


Further contributions to the early Miocene forest vegetation of the Galatian Volcanic Province, Turkey
Article number: 21.3.40
https://doi.org/10.26879/816
Copyright Paleontological Society, November 2018
Author biographies
Plain-language and multi-lingual abstracts
PDF version
Submission: 4 September 2017. Acceptance: 2 November 2018
{flike id=2353}
ABSTRACT
Ten new petrified forest sites have been discovered on the Galatian Volcanic Province (GVP). The silicified woods, preserved in the lower Miocene Hançili and Pazar formations, represent a diverse plant assemblage that grew across western and central GVP during the Burdigalian, ca. 20-17 Ma. The assemblage comprises a total of 233 petrified woods collected from the 10 sites in and around Ankara (Beypazarı, Çamlıdere, Kızılcahamam) and Bolu (Kıbrısçık). Taxonomic identifications of the fossil wood floras suggest four distinct vegetational associations can be recognized on the GVP: (1) swamp associations of Glyptostrobus Endlicher, Sequoia Endlicher and Taxodium Richard; (2) riparian forest comprising palms, Salix L., Liquidambar L. and Quercus L. section Ilex ; (3) mesic-xeric woodland comprising Acer L., Quercus L., Pinus L. and Juniperus L.; and (4) mixed coniferous-angiosperm forest comprising Picea A. Dietrich, Cedrus Trew., Pinus, Juniperus L. and Podocarpus L’Herit (ex Pers.) alongside. These associations support the idea that the landscape was varied and the climate warm and frost free.
N. Neslihan Acarca Bayam. Çapa Science College, 34104 Çapa-İstanbul, Turkey. nneslihan.acarca@gmail.com
Ünal Akkemik. İstanbul University, Faculty of Forestry, Department of Forest Botany, 34473 Bahçeköy-Sarıyer, İstanbul, Turkey. uakkemik@istanbul.edu.tr
Imogen Poole. Aberdeen, United Kingdom. poole.imogen@gmail.com
Ferdi Akarsu. İstanbul University, Faculty of Forestry, Department of Forest Botany, 34473 Bahçeköy-Sarıyer, İstanbul, Turkey. fakarsu@istanbul.edu.tr
Keywords: Miocene; silicified wood; petrified forest; wood identification; Galatian Volcanic Province; Köroglu Volcanics, Turkey
Final citation: Acarca Bayam, N. Neslihan, Akkemik, Ünal, Poole, Imogen, and Akarsu, Ferdi 2018. Further contributions to the early Miocene forest vegetation of the Galatian Volcanic Province, Turkey. Palaeontologia Electronica 21.3.40A 1-42. https://doi.org/10.26879/816
palaeo-electronica.org/content/2018/2353-trees-of-gvp-in-miocene
Copyright: November 2018 Paleontological Society.
This is an open access article distributed under the terms of Attribution-NonCommercial-ShareAlike 4.0 International (CC BY-NC-SA 4.0), which permits users to copy and redistribute the material in any medium or format, provided it is not used for commercial purposes and the original author and source are credited, with indications if any changes are made.
creativecommons.org/licenses/by-nc-sa/4.0/
INTRODUCTION
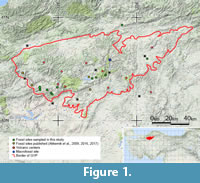 The Galatean Volcanic Province (GVP) (or Köroglu Volcanics) is a major Neogene volcanic belt covering an area of approximately 12,800 km2 (Koçyiğit et al., 2003) and located in central Anatolia in the northwestern part of Turkey (Toprak et al., 1996). The GVP and three other volcanic provinces in western, central and eastern Anatolia together form the four main Neogene and Quaternary volcanic fields in Turkey (Kazancı, 2012) that evolved as a consequence of the tectonic collision between the African Plate and the Eurasian Plate (Wilson et al., 1997; Tankut et al., 1998; Rögl, 1999; Koçyiğit et al., 2003). Most of the volcanic products of the GVP belong to the eruptions from nine eruption centers (Figure 1) so far identified, which grade from basalts to andesite and rhyolites (Kazancı, 2012). Rocks were deposited during three major volcanic phases: (1) late Campanian-Lutetian subductional phase; (2) post-Lutetian collisional phase; and (3) Oligocene-Miocene post-collisional phase (Koçyiğit et al., 2003).
The Galatean Volcanic Province (GVP) (or Köroglu Volcanics) is a major Neogene volcanic belt covering an area of approximately 12,800 km2 (Koçyiğit et al., 2003) and located in central Anatolia in the northwestern part of Turkey (Toprak et al., 1996). The GVP and three other volcanic provinces in western, central and eastern Anatolia together form the four main Neogene and Quaternary volcanic fields in Turkey (Kazancı, 2012) that evolved as a consequence of the tectonic collision between the African Plate and the Eurasian Plate (Wilson et al., 1997; Tankut et al., 1998; Rögl, 1999; Koçyiğit et al., 2003). Most of the volcanic products of the GVP belong to the eruptions from nine eruption centers (Figure 1) so far identified, which grade from basalts to andesite and rhyolites (Kazancı, 2012). Rocks were deposited during three major volcanic phases: (1) late Campanian-Lutetian subductional phase; (2) post-Lutetian collisional phase; and (3) Oligocene-Miocene post-collisional phase (Koçyiğit et al., 2003).
