

 A new non-pterodactyloid monofenestratan pterosaur from the Mörnsheim Formation of southern Germany
A new non-pterodactyloid monofenestratan pterosaur from the Mörnsheim Formation of southern Germany
Article number: 28.3.a42
https://doi.org/10.26879/1542
Copyright Paläontologische Gesellschaft, September 2025
Author biographies
Plain-language and multi-lingual abstracts
PDF version
Submission: 25 February 2025. Acceptance: 9 September 2025.
ABSTRACT
In addition to the numerous pterosaur specimens and species found in the fossilferous limestone beds of southern Germany, recent discoveries there have shown that the intermediate non-pterodactyloid monofenestratan grade of pterosaurs were also present. Here we describe and identify a new genus and species of pterosaur from this grade. This new animal Makrodactylus oligodontus gen. and sp. nov. is smaller than the species currently described from this grade in Germany, but is clearly at, or near, adult status. Diagnostic traits include a number of unusually long teeth in the jaw, and the presence of a long wing finger compared to the proximal arm. This is now the fourth member of this grade known from the region, and the third from the Mörnsheim Formation which are apparently near absent elsewhere, suggesting that these animals inhabited a different environment to the more commonly found pterosaurs.
David W.E. Hone. School of Biological and Behavioural Sciences, Queen Mary University of London, Mile End Road, London, E1 4NS, UK. d.hone@qmul.ac.uk
René Lauer. Lauer Foundation for Paleontology, Science and Education, Wheaton, Illinois, USA. rene@lauerfoundationpse.org
Bruce Lauer. Lauer Foundation for Paleontology, Science and Education, Wheaton, Illinois, USA. bruce@lauerfoundationpse.org
Frederik Spindler. Dinosaurier Museum Altmühltal, Dinopark 1, 85095 Denkendorf, Germany. mail@frederik-spindler.de
Keywords: Pterosauria; Wukongopteridae; Darwinoptera; Pterodactyliformes; Solnhofen; Mörnsheim; new genus; new species
Final citation: Hone, David W.E., Lauer, René, Lauer, Bruce, and Spindler, Frederik. 2025. A new non-pterodactyloid monofenestratan pterosaur from the Mörnsheim Formation of southern Germany. Palaeontologia Electronica, 28(3):a42.
https://doi.org/10.26879/1542
palaeo-electronica.org/content/2025/5661-a-new-german-monofenestratan-pterosaur
Copyright: September 2025 Paläontologische Gesellschaft
This is an open access article distributed under the terms of Attribution-NonCommercial-ShareAlike 4.0 International (CC BY-NC-SA 4.0), which permits users to copy and redistribute the material in any medium or format, provided it is not used for commercial purposes and the original author and source are credited, with indications if any changes are made.
creativecommons.org/licenses/by-nc-sa/4.0/
https://zoobank.org/CB90AA93-3F07-41F0-8997-5FE130062696
INTRODUCTION
The Solnhofen region of southern Germany has been an important area for Upper Jurassic fossils for centuries and has yielded innumerable fossils from the various geological formations that are quarried there. In particular, it represents a hotspot for the diversity of pterosaurs (Barrett et al., 2008) with numerous specimens of multiple species being known for both the early branching non-pterodactyloids (Wellnhofer, 1975) and more derived pterodactyloids (Wellnhofer, 1970).
The pterosaurs uncovered from these deposits provide a rich history, as they include some of the oldest and most important pterosaur specimens ever found (e.g., Collini, 1784; Frey et al., 2003; Ősi and Prondvai, 2009), and key papers relying on large datasets are possible because of the sample sizes (e.g., Bennett, 1996; Hone et al., 2021) and the quantity and quality of preservation. New specimens and new species continue to be found (e.g., Balaenognathus, Martill et al., 2023; Petrodactyle, Hone et al., 2023) and this remains one of the most important areas for our understanding of pterosaur biology and evolution.
Until recently though, this area was surprisingly devoid of non-pterodactyloid monofenestratans (hereafter ‘early monofenestratans’). In 2010, Lü et al. named the new taxon Darwinopterus that showed intermediate features between the earlier non-pterodactyloids and the derived pterodactyloids. This animal has a large head, bearing a confluent nasoantorbital fenestra and a long neck that was formerly seen only in the pterodactyloids, but it retained a short wing metacarpal, a long fifth toe and long tail of the non-pterodactyloids. Since then, other earlier named taxa have been referred to this grade (e.g., Wukongopterus lii, Wang et al., 2009), a large number of taxa named to this group, and new taxa are still being found (e.g., Kunpengopterus antipollicatus, Zhou et al., 2021; Ceoptera evensae, Martin-Silverstone et al., 2024).
The early monofenestratans recovered in varying analyses as a small clade that radiated prior to the origins of the pterodactyloids (e.g., Martin-Silverstone et al., 2024) but more often as a grade (Wang et al., 2017; Zhou et al., 2021). In particular, Hone et al. (2024) recovered them as a grade that included Dearc (previously regarded as a rhamphorhynchine - Jagielska et al., 2022) and others as early branching members, making this grade more inclusive than some other analyses (e.g., Darwinoptera of Andres et al., 2014), and with the closest taxa to the pterodactyloids including Skiphosoura (Hone et al., 2024) and Propterodactylus (Spindler, 2024) from the Solnhofen region.
 Despite these discoveries, early monofenestratans remain a predominantly Middle Jurassic group (Sullivan et al., 2014), but species have been found in the Late Jurassic (Martill and Etches, 2012), including now in the Solnhofen region beds (Rauhut, 2012; Tischlinger and Frey, 2013; Hone et al., 2024) revealing a hidden diversity from this region.
Despite these discoveries, early monofenestratans remain a predominantly Middle Jurassic group (Sullivan et al., 2014), but species have been found in the Late Jurassic (Martill and Etches, 2012), including now in the Solnhofen region beds (Rauhut, 2012; Tischlinger and Frey, 2013; Hone et al., 2024) revealing a hidden diversity from this region.
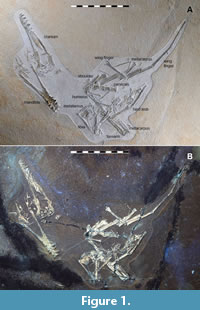
Here we add to this recently uncovered diversity with a specimen of a small pterosaur that represents a new genus of early monofenestratan (Figure 1). Among other features, this animal has unusual proportions to its wing metacarpal and wing finger which define it as a unique taxon.
Institutional Abbreviations
BSPG, Bavarian State Collection for Palaeontology and Geology, Munich, Germany;
LF, Lauer Foundation for Paleontology, Science and Education, Wheaton, Illinois, USA;
NHM-UK, Natural History Museum, London, UK.
METHODS
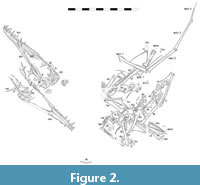 Digital photography of the specimen was obtained in both “daylight” with the use of color balanced LED lights and also under ultraviolet (UV) wavelength lights. UV lights were directed onto the specimen and were sufficient to energize fluorescent mineralizations, (if present in the specimen) to create a UV induced fluorescence (UVIF) response which could be seen in the visible light spectrum and photographically documented. This method provided the ability to discern fine elements such as bones, skin, teeth, and soft tissue preservation which may be poorly discerned or not discernable at all in the visible light spectrum alone. (no soft tissue preservation was present in this specimen), (Figure 2). This method also helped to identify breaks in the matrix and manmade artifacts, such as those which often occur during fossil preparation.
Digital photography of the specimen was obtained in both “daylight” with the use of color balanced LED lights and also under ultraviolet (UV) wavelength lights. UV lights were directed onto the specimen and were sufficient to energize fluorescent mineralizations, (if present in the specimen) to create a UV induced fluorescence (UVIF) response which could be seen in the visible light spectrum and photographically documented. This method provided the ability to discern fine elements such as bones, skin, teeth, and soft tissue preservation which may be poorly discerned or not discernable at all in the visible light spectrum alone. (no soft tissue preservation was present in this specimen), (Figure 2). This method also helped to identify breaks in the matrix and manmade artifacts, such as those which often occur during fossil preparation.
