

 The morphological diversity and distribution of the genus Menispermites (Magnoliopsida) in the Cretaceous of Northern Asia
The morphological diversity and distribution of the genus Menispermites (Magnoliopsida) in the Cretaceous of Northern Asia
Article number: 28.1.a9
https://doi.org/10.26879/1441
Copyright Palaeontological Association, February 2025
Author biographies
Plain-language and multi-lingual abstracts
PDF version
Submission: 21 August 2024. Acceptance: 5 February 2025.
ABSTRACT
Eleven species of Menispermites from Kazakhstan, Siberia, North-East of Russia and Sakhalin distributed from the upper Albian up to Maastrichtian–Danian were restudied based on original materials and published data to analyze their morphological diversity and stratigraphic and geographical occurrence. English diagnoses of five species are presented for the first time. A new species, M. temlyanensis Zolina, Golovneva et Grabovskiy, sp. nov. from the Maastrichtian–Danian Tanyurer Formation of Chukotka is described. The greatest number of species is confined to the North-East of Russia. In this territory, short-lived species with limited ranges predominated. M. sibiricus had the widest geographical and stratigraphic range: from Kazakhstan to Chukotka and from the Cenomanian to Coniacian. In other regions of Northern Asia, the stratigraphic distribution of Menispermites was limited. In Kazakhstan and Siberia, it is known from the Cenomanian to Turonian. On Sakhalin, Menispermites was recorded from the Campanian to the Maastrictian–Danian. The greatest number of species and morphological diversity of Menispermites occured in the Cenomanian–Turonian of the North-East of Russia.
A.A. Zolina. Komarov Botanical Institute, Russian Academy of Sciences, 197022, prof. Popov str. 2, St. Petersburg, Russia. azolina@binran.ru
L.B. Golovneva. Komarov Botanical Institute, Russian Academy of Sciences, 197022, prof. Popov str. 2, St. Petersburg, Russia. golovneva@binran.ru
A.A. Grabovskiy. Komarov Botanical Institute, Russian Academy of Sciences, 197022, prof. Popov str. 2, St. Petersburg, Russia. agrabovskiy@binran.ru
Keywords: new species; Menispermites; Menispermaceae; Ranunculales; North-East of Russia; paleogeography
Final citation: Zolina, A.A., Golovneva, L.B., and Grabovskiy, A.A. 2025. The morphological diversity and distribution of the genus Menispermites (Magnoliopsida) in the Cretaceous of Northern Asia. Palaeontologia Electronica, 28(1):a9.
https://doi.org/10.26879/1441
palaeo-electronica.org/content/2025/5451-menispermites-in-northern-asia
Copyright: February 2025 Palaeontological Association.
This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
creativecommons.org/licenses/by/4.0
INTRODUCTION
The genus Menispermites Lesquereux was described from the Albian–Cenomanian Dakota Formation, USA (Lesquereux, 1874). The leaves are characterized by lobate or unlobate, usually peltate leaves with actinodromous venation and numerous primary veins. Menispermites originally included three species (Lesquereux, 1874). One of them, M. obtusilobus Lesquereux, was chosen later as a type species (Knowlton, 1919). This species has broadly ovate, trilobate peltate leaves with obtuse apex, low pelta, truncate base and crenate margin.
Menispermites were widely distributed in the Late Cretaceous and Paleogene. Nowadays this genus includes more than 30 fossil species. About 15 of them are known from North and South America (Lesquereux, 1874, 1892; Hollick, 1930; Berry, 1916; Hollick, 1930; Bell, 1956, 1962; Doyle and Hickey, 1976; Doria, et al., 2008; Guzmán-Vázquez et al., 2018; Del Rio et al., 2021). In the Upper Cretaceous deposits of Europe, the genus was represented by M. summesbergeri Herman et J. Kvaček from the lower Campanian Grünbach Formation (Herman and Kvaček, 2010). More than 10 species were described from Northern Asia (Vachrameev, 1952; Kryshtofovich and Baikovskaya, 1960; Budantsev, 1968; Zhilin, 1974; Krassilov, 1979; Shilin, 1986; Herman and Lebedev, 1991; Philippova and Abramova, 1993; Golovneva, 2006; Golovneva et al., 2011; Moiseeva, 2014; Golovneva, 2019). The greatest number of Menispermites species comes from the Albian and Cenomanian. The latest Cretaceous and Paleogene representatives are less numerous.
Menispermites have a significant morphological resemblance to leaves of modern genus Menispermum L. (Menispermaceae, Ranunculales), that are characterized by a mostly entire margin, actinodromous primary and brochidodromous secondary venation (Ortiz et al., 2007; Doria et al., 2008).
The oldest clearly attributed fossil evidence of Menispermaceae is an endocarp of the extant genus Stephania Loureiro found in the lower Paleocene of Patagonia (Jud et al., 2018). Endocarps of different Menispermaceae (Stephania, Palaeosinomenium Chandler and Tinospora Miers) were also found in the lower Paleocene of Paris Basin, Europe (Kara et al., 2023). Since the Eocene, endocarps of Menispermaceae are abundant in Europe and North America (Chandler, 1964; Collinson, 1983; Manchester, 1994; Mai, 1995; Jacques and De Franceschi, 2005). The earliest record, which was interpreted as Menispermaceae wood, comes from the latest Cretaceous–early Paleocene Deccan Intertrappean beds, India (Bonde, 1997). This material is characterized by alternating rings of phloem and xylem that resemble modern lianas.
Therefore, the Paleogene records of Menispermites are usually assigned to the family Menispermaceae and this attribution is confirmed by findings of corresponding reproductive structures in the same deposits (Doria, et al., 2008; Guzmán-Vázquez et al., 2018; Del Rio et al., 2021). But the attribution of Cretaceous finds of Menispermites to Menispermaceae is debatable. Indeed, the leaves of Menispermites are also similar to leaves of some modern representatives of Nymphaeaceae and Piperaceae. However, the Cretaceous representatives of these families differ rather well from Menispermites. Besides, the relationships of the Cretaceous finds with modern families Nymphaeaceae and Piperaceae is also questionable, as is the relationships of Menispermites and Menispermaceae. Thus, some authors consider Menispermites as a morphological taxon of unclear systematic position (Herman et al., 2016).
The first endocarps with affinity to Menispermaceae were recorded from the Turonian of Europe (Knobloch and Mai, 1984) and were attributed to the genus Prototinomiscium Knobloch et Mai. However, endocarps of Prototinomiscium do not have a well-developed condyle, and this makes the assignment of this genus to Menispermaceae doubtful (Herrera et al., 2011; Wefferling et al., 2013).
Actually, many Cretaceous genera have characters that preclude their direct assignment to the extant families. For example, small actinomorphic pistillate flowers of Callicrypta chlamydea Krassilov et Golovneva from the Cenomanian Timmerdyakh Formation of the Lena-Vilyui depression are characterized by a significant similarity to the flowers of Menispermaceae but also share diagnostic features with several other ranunculid families (Krassilov and Golovneva, 2004). Floral and fruit morphology of the early eudicot Ranunculaecarpus quinquecarpellatus Samylina from the lower-middle Albian Buor-Kemyus Formation of the North-East of Russia support possible affinity of this species to extant Ranunculaceae, but it is distinguished from modern members of the family by some features of the perianth and pollen (Manchester et al., 2018).