Keller et al. (1992), Wilson et al. (1997) and Adıyaman et al. (2001) suggest that during the Oligocene-Miocene volcanic phase three episodes of volcanism took place across the GVP: the first around 22-19 Ma, followed by a second episode between 18-17 Ma and a third episode of volcanism during the late Miocene (10-9 Ma). Potassium-argon dating of volcanic rocks determined that the major phase of volcanic activity took place during the early Miocene (20-17 Ma; Wilson et al., 1997), which resulted in the accumulation of thick volcanic and volcaniclastic rocks intercalated with continental sedimentary rock (Toprak et al., 1996).
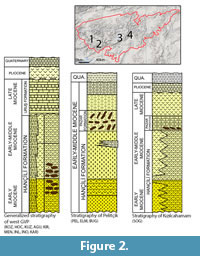 The fossil wood material from the western part of the GVP originated from the lower part of the early to early-middle Miocene Hançili Formation (Akbaş et al., 2002; Altun et al., 2002) (Figure 2), a sequence of conglomerates, coals and organic-rich shales and claystones reflecting deposition in lacustrine and coastal environments under warm, humid climatic conditions (Vosoughi Moradi et al., 2016). Radiometric dating of the early Miocene volcanic basalts across the western part of the GVP have been undertaken by several workers: Toprak et al. (1996), summarizing Türkecan et al. (1991) and Keller et al. (1992), give the age of the exposed volcanoclastic rocks found in the south-western part of the Kartalkaya-Köroglu Complex near Seben (Figure 1) in the western part of the GVP as ranging from 24.83 ± 2.01 to 21.79 ± 4.93 Ma. Subsequently Akkemik et al. (2016) undertook radiometric dating of volcanic rocks taken from around the silicified wood samples that originated from the Seben-Hoçaş Fossil Forest (Bolu Province, Figure 1), just west of the sites sampled in this study. These returned an age of 18.2 ± 0.8 Ma (early-middle Burdigalian). These data suggest an age range of ca. 24.8-18 Ma (Aquitanian-Burdigalian) for the fossil woods found in the western part of the GVP.
The fossil wood material from the western part of the GVP originated from the lower part of the early to early-middle Miocene Hançili Formation (Akbaş et al., 2002; Altun et al., 2002) (Figure 2), a sequence of conglomerates, coals and organic-rich shales and claystones reflecting deposition in lacustrine and coastal environments under warm, humid climatic conditions (Vosoughi Moradi et al., 2016). Radiometric dating of the early Miocene volcanic basalts across the western part of the GVP have been undertaken by several workers: Toprak et al. (1996), summarizing Türkecan et al. (1991) and Keller et al. (1992), give the age of the exposed volcanoclastic rocks found in the south-western part of the Kartalkaya-Köroglu Complex near Seben (Figure 1) in the western part of the GVP as ranging from 24.83 ± 2.01 to 21.79 ± 4.93 Ma. Subsequently Akkemik et al. (2016) undertook radiometric dating of volcanic rocks taken from around the silicified wood samples that originated from the Seben-Hoçaş Fossil Forest (Bolu Province, Figure 1), just west of the sites sampled in this study. These returned an age of 18.2 ± 0.8 Ma (early-middle Burdigalian). These data suggest an age range of ca. 24.8-18 Ma (Aquitanian-Burdigalian) for the fossil woods found in the western part of the GVP.
The fossils found in the central part of the GVP were recovered from the Pazar Formation (Figure 2), a sequence of argillaceous limestones, marl and claystones with locally cherty limestone, sandstone and tuff, and occasional thin coal seams (Süzen and Türkmenoglu, 2000; Yavuz-Işık and Demirci, 2009; Sen et al., 2017). Tatlı (1975) stated that Pazar Formation was formed in lake and swamp environments. Sen et al. (2017) suggested that the Pazar Formation was deposited during the early and early middle Miocene. The fossils originated from the basal layers of the Pazar Formation at Kizilcahamam, but were more evenly dispersed throughout the formation at the Pelitcik sites (Figure 2). Representative basalt samples from the volcanic layers covering the central part of the GVP, near the towns of Özmus and Güdül (Figure 1), were analyzed by Wilson et al. (1997) and returned ages of 18.2 ± 0.5 Ma and 18.8 ± 0.6 Ma (both middle Burdigalian), respectively. Further basalt samples from Güvem and Çamlıdere (Figure 1) returned an age of middle to late Burdigalian (18.2 [± 0.5] - 16.9 [± 0.5] Ma, respectively) (Table 1) (Tankut et al., 1995; Wilson et al., 1997). Based on this evidence we consider the age of the fossils preserved in the same volcanic rocks covering the central part of the GVP to be middle to late Burdigalian and thus coeval or slightly younger that those fossils found in the west. Given that the two regions are interpreted to have formed from the same volcanic processes, and that there exists a degree of overlap in the ages of these two regions, until further dating is undertaken we consider the material found in the western and the central region of the GVP to be relatively contemporaneous.