The photographic images were captured with the following equipment: Nikon D4 DSLR and Nikon Z9 mirrorless cameras; FTZ II adaptor as necessary; and lenses: AF Nikkor 28 mm f2.8, AF-S micro Nikkor 60 mm 2.8G ED, and Nikkor 105 mm lens. Filters included: a circular linear polarizing filter (used for daylight and UVIF photography), MidOpt PRO 32-62 (+ polarizing film on the lamp for visible light images for cross polarization to reduce unwanted glare on shiny surfaces and improve clarity and definition); and a color correction filter (for UVIF photography), MidOpt LAT20-62 (orange). Lamps: Daylight images captured with Raleno LEDlights (color temperature of 5500K) with diffuser and polarizing film (used in combination with the circular linear polarizing lens filter to reduce unwanted glare and provide definition): UVIF images utilized “Way Too Cool Quad High-Performance UV Lamp”, equipped with 95 watt bulbs: wavelengths UV A (long wavelength 320 - 400 nm), UV B (mid wavelength 280 - 320 nm), and UV C (short wavelength - 280 -100 nm), with built in Hoya U-325 and Komodo UV-400 filters. NOTE: UV A, B and C wavelengths were utilized together at once. Additional inspection of the specimen was done with a Convoy S2+ 365 nm UV with LG LED flashlight.
NOTE: Safety procedures were followed when UV lights were in use to protect eyes and skin with UV certified safety glasses, and skin was covered.
 All images were captured in a darkened room with the lamps (daylight and UV) as the sole light source. Focus stacked images were created for some figures with use of Helicon Focus software.
All images were captured in a darkened room with the lamps (daylight and UV) as the sole light source. Focus stacked images were created for some figures with use of Helicon Focus software.
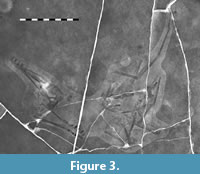
Radiograph images were captured with an X-ray machine by Dr. Christopher Vittore at a hospital in Rockford, Illinois (Figure 3). The X-rays also confirm the original breaks in the slab which were restored during the preparation work, and coupled with the UV images confirm the distal end of the fourth wing phalanx was not recovered, it had been restored.
SYSTEMATIC PALAEONTOLOGY
Pterosauria Kaup, 1834
Breviquartossa Unwin, 2003
Monofenestrata Lü et al., 2010
Makrodactylus gen. nov.
oligodontus sp. nov.
zoobank.org/CB90AA93-3F07-41F0-8997-5FE130062696
Diagnosis
A moderately sized (c. 0.6 m wingspan), non-pterodactyloid monofenestratan pterosaur with the following unique traits for members of this grade: small and knob-like expansion to the descending process of the nasal; low tooth count with only seven teeth each side of the upper jaw and six in the lower; short and broad jaw with long symphysis that is close to half the length of the jaw; a very short wing metacarpal compared to humerus (ratio of 1.75, other early monofenestratans are 1.55 or less); wing finger phalanges that are subequal in length; a long wing finger compared to the proximal wing (humerus + ulna + wing metacarpal vs wing finger ratio of 0.62).
In addition, it can be further separated from other recently named and described specimens from the region by the following traits: It has a much longer humerus compared to the femur and lacks the proportionally long tibia of the Propterodactylus (Tischlinger and Frey, 2013; Spindler, 2024); it lacks the long tooth row at the back of the skull and short humerus to femur ratio of Skiphosoura (Hone et al., 2024); it lacks the large deltopectoral crest on the humerus and the long wing phalanx 2 of the ‘Rhamphodactylus’ (Rauhut, 2012); it lacks the long wing phalanx 1 and short tibia of the Propterodactylus (Tichlinger and Frey, 2013; Spindler, 2024).
Holotype
Specimen LF 1370 (Figure 1) is a partial skeleton consisting of a mostly complete cranium and mandible, some cervical and dorsal vertebrae, partial pectoral and pelvis girdles and most of all four limbs. The specimen is largely compressed into two dimensions.
Etymology
Makrodactylus from the Ancient Greek, from ‘makros’ meaning ‘long’ and ‘dactylus’ meaning finger, for the proportionally long wing finger of this animal. The species name oligodontus is from the same language with ‘oligo’ meaning ‘few’ and ‘dont’ for teeth, for the low tooth count that is unique to the species.
Locality Information
The specimen was found on the 13th of April, 2014 in the private section of the Schaudiberg Quarry near Mühlheim, Bavaria, Germany. The Schaudiberg Quarry is located near the town of Mühlheim which is part of the community of Mörnsheim/Gailachtal at the western end of the district of Eichstätt. This is in the region of Franconia between Nuremberg and Munich, in the state of Bavaria, Germany.
The specimen was found in the Mörnsheim Formation, and was collected at the following coordinates: 48° 51’14” North, 10° 59’20” East and with an approximate elevation of 500 m. The Mörnsheim Formation is a series of laminated, lithographic limestones that are Lower Tithonian in age (Upper Jurassic) and is considered part of the Malm Zeta 2b-3 zone (Werner, 2005). From our discussion with the quarry owners we are informed that this specimen likely comes from the same specific layer that yielded the ‘Rhamphodactylus’ specimen (Rauhut, 2012), but this has not been confirmed.
The specimen was discovered by Roland Pöschl and was prepared by Rolf Kriegerbarthold from April to July of 2014. The specimen was prepared under microscopy using pneumatic tools, Hardy Winkler Airscribes and Teflon Airscribe as well as pin and needle tools. Binding agents of Starbond Super Thin Fast and Akemi 1000 were used to consolidate bones and matrix as appropriate.
The specimen was acquired by the Lauer Foundation for Paleontology, Science and Education, Wheaton, Illinois, U.S.A. in September 2014, and is permanently housed in the Lauer Foundation collection as LF 1370. Permanent access to type and figured specimens, as well as specimens listed or cited in publications together with other scientifically important specimens is guaranteed.
Taphonomy
As with other pterosaurs from this locality (see Rauhut, 2012; Hone et al., 2023, 2024) the specimen is somewhat disarticulated, but many of the elements lie close to their original positions. For example, although the cranium has separated into various elements, the parts are largely in situ, and the elements of the wings and legs are close to their original articulations. Small elements like ribs, the free metacarpals, and prepubes lie close to the larger elements with which they would usually articulate, despite some separation. Therefore, as with previous studies we infer from this pattern that the specimen was at rest on the substrate for a considerable period of time before burial in order to have decayed and separated as much as it did, but that it did not suffer much or any scavenging or show evidence of any currents or water movement which would separate and remove the small elements or move them all into a a similar orientation.
Description
The specimen is on a slab of pale limestone that is approximately 0.45 by 0.55 m. The slab was broken when it was recovered (a series of major breaks are visible in the X-rays and UV images) though with limited damage to the bones (Figure 3). The distal half of the fourth wing phalanx was damaged or lost and it has been restored.
The specimen is moderately well-preserved and mostly complete (Figure 1 and Figure 2). Some of the surface texture is rough and grainy and the details of some small elements are not well preserved, especially where elements overlap. The specimen is also very flat and is more similar to typical crushed Solnhofen region pterosaurs, and less like the more 3D preserved ones normally seen in this quarry (see Hone et al., 2023, 2024). Nearly all of the skull, cervical series, and limbs are present as well as most of the pectoral and pelvic girdles and some dorsal vertebrae, ribs and other small elements. There is no evidence of any soft tissues preserved, either observed under natural light or with various bands of UV wavelength light (Figure 2). The total wingspan is 606 mm, calculated as twice the total length of the humerus, radius, wing metacarpal and wing finger elements.
The orientation of the specimen is inconsistent, with various elements rotated and the limbs folded over and alongside one another (e.g., the mandible is rotated, facing the opposite direction to the cranium, and both are separated from the rest of the specimen (Figure 1). As with other pterosaur specimens from the quarry (e.g., see Hone et al., 2023, 2024), LF 1370 shows separation of many elements, but they are not entirely disarticulated, and many small parts are either in articulation or close to their natural positions (e.g., cranial elements, metacarpals, the hindlimbs).