Similarity of Menispermites with Menispermaceae in leaf morphology and association of Paleogene representatives of Menispermites with reproductive structures of Menispermaceae suggest that the Cretaceous representatives of this genus could also be related to ancestral forms of Menispermaceae. Their detailed study could allow us to consider their taxonomic affinity more precisely. Unfortunately, many Cretaceous species, especially from North America, are represented by fragmentary material, have nomenclature problems and need reinvestigation. The Cretaceous records of Menispermites from Northern Asia are represented by numerous whole leaves. In this paper, we attempt to analyze changes in the morphological diversity and stratigraphic and geographical distribution of the genus Menispermites at the end of the Early Cretaceous and during the Late Cretaceous. In addition, we describe a new species, M. temlyanensis Zolina, Golovneva et Grabovskiy, sp. nov. from the Maastrichtian–Danian Tanyurer Formation of Chukotka.
MATERIAL AND METHODS
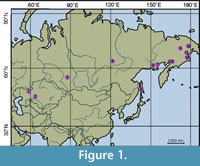 Species of Menispermites from Northern Asia were restudied based on original material and published data. Full and abbreviated names of institutions, where studied specimens of Menispermites are stored, are given in Table 1, and all localities are shown in Figure 1.
Species of Menispermites from Northern Asia were restudied based on original material and published data. Full and abbreviated names of institutions, where studied specimens of Menispermites are stored, are given in Table 1, and all localities are shown in Figure 1.
Four species of Menispermites come from the upper Albian–lower Turonian Krivorechenskaya Formation, Grebenka River, Anadyr River basin (Philippova and Abramova, 1993; Golovneva et al., 2015): M. minutus (Kryshtofovich) Shczepetov, Herman et Belaya, M. vasetskii Philippova, M. marcovoensis Philippova, and M. orientalis Golovneva. The last species was also found in the upper Turonian–Coniacian Zarya and Arman formations of the Magadan Region, and in the lower Coniacian Poperechnenskaya Formation, Pekulney Ridge, Chukotka (Philippova, 2010; Golovneva et al., 2015).
M. ginterensis Herman is present in the Cenomanian Ginter Formation in the northeastern part of the Koryak Upland (Herman, 1988; Moiseeva, 2010).
Leaves of M. sibiricus (Heer) Golovneva were originally described from the Cenomanian–Turonian Simonovo Formation of Eastern Siberia under the name Acer sibiricum Нееr (Heer, 1878; Golovneva, 2006; Golovneva and Nosova, 2012). Later, this species was also found in the Cenomanian Timmerdyakh Formation of the Lena-Vilyui depression (Golovneva, 2006), in the Cenomanian Altykuduk Formation and in the Turonian Zhirkindek Formation of Kazakhstan (Vachrameev, 1952; Shilin, 1986; Golovneva, 2006), in the upper Turonian–Coniacian Valizhgen Formation of northwestern Kamchatka (Herman and Lebedev, 1991), and in the upper Turonian–Coniacian Chingandzha and Arman formations of the Magadan Region (Golovneva et al., 2011; Herman et al., 2016; Golovneva, 2019), In this paper, we also referred to this species leaves from the Coniacian Poperechnenskaya Formation.
Two species of Menispermites were described from Sakhalin Island. М. sachalinensis Kryshtofovich ex Zhilin comes from the Campanian Zhonk’er Formation (Kryshtofovich and Baikovskaya, 1960; Zhilin, 1974; Krassilov, 1979). М. favosus Krassilov was reported from the Maastrichtian–Danian Boshnyakovo Formation (Vachrameev, 1966; Krassilov, 1979).
M. lebedevii Moiseeva comes from the Santonian–Campanian Emuneret Formation, Enmyvaam River, Chukotka. A leaf fragment of Menispermites, described as М. cf. sachalinensis (Moiseeva and Sokolova, 2007) was found from the Campanian Barykov Formation of the Ugolnaya Bay, North-East of Russia.
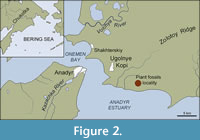 Leaves of the new species, M. temlyanensis, sp. nov., were collected by A. Grabovskiy from the Tanyurer Formation, distributed in the northern part of the Pekulney Ridge and between the Pekulney Ridge and the Zolotoy Ridge near Anadyr town, Chukotka. This formation is 550–2200 m thick and consists of andesites, pyroxene and olivine basalts, tuffs and volcano-sedimentary deposits (Zvizda et al., 1974). In the lower part of the section, there are beds of conglomerates with interlayers of coaly siltstones and grey sandstones. Plant fossils are confined to the volcano-sedimentary layers in the base of the Tanyurer Formation, which are exposed in the road quarry near Ugolnye Kopi settlement (Figure 2).
Leaves of the new species, M. temlyanensis, sp. nov., were collected by A. Grabovskiy from the Tanyurer Formation, distributed in the northern part of the Pekulney Ridge and between the Pekulney Ridge and the Zolotoy Ridge near Anadyr town, Chukotka. This formation is 550–2200 m thick and consists of andesites, pyroxene and olivine basalts, tuffs and volcano-sedimentary deposits (Zvizda et al., 1974). In the lower part of the section, there are beds of conglomerates with interlayers of coaly siltstones and grey sandstones. Plant fossils are confined to the volcano-sedimentary layers in the base of the Tanyurer Formation, which are exposed in the road quarry near Ugolnye Kopi settlement (Figure 2).
The meaning of Latin abbreviations using in chapter “Systematic Palaeontology” is given in Table 2. The terminology follows Ellis et al. (2009), except we use 'entire' in the sense of being unlobed. Photographs of the specimens were made using a Canon 6D camera with low angle illumination.
Registry
New species name Menispermites temlyanensis Zolina, Golovneva et Grabovskiy, described in this paper, is registered with a unique PFN number in the Plant Fossil Names Registry, hosted and operated by the National Museum, Prague for the International Organisation of Palaeobotany.
SYSTEMATIC PALAEONTOLOGY
Class MAGNOLIOPSIDA Brongniart, 1843
MAGNOLIOPSIDA incertae sedis
Genus MENISPERMITES Lesquereux, 1874
Menispermites sibiricus (Heer) Golovneva
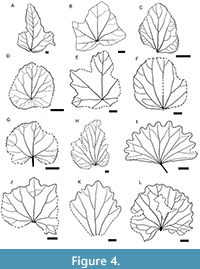
Figure 3A–G, Figure 4E
1878 Acer sibiricum Heer, p. 46, pl. 10, figure 4b, 5a, pl. 12, figure 1b.
1878 Nymphaeites tener Heer, p. 44, pl. 13, figure 7.
1952 Menispermites kryshtofovichii Vachrameev, p. 194, pl. 14, figure 5, 6, pl. 15, figure 6, text-figure 41.
1955a Menispermites sibiricus (Heer) I. Lebedev non rite publ., p. 199, pl. 30, figure 1.