The GVP is one of the most important areas for fossil forest preservation in Turkey (Akkemik et al., 2016 and references cited therein) at a time when particularly warm conditions favoured the growth of rich forest vegetation across the Northern Hemisphere (Mai, 1995; Zachos et al., 2008; Denk et al., 2011, 2017a, 2017b, 2017c). Fossilized animal remains alongside petrified wood, leaf, fruit and cones are commonly found in the lower Miocene volcano-sedimentary units in the central part of the GVP (Kasaplıgil 1977; Dernbach et al., 1996; Akkemik et al., 2009, 2016, 2017; Denk et al., 2017b). Here evidence supports a mixed conifer-deciduous and evergreen angiosperm early Miocene vegetation growing in the Kizilcahamam-Camlidere-Güvem area (Figure 1), with Sequoia, Taxodium Rich., Quercus section Ilex, Acer, Salix, Populus, Ulmus, Zelkova, Alnus Mill. Betula L. Fabaceae, Engelhardioideae (Akkemik et al., 2009; Dernbach et al., 1996; Kasaplıgil, 1977; Denk et al., 2017b). To the south, near Güdül (Figure 1), Kasapligil (1977) and Akkemik et al. (2017) identified leaves and woods belonging to Glyptostrobus and Glyptostroboxylon Conwentz and concluded this taxon formed a main constituent of a prevailing swamp forest vegetation. Further north, near Pelitçik (Figure 1), a Sequoia type wood along with wood of Taxodium were identified as representative forest trees (Akkemik et al., 2009). The ‘Sequoia type’ wood is distinct in that it includes anatomical features of Sequoia, Sequoiadendron J. Buchholz and Metasequoia Miki and thus cannot be identified as belonging to either one of these modern genera; with better preserved samples a closer identification may be possible.
In the western part of the GVP, Akkemik et al. (2016) also described wood samples recovered from two early Miocene sites: one near the village of Hoçaş and the second near the village of Kozyaka (Figure 1). Both sites lie at an altitude of ~900 m close to the town of Seben (Figure 1) in the province of Bolu, and together are referred to as the Seben Fossil Forest. The Hoçaş site had preserved both in situ and in situ samples that include Palmae, Salix L., Liquidambar, Quercus section Ilex and Juniperus, Acer, Cedrus and Picea. The Kozyaka site also yielded samples of Acer, Quercus L. (Ilex type), Cedrus, along with Salix/Populus L., Ulmus L., and Pinus (Akkemik et al., 2016).
Although these studies have provided an indication of the local vegetation that grew across the western and central parts of the GVP during the early Miocene, ideas pertaining to the flora that grew here are still limited. The discovery of new petrified wood sites and the subsequent analyses of the wood samples discussed in context of the vegetational hypotheses presented in previous publications, forms the basis of this work and helps further understanding of the flora and ecological dynamics that prevailed across Anatolia during the Miocene.
MATERIAL AND METHODS
 Ten new petrified forest sites within GVP have been identified (Figure 1, Table 1). From these sites a total of 233 silicified wood samples were collected ranging from 3 to 30 cm in length/height and 0.5 to 17 cm in diameter. The woods comprise both in situ stumps (Figure 3) decapitated by a volcanic eruption, but still in their life position (authochthonous), or relatively small pieces of in situ (allochthonous) material including those specimens from trees felled by volcanic activity and subsequently moved from the original growth position (Figure 4). Preservation is variable with some specimens showing good preservation, whilst others are poorly preserved. The in situ fossil wood specimens collected from the Çamlıdere-Elmalı Village site (ELM) were generally too badly preserved for any further analysis.
Ten new petrified forest sites within GVP have been identified (Figure 1, Table 1). From these sites a total of 233 silicified wood samples were collected ranging from 3 to 30 cm in length/height and 0.5 to 17 cm in diameter. The woods comprise both in situ stumps (Figure 3) decapitated by a volcanic eruption, but still in their life position (authochthonous), or relatively small pieces of in situ (allochthonous) material including those specimens from trees felled by volcanic activity and subsequently moved from the original growth position (Figure 4). Preservation is variable with some specimens showing good preservation, whilst others are poorly preserved. The in situ fossil wood specimens collected from the Çamlıdere-Elmalı Village site (ELM) were generally too badly preserved for any further analysis.
Each petrified wood sample is coded using the first three letters of the site name (Table 1) and numbered sequentially. Those samples found in situ are identified by ‘D’ following the site name identifier (and prefixing the sample number). Samples are deposited in the Wood Anatomy Laboratory of the Forest Botany Department, Faculty of Forestry at Istanbul University.