The ontogenetic status of the animal is that of an adult. It lacks the fine-grained bone texture of very young specimens and shows a pattern of fusion that is instead consistent with an animal that is close to or at skeletal maturity (e.g., Bennett, 1996). Although the skull is not well fused and many elements have separated, this may be normal for even adult early monofenestratans as this is also seen in Skiphosoura (Hone et al., 2024) and a similar pattern is also found in ctenochasmatid pterosaurs (Prondvai and Ősi, 2011). Therefore LF 1370 is at, or close to, full osteological maturity showing multiple features from Kellner’s (2015) ontogenetic categories 5 and 6, which represent the most mature individuals. The mandibular symphysis is well sutured, although there is a faint line in the midline which could be incomplete obliteration of the suture. The scapulae and coracoids are fully fused into their respective pairs. No clear epiphyses on any long elements (e.g., humerus, femur, tibia). The carpals are fully fused into a proximal and distal syncarpal pair. The pelvis is poorly preserved, but there is no evidence of sutures of the major elements. The extensor tendon processes on the first wing phalanges are fully fused. This is therefore an adult specimen, with perhaps only limited further growth.
 Skull. The cranium is preserved in right lateral view (Figure 4). It is partially disarticulated and crushed such that the posterior part of the skull is in ventral (palatal) view, and some elements have separated and are present around the posterior part of the cranium but also around the posterior part of the mandible. Many of these elements are poorly preserved and even under UV light their details are not clear. Some of the following identifications of elements are therefore tentative and some pieces cannot be identified. The skull is roughly triangular in outline being 109 mm long and around 28-30 mm tall at the posterior end (this value is approximate given the crushing and displacement of posterior cranial elements). This shape is notably short and tall, similar to that of the Propterodactylus (Tischlinger and Frey, 2013; Spindler, 2024) and Darwinopterus modularis (Lü et al., 2010), but not Darwinopterus linglongtaensis (Wang et al., 2010), Darwinopterus camposi (Cheng et al., 2025) or Cuspicephalus (Martill and Etches, 2011) or as inferred for Melkamter (Fernanes et al., 2024). The nasoantorbital fenestra is around 38 mm long (including what appears to be a partial fossa anteriorly). The orbit is subcircular in shape (as is common for early monofenestratans) and is about 20 mm wide by 17 mm high. The other cranial fenestrae cannot be made out.
Skull. The cranium is preserved in right lateral view (Figure 4). It is partially disarticulated and crushed such that the posterior part of the skull is in ventral (palatal) view, and some elements have separated and are present around the posterior part of the cranium but also around the posterior part of the mandible. Many of these elements are poorly preserved and even under UV light their details are not clear. Some of the following identifications of elements are therefore tentative and some pieces cannot be identified. The skull is roughly triangular in outline being 109 mm long and around 28-30 mm tall at the posterior end (this value is approximate given the crushing and displacement of posterior cranial elements). This shape is notably short and tall, similar to that of the Propterodactylus (Tischlinger and Frey, 2013; Spindler, 2024) and Darwinopterus modularis (Lü et al., 2010), but not Darwinopterus linglongtaensis (Wang et al., 2010), Darwinopterus camposi (Cheng et al., 2025) or Cuspicephalus (Martill and Etches, 2011) or as inferred for Melkamter (Fernanes et al., 2024). The nasoantorbital fenestra is around 38 mm long (including what appears to be a partial fossa anteriorly). The orbit is subcircular in shape (as is common for early monofenestratans) and is about 20 mm wide by 17 mm high. The other cranial fenestrae cannot be made out.
The premaxilla-maxilla complex is relatively well-preserved and makes up the majority of the anterior two thirds of the skull. This is seen in ventrolateral view for the right side, and the ventral and medial side of the left, as the posterior part of the right maxilla is missing. The rostrum (anterior to the NAOF) is relatively low and straight with perhaps a slight dorsal expansion as a small ridge above the third and fourth teeth, similar to the condition in Kunpengopterus antipollicatus (Zhou et al., 2021 - their figure 1). There are seven teeth across premaxilla-maxilla, which is much less than any of the early monofenestratans, even those with relatively few teeth such as Skiphosoura (12, Hone et al., 2024) and less even that the Propterodactylus (11, Tischlinger and Frey, 2013; Spindler, 2024). The split of teeth between these elements cannot be determined because the suture between the elements is not seen. The tooth row is unusually short, finishing anterior to the nasoantorbital fenestra (NAOF) which is similar to the pattern seen in e.g., Kunpengopterus sinensis (Wang et al., 2010), and Darwinopterus robustodens (Lü et al., 2011), but very different from that of Melkamter which has teeth all the way to the back of the NAOF (Fernanes et al., 2024).
Part of a large and broad palatine is visible in the NAOF, but it is broken posteriorly and no details of the other palatal elements or openings in the palatal complex are visible. A similar plate-like piece is seen in the anterior part of the NAOF of Propterodactylus (Tischlinger and Frey, 2013, their figures 4 and 5). Two more palatine pieces, or perhaps other parts of the palate, lie behind the mandible. These are long and very thin plates or splinters of bone and would have formed the posterior parts of the palatines or the ectopterygoids.
The skull roof is largely present as the paired frontals and parietals that are visible in ventral view (Figure 4). The frontals as a pair are rather oval in shape with slightly thickened margins and midline, and the concavities on their posterior parts show the dorsal margin of the orbits. Behind the frontals are the paired parietals and probably part of the dorsal braincase. This part is not well preserved and is difficult to make out any details beyond a midline concavity that is surrounded by a raised ridge. Its overall shape is similar to that of the ‘Rhamphodactylus’ (Rauhut, 2011, his figure 2), though this complex is seen in dorsal view.
On the dorsal face of the left ventral ramus of the maxilla is the left jugal. This can be seen to be a thin and triradiate element, though the anterior and posterior rami are mostly covered by the maxilla and are exposed only as very thin rods. The ascending process is thin and directed anteriorly and is 7 mm long. This anterior deflection is also seen in the Propterodactylus (Tischlinger and Frey, 2013, their figure 4) and Skiphosoura (Hone et al., 2024), but it is closer to a vertical direction in e.g., Darwinopterus linglongtaensis (Wang et al., 2010, their figure 2) and D. robustodens (Lü et al., 2011, their figure 2). Behind the jugal, lying parallel to the ventral ramus of the maxilla, lie a series of rod-like elements. These are the right (lateral view) and left (medial view) quadrates, and dorsal to them, the more triangular right postorbital seen in lateral view. The left postorbital is also preserved, but lying between the rami of the mandible and no details are visible. There are some indistinct bony patches between these and the parietals, but these cannot be positively identified, though one curved element near the postorbital may be a squamosal.
Sitting adjacent to the frontals, but clearly separated from the skull, is a lacrimal. This is a small triangular element with a short projection at each point, and with a small foramen in the center. Both nasals are preserved. One of them sits just next to the lacrimal, with the other sitting some distance away next to the right ramus of the mandible. The nasals are small and triradiate elements and both show a knob-like expansion at the tip of the descending process, which is unique in pterosaurs.
The mandible in presented dorsal view (Figure 4). The mandibular symphysis is very long (39 mm), and is nearly half the length of the mandible and has a sharp V-shaped incision in the midline, such that the lateral parts of this extend even further back onto the dentaries. The jaw is relatively short (three times as long as wide at the posterior end, where it is about four times as long in Skiphosoura, Hone et al., 2024 - the only other early monofenestratan where the jaw is similarly preserved for comparison) which would give the skull short and wide proportions. Each dentary bears six teeth, with teeth 2-5 being larger than the first and last teeth. There may be a seventh tooth present, as there is a piece of broken bone at the back of the jaw that is about the right size and shape to be a tooth, but it is not well-preserved and shows no trace of enamel, so its exact identity cannot be determined. Even if this was a seventh tooth, this would still be a very low tooth count as a whole. The tooth row is short, finishing approximately in line with the posterior end of the symphysis. The sutures at the posterior part of the mandible are not very clear, although the surangular is clearly visible. This is a long, thin and triangular element, with a medial posterior expansion. One plate-like and long piece of bone at the posterior end of the right ramus is considered to be a separated postdentary element, possibly the right surangular.
The teeth are generally in situ in the jaws, although one isolated tooth is present in the middle of the slab. The teeth are mostly long, thin and curved, and the longest teeth (3rd cranial and 4th dentary) are about 6-7 mm long and 2 mm wide. The low number of teeth, that are also well-spaced and thin and curved are markedly similar to Propterodactylus (Spindler, 2024), and similar to those of Skiphosoura (Hone et al., 2024), and are generally unlike the more numerous and conical teeth of early monofenestratans (Wang et al., 2010). The posteriormost teeth in both upper and lower jaws here are smaller, being as little as 4 mm long but a similar width. The spacing of the teeth is generally even being around 2-4 mm between each tooth in both upper and lower jaws, though this is harder to measure in the lower jaw where some teeth are clearly displaced. The teeth in the upper jaw are vertically orientated, but those in the lower jaw are slightly splayed laterally, though again with some crushing and displacement which may have affected this.