1955b Cissites sibirensis I. Lebedev, p. 203, pl. 31, figure 4.
1962 Menispermites sibiricus (Heer) I. Lebedev non rite publ. p. 258, pl. 49, figure 1.
1962 Menispermites simonovskiensis I. Lebedev, p. 258, pl. 65, figure 2.
1962 Menispermites tschulimensis I. Lebedev, p. 258, pl. 49, figure 5.
1968 Menispermites ribesifolius Budantsev, p. 11, pl. 2, figure 3.
1968 Menispermites stellatus Budantsev, p. 12, pl. 2, figure 4.
1974 Menispermites kryshtofovichii Zhilin, p. 99, pl. 37, figure 5, pl. 40, figure 7.
1974 Protoacerophyllum sibiricum (Heer) Iljinskaja, p. 151, text-figure 96, figure 4.
1975 Menispermites sp., Philippova, p. 64, pl. 9, fig. 1; text-figure 3.
1986 Menispermites kryshtofovichii Shilin, pl. 8, figure 2, 4, pl. 9, figure 1.
1986 Menispermites syrdariensis Shilin, p. 113, pl. 9, figure 3, pl. 10, figure 4.
1991 Menispermites kryshtofovichii Vachrameev; Herman and Lebedev, p. 67, pl. 2, figure 5, text-figure 13.
2006 Menispermites sibiricus (Heer) Golovneva, p. 1733, pl. 1, figure 1–3; 2010 Cissites cf. inscissus auct. non Herman; Philippova, pl. 28, figure 2.
2012 Menispermites sibiricus (Heer) Golovneva; Golovneva and Nosova, p. 115, pl. 43, figure 1–7, pl. 84, figure 5–9, text-figure 8.1, 8.2.
2016 Menispermites sibiricus (Heer) Golovneva; Herman et al., p. 727, pl. 23, figure 10, 11, text-figure 18.
2019 Menispermites sibiricus (Heer) Golovneva, p. 66, pl. 41, figure 1–5.
 Lectotype (designated by Golovneva, 2006). Specimen GIN 1800/11, Eastern Siberia, Chulym River basin, Simonovo Formation, Cenomanian–Turonian; figure 3C.
Lectotype (designated by Golovneva, 2006). Specimen GIN 1800/11, Eastern Siberia, Chulym River basin, Simonovo Formation, Cenomanian–Turonian; figure 3C.
Diagnosis (Golovneva, 2006). Leaves rounded or broadly ovate in outline, usually broader than long, 3–7 lobate or entire, with peltate, rounded, cordate or truncate base and broadly triangular obtuse apex; lobes radially divergent, decreasing downwards, with additional secondary lobes or lobe-like teeth, which are rounded or triangular with obtuse apex; sinuses between lobes rounded; margin entire or irregularly coarsely dentate, varying in different leaves; venation palmate with 5–7 straight basal veins, which have several lateral branches on each side in the distal part; basal veins usually craspedodromous, other ones form loops near the margin; peltate base innervated by 2–5 thin veins, running from the beginning of basal veins.
Range and occurrence. Kazakhstan, Altykuduk Formation, Cenomanian, Zhirkindek Formation, Turonian; Western Siberia, Simonovo Formation, Cenomanian–Turonian; Eastern Siberia, Timmerdyakh Formation, Cenomanian; North-East of Russia, Valizhgen, Chingandzha and Arman formations, upper Turonian–Coniacian, Poperechnenskaya Formation, lower Coniacian.
Menispermites minutus (Kryshtofovich) Shczepetov, Herman et Belaya
Figure 4G, Figure 5D, F
1958 Menispermites septentrionalis auct. non Hollick; Kryshtofovich, p. 53, pl. 11, figure 2, 3.
1958 Populus minuta Kryshtofovich, p. 42, text-figure 19.
1992 Menispermites ex gr. septentrionalis Hollick; Shczepetov et al., pl. 41, figure 2, pl. 42, figure 1, pl. 44, figure 1, pl. 49, figure 1, pl. 50, figure 3, 4, pl. 59, figure 4, 5.
1992 Menispermites minutus (Kryshtofovich) Shczepetov, Herman et Belaya, p. 44, pl. 41, figure 6.
1993 Menispermites septentrionalis auct. non Hollick; Philippova and Abramova, pl. 31, figure 1–3. 2015 Menispermites minutus Golovneva et al., p. 431, pl. 7, figure 1–7, pl. 8, figure 1–3.
Neotype (designated by Golovneva et al., 2015). Specimen NEISRI PF1/688a, North-East of Russia, Anadyr River basin, Grebenka River, Krivorechenskaya Formation, upper Albian–lower Turonian; Golovneva et al., 2015, pl. 7, figure 4.
 Emended diagnosis (Golovneva et al., 2015). Leaves entire, broadly ovate, sometimes with lobe-like extensions in upper part, with cordate, rounded or truncate base and broadly triangular obtuse or attenuate apex; margin dentate; teeth broadly triangular, with concave sides, an acute apex with glands and wide low sinuses; venation palmate, craspedodromous, with 7⎯9 basal veins; midvein straight with 2–3 pairs of secondary branches in upper part; other basal veins curved, decreasing downwards, with 2–3 basi- and 1–3 acroscopic branches or dichotomizing 1–3 times.
Emended diagnosis (Golovneva et al., 2015). Leaves entire, broadly ovate, sometimes with lobe-like extensions in upper part, with cordate, rounded or truncate base and broadly triangular obtuse or attenuate apex; margin dentate; teeth broadly triangular, with concave sides, an acute apex with glands and wide low sinuses; venation palmate, craspedodromous, with 7⎯9 basal veins; midvein straight with 2–3 pairs of secondary branches in upper part; other basal veins curved, decreasing downwards, with 2–3 basi- and 1–3 acroscopic branches or dichotomizing 1–3 times.
Range and occurrence. North-East of Russia, Krivorechenskaya Formation, upper Albian–lower Turonian.
Menispermites orientalis Golovneva
Figure 4L, Figure 5G, H
1993 Menispermites septentrionalis auct. non Hollick; Philippova and Abramova, pl. 83, figure 1.
 2010 Menispermites efimovae Philippova, non rite publ., p. 94, pl. 17, figure 5, 6.
2010 Menispermites efimovae Philippova, non rite publ., p. 94, pl. 17, figure 5, 6.
2015 Menispermites orientalis Golovneva; Golovneva et al., p. 434, pl. 8, figure 4–6.
2016 Menispermites orientalis Golovneva; Herman et al., p. 726, pl. 23, fig. 9, text-figure 17.
Holotype. Specimen NEISRI PF1/505; North-East of Russia, Anadyr River basin, Grebenka River, Krivorechenskaya Formation, upper Albian–lower Turonian Golovneva et al., 2015, pl. 8, figure 4.