 Sixty-two of the samples were thin sectioned along the three planes of section (transverse section, TS, radial longitudinal section, RLS, and tangential longitudinal section, TLS), and identified using transmitted light microscopy. Descriptions follow the terminology of the IAWA Committee (1989) for hardwood identification and IAWA Committee (2004) for softwood identification wherever possible. Measurements were taken and documented using guidelines recommended by the IAWA Committee (1989, 2004) (i.e., the mean followed by minimum and maximum measurements in parentheses). Identifications were made following reference to well-described extant wood, such as vouchered specimens housed in Forest Botany Department of Faculty of Forestry, University of Istanbul, comparisons with published literature (Jacquiot, 1955; Greguss, 1955, 1967; Barefoot and Hankins, 1982; Fahn et al., 1986; Schweingruber, 1990, 1993; Tidwell, 1998; Eliçin, 1977; Vassio et al., 2008; Wheeler, 2011; Akkemik and Yaman, 2012; Uhl et al., 2014) and online databases such as InsideWood for angiosperm wood (http://insidewood.lib.nsc.edu).
Sixty-two of the samples were thin sectioned along the three planes of section (transverse section, TS, radial longitudinal section, RLS, and tangential longitudinal section, TLS), and identified using transmitted light microscopy. Descriptions follow the terminology of the IAWA Committee (1989) for hardwood identification and IAWA Committee (2004) for softwood identification wherever possible. Measurements were taken and documented using guidelines recommended by the IAWA Committee (1989, 2004) (i.e., the mean followed by minimum and maximum measurements in parentheses). Identifications were made following reference to well-described extant wood, such as vouchered specimens housed in Forest Botany Department of Faculty of Forestry, University of Istanbul, comparisons with published literature (Jacquiot, 1955; Greguss, 1955, 1967; Barefoot and Hankins, 1982; Fahn et al., 1986; Schweingruber, 1990, 1993; Tidwell, 1998; Eliçin, 1977; Vassio et al., 2008; Wheeler, 2011; Akkemik and Yaman, 2012; Uhl et al., 2014) and online databases such as InsideWood for angiosperm wood (http://insidewood.lib.nsc.edu).
SYSTEMATIC PALAEOBOTANY
A total of 14 different genera were identified from 10 fossil sites across the GVP. Seven of them were conifers, and the rest angiosperms. A summary of the wood characters of each taxon outlined below is based on all samples listed. Measurements for each individual sample are given in Appendix 1 and Appendix 2.
Family CUPRESSACEAE Rich. ex Bartling, 1830
Genus JUNIPERUS Linnaeus, 1753
Juniperus sp.
Material. AGU04, AGU05, AGU09, AGU11 (in situ); AGUD3, AGUD7, AGUD8, AGUD10 (in situ).
Locality. Near to the village of Beypazarı-Aşağıgüney (AGU).
Description. Ex-situ and in-situ material with dimensions ranging from 4 to 16 cm in height/length, and 2 to 8 cm in diameter.
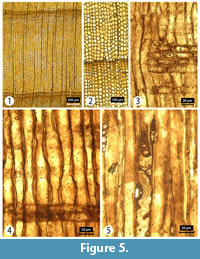 Transverse section (TS). Growth ring widths 0.2-1.5 mm with distinct boundaries. Transition from earlywood to latewood gradual with latewood tracheids forming a layer 1-2 rows deep; false rings common. Resin canals absent. Intercellular spaces obvious and occur throughout the wood. Tracheid tangential and radial diameters measure 15 (5-25) µm and 16 (3-28) µm, respectively, in earlywood, and 9 (2-17) µm and 5 (1-13) µm, respectively, in the latewood. Tracheid wall thickness measures 4 (1-9) µm (thin walled) in earlywood and 6 (2-12) µm (thick walled) in latewood. Axial parenchyma is common, and both diffuse (in the transition from earlywood to latewood and within the latewood itself) and tangentially zonate (Figure 5).
Transverse section (TS). Growth ring widths 0.2-1.5 mm with distinct boundaries. Transition from earlywood to latewood gradual with latewood tracheids forming a layer 1-2 rows deep; false rings common. Resin canals absent. Intercellular spaces obvious and occur throughout the wood. Tracheid tangential and radial diameters measure 15 (5-25) µm and 16 (3-28) µm, respectively, in earlywood, and 9 (2-17) µm and 5 (1-13) µm, respectively, in the latewood. Tracheid wall thickness measures 4 (1-9) µm (thin walled) in earlywood and 6 (2-12) µm (thick walled) in latewood. Axial parenchyma is common, and both diffuse (in the transition from earlywood to latewood and within the latewood itself) and tangentially zonate (Figure 5).
Tangential longitudinal section (TLS). Rays exclusively uniseriate. Ray height is 2-5 (max.12) cells or 45 (9-226) μm; ray width is 8 (4-18) µm in width. Transverse end walls of axial parenchyma cells are clearly, or slightly, nodular (Figure 5). Tracheid pitting in tangential walls is common.
Radial longitudinal section (RLS). Rays homogeneous. Ray tracheids absent. End walls of ray parenchyma cells nodular and indentures are present. Nodular end walls present. Cross-field pitting is cupressoid with 3-5 pits per cross-field. Tracheid pitting in radial walls of the earlywood is predominantly uniseriate (Figure 5).