Vertebral column. Apart from the sacrum, much of the vertebral series is preserved in LF 1370, although it is mostly disarticulated, and the various elements are seen in different parts of the specimen and in variety of orientations. Some individual elements are well-preserved, but the details of most of them are difficult to ascertain. The tail is largely concealed behind other elements.
A probable atlas-axis complex sits behind and slightly below the skull. It is an irregularly shaped, but rounded piece of bone, but few details are visible. There is a distinct semi-circular notch on one side with an apparent ridge around the edge, and a small hemispherical bulb protrudes close to this notch.
 One cervical vertebra lies close to one of the legs (Figure 2) and is in dorsal view, with possibly a second one next to it in partial articulation, but this is very poorly preserved. The centra is 10 mm long and the zygapophyses extend around 3 mm anteriorly and posteriorly. The appearance of the neural arch is short and broad in dorsal view. An articulated series of at least four cervical vertebrae lie together, the anteriormost of these is in dorsolateral view and the three posterior to it are in lateral view (Figure 5). The series is partially covered by a scapulocoracoid and it appears that one relatively short element may be largely or entirely covered. In total six are preserved here, although only five of them (presumably 3, 5-8) are visible. The longest of the series has a centrum that is 11 mm long, 5 mm tall and has a slightly convex ventral margin in lateral view. Their orientation and overlap make it difficult to compare these to other taxa, but these appear to be proportionally shorter and broader than those of Skiphosoura (Hone et al., 2024) or K. sinensis (Cheng et al., 2017), and are similar to e.g., D. robustodens (Lü et al., 2011) and Propterodactylus (Spindler, 2024).
One cervical vertebra lies close to one of the legs (Figure 2) and is in dorsal view, with possibly a second one next to it in partial articulation, but this is very poorly preserved. The centra is 10 mm long and the zygapophyses extend around 3 mm anteriorly and posteriorly. The appearance of the neural arch is short and broad in dorsal view. An articulated series of at least four cervical vertebrae lie together, the anteriormost of these is in dorsolateral view and the three posterior to it are in lateral view (Figure 5). The series is partially covered by a scapulocoracoid and it appears that one relatively short element may be largely or entirely covered. In total six are preserved here, although only five of them (presumably 3, 5-8) are visible. The longest of the series has a centrum that is 11 mm long, 5 mm tall and has a slightly convex ventral margin in lateral view. Their orientation and overlap make it difficult to compare these to other taxa, but these appear to be proportionally shorter and broader than those of Skiphosoura (Hone et al., 2024) or K. sinensis (Cheng et al., 2017), and are similar to e.g., D. robustodens (Lü et al., 2011) and Propterodactylus (Spindler, 2024).
The neural spines are short and rectangular and measures 9 mm by 3 mm high in the largest cervical. Overall, these are relatively short and deep and broad elements with well-rounded pre- and postzygapophyses. None extend as far from the centrum as seen in the ‘Rhamphodactylus’ (BSPG 2011 I 133) and are more similar to the condition seen in Propterodactylus (Spindler, 2024) and other earlier branching monofenestratans. There is no evidence of cervical ribs on these elements. Collectively, they suggest that the neck was relatively short and robust and the total lengths of all the elements mentioned above would be only around 70 mm, which is considerably shorter than the length of the skull. There are no postexapophyses as seen in Skiphosoura (Hone et al., 2024) though this may be a result of the much smaller size of the specimen here.
 There are three poorly preserved dorsal vertebrae present that are visible in anterior or posterior view (two lie alongside a femur, and the other is close to the cervical series). These are hard to make out in natural light, but they are relatively clear under UV light (Figure 6) and are apparently only preserved as neural arches with the centra covered or lost. They have neural spines and lateral processes that are subequal in length to each other (c. 3 mm). The lateral processes project at an angle of approximately 30 degrees and the neural canal is slightly triangular in shape (with the apex under the neural spine) and is c. 2 mm in diameter. The shape of the neural arch is fundamentally similar to that seen in Ceoptera (Martin-Silverstone et al., 2024), K. sinensis (Cheng et al., 2017), and Skiphosoura (Hone et al., 2024). A series of poorly preserved elements lying close to these, but mostly hidden below the left femur (Figure 5), are likely a series of dorsal neural arches seen in lateral view. They are small and triangular and with possible lateral processes projecting from their bases.
There are three poorly preserved dorsal vertebrae present that are visible in anterior or posterior view (two lie alongside a femur, and the other is close to the cervical series). These are hard to make out in natural light, but they are relatively clear under UV light (Figure 6) and are apparently only preserved as neural arches with the centra covered or lost. They have neural spines and lateral processes that are subequal in length to each other (c. 3 mm). The lateral processes project at an angle of approximately 30 degrees and the neural canal is slightly triangular in shape (with the apex under the neural spine) and is c. 2 mm in diameter. The shape of the neural arch is fundamentally similar to that seen in Ceoptera (Martin-Silverstone et al., 2024), K. sinensis (Cheng et al., 2017), and Skiphosoura (Hone et al., 2024). A series of poorly preserved elements lying close to these, but mostly hidden below the left femur (Figure 5), are likely a series of dorsal neural arches seen in lateral view. They are small and triangular and with possible lateral processes projecting from their bases.
A number of ribs and gastralia are present in several places on the slab (Figure 5, Figure 6). The ribs are generally long, thin and straight elements that are about 30 mm long. A cluster of shorter (<15 mm) and thinner elements lie together next to the right femur and these are most likely a set of gastralia. Some small elements lying close to these could be sternal ribs, but they are indistinct.
No sacral vertebrae can be seen. However, given the fusion of the skeleton as a whole, it is likely that it was fused to the pelvic elements and so it is hidden behind the pelvic plates (Figure 6B). There are some indistinct subcircular elements close to the prepubes which may be proximal caudal centra based on their size, and similarly, it is possible that one or two of the small indeterminate elements on the slab represent anterior caudal vertebrae. However, there is a series of very small caudal elements preserved by the right tibia. The shape of these is difficult to make out but they are tiny and simple elements that are 4 mm long and only 1 mm tall and apparently lack elongate zygapophyses. The total length of the series is only 15 mm, but both ends of this are covered so it would have been longer. Given their size they may represent distal caudals, but even so there is no evidence of the elongate zygapophyses and chevrons that are present to near the tip of the tail in Skiphosoura (Hone et al., 2024), Douzhanopterus (Wang et al., 2017) and other earlier branching monofenestratans. and suggesting that the tail here is simple and structured more like that of the Propterodactylus (Spindler, 2024) or perhaps with still greater reduction in the bounding filaments since these are short in that taxon, but these are apparently absent here (or are so small as to have not preserved well).
Girdles. Both of the scapulacoracoids are present and are seen in anterior view to give them a V-shape, with the two rami meeting at an angle of approximately 160 degrees (Figure 5). The right sits on top of the cervical series, and the left is partially covered by gastralia, but both are fairly clearly visible, although the preservation of the glenoid is indistinct in both cases. As is common in early pterosaurs, the scapula is longer and much thicker than the coracoid (34 mm long with a midshaft of 4 mm, compared to 24 mm and 2 mm). This is also seen in e.g., Skiphosoura (Hone et al., 2024) and Ceoptera (Martin-Silversone et al., 2024), though in the latter, the difference in length between the two is less extreme. Here the two elements are also more different in their thickness than in D. linglontaensis (Wang et al., 2010) at least, though in D. camposi the shaft of the scapula appears to be potentially thinner than that of the coracoid (Cheng et al., 2025, their figure 1).
A small sternal plate (15 mm by 15 mm) is present, although the details on this are very unclear except under UV light and even here the lateral and distal margins are broken and not well-preserved (Figure 6). The sternal plate has straight lateral margins with a somewhat semi-circular expansion and posterior margin, which is broadly similar in shape to that seen in other early monofenestratans (Hone, 2023). Anteriorly, there is a slight ridge close to the midline, which may be taphonomic, but we consider to be part of the base of the cristospine. The cristospine is not visible and may be separate and lost, or intact and hidden below the tibia.
