome 7 Crustacés Fascicule 3A Péracarides. Mémories de l’Institut Océanographique, Monaco, 19:279–352.
Vogt, P., Eshelman, R.E., and Godfrey, S.J. 2018. Calvert Cliffs: eroding mural escarpment, fossil dispensary, and paleoenvironmental archive in space and time. Smithsonian Contributions to Paleobiology, 100:3–44.
https://lccn.loc.gov/2017059091
Wägele, J.W. 1989. Evolution und phylogenetisches System der Isopoda. Zoologica (Stuttgart), 140:1–262.
Ward, L.W. 1992. Molluscan biostratigraphy of the Miocene, Middle Atlantic Coastal Plain of North America. Virginia Museum of Natural History Memoir 2:1–159.
Ward, L.W. and Andrews, G.W. 2008. Stratigraphy of the Calvert, Choptank, and St. Marys Formations (Miocene) in the Chesapeake Bay area, Maryland and Virginia. Virginia Museum of Natural History Memoir, 9:1–169.
Weems, R.E. 2018. Crocodilians of the Calvert Cliffs. Smithsonian Contributions to Paleobiology, 100:213–240.
https://lccn.loc.gov/2017059091
Weems, R.E. and Edwards, L.E. 2007. Post-Middle Miocene origin of modern landforms in the Eastern Piedmont of Virginia. Stratigraphy, 4(1):35–48.
Weems, R.E., Edwards, L.E., and Landacre, B. 2017. Geology and biostratigraphy of the Potomac River cliffs at Stratford Hall, Westmoreland County, Virginia. In Bailey, C.M. and Jaye, S. (eds.), From the Blue Ridge to the Beach: Geological Field Excursions across Virginia. Geological Society of America Field Guide, 47:125–152.
https://doi:10.1130/2017.0047(05)
Wetmore, A. 1943. The occurrence of feather impressions in the Miocene deposits of Maryland. The Auk, 60:440–441.
Wieder, R.W. and Feldmann, R.M. 1992. Mesozoic and Cenozoic fossil isopods of North America. Journal of Paleontology, 66:958–972.
https://www.jstor.org/stable/1305950
WoRMS Editorial Board (2023). World register of marine species. Available from https://www.marinespecies.org at VLIZ. Accessed 2023-12-29.
https://doi.org/10.14284/170