Comments. The combination of wood features described here is consistent with that of Juniperus and are similar to those of the eight juniper species growing today in Turkey. All eight species grow across the whole of Anatolia with three of them, Juniperus excelsa M. Bieb., J. drupacea Labill. and J. oxycedrus L., being arboreal in habit (Akkemik and Yaman, 2012). Akkemik et al. (2016) also identified Juniperus wood from the Hoçaş Fossil Forest site and given the close anatomical similarity and geographical proximity, we conclude the material described here was probably derived for the same taxon as that found in the Hoçaş Fossil Forest site. Therefore, we assign the material described here to Juniperus species. Recently, Denk et al. (2017b) reported twigs of Juniperus from early Burdigalian deposits of Beskonak (Güvem), which supports evidence for juniper growing in this region during the early Miocene.
Genus SEQUOIA (D. Don) Endlicher, 1847
Sequoia sp.
Material. INO04, BUG20 and SOG04.
Locality. Within the Beypazarı Inozu Valley on the south side (INO), and near the villages of Çamlıdere-Buğralar (BUG) and Kızılcahamam-Soğuksu (SOG).
Description. The following description was based on ex-situ material collected from three sites with dimensions ranging from 4 to 6.5 cm in diameter by 6 to 10.5 cm in height.
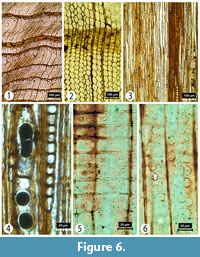 Transverse section (TS). Growth ring widths 0.5-3 mm with distinct boundaries. Transition from earlywood to latewood abrupt with a wide zone (3-22 cells) of latewood tracheids with reduced radial diameters. No resin canals observed. Tracheid tangential and radial diameters measure 26 (11-42) µm and 29 (14-44) µm, respectively, in the earlywood and 17 (5-38) µm and 10 (1-27) µm respectively in the latewood. Tracheid wall thickness measures 3 (1-9) µm (thin walled) in earlywood and 9 (1-18) µm (thick walled) in latewood. Axial parenchyma common and diffuse to tangentially zonate in the transition zone and latewood. Vascular traces absent (Figure 6).
Transverse section (TS). Growth ring widths 0.5-3 mm with distinct boundaries. Transition from earlywood to latewood abrupt with a wide zone (3-22 cells) of latewood tracheids with reduced radial diameters. No resin canals observed. Tracheid tangential and radial diameters measure 26 (11-42) µm and 29 (14-44) µm, respectively, in the earlywood and 17 (5-38) µm and 10 (1-27) µm respectively in the latewood. Tracheid wall thickness measures 3 (1-9) µm (thin walled) in earlywood and 9 (1-18) µm (thick walled) in latewood. Axial parenchyma common and diffuse to tangentially zonate in the transition zone and latewood. Vascular traces absent (Figure 6).
Tangential longitudinal section (TLS). Rays uniseriate and (low) medium to high (2-27 cells), rarely partly-biseriate. Ray height is 174 (28-437) μm; ray width is 13 (6-27) µm. Helical thickenings absent. Axial parenchyma present with either slightly nodular or smooth transverse end walls; brown contents (possible preservational feature) present in each parenchyma cell (Figure 6).
Radial longitudinal section (RLS). Tracheid pitting generally uniseriate in the latewood, and uni- to biseriate and opposite in the earlywood. Crassulae commonly associated with both uniseriate and biseriate pitting. Two to six taxodioid pits per cross-field and horizontally arranged in one or two rows. No ray tracheids observed. Ray parenchyma walls smooth. Axial parenchyma cells with (brown) contents (Figure 6).
Comments. The combination of wood characters described here is consistent with that of the extant Sequoia/Sequoiadendron/Metasequoia complex and very similar to those fossil woods also identified as Sequoia from the Pelitçik Fossil Forest site located between Beypazarı Inozu Valley (INO) and Kızılcahamam-Soğuksu (SOG) (Akkemik et al., 2009). Further evidence for the presence of Sequoia/Sequoiadendron/Metasequoia comes from leaf imprints and pollen identified from the Güvem area (Kasaplıgil, 1977; Denk et al., 2017b). Sequoia and the Sequoia/Sequoiadendron/Metasequoia complex are the most common tree taxa found growing in Anatolia during the Oligocene (Akkemik et al., 2005; Akkemik and Sakınç, 2013) and into the early to mid-Miocene (Kasaplıgil, 1977; Akkemik et al., 2009; Denk et al., 2017b).
Genus TAXODIUM Richard, 1810
Taxodium sp.
Material. BUG01.
Locality. Near to the village of Çamlıdere-Buğralar
Description. The following description is based on one poorly preserved ex-situ sample measuring 12 cm in diameter by 14 cm in length.