Adjacent to the sternum, one of the prepubes is present and lies close to the rest of the pelvis and hindlimbs (Figure 6). The prepubis is roughly semi-circular, though with a somewhat flattened anterior margin, and with a short extension in the middle of the posterior straight edge as is common for taxa in this grade (Hone et al., 2024). Alongside this is an ischium, apparently separate from the ilium and pubis given its small size and overall shape. It is only slightly larger than the prepubis and with an overall similar morphology though with a distinct sharp corner and flat posterior margin which in other members of the group usually shows a knob-like expansion (e.g., Propterodactylus, Spindler, 2024; Kunpengopterus sinensis, Cheng et al., 2017).
A large flat piece of bone lying next to these and adjacent to the head of the femur is considered likely to be one of the two ischiopubic plates. This is indistinct and not well preserved, but its size, shape and position would conform to it being a major part of the pelvis.
Limbs. The wings are nearly complete and missing only the pteroid and most of the elements of the free digits (Figure 2). They are, however, somewhat disarticulated and while the elements all lie close to a natural position, many of the joints have separated and adjacent elements are at unnatural angles to one another.
The humerus is a simple and straight element that is relatively broad (42 mm long, with a minimum width of 4 mm) though the two ends are slightly wider than the midpoint. One is preserved with the proximal end more visible and the other with the distal end better exposed. The deltopectoral crest as seen is a simple semicircle that extends less than quarter of the way down the shaft. This is similar to that seen in Propterodactylus (Spindler, 2024) and Skiphosoura (Hone et al., 2024), but otherwise is a smaller and more rounded crest than seen in other early monofenestratans that tend to have a more triangular and distally extended deltopectoral crest. The shaft of the humerus here is robust, with the distal end expanding medially to a width of 8 mm, and it is notably broader for its length than in the ‘Rhamphodactylus’ (Rauhut, 2012) and D. linglongtaensis (Wang et al., 2010) but is otherwise generally similar to other early monofenestratans. The radii and ulnae are preserved in their respective pairs, but all four elements are partly covered by other elements and their ends are not clear. As is usual in pterosaurs, these are long, straight and simple elements with the ulnae being the more robust elements.
The carpals appear to be fully fused into the proximal and distal syncarpals of adult pterosaurs, as no sutures are visible. However, these are not well preserved and the details of these complex elements are difficult to make out. On the right wing, only the proximal syncarpal can be seen and the distal element is either hidden or missing. On the left wing, the proximal element is visible in anterior or posterior view and it is nearly square in outline with sides that are 7 mm long. The pteroids are not preserved.
The free metacarpals of digits 1-3 are present and these are long (23 mm) thin, straight elements that are subequal in length to one another and the wing metacarpal, as is normal for non-pterodactyloid pterosaurs (Figure 5). The free metacarpals have slightly expanded distal ends which are subcircular. These elements are very thin (less than 1 mm), similar to the condition in Propterodactylus (Spindler, 2024), but these are notably much more robust in many other early monofenestratans, either all three of the elements (e.g., Changchengopterus, Zhou and Schoch, 2011 and Wukongopterus Wang et al., 2009) or just one of them (e.g., K. sinensis Cheng et al., 2017, D. robustodens Lü et al., 2011).
The wing metacarpal is proportionally short in Makrodactylus compared to other members of the grade. It is robust with a broad proximal end that then tapers gently along its length before expanding distally to form the ginglymoid articulation with the first wing phalanx. Few pterosaurs from this grade preserve the wing metacarpal in the same orientation seen here, though K. sinensis (Cheng et al., 2017) has a similarly preserved element and this shows a similar shape and tapering.
There are several small phalanges preserved on the slab. Two of these are close to one of the metacarpal sets and are relatively large compared to the metatarsals, suggesting that these are from the hands rather than the feet. They are small and typical of pterosaur free phalanx elements, but it cannot be determined to which fingers they might have originally belonged. No manual unguals are visible.
The four wing phalanges are long, straight elements (Figure 1). The first phalanx is the shortest and shows a small, but fully fused (i.e., obliterated suture) extensor tendon process on its anterior face close to the proximal end. Unusually, the first three elements are all very similar in length (see Table 1 for measurements). The fourth phalanx is also straight and shows no evidence of curvature along its preserved length, as is seen in some pterosaurs (Hone et al., 2015). As noted above, this element has been restored (Figure 3)and the original length and terminal shape of the bone is not known. The length of the incomplete fourth wing phalanx of the wing was estimated by overlaying the same element from Propterodactylus as the two are close relatives and have a similar proximal shape and taper to the phalanges, and this method suggests that the element is c. 44 mm long. This is slightly shorter than it appears on the slab where it is restored as being 48 mm long.
Both hindlimbs are present but the right only had a femur and tibia, where the left also has a partial pes preserved (Figure 1). The femur has a small semicircular head attached to a thin neck which is offset from the femoral shaft to an angle of approximately 50 degrees. The femur is rather robust (midshaft width of 4 mm) and is slightly broader than the tibia, which is only a little longer than the former element (46 vs 38 mm). The femoral shaft is straight with a slight distal expansion to a pair of condyles and is similar to other early monofenestratans, but is notably more robust than Ceoptera (Martin-Silversone et al., 2024) where the femur also notably expands distally. The tibia in Makrodactylus is also straight and with a distal expansion. The tarsals have not fused to the end of the tibia to form a tibiotarsus, but are separate, if indistinct, elements. There is a partial fibula preserved on the left tibia, this is very thin and rod like and may be fused to the tibia. It is broken distally and so its complete extent is not known, but it is at least half the length of the tibia.
There are four metatarsals preserved on the right foot. These are all long, thin and subequal in length and remain tightly packed together and subparallel to each other, indicating that they represent metatarsals I-IV. A number of small phalangeal elements are present close to the distal end of the metatarsals, but as with the free manual elements these cannot be assigned to a given digit.
DISCUSSION
Identity and comparisons
This animal is clearly a monofenestratan given the proportionally large head with the distinctive confluent nasoantorbital fenestra (Lü et al., 2010). However, the proportionally short wing metacarpal and overall short wing phalanx 1 are very clear characters that show that the animal is an early monofenestratan, and not a pterodactyloid (Lü et al., 2010). Within this context, its exact position it difficult to determine - for example, although poorly preserved, the tail appears to be a simple one which suggests that this is a more derived early monofenestratan and close to pterodactyloids (e.g., see Hone et al., 2024), although the moderate elongation of the wing metacarpal compared to non-monofenestratans (just over half the length of the humerus) would suggest an earlier branching position. Other characters however generally support the more derived state of this animal, such as the anterior projection of the jugal being deflected, the reduced free metacarpals, wing phalanx one being the proportionally longest of any non-pterodactyloid, and the robust femur are all traits that would link this to the derived Propterodactylus (Spindler, 2024) but are absent in most of the earlier branching taxa. As noted by Hone et al. (2024), despite a relatively clear transition of key characteristics from the non-monofenestratans to the pterodactyloids, there may be a more mosaic pattern of character states, with some taxa showing a mix of ancestral and derived traits. The mixture of traits seen here would present such a case.
With a total wingspan of just over 0.6 m (including the estimated length of the incomplete 4th wing phalanx - see Table 1 and above), this was a moderately sized adult pterosaur for the Jurassic, and comparable to a number of other small species from the Solnhofen region (Table 2). However, compared to other early monofenestratans, it is rather small. Skiphosoura is very large, with a wingspan nearly three times as large at 1.7 m, and the ‘Rhamphodactylus’ is c. 1 m., Propterodactylus is only a little smaller than Makrodactylus here, with a wingspan of 0.52 m, however as this is considered to be a juvenile animal (Tischlinger and Frey, 2013; Spindler, 2024), then this would likely have been considerably larger than LF 1370 as an adult. Makrodactylus is therefore the smallest early monofenestratan in the region and smaller than nearly all other specimens from this grade, as well as being the smallest known mature specimen.
There is variable maturity in pterosaurs including in Rhamphorhynchus, (e.g., Prondvai et al., 2014), so that animals of similar ages could be of rather different sizes (Chinsamy et al., 2008). Furthermore, adult animals may vary considerably in size (e.g., see Hone and McDavid, 2024), including sexual size dimorphism (see Bennett, 2001). Nevertheless, animals that are considerably different in size, if they are at similar stages of maturity, will likely represent those which are different sizes as an adult.