Diagnosis (Golovneva et al., 2015). Leaves entire, broadly ovate, with cordate base and obtuse or rounded apex; margin crenate; teeth rounded or broadly triangular, with convex sides and rounded apex with glands; venation palmate, craspedodromous, with 9 basal veins; midvein straight with 1–2 pairs of secondary branches; other basal veins curved, decreasing downwards, with 2–3 basiscopic branches or dichotomizing 1–2 times.
Range and occurrence. North-East of Russia, Krivorechenskaya Formation, upper Albian–lower Turonian, Zarya and Arman formations, upper Turonian–Coniacian, Poperechnenskaya Formation, lower Coniacian.
Menispermites vasetskii Philippova
Figure 4K
1993 Menispermites vasetskii Philippova; Philippova and Abramova, p. 122, pl. 32, fig. 1, pl.38, figure 5, 6, pl. 40, figure 4, pl. 56, figure 5, pl. 57, figure 1, 2.
Holotype. Specimen TFGI 622-1-40/225a, North-East of Russia, Bystraya River, Krivorechenskaya Formation, upper Albian–lower Turonian; Philippova and Abramova, 1993, pl. 32, fig. 1.
Diagnosis (presented here). Leaves entire, broadly ovate, with cordate or truncate base and rounded or acute apex; margin dentate; teeth evenly spaced, high, triangular, with rounded apex; sinuses rounded; venation palmate, craspedodromous, with 5–7 basal veins, midvein with 1–2 pairs of dichotomizing secondary branches in upper part; lateral veins dichotomously branching 2–3 times.
Range and occurrence. North-East of Russia, Krivorechenskaya Formation, upper Albian–lower Turonian.
Menispermites marcovoensis Philippova
Figure 4I, Figure 5E
1978 Menispermites marcovoensis Philippova, p. 141, pl. 10, figure 1, 2.
1993 Menispermites marcovoensis Philippova; Philippova and Abramova, p. 121, pl. 32, figure 3–5, pl. 56, figure 4.
2010 Menispermites marcovoensis; Philippova Moiseeva, pl. 15, figure 12, 13, text-figure 3l–3n.
Holotype. Specimen NEISRI PF1/505; North-East of Russia, Anadyr River basin, Grebenka River, Krivorechenskaya Formation, upper Albian–lower Turonian; Golovneva et al., 2015, pl. 8, figure 4.Holotype. Specimen NEISRI PF1/505; North-East of Russia, Anadyr River basin, Grebenka River, Krivorechenskaya Formation, upper Albian–lower Turonian; Golovneva et al., 2015, pl. 8, figure 4.
Diagnosis (presented here). Leaves entire, fan-shaped, usually the width is twice the length, with broadly cuneate or truncate base and obtuse rounded apex; margin dentate; teeth evenly spaced, high, broadly triangular with rounded apex; sinuses rounded; venation palmate, craspedodromous, with 7–11 basal veins dichotomously branching in upper part.
Range and occurrence. North-East of Russia, Krivorechenskaya Formation, upper Albian–lower Turonian, Ginter Formation, Cenomanian.
Menispermites ginterensis Herman
Figure 4C
2010 Menispermites ginterensis Herman, 1988, p. 112, text-fig. 2, 3. - Moiseeva, pl. 15, fig. 11, text-figure 4g.
Holotype. Specimen GIN 3385/203, North-East of Russia, Koryak Upland, Ugol’naya Bay, Ginter Formation, Cenomanian; Herman, 1988, text-figure 2, 3.
Diagnosis (presented here). Leaves entire, widely ovate, with truncate base and obtuse apex; margin dentate; teeth small, no evenly spaced, triangular, with acute or rounded apex; sinuses rounded; venation palmate, brochidodromous or semi craspedodromous in lower part of lamina and craspedodromous in its upper part, with 5 basal veins; midvein with 3 pairs of secondary branches in upper part; lower basal veins curved, with several basiscopic branches.
Range and occurrence. North-East of Russia, Ginter Formation, Cenomanian.
Menispermites lebedevii Moiseeva
Figure 4D
2014 Menispermites lebedevii Moiseeva, p. 681, pl. 15, figure 1–5, 9.
Holotype. Specimen GIN 3392/68, North-East of Russia, Enmyvaam River, Emuneret Formation, Santonian–Campanian; Moiseeva, 2014, pl. 15, fig. 1, 5, text-figure 3b.
Diagnosis (Moiseeva, 2014). Leaves entire, rounded or ovate, slightly asymmetric; leaf base peltate eccentric; leaf apex rounded; margin irregularly dentate, partly undulate; teeth usually flat, rounded-triangular with rounded or more rarely acute apex; venation palmate, craspedodromous or semicraspedodromous, with 7–9 basal veins decreasing downwards; midrib prominent, with 2–3 pairs of secondary veins; lower basal veins thin and short, running to base of leaf lamina into pelta.
Range and occurrence. North-East of Russia, Emuneret Formation, Santonian–Campanian.
Menispermites sachalinensis Kryshtofovich
ex Zhilin
Figure 4J, Figure 5C
1918 Hedera macclurii auct non Heer; in Kryshtofovich, p. 59, text-figure 14.
1960 Menispermites sachalinensis Kryshtofovich, non rite publ., designat. typi omissa; Kryshtofovich and Baikovskaya, p. 81, pl. 17, fig. 2, text-figure 28.
1974 Menispermites sachalinensis Kryshtofovich ex Zhilin; Zhilin p. 100, pl. 38, figure 6.
1979 Menispermites sachalinensis Kryshtofovich ex Zhilin; Krassilov, p. 108, pl. 30, figure 11.
Lectotype (designated by Zhilin, 1974). Specimen TSNIGRM 6256/149, Sakhalin, Cap Jonquiere, Zhonk’er Formation, Campanian; Zhilin, 1974, pl. 38, figure 6.
Diagnosis (presented here). Leaves entire, rounded in outline, with cordate base, apex acute with big rounded tooth in the top; margin dentate or undulate; teeth large, distantly spaced, broadly triangular with rounded apices, different in size; sinuses rounded; venation palmate, craspedodromous, with 5–7 basal veins; midvein with 1–2 pairs of secondary veins in upper part; lateral veins producing 1–3 basiscopic branches or dichotomizing.
Range and occurrence. Sakhalin, Zhonk'er Formation, Campanian.
Menispermites favosus Krassilov
Figure 4F, Figure 5A, B
1979 Menispermites favosus Krassilov, p. 108, pl. 31, figure 1.
Holotype. Specimen FSCEATB 330/377, Sakhalin, Avgustovka River, Boshnyakovo Formation, Maastrichtian–Danian; Krassilov, 1970, pl. 31, figure 1.
Diagnosis (presented here). Leaves entire, broadly ovate; base rounded or truncate, peltate or nonpeltate; pelta (if present) small, 2–3 mm high; apex rounded with small triangular tooth in top; margin undulate; venation palmate, craspedodromous, with 5–7 basal veins; midvein producing 2–3 pairs of secondary veins; outer lateral basal veins providing 3–5 basiscopic brochidodromous branches.