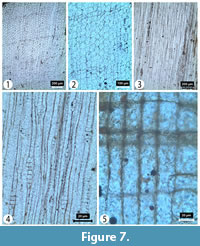 Transverse section (TS). Growth ring boundaries indistinct with ring widths ranging from 0.7 to 2 mm. Transition from earlywood to latewood gradual with a narrow zone (1-3, rarely more) of thin walled, latewood tracheids. Tracheid tangential and radial diameters measure 39 (26-53) µm and 50 (33-63) µm, respectively, in the earlywood and 31 (19-50) µm and 27 (20-37) µm, respectively, in the latewood. Tracheid wall thickness measures 3 (1-5) µm (thin walled) in earlywood and 3 (1-4) µm (thin walled) in latewood. Axial parenchyma diffuses and rare. Resin canals absent (Figure 7).
Transverse section (TS). Growth ring boundaries indistinct with ring widths ranging from 0.7 to 2 mm. Transition from earlywood to latewood gradual with a narrow zone (1-3, rarely more) of thin walled, latewood tracheids. Tracheid tangential and radial diameters measure 39 (26-53) µm and 50 (33-63) µm, respectively, in the earlywood and 31 (19-50) µm and 27 (20-37) µm, respectively, in the latewood. Tracheid wall thickness measures 3 (1-5) µm (thin walled) in earlywood and 3 (1-4) µm (thin walled) in latewood. Axial parenchyma diffuses and rare. Resin canals absent (Figure 7).
Tangential longitudinal section (TLS). Rays uniseriate, rarely partly biseriate and homogenous. Rays measure 422 (303-569) μm in height and 18 (6-27) µm in width. Tracheidal pitting rare, uniseriate and non-contiguous. Helical thickening absent (Figure 7).
Radial longitudinal section (RLS). Tracheid pitting uniseriate or biseriate (rarely triseriate) and opposite. Rays low. Three to seven seemingly taxodioid pits per cross field both horizontally and randomly arranged. Ray tracheids absent. Ray parenchyma walls smooth (Figure 7).
Comments. The combination of wood characters described here is consistent with that of Taxodium. Akkemik et al. (2009) identified this genus from Pelitçik, a site proximal to Çamlıdere-Buğralar Village (Figure 1). Taxodium is one of the more frequently encountered tree genera from the early Miocene with micro- and macrofossils having been identified from Anatolia (Kasaplıgil, 1977; Akkemik et al., 2009), Italy (Biondi and Brugiapaglia, 1991) and Greece (Velitzelos et al., 2014).
Family PINACEAE Sprengel ex F. Rudolphi, 1830
Genus CEDRUS Trew, 1757
Cedrus sp.
Material. INL03, INO05 and INO06.
Localities. Within the Beypazarı-Inozu Valley on the south side (INO) and north side (INL).
Description. All samples are ex-situ with dimensions ranging from 7 to 10 cm in length and 2 to 4.5 cm in diameter.
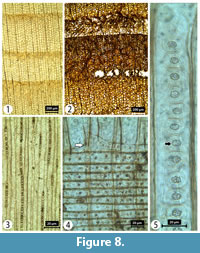 Transverse section (TS). Growth ring width ranging from 0.4 to 1.5 mm. Growth ring boundaries distinct with transition from earlywood to latewood gradual. Latewood zone is generally wide (5-30 cells in radial direction). Tangential and radial tracheid diameters measure 21 (6-38) µm and 30 (15-52) µm, respectively, in the earlywood and 13 (4-26) µm and 7 (1-15) µm, respectively, in latewood. Tracheid wall thickness measures 5 (1-9) µm (thin walled) in earlywood and 12 (5-21) µm (thick walled) in latewood. Axial parenchyma marginal, diffuse or in 2-3 rows. Resin canals absent although traumatic resin canals in both the latewood and transition zones of some rings are common (Figure 8).
Transverse section (TS). Growth ring width ranging from 0.4 to 1.5 mm. Growth ring boundaries distinct with transition from earlywood to latewood gradual. Latewood zone is generally wide (5-30 cells in radial direction). Tangential and radial tracheid diameters measure 21 (6-38) µm and 30 (15-52) µm, respectively, in the earlywood and 13 (4-26) µm and 7 (1-15) µm, respectively, in latewood. Tracheid wall thickness measures 5 (1-9) µm (thin walled) in earlywood and 12 (5-21) µm (thick walled) in latewood. Axial parenchyma marginal, diffuse or in 2-3 rows. Resin canals absent although traumatic resin canals in both the latewood and transition zones of some rings are common (Figure 8).
Tangential longitudinal section (TLS). Rays entirely uniseriate, 13-22 cells high or 215 (27-1004) μm in height and 12 (4-31) µm in width. Many traumatic vertical resin canals observed, rays with horizontal resin canals absent (Figure 8).
Radial longitudinal section (RLS). Rays are heterogeneous. Smooth walled ray tracheids are common and form 1-2 rows with enlarged cells when situated towards the outside of the rays. End- and horizontal walls of ray parenchyma cells are distinctly pitted. Tracheid pitting in radial walls (in early wood) is predominantly uniseriate with scalloped tori. Cross-field pitting is cupressoid with 1-4 pits per cross-field (Figure 8).