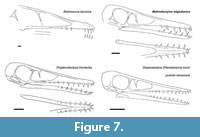 The skull of Makrodactylus is notably proportionally short and tall, unlike the longer and so more slender rostra that is typical of the early monofenestratans (Figure 7). Jiang et al. (2021) noted the rostrum extends in ontogeny in early monofenestratans. The skull here does have a similarly short and deep proportions to that of a juvenile specimen of Kunpengopterus (Jiang et al., 2021) and the sole specimen of Propterodactylus which is also not an adult animal (Spindler, 2024). Here Makrodactylus is more mature so this short and tall head appears to be a genuine feature, and this juvenile-shaped skull in an animal that is otherwise considered an adult may be an example of paedomorphosis. The tooth count and the length of the tooth row does not appear to change between juvenile and adult Kunpengopterus (Jiang et al., 2021), so the short row with few teeth seen here is considered to be a genuine feature of the animal.
The skull of Makrodactylus is notably proportionally short and tall, unlike the longer and so more slender rostra that is typical of the early monofenestratans (Figure 7). Jiang et al. (2021) noted the rostrum extends in ontogeny in early monofenestratans. The skull here does have a similarly short and deep proportions to that of a juvenile specimen of Kunpengopterus (Jiang et al., 2021) and the sole specimen of Propterodactylus which is also not an adult animal (Spindler, 2024). Here Makrodactylus is more mature so this short and tall head appears to be a genuine feature, and this juvenile-shaped skull in an animal that is otherwise considered an adult may be an example of paedomorphosis. The tooth count and the length of the tooth row does not appear to change between juvenile and adult Kunpengopterus (Jiang et al., 2021), so the short row with few teeth seen here is considered to be a genuine feature of the animal.
This new taxon is now the third early monofenestratan from the Mörnsheim Formation alongside Skiphosoura (Hone et al., 2024) and the ‘Rhamphodactlyus’ (Rauhut, 2012). Pterosaurs have been excavated in the region of southern Germany for nearly 250 years, with hundreds of specimens representing around 20 species. But only since 2012 have any from this grade of pterosaurs been found, and three of the four known so far are from the same formation. As noted above, the preservation of this site is unusual compared to other local plattenkalk beds, and suggests that conditions of the local environment were different from the rest of the system or are older beds.
This in turn suggests that these animals had a fundamentally different ecology to their predecessors if they were common only in this locality. Most early pterosaurs are associated with marine or freshwater aquatic environments. The Solnhofen was an archipelago surrounded by lagoons (Barthel et al., 1990) and the pterosaur faunas are dominated by piscivorous rhamphorhynchids for the non-monofenestratan taxa, and filter-feeding ctenochasmatids for the pterodactyloids (Witton, 2013). We suggest therefore, that the early monofenestratans may have been more terrestrial in their habits which would match with their general rarity in the Solnhofen archipelago. This would fit with the patterns seen elsewhere as for example, their frequency in various terrestrial settings in the Chinese Jurassic faunas (Sullivan et al., 2014) where rhamphorhynchids are rare (only one specimen is known - Qinglongopterus, Lü et al., 2012) and the anurognathids (also widely considered to be terrestrial) are common (e.g., see Hone, 2020). The anurognathids are present in the Solnhofen archipelago, but like the early monofenestratans are rare in the traditional Solnhofen archipelago beds, with only two specimens known (Bennett, 2007), despite these first being found in these beds over a century ago.
In this context, it suggests that the fauna preserved in the Mörnsheim beds - although clearly deposited in a marine environment - were fundamentally different to other formations (or at least localities) in Southern Germany. The unique fauna seen here may be because this was a subtly different time and represent a fundamental shift from earlier formations, but may also simply be because there was more land nearby that meant there was a tendency for more terrestrial animals to become buried here.
Wing finger proportions
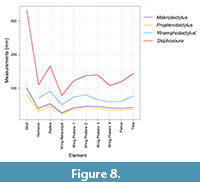 Pterodactyloid pterosaurs have a characteristic pattern of the lengths of the bones in the wing finger where phalanx 1>2>3>4 (with the exception of Forfexopterus, Jiang et al., 2016). Non-pterodactyloids have wing finger phalanges 2 and 3 as the largest, then phalanx 1 and with 4 almost always being the shortest. However, here we see an apparently unique pattern where the first three of these elements are almost subequal being respectively 44, 48, and 46 mm long. (The exact length of the restored phalanx 4 being unknown, but with 27 mm preserved and not yet apparently beginning to taper to a point, it cannot have been too much shorter than these others and we restore this as being c. 44 mm in length). Although it is impossible to rule out intraspecific variation, the high level of conservatism of the components of the pterosaur skeleton including the wing finger, (in some taxa at least - Habib and Hone, 2024) argue that such proportions are likely genuine and represent a distinct anatomical (and therefore taxonomic) feature (Figure 8).
Pterodactyloid pterosaurs have a characteristic pattern of the lengths of the bones in the wing finger where phalanx 1>2>3>4 (with the exception of Forfexopterus, Jiang et al., 2016). Non-pterodactyloids have wing finger phalanges 2 and 3 as the largest, then phalanx 1 and with 4 almost always being the shortest. However, here we see an apparently unique pattern where the first three of these elements are almost subequal being respectively 44, 48, and 46 mm long. (The exact length of the restored phalanx 4 being unknown, but with 27 mm preserved and not yet apparently beginning to taper to a point, it cannot have been too much shorter than these others and we restore this as being c. 44 mm in length). Although it is impossible to rule out intraspecific variation, the high level of conservatism of the components of the pterosaur skeleton including the wing finger, (in some taxa at least - Habib and Hone, 2024) argue that such proportions are likely genuine and represent a distinct anatomical (and therefore taxonomic) feature (Figure 8).
It was originally argued that the early monofenestratans were a distinct intermediate between the then known pterodactyloids and non-pterodactyloids (Lü et al., 2010). However, subsequent discoveries have begun to break down this distinction with more variation seen in features like the length of the wing metacarpals and tails (Sullivan et al., 2014; Hone et al., 2024). This shows that there was, perhaps inevitably, less of a disjunct between early pterosaurs and the first monofenestratans, and between these and the pterodactyloids. As such, some variation and plasticity, and perhaps evolutionary experimentation with this intermediate bauplan is to be expected, and may have resulted in the unusual proportional sizes of some elements. The evolution from the non-pterodactyloid to the pterodactyloid condition requires the elongation of phalanx 1 and reduction of 4, and it is reasonable that at some point there were derived non-pterodactyloids that would show a transitional proportion of a wing with phalanx 1 shorter than their descendants, but longer than their ancestors, and this may be the case here. Other pterosaurs in this grade may add further to unravelling this transition as, for example, Darwinopterus robustodens also has an unusually long wing phalanx 4 that is longer than 1, although both are much shorter than 2 and 3 (Lü et al., 2011).
The distal wing likely functioned as a single unit as the joints between the individual phalanges was stiff and would not normally flex in pterosaurs (anurognathids are an apparent exception to this - Hone, 2020). Longer elements would be more vulnerable to bending, and forces may be greater on distal part of the wings though leverage. This may explain the pterodactyloid condition of a long phalanx 1 through to a short 4, keeping lower stresses on the longer wing elements that would be more proximal, while the distal phalanges that would have the highest forces are kept short and therefore strong. This may have been important for the pterodactyloids which were generally larger and heavier animals and so would experience higher stresses in flight (the largest pterosaurs, the azhdarchids notably have a greatly reduced distal wing phalanges - Witton, 2013). This, though, does not explain why this was adopted so early on in pterodactyloid evolution in the generally small ctenochasmatoids which were comparable in size or even smaller than contemporary non-pterodactyloids.
An alternate explanation is the move into more terrestrial environments by the monofenestratans. If these animals were living in more inland ecosystems, then they were likely flying in cluttered systems and with the potential for impacts on the wing phalanges during flight and potentially during take-offs and landing. Such a habitat preference has now been suggested with all but the most derived early monofenestratans showing affinities for climbing (Smyth et al., 2024). As above, longer elements would be more vulnerable to damage and the distal wing elements would naturally be more likely to impact on objects than the proximal ones (see also Hone et al., 2015 on pterosaur wingtips). Therefore, shortening the distal phalanges while lengthening the proximal ones would mean that the wing finger could be the same functional length, while the distal ones became less likely to be damaged if they struck something.