Range and occurrence. Sakhalin, Boshnyakovo Formation, Danian.
Menispermites temlyanensis Zolina, Golovneva et Grabovskiy, sp. nov.
Figure 6A–E, Figure 7A–E, Figure 8A–F
Plant Fossil Registration number PFN003476
Etymology. After Temlyan Mountain.
 Holotype (designated here). Specimen BIN 1588/263, North-East of Russia, Anadyr district of Chukotka, Tanyurer Formation, Maastrichtian–Danian; figure 6A, 8B.
Holotype (designated here). Specimen BIN 1588/263, North-East of Russia, Anadyr district of Chukotka, Tanyurer Formation, Maastrichtian–Danian; figure 6A, 8B.
Material. Collection BIN 1588, specimens 218, 221, 241, 250–266, 410, 411.
Diagnosis. Leaves trilobate or unlobate, broadly ovate with rounded, peltate base with high pelta; lobes (if presents) small, broadly triangular with rounded apices; central lobe bigger than lateral ones; apex of unlobate leaves attenuate; margin entire; venation palmate, with 3–5 basal veins; midvein producing 3–5 pairs of secondary veins; inner lateral veins straight, terminated in the lobes apices; outer lateral basal veins curved, connected with inner lateral veins; pelta innervated by 2–4 pairs of thin veins.
 Description. Leaves are simple, 3–12 cm in length and 3–9 cm in wide. The leaf lamina is broadly ovate in outlines, trilobate, more rarely unlobate. Lobes (if presents) are small, broadly triangular with rounded apices and strait or slightly convex sides (Figure 6A, D, Figure 7C, Figure 8B, C, E). Central lobe is significantly bigger than lateral ones. Unlobate leaves have attenuate apex with rounded tip (Figure 7A, B). The leaf base is rounded, peltate. The height of the pelta ranges from one quarter to one third of the leaf length. The margin is entire (Figure 6C).
Description. Leaves are simple, 3–12 cm in length and 3–9 cm in wide. The leaf lamina is broadly ovate in outlines, trilobate, more rarely unlobate. Lobes (if presents) are small, broadly triangular with rounded apices and strait or slightly convex sides (Figure 6A, D, Figure 7C, Figure 8B, C, E). Central lobe is significantly bigger than lateral ones. Unlobate leaves have attenuate apex with rounded tip (Figure 7A, B). The leaf base is rounded, peltate. The height of the pelta ranges from one quarter to one third of the leaf length. The margin is entire (Figure 6C).
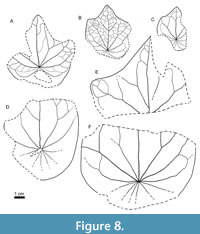 Venation is palmate, with 3–5 basal veins. The midvein is straight or slightly curved. It is thicker and longer that other veins, running to the leaf apex, producing 3–5 pairs of alternate or subopposite secondary veins in the upper one-quarter of the lamina. These secondary veins are connected to each other and form series of loops near leaf apex. Inner basal lateral veins are thick, straight, diverging from the midvein at an angle 35–45°, running to the lobe’s apices. They produce 1–3 pairs of short curved brochidodromous secondary veins in upper part, which innervate lobes. The lowest acroscopic secondary veins are connected with the lowest secondary veins extending from the midvein. Outer lateral basal veins are curved, thinner and shorter than inner lateral basal veins. They diverge from the midvein at an angle about 75–85° and join with inner lateral veins near the leaf margin. They provide 3–5 thin basiscopic brochidodromous branches. Pelta is innervated by 2–4 pairs of thin, radiating, dichotomously branching veins (Figure 6A, B, E, Figure 7D, B, E, Figure 8B, D, F). Tertiary veins form an alternating series of meshes (Figure 6C, Figure 8B).
Venation is palmate, with 3–5 basal veins. The midvein is straight or slightly curved. It is thicker and longer that other veins, running to the leaf apex, producing 3–5 pairs of alternate or subopposite secondary veins in the upper one-quarter of the lamina. These secondary veins are connected to each other and form series of loops near leaf apex. Inner basal lateral veins are thick, straight, diverging from the midvein at an angle 35–45°, running to the lobe’s apices. They produce 1–3 pairs of short curved brochidodromous secondary veins in upper part, which innervate lobes. The lowest acroscopic secondary veins are connected with the lowest secondary veins extending from the midvein. Outer lateral basal veins are curved, thinner and shorter than inner lateral basal veins. They diverge from the midvein at an angle about 75–85° and join with inner lateral veins near the leaf margin. They provide 3–5 thin basiscopic brochidodromous branches. Pelta is innervated by 2–4 pairs of thin, radiating, dichotomously branching veins (Figure 6A, B, E, Figure 7D, B, E, Figure 8B, D, F). Tertiary veins form an alternating series of meshes (Figure 6C, Figure 8B).
Remarks. The type species of the genus Menispermites, M. obtusilobus (Figure 4A) from the Dakota Formation has broadly ovate, trilobate peltate leaves with obtuse apex, low pelta (3–4 mm), truncate base and crenate or undulate margin (Lesquereux, 1874). M. temlyanensis is similar to the type species in its trilobate shape and in the presence of a pelta. The new species differs from M. obtusilobus by its small lobes, high pelta (2–4 cm) and entire margin. Two species M. salinensis Lesquereux and M. acerifolius Lesquereux were described from the Dakota Formation together with the type species (Lesquereux, 1874). Both species are characterized by nonpeltate leaves.
Most species of Menispermites have entire leaves. Since the leaves of the new species are trilobate, they have the greatest resemblance to M. sibiricus. This species has broadly ovate 3–5 lobate leaves, with rounded, cordate or truncate base, broadly triangular obtuse lobes and dentate margin (Figure 4). Both species are characterized by well-developed pelta. However, M. temlyanensis is distinguished from M. sibiricus by less developed lobes and by entire margin
Range and occurrence. North-East of Russia, Tanyurer Formation, Maastrichtian–Danian.
Menispermites sp.
Figure 4H
2007 Menispermites cf. sachalinensis Kryshtofovich ex Zhilin; Moiseeva and Sokolova, pl. 4, figure 12, text-figure 6i.
Range and occurrence. North-East of Russia, Barykov Formation, Campanian.
DISCUSSION
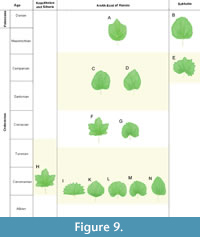 The earliest occurrences of Menispermites in the North-East of Russia appeared in the upper Albian–lower Turonian Krivorechenskaya Formation. Here four species were recorded: M. minutus, M. vasetskii, M. marcovoensis, and M. orientalis. The first three of them were found only in this formation. Thus, they are endemic to the Grebenka flora (Shczepetov et al., 1992; Philippova and Abramova, 1993), while M. orientalis was also discovered in several late Turonian–Coniacian floras in the North-East of Russia. The leaves of these four species are rather diverse in morphology, from fan-shaped with truncate base to widely ovate with deeply cordate base. However, all of them are entire (have no lobes) and have no pelta (Figure 9).