Comments. Woods with resin canals also occur in wood of Pinus and Picea but differ in that they are arranged in one row within each growth ring. The combination of wood characters described here is consistent with that of Cedrus. Cedrus fossils have been described from the central part of the GVP. Denk et al. (2017b) reported a seed of Cedrus at Güvem and pollen, which is considered to be relatively common in the Güvem area, by Yavuz-Isik (2008). At the Hoçaş Fossil Forest site in the west Akkemik et al. (2016) identified the wood of Cedrus. The published wood characteristics of extant Cedrus libani A. Rich. (Akkemik and Yaman, 2012) are virtually identical to those of the petrified woods but although identical at species level, when the age of the material is considered we cannot ignore the fact that this material might be a representative of an ancestral lineage (Akkemik et al., 2016) and for this reason these specimens are referred to Cedrus. as Cedrus cf. libani. Today one species, Cedrus libani (Taurus cedar), grows in the Mediterranean Region of Turkey. However, two more species (C. brevifolia A. Henry ex Elwes et A. Henry and C. atlantica [Endl.] Manetti ex Carrière) are native to land surrounding the Mediterranean Sea.
Genus PICEA A. Dietrich, 1824
Picea sp.
Material. AGUD2 and KUZ20.
Localities. Near the villages of Beypazarı-Aşağıgüney and Bolu-Kıbrısçık-Kuzca- Alanhümmet.
Description. The following description is based on two specimens, one in situ AGUD2 in life position whereas the second, KUZ20, is in situ. Dimensions of the samples range from 11-17 cm in height/length and 5.5-7.5 cm in diameter.
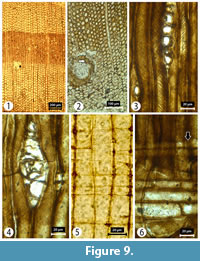 Transverse section (TS). Growth ring boundaries distinct with transition from earlywood to latewood gradual. Growth ring width is very wide, 3.5-9 mm. Tracheid tangential and radial diameters measure 17 (8-26) µm and 24 (9-54) µm, respectively, in earlywood, and 8 (4-16) µm and 7 (1-18) µm, respectively, in latewood. Tracheid wall thickness measures 3 (1-6) µm (thin walled) in earlywood and 5 (2-9) µm (thick walled) in latewood. Resin canals common and surrounded by epithelial cells of varying dimensions (Figure 9), sometimes paired and generally restricted to transition zone; tangential and radial diameters measure 129 (75-192) µm and 141 (112-193) µm, respectively.
Transverse section (TS). Growth ring boundaries distinct with transition from earlywood to latewood gradual. Growth ring width is very wide, 3.5-9 mm. Tracheid tangential and radial diameters measure 17 (8-26) µm and 24 (9-54) µm, respectively, in earlywood, and 8 (4-16) µm and 7 (1-18) µm, respectively, in latewood. Tracheid wall thickness measures 3 (1-6) µm (thin walled) in earlywood and 5 (2-9) µm (thick walled) in latewood. Resin canals common and surrounded by epithelial cells of varying dimensions (Figure 9), sometimes paired and generally restricted to transition zone; tangential and radial diameters measure 129 (75-192) µm and 141 (112-193) µm, respectively.
Tangential longitudinal section (TLS). Rays generally uniseriate unless with horizontal resin canals. Rays with resin canal uniseriate to partly biseriate. Rays measure 298 (128-405) μm in height when resin canals are present, and 144 (28-353) μm in those rays without resin canal; ray widths measure 44 (38-48) µm in those rays with resin canals, and 12 (6-20) μm in rays without resin canals. When present in rays, resin canals are surrounded by a layer of epithelial cells. Helical thickenings are very common in AGUD2 but absent in KUZ20 (Figure 9).
Radial longitudinal section (RLS). Rays heterogeneous. Ray tracheids common in 1-2 rows with smooth, thin walls, rarely slightly wavy. Both horizontal and end walls of ray parenchyma cells are distinctly pitted. Cross-field pitting is cupressoid with 1-3 pits per cross-field. Tracheid pitting in radial walls of the early wood is predominantly uniseriate (Figure 9).
Comments. The combination of wood characters described here is consistent with that of Picea. Today Picea is represented by three species around the Mediterranean and Europe (i.e., Picea abies (L.) Karsten, P. omorika (Panèiæ) Purkyne, and P. orientalis (L.) Link.), but identification to species level is not always possible using wood characters alone. Within these three species, Schweingruber (1990) notes that cross-field pitting is generally cupressoid in Picea abies (and Picea omorika, whereas Akkemik and Yaman (2012) have observed piceoid cross-field pitting in Picea orientalis. Our samples exhibit predominantly cupressoid cross-field pitting suggesting that the material is probably more similar to European Picea species, but the fossil wood differs from all extant European and Mediterranean species by the presence of helical thickening. Therefore, the fossil material described here is assigned to Picea sp.
Genus PINUS Linnaeus, 1753
Pinus sp.