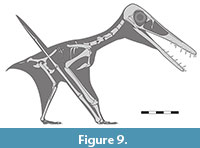 On a similar note, ancestrally, reptiles (including archosaurs) have a pterodactyloid-like pattern in the lengths of the phalanges of manual digit IV with each shorter than the preceding elements (Romer, 1956, figures 182, 183, 184), with the exception of the ungual, though this is absent in pterosaurs on this digit. This means that there must have been some strong selective pressure in ancestral pterosaurs to have evolved phalanges 2 and 3 to become longer than 1, and to retain this pattern until the origin of the pterodactyloids. This would represent a developmental shift in the growth of the phalanges, and that this was selected because of a functional reason that was important in early pterosaurs. The stresses and functions of different wing phalanx lengths are beyond the scope of this work, but the unique arrangement seen here in Makrodactylus oligodontus (Figure 9), and the functions of differing lengths of wing phalanges is worthy of further investigation.
On a similar note, ancestrally, reptiles (including archosaurs) have a pterodactyloid-like pattern in the lengths of the phalanges of manual digit IV with each shorter than the preceding elements (Romer, 1956, figures 182, 183, 184), with the exception of the ungual, though this is absent in pterosaurs on this digit. This means that there must have been some strong selective pressure in ancestral pterosaurs to have evolved phalanges 2 and 3 to become longer than 1, and to retain this pattern until the origin of the pterodactyloids. This would represent a developmental shift in the growth of the phalanges, and that this was selected because of a functional reason that was important in early pterosaurs. The stresses and functions of different wing phalanx lengths are beyond the scope of this work, but the unique arrangement seen here in Makrodactylus oligodontus (Figure 9), and the functions of differing lengths of wing phalanges is worthy of further investigation.
ACKNOWLEDGEMENTS
We thank the Lauer Foundation for support on this project. We thank Roland Pöschl for finding the specimen, Rolf Kriegerbarthold for preparation work, and Uli Leonhardt for help on identification of the exact location where the specimen was recovered. Thanks to Dr. Christopher Vittore, MD, of the Radiology Department at Rockford Memorial Hospital, Rockford, Illinois, USA for taking the X-ray used in Figure 3. We thank Adam Fitch for discussion of pterosaur anatomy and diagnoses. We thank the editor and two anonymous referees for comments that helped to improve an earlier version of this manuscript.
REFERENCES
Andres, B., Clark, J., and Xu, X. 2014. The Earliest Pterodactyloid and the Origin of the Group. Current Biology, 24:1011-6.
https://doi.org/10.1016/j.cub.2014.03.030
Barrett, P.M., Butler, R.J., Edwards, N.P. and Milner, A.R. 2008. Pterosaur distribution in time and space: an atlas. Zitteliana, 28:61-107.
Barthel, K.W., Swinburn, N.H.M., and Conway Morris, S. 1990. Solnhofen: a study in Mesozoic palaeontology. Cambridge University Press, Cambridge, UK.
Bennett, S.C. 1996. Year-classes of pterosaurs from the Solnhofen Limestone of Germany: taxonomic and systematic implications. Journal of Vertebrate Paleontology, 16:432-444.
https://doi.org/10.1080/02724634.1996.10011332
Bennett, S.C. 2001. The Osteology and Functional Morphology of the Late Cretaceous Pterosaur Pteranodon Part I. General Description of Osteology. Palaeontographica Abteilung A: Palaozoologie, 260:1-112.
https://doi.org/10.1127/pala/260/2001/1
Bennett, S.C. 2007. A second specimen of the pterosaur Anurognathus ammoni. Paläontologische Zeitschrift, 81:376-398.
Cheng, X., Jiang, S., Wang, X. and Kellner, A.W. 2017. Premaxillary crest variation within the Wukongopteridae (Reptilia, Pterosauria) and comments on cranial structures in pterosaurs. Anais da Academia Brasileira de Ciências, 89:119-130.
https://doi.org/10.1590/0001-3765201720160742
Cheng, X., Jiang, S., Bantim, R.A., Sayão, J.M., Saraiva, A. Á., Meng, X., Kellner, A.W. and Wang, X. 2025. A new species of Darwinopterus (Wukongopteridae, Pterosauria) from western Liaoning provides some new information on the ontogeny of this clade. Anais da Academia Brasileira de Ciências, 97:p.e20240707.
https://doi.org/10.1590/0001-3765202520240707
Chinsamy, A., Codorniú, L. and Chiappe, L. 2008. Developmental growth patterns of the filter-feeder pterosaur, Pterodaustro guinazui. Biology Letters, 4:282-285.
https://doi.org/10.1098/rsbl.2008.0004
Collini, C.A. 1784. Sur quelques Zoolithes du Cabinet d’Histoire naturelle de SASE Palatine & de Bavière, à Mannheim. Acta Academiae Theodoro Palatinae, Mannheim, Pars Physica, 5:58-103.
Fernandes, A.E., Pol, D. and Rauhut, O.W. 2024. The oldest monofenestratan pterosaur from the Queso Rallado locality (Cañadón Asfalto Formation, Toarcian) of Chubut Province, Patagonia, Argentina. Royal Society Open Science, 11:p.241238.
https://doi.org/10.1098/rsos.241238
Frey, E., Tischlinger, H., Buchy, M.C. and Martill, D.M. 2003. New specimens of Pterosauria (Reptilia) with soft parts with implications for pterosaurian anatomy and locomotion. Geological Society, London, Special Publications, 217:233-266.
Habib, M.B. and Hone, D.W.E. 2024. Intraspecific variation in the pterosaur Rhamphorhynchus muensteri - implications for flight and socio-sexual signaling. PeerJ, 12:17524.
Hone, D.W.E., Van Rooijen, M.K. and Habib, M.B. 2015. The wingtips of the pterosaurs: anatomy, aeronautical function and ecological implications. Palaeogeography, Palaeoclimatology, Palaeoecology, 440:431-439.
https://doi.org/10.1016/j.palaeo.2015.08.046
Hone, D.W. 2020. A review of the taxonomy and palaeoecology of the Anurognathidae (Reptilia, Pterosauria). Acta Geologica Sinica-English Edition, 94:1676-1692.
Hone, D.W., Ratcliffe, J.M., Riskin, D.K., Hermanson, J.W. and Reisz, R.R. 2021. Unique near isometric ontogeny in the pterosaur Rhamphorhynchus suggests hatchlings could fly. Lethaia, 54:106-112.
https://doi.org/10.1111/let.12391
Hone, D.W. 2023. The anatomy and diversity of the pterosaurian sternum. Palaeontologia Electronica, 26:1-34.
https://doi.org/10.26879/1261
Hone, D.W., Lauer, R., Lauer, B. and Spindler, F. 2023. Petrodactyle wellnhoferi gen. et sp. nov.: A new and large ctenochasmatid pterosaur from the Late Jurassic of Germany. Palaeontologia Electronica, 26:1-28.
https://doi.org/10.26879/1251
Hone, D.W.E., Fitch, A.J., Selzer, S., Lauer, R., and Lauer, B. 2024. A new and large darwinopteran reveals the evolutionary transition to the pterodactyloid pterosaurs. Current Biology. 34:p5607-5614.
https://doi.org/10.1016/j.cub.2024.10.023
Hone, D.W.E., and McDavid, S. 2024. A giant specimen of Rhamphorhynchus muensteri and comments on the ontogeny of rhamphorhynchines. PeerJ, 13:e18587.
https://doi.org/10.7717/peerj.18587
Jagielska, N., O’Sullivan, M., Funston, G.F., Butler, I.B., Challands, T.J., Clark, N.D., Fraser, N.C., Penny, A., Ross, D.A., Wilkinson, M. and Brusatte, S.L. 2022. A skeleton from the Middle Jurassic of Scotland illuminates an earlier origin of large pterosaurs. Current Biology, 32:1446-1453.
https://doi.org/10.1016/j.cub.2022.01.073
Jiang, S., Cheng, X.; Ma, Y., and Wang, X. 2016. A new archaeopterodactyloid pterosaur from the Jiufotang Formation of western Liaoning, China, with a comparison of sterna in Pterodactylomorpha. Journal of Vertebrate Paleontology, 36:e1212058
https://doi.org/10.1080/02724634.2016.1212058
Jiang, S., Wang, X., Zheng, X., Cheng, X., Zhang, J. and Wang, X. 2021. An early juvenile of Kunpengopterus sinensis (Pterosauria) from the Late Jurassic in China. Anais da Academia Brasileira de Ciências, 93:p.e20200734.
https://doi.org/10.1590/0001-3765202120200734
Kellner, A.W. 2015. Comments on Triassic pterosaurs with discussion about ontogeny and description of new taxa. Anais da Academia Brasileira de Ciências, 87:669-689.