The earliest occurrences of Menispermites in the North-East of Russia appeared in the upper Albian–lower Turonian Krivorechenskaya Formation. Here four species were recorded: M. minutus, M. vasetskii, M. marcovoensis, and M. orientalis. The first three of them were found only in this formation. Thus, they are endemic to the Grebenka flora (Shczepetov et al., 1992; Philippova and Abramova, 1993), while M. orientalis was also discovered in several late Turonian–Coniacian floras in the North-East of Russia. The leaves of these four species are rather diverse in morphology, from fan-shaped with truncate base to widely ovate with deeply cordate base. However, all of them are entire (have no lobes) and have no pelta (Figure 9).
M. ginterensis Herman was found in the Cenomanian Ginter Formation and is not recorded in other places.
In western Kazakhstan and Siberia, Menispermites appeared in the Cenomanian and is represented by M. sibiricus. This species is characterized by very variable peltate and lobate leaves with entire or dentate margin. It persisted in Kazakhstan and Siberia up to the Turonian and appeared in the North-East of Russia in the upper Turonian–Coniacian.
After the Turonian, Menispermites did not occur in Kazakhstan and Siberia. In the North-East of Russia two species of Menispermites were found in the Santonian and Campanian: M. lebedevii from the Emuneret Formation and Menispermites sp. from the Barykov Formation (Figure 9). The first species is characterized by entire ovate leaves with dentate margin and high pelta, and second species had also entire ovate leaves, but with entire margin and nonpeltate.
In the Campanian Menispermites first appears on Sakhalin, where two species are known. М. sachalinensis, with entire widely ovate nonpeltate leaves, was described from the Campanian Zhonk'er Formation. The second species, М. favosus, with entire ovate leaves and with very small pelta, was found in the Maastrichtian–Danian Boshnyakovo Formation.
At the same time, M. temlyanensis was distributed in the North-East of Russia. Its lobate leaves with very high pelta and entire margin were found in the Maastrichtian–Danian Tanyurer Formation. Later findings of Menispermites in Northern Asia are not known. Menispermites from Northern Asia are characterized by significant morphological diversity.
Among the representatives of Menispermites from Northern Asia, four species have a pelta: M. sibiricus, M. lebedevii, M. temlyanensis, and М. favosus. This suggests that they may have been lianas like many modern Menispermaceae, which also have peltate leaves (Givnish and Vermeij, 1976). Leaves with this morphological feature are observed from the Cenomanian to the Maastrichtian–Danian. Nowadays, lianas are mainly distributed in tropical climates, although they are also found in both subtropical and temperate climates. During the Late Cretaceous, the climate in the different parts of Northern Asia was humid, subtropical or temperate (Moiseeva and Sokolova, 2007; Spicer and Herman, 2010; Zolina et al., 2020). The analysis of fossil floras suggests that polydominant deciduous forests with an admixture of evergreen species were widespread in this territory (Spicer and Herman, 2010; Golovneva and Nosova, 2012; Golovneva, 2019).
Thus, Menispermites in Northern Asia is known from the late Albian up to the Maastrichtian–Danian. The greatest number of species is confined to the North-East of Russia.
In other regions of Northern Asia, stratigraphic distribution of Menispermites was limited. In Kazakhstan and Siberia, it is known from the Cenomanian to Turonian. On Sakhalin, Menispermites is known from the Campanian to the Maastrictian–Danian.
Eleven species of Menispermites are known from Northern Asia. Among them, M. sibiricus had the widest geographical and stratigraphic distribution. The other species of the genus had narrow geographic and stratigraphic ranges.
In other regions of Eurasia, the species diversity of Menispermites was significantly lower. Only one species was described from the Upper Cretaceous of Europe — M. summesbergeri from the Campanian Grünbach Formation (Herman and Kvaček, 2010). In North America, the greatest species diversity of the genus occurs in the late Albian–Cenomanian (Lesquereux, 1874; Knowlton, 1919; Doria et al., 2008). Later finds are rare. The youngest Cretaceous species is M. olmosensis from the Campanian–Maastrichtian deposits of Mexico (Guzmán-Vázquez et al., 2018).
CONCLUSIONS
In Northern Asia, Menispermites was distributed from the late Albian up to the Maastrichtian–Danian. During this time, the territory of the North-East of Russia was the center of species diversity of this genus. Short-lived species with limited ranges predominated. Members of Menispermites had diverse leaf morphology. M. sibiricus had the widest geographical and stratigraphic ranges from Kazakhstan to Chukotka and from Cenomanian to Coniacian. The greatest species and morphological diversity of Menispermites occurs in the Cenomanian–Turonian of the North-East of Russia.
ACKNOWLEDGEMENTS
The authors are grateful to Dr. Mike Pole, Dr. Tatiana Kodrul, and anonymous reviewers for their helpful comments. This study was supported by the Russian Science Foundation (grant number 23–77-01087).
REFERENCES
Angiosperm Phylogenetic Group (APG). 2016. An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG IV. Botanical Journal of the Linnean Society, 141:399–436.
https://doi.org/10.1046/j.1095-8339.2003.t01-1-00158.x
Bell, W.A. 1956. Lower Cretaceous floras of western Canada. Geological Survey of Canada Memoir, 285:1–153.
https://doi.org/10.4095/101511
Bell, W.A. 1962. Upper Cretaceous floras of the Dunvegan, Bad Heart, and Milk River Formations of Western Canada. Geological Survey of Canada, 94:1–76.
https://doi.org/10.4095/100613
Berry, E.W. 1916. The lower Eocene floras of southeastern North America. U.S. Geological Survey Professional Paper 91, Washington, D.C., USA.
https://doi.org/10.3133/pp91
Berchtold, B.V. von and Presl, J.S. 1820. O Prirozenosti Rostlin. Krala Wiljma Endersa, Prague.
Bonde, S. 1997. Fossil dicotyledonous liana Anamirta pfeifferi sp. nov. (Menispermaceae) from the Deccan Intertrappean beds of India. Journal of Palaeosciences, 46(3):89–94.
https://doi.org/10.54991/jop.1997.1352
Brongniart, A.1843. Énumération des genres de plantes cultives au Muséum d’Histoire Naturelle de Paris, Saivant l’Orde établi dan l’école de Botanique en 1843. Paris.
Budantsev, L.Yu. 1968. Late Cretaceous flora of the Vilyui Depression. Botanicheskii Zhurnal, 53(1):3–16. [In Russian]
Chandler, M.E.J. 1964. The Lower Tertiary Floras of Southern England. Volume IV. A Summary and Survey of Findings in Light of Recent Botanical Observations. British Museum (Natural History), London.
Collinson, M.E. 1983a. Fossil Plants of the London Clay. Palaeontological Association Field Guides to Fossils no.1, London.
Cronquist, A. 1981. An integrated system of classification of flowering plants. Columbia University Press, New York, USA.