Lü, J. 2009. A new non-pterodactyloid pterosaur from Qinglong County, Hebei Province of China. Acta Geologica Sinica (English Edition), 83:189-199.
https://doi.org/10.1111/j.1755-6724.2009.00062.x
Lü, J.C. and Fucha, X.H. 2010. A new pterosaur (Pterosauria) from Middle Jurassic Tiaojishan formation of western Liaoning, China. Global Geology, 13:113-118.
https://doi.org/10.3969/j.issn.1673-9736.2010.03.04.01
Lü, J., Unwin, D.M., Jin, X., Liu, Y. and Ji, Q. 2010. Evidence for modular evolution in a long-tailed pterosaur with a pterodactyloid skull. Proceedings of the Royal Society B: Biological Sciences, 277:383-389.
https://doi.org/10.1098/rspb.2009.1603
Lü, J., Unwin, D.M., Deeming, D.C., Jin, X., Liu, Y., and Ji, Q. 2011a. An egg-adult association, gender, and reproduction in pterosaurs. Science, 331:321-324.
https://doi.org/10.1126/science.1197323
Lü, J., Xu, L., Chang, H. and Zhang, X. 2011b. A new darwinopterid pterosaur from the Middle Jurassic of western Liaoning, northeastern China and its ecological implications. Acta Geologica Sinica-English Edition, 85:507-514.
Lü, J., Unwin, D.M., Zhao, B., Gao, C. and Shen, C. 2012. A new rhamphorhynchid (Pterosauria: Rhamphorhynchidae) from the Middle/Upper Jurassic of Qinglong, Hebei Province, China. Zootaxa, 3158:1-19.
Martill, D.M. and Etches, S. 2012. A new monofenestratan pterosaur from the Kimmeridge Clay Formation (Kimmeridgian, Upper Jurassic) of Dorset, England. Acta Palaeontologica Polonica, 58:285-294.
https://doi.org/10.4202/app.2011.0071
Martill, D.M., Frey, E., Tischlinger, H., Mäuser, M., Rivera-Sylva, H.E. and Vidovic, S.U. 2023. A new pterodactyloid pterosaur with a unique filter-feeding apparatus from the Late Jurassic of Germany. PalZ, 97:383-424.
https://doi.org/10.1007/s12542-022-00644-4
Martin-Silverstone, E., Unwin, D.M., Cuff, A.R., Brown, E.E., Allington-Jones, L. and Barrett, P.M., 2024. A new pterosaur from the Middle Jurassic of Skye, Scotland and the early diversification of flying reptiles. Journal of Vertebrate Paleontology, 43:p.e2298741.
https://doi.org/10.1080/02724634.2023.2298741
Ősi, A. and Prondvai, E. 2009. Forgotten pterosaurs in Hungarian collections: first description of Rhamphorhynchus and Pterodactylus specimens. Neues Jahrbuch für Geologie und Paläontologie-Abhandlungen, 252:167-180.
https://doi.org/10.1127/0077-7749/2009/0252-0167
Prondvai, E. and Ősi, A. 2011. Potential for intracranial movements in pterosaurs. The Anatomical Record, 294: 813-830.
https://doi.org/10.1002/ar.21376
Prondvai, E., Bodor, E.R. and Ősi, A. 2014. Does morphology reflect osteohistology-based ontogeny? A case study of Late Cretaceous pterosaur jaw symphyses from Hungary reveals hidden taxonomic diversity. Paleobiology, 40:288-321.
https://doi.org/10.1666/13030
Rauhut, O.W., Heyng, A.M. and Leonhardt, U. 2011. Neue Reptilfunde aus der Mörnsheim− Formation von Mühlheim. Freunde der Bayerischen Staatssammlung für Paläontologie und Historische Geologie, München, 39:61-71.
Rauhut, O.W.M. 2012. Ein “Rhamphodactylus” aus der Mörnsheim-Formation von Mühlheim. Jahresbericht 2011 und Mitteilungen der Freunde der Bayerischen Staatssammlung für Paläontologie und Historische Geologie München, 40:69-74.
Romer, A.S. 1965. The osteology of the reptiles (second impression). University of Chicago Press, Chicago. 772pp.
Smyth, R.S., Breithaupt, B.H., Butler, R.J., Falkingham, P.L. and Unwin, D.M. 2024. Hand and foot morphology maps invasion of terrestrial environments by pterosaurs in the mid-Mesozoic. Current Biology, 34:4894-4907.
https://doi.org/10.1016/j.cub.2024.09.014
Spindler, F. 2024. A pterosaurian connecting link from the Late Jurassic of Germany. Palaeontologica Electronica, 27:a35.
https://doi.org/10.26879/1366
Sullivan, C., Wang, Y., Hone, D.W., Wang, Y., Xu, X. and Zhang, F. 2014. The vertebrates of the Jurassic Daohugou Biota of northeastern China. Journal of Vertebrate Paleontology, 34:243-280.
https://doi.org/10.1080/02724634.2013.787316
Tischlinger, H., and Frey, E. 2013. A new pterosaur with mosaic characters of basal and pterodactyloid pterosauria from the Upper Kimmeridgian of Painten (Upper Palatinate, Germany). Archaeopteryx, 31:1-13.
Wang, X., Kellner, A.W., Jiang, S. and Meng, X. 2009. An unusual long-tailed pterosaur with elongated neck from western Liaoning of China. Anais da Academia Brasileira de Ciências, 81:793-812.
https://doi.org/10.1590/S0001-37652009000400016
Wang, X., Kellner, A.W., Jiang, S., Cheng, X., Meng, X. and Rodrigues, T. 2010. New long-tailed pterosaurs (Wukongopteridae) from western Liaoning, China. Anais da Academia Brasileira de Ciências, 82:1045-1062.
https://doi.org/10.1590/S0001-37652010000400024
Wang, X., Jiang, S., Zhang, J., Cheng, X., Yu, X., Li, Y., Wei, G. and Wang, X. 2017. New evidence from China for the nature of the pterosaur evolutionary transition. Scientific Reports, 7:1-9.
https://doi.org/10.1038/srep42763
Wellnhofer, P. 1970. Die Pterodactyloidea (Pterosauria) der Oberjura-Plattenkalke Süddeutschlands. - Bayerische Akademie der Wissenschaften, Mathematisch- Wissenschaftlichen Klasse, Abhandlugen, 141:1-133.
Wellnhofer, P. 1975. Die Rhamphorhynchoidea (Pterosauria) der Oberjura-Plattenkalke Süddeutschlands. Palaeontographica, Abt. A, 148:1-13.
Wilkin, J.T.R. 2019. taphonomy of Tithonian Fishes from the Mörnsheim Formation of southern Germany. Zitteliana, 93:83-88.
Witton, M.P. 2013. Pterosaurs. Princeton University Press, Los Angeles, USA.
https://doi.org/10.1515/9781400847655
Zhou, C.F. and Schoch, R.R. 2011. New material of the non-pterodactyloid pterosaur Changchengopterus pani Lü, 2009 from the Late Jurassic Tiaojishan Formation of western Liaoning. Neues Jahrbuch fur Geologie und Paläontologie-Abhandlungen, 260:265-275.
https://doi.org/10.1127/0077-7749/2011/0131
Zhou, X., Pêgas, R.V., Ma, W., Han, G., Jin, X., Leal, M.E., Bonde, N., Kobayashi, Y., Lautenschlager, S., Wei, X. and Shen, C. 2021. A new darwinopteran pterosaur reveals arborealism and an opposed thumb. Current Biology, 31:2429-2436.
https://doi.org/10.1016/j.cub.2021.03.030