Del Rio, C., Huang, J., Liu, P., Deng, W.Y.D., Spicer, T.E.V., Wu, F.X., Zhou, Z.K., and Su, T. 2021. New Eocene fossil fruits and leaves of Menispermaceae from the central Tibetan Plateau and their biogeographic implications. Journal of Systematics and Evolution, 59(6):1287–1306.
https://doi.org/10.1111/jse.12701
Doyle, J.A. and Hickey, L.J. 1976. Pollen and leaves from the Mid-Cretaceous Potomac Group and their bearing on early angiosperm evolution, p. 139–206. In Beck, C.B. (ed.), Origin and early evolution of Angiosperms. Columbia University Press, New York, USA.
Ellis, B., Daly, D., Hickey, L.J., Johnson, K., Mitchell, J., Wilf, P., Wing, S. (eds.), 2009. Manual of leaf architecture. Cornell University Press, New York, 201 pp.
Friis, E.M., Crane, P.R., and Pedersen, K.R. 2011. The Early Flowers and Angiosperm Evolution. Cambridge University Press, Cambridge.
https://doi.org/10.1017/CBO9780511980206
Givnish, T.J. and. Vermeij, G.J. 1976. Sizes and shapes of liane leaves. The American Naturalist, 110(975):743–778.
https://doi.org/10.1086/283101
Golovneva, L.B. 2006. The genus Menispermites in the Cretaceous floras of Northern Asia. Botanicheskii Zhurnal, 91(11):1731–1740. (In Russian with English abstract)
Golovneva, L.B. 2019. The Chingandzha flora of the Okhotsk-Chukotka volcanic belt. Palaeobotany, 10:13–179.
https://doi.org/10.31111/palaeobotany/2019.10.13
Golovneva, L.B., Shczepetov, S.V., and Alekseev, P.I. 2011. The Chingandzha flora (the Late Cretaceous, North-East of Russia): systematic composition, palaeoecological features and stratigraphic significance. Lectures in memory of A. N. Kryshtofovich, 7:37–61. [In Russian with English abstract]
Golovneva, L.B. and Nosova, N.B. 2012. Albian–Cenomanian flora of Western Siberia.Marathon, Saint–Petersburg. [In Russian]
https://doi.org/10.1134/S0031030115030065
Golovneva, L.B. and Grabovskiy, A.A. 2015. Stratigraphic position and localities of the Temlyan flora from the Anadyr river lower course basin. Palaeobotany, 6:48–67. (In Russian with English abstract)
https://doi.org/10.31111/palaeobotany/2015.6.48
Golovneva, L.B., Herman, A.B., and Shczepetov, S.V. 2015. The genus Menispermites Lesquereux (Angiosperms) in the Cretaceous Grebenka flora of Northeastern Russia. Paleontological Journal, 49(4):429–437.
Golovneva, L.B. and Grabovskiy, A.A. 2019. The genus Hausmannia (Dipteridaceae) in the Cretaceous of the North-East of Russia and its paleobiogeographic implications. Cretaceous Research, 93:22–32.
https://doi.org/10.1016/j.cretres.2018.09.001
Guzmán–Vázquez, I., Calvillo–Canadell, L., and Sánchez–Beristain, F. 2018. Leaves of Menispermaceae and Dioscoreaceae from the Olmos Formation (Upper Cretaceous) from the state of Coahuila, Northern Mexico. Review of Palaeobotany and Palynology, 258:73–82.
https://doi.org/10.1016/j.revpalbo.2018.06.014
Heer, O. 1878. Beiträge zur fossilen Flora Sibiriens und des Amurlandes. Flora Fossilis Arctica, 5:1–58.
Herman, A.B. 1988. Cenomanian Flora of Ugol’naya Bay (Northeastern USSR). Izvestia Akademii Nauk SSSR, Seria Geolologia, 11:110–114. [In Russian]
Herman, A.B. and Lebedev, E.L. 1991. Stratigraphy and flora of Cretaceous deposits of the North-West Kamchatka. Nauka, Moscow. [In Russian]
Herman, A.B. and Kvaček, J. 2010. Late Cretaceous Grünbach Flora of Austria. Verlagdes Naturhistorisches Museum in Wien, Wien.
Herman, A.B., Golovneva, L.B., Shczepetov, S.V., and Grabovsky, A.A. 2016. The Late Cretaceous Arman Flora of Magadan Oblast, Northeastern Russia. Stratigraphy and Geological Correlation, 24(7):651–760.
https://doi.org/10.1134/S0869593816070029
Herrera, F., Manchester, S.R., Hoot, S.B., Wefferling, K.M., Carvalho M.R., Jaramillo, C. 2011. hytogeographic implications of fossil endocarps of Menispermaceae from the Paleocene of Colombia. American Journal of Botany, 98:2004–2017.
Hollick, A. 1930. The Upper Cretaceous floras of Alaska. United States Geological Survey professional paper, 159:1–119.
Hoot, S.B., Zautke, H., Harris, D.J., Crane, P.R., and Neves, S.S. 2009. Phylogenetic patterns in Menispermaceae based on multiple chloroplast sequence data. Systematic Botany, 34:44–56.
https://doi.org/10.1600/036364409787602339
Iljinskaja, I.A. 1974. Protoacerophyllum Romanova, p. 150–152. In Takhtajan, A.L. (ed.), Magnoliophyta Fossilia URSS 1. Nauka, Leningrad. [In Russian]
Jacques, F.M.B. 2009. Fossil history of the Menispermaceae (Ranunculales). Annals de Paléontologie, 95:53–69.
https://doi.org/10.1016/j.annpal.2009.03.001
Jacques, F.M.B. and De Franceschi, D. 2005. Endocarps of Menispermaceae from Le Quesnoy outcrop (Sparnacian facies, Lower Eocene, Paris Basin). Review of Palaeobotany and Palynology, 135:61–70.
https://doi.org/10.1016/j.revpalbo.2005.02.005
Jacques, F.M.B., Wang, W., Ortiz, R.D.C., Li, H.‐L., Zhou, Z.‐K., and Chen, Z.‐D. 2011. Integrating fossils in a molecular-based phylogeny and testing them as calibration points for divergence time estimates in Menispermaceae. Journal of Systematics and Evolution, 49:25–49.
https://doi.org/10.1111/j.1759–6831.2010.00105.x
Jud, N.A., Iglesias, A., Wilf, P., Gandolfo, M.A. 2018. Fossil moon seeds from the Paleogene of West Gondwana (Patagonia, Argentina). America Journal of Botany, 105: 27–942.
Kara, E., Bardin, J., De Franceschi, D., and Del Rio, C. 2023. Fossil endocarps of Menispermaceae from the late Paleocene of Paris Basin, France. Journal of Systematics and Evolution, 62(4):809–828.
https://doi.org/10.1111/jse.13033
Kessler, P.J.A. 1993. Menispermaceae, p. 402–418. In Kubitzki, K., Rohwer, J.G., and Bittrich, V. (eds.), The families and genera of vascular plants. Springer, Berlin.
Knobloch, E. and Mai, D.H. 1984. Neue Gattungen nach Früchten und Samen aus dem Cenoman bis Maastricht (Kreide) von Mitteleuropa. Feddes Repertorium, 95:3–41.
https://doi.org/10.1002/fedr.4910950103
Knowlton, F.H. 1919. Catalogue of Mesozoic and Cenozoic plants of North America.Bulletin of the United States Geological Survey, 696:1–815.
Krassilov, V.A. 1979. The Cretaceous flora of Sakhalin. Nauka, Moscow. [In Russian]
Krassilov, V.A. and Golovneva, L.B. 2004. A minute mid–Cretaceous flower from Siberia and implications for the problem of basal angiosperms. Geodiversitas, 26 (1):5–15.
Krishtofovich, A.N. 1958. Cretaceous flora of the river basin Anadyr. Proceedings of the BIN USSR Academy of Sciences, 8(3):7–68. [In Russian]
Kryshtofovich, A. 1918. On the Cretaceous flora of Russian Sakhalin. J. college of sc., Imperial University of Tokyo, 40(8):1–73.
Kryshtofovich, A.N. and Baikovskaya, T.N. 1960. Cretaceous Flora of Sakhalin Island. Akad. Nauk SSSR, Moscow. [In Russian] Lebedev, I.V. 1955a. Cretaceous system, p. 183–186. In Atlas of leading forms of fossil fauna and flora Western Siberia. Gosgeoltekhizdat, Moscow. [In Russian]
Lebedev, I.V. 1955b. Kassky complex, p. 196–205. In Atlas of leading forms of fossil fauna and flora Western Siberia. Gosgeoltekhizdat, Moscow. [In Russian]
Lebedev, I.V. 1962. Upper Cretaceous plants. Biostratigraphy of Mesozoic and Tertiary sediments of Western Siberia. Proceedings SNIIGGIMS, 22:237–282. [In Russian]
Lesquereux, L. 1874. Contributions to the fossil flora of the Western Territories. Part I. Cretaceous flora. United States Geological and Geographical Survey of the Territories, 6:1–136.
https://doi.org/10.5962/bhl.title.7726
Mai, D.H. 1995. Tertiäre Vegetationsgeschichte Europas. Gustav Fischer Verlag, Jena, Stuttgart, New York.
Manchester, S.R. 1994. Fruits and seeds of the Middle Eocene Nut Beds Flora, Clarno Formation, Oregon. Palaeontographica Americana, 58:1–205.
Manchester, S.R.,Golovneva, L.B., Sokoloff, D.D., and Friis, E.M. 2018. Early eudicot reproductive structure: Fruit and flower morphology of Ranunculaecarpus Samyl. from the Early Cretaceous of eastern Siberia. Acta Palaeobotanica, 58(2):121–133.
https://doi.org/10.2478/acpa–2018–0017
Moiseeva, M.G. 2010. New data on the Cenomanian Flora of the Ugol’naya bay (Northeastern Russia). Paleontological Journal, 44(2):226–239.
https://doi.org/10.1134/S0031030110020140
Moiseeva, M.G. 2014. New Angiosperms from the Late Cretaceous Ust’Emuneret flora of Central Chukotka. Paleontological Journal, 48(6):676–687.
https://doi.org/10.1134/S0031030114060112
Moiseeva, M.G. and Sokolova, A.V. 2007. Campanian flora of the Ugolnaya Bay area (North–Eastern Russia): systematics and paleoclimate. Lectures in memory of A. N. Kryshtofovich, 7:62–87. [In Russian]
Ortiz, R.D.C., Kellogg, E.A., and Werff, H.V.D. 2007. Molecular phylogeny of the moonseed family (Menispermaceae): implications for morphological diversification. American Journal of Botany, 94:1425–1438.
https://doi.org/10.3732/ajb.94.8.1425
Philippova, G.G. 1978. The Cretaceous angiosperms from the Anadyr River basin. Paleontological Journal, 1:138–144. [In Russian]
Pilippova, G.G. 2010. Cretaceous Stratigraphy and Floral Assemblages of the Northern Pekulney Range (Chukotka). Northeastern Integrated Scientific Research Institute, Magadan. [In Russian]
Pilippova, G.G. and Abramova, L.N. 1993. The Late Cretaceous flora of the North-East of Russia. Nedra, Moscow. [In Russian]
Shczepetov, S.V., Herman, A.B., and Belaya, B.V. 1992. Middle Cretaceous flora on the right bank of the Anadyr River: stratigraphic setting, taxonomic composition, and atlas of fossil plants. SVKNII DVO RAN, Magadan. [In Russian]
Shilin, P.V. 1986. Late Cretaceous flora of Kazakhstan: systematic composition, history of development, stratigraphic significance. Nauka, Alma-Ata. [In Russian]
Spicer, R.A. and Herman, A.B., 2010. The Late Cretaceous Environment of the Arctic: A quantitative reassessment using plant fossils. Palaeogeography, Palaeoclimatolgy, Palaeoecology, 295:423–442.
https://doi.org/10.1016/j.palaeo.2010.02.025
Vachrameev, V.A. 1952. Stratigraphy and fossil flora of the Jurassic–Cretaceous beds of western Kazakhstan. Regional Stratigraphy of the USSR, 1. Akademia Nauk SSSR, Moscow. [In Russian]
Vachrameev, V.A. 1966. Late Cretaceous floras of the Pacific coast of the USSR, features of their composition and stratigraphy in some positions. Proceedings of the USSR Academy of Sciences, Seria Geology, 3:76–87. [In Russian]
Wang, W., Ortiz, R.D.C., Jacques, F.M.B., Xiang, X.-G., Li, H.-L., Lin, L., Li, R.-Q., Liu, Y., Soltis, P.S., Soltis, D.E., and Chen, Z.-D. 2012. Menispermaceae and the diversification of tropical rainforests near the Cretaceous–Paleogene boundary. New Phytologist, 195:470–478.
https://doi.org/10.1111/j.1469–8137.2012.04158.x
Wefferling, K.M., Hoot, S.B., and Neves, S.S. 2013. Phylogeny and fruit evolution in Menispermaceae. American Journal of Botany, 100:883–905.
https://doi.org/10.3732/ajb.1200556
Zhilin, S.G. 1974. Menispermites Lesq., p. 97–102. In Takhtajan, A.L. (ed.), Magnoliophyta Fossilia URSS 1. Nauka, Leningrad. [In Russian]
Zolina, A.A., Golovneva, L.B., and Spicer, R.A., 2020. Latest Cretaceous (Maastrichtian) climate of the Koryak Upland of North-East Russia based on a quantitative analysis of a palaeo-polar flora. Palaeogeography, Palaeoclimatology, Palaeoecology, 560:1e12.
https://doi.org/10.1016/j.palaeo.2020.109997
Zvizda, T.V., Papir, A.A., and Chubarova, Z.F. 1974. Report on hydrological and engineering–geological survey of 1:200,000 scale on the territory of sheets Q–60–XXIUSH, XXX1U for 1971–72. Anadyr. [In Russian]