Material. AGU01, AGU22, AGU23, INO12, KIR21, KUZ01, KUZ07, KUZ09, KUZ17, MEN32 and MEN33 (in situ); AGUD12 and KUZ01 (in situ).
Localities. Near the villages of Beypazarı-Aşağıgüney (AGU), Beypazarı-Nuhhoca (Kıraluç Site) (KIR), Bolu-Kıbrısçık-Kuzca- Alanhümmet (KUZ) and Beypazarı-Mençeler (MEN), and in the Beypazarı Inozu Valley (south side) (INO).
Description. In situ and in situ material with dimensions ranging from 9 to 16 cm in height/length by 2 to 8.5 cm in diameter.
T  ransverse section (TS). Growth ring boundaries distinct with transition from earlywood to latewood abrupt in AGU22 and KIR21, and gradual in the others. Width of growth rings range from 1-3.5 to 7 mm with a mode of 4-5 mm; i.e., generally wide except in specimen AGU22. Tracheid tangential and radial diameters measure 25 (6-59) µm and 29 (9-55) µm, respectively, in the earlywood and 10 (1-38) µm and 7 (1-22) µm, respectively, in the latewood; tracheid wall thickness measure 4 (1-15) µm (thin walled) in earlywood and 6 (1-19) µm (thick walled), respectively, in latewood. Resin canals present, frequent in the earlywood-latewood transition zone; thin-walled epithelial cells observed surrounding some resin canals (Figure 10). Tangential and radial diameters of resin canals are 138 (41-354) µm and 139 (61-295) µm, respectively.
ransverse section (TS). Growth ring boundaries distinct with transition from earlywood to latewood abrupt in AGU22 and KIR21, and gradual in the others. Width of growth rings range from 1-3.5 to 7 mm with a mode of 4-5 mm; i.e., generally wide except in specimen AGU22. Tracheid tangential and radial diameters measure 25 (6-59) µm and 29 (9-55) µm, respectively, in the earlywood and 10 (1-38) µm and 7 (1-22) µm, respectively, in the latewood; tracheid wall thickness measure 4 (1-15) µm (thin walled) in earlywood and 6 (1-19) µm (thick walled), respectively, in latewood. Resin canals present, frequent in the earlywood-latewood transition zone; thin-walled epithelial cells observed surrounding some resin canals (Figure 10). Tangential and radial diameters of resin canals are 138 (41-354) µm and 139 (61-295) µm, respectively.
Tangential longitudinal section (TLS). Rays with and without horizontal resin canals. Rays without horizontal canals are uniseriate, 144 (29-425) µm in height and 14 (4-30) µm in width. Rays with resin canals present measure 301 (83-501) µm in height and 41 (13-67) µm in width (Figure 10).
Radial longitudinal section (RLS). Rays heterogeneous. Ray tracheids commonly in 1-4 rows with smooth walls, rarely slightly wavy. Both transverse walls and end walls of ray parenchyma cells are unpitted. Cross-field pitting is clearly pinoid with 1-2 pits per cross-field. Tracheid pitting uniseriate (Figure 10).
Comments. The combination of wood characters such as resin canals, heterogeneous rays and pinoid type cross-field pitting place this morphotaxon within the genus Pinus. Today Turkey is represented by five species of Pinus, namely P. nigra Arnold, P. sylvestris L., P. brutia Ten., P. pinea L. and P. halepensis Miller. The fossils show greatest anatomical similarity to Pinus pinea with its smooth ray tracheids and 1-2 pinoid type of pits per cross-field (Akkemik and Yaman, 2012). Pinus wood has also been identified from the Kozyaka Fossil Forest site (Figure 1) in the west part of GVP (Akkemik et al., 2016), and other macrofossil (leaf and cone) evidence (Kasaplıgil, 1977; Denk et al., 2017b) found at both the Güvem and Hoçaş sites (Figure 1) that revealed Pinus comprised both the three needled and five needled varieties. With this in mind, it cannot be assumed that all Pinus fossil evidence originated from the same taxon. Moreover, since identification to species based on wood anatomy alone is often inconclusive, we refer the specimens described herein to Pinus.
Family PODOCARPACEAE Endlicher, 1847
Genus PODOCARPUS L'Heritier ex Persoon, 1807
Podocarpus sp.
Material. KIR40, KIR70, KIR73 and MEN19.
Localities. Near the village of Beypazarı-Nuhhoca (Kıraluç Site) (KIR) and in the Beypazarı-Mençeler Valley (MEN).
Description. All samples are ex-situ and dimensions range from 6-7cm in height and 1-4 cm in diameter. One sample, KIR40, is very poorly preserved, and some features were not observed.
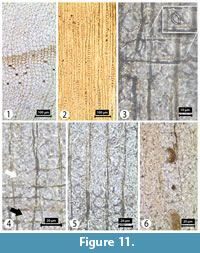 Transverse section (TS). Growth rings boundaries distinct with ring widths ranging from 0.4 to 1.2 mm; transition from earlywood to latewood almost abrupt, with 4-7 rows of latewood tracheids in KIR73 but gradual in KIR70. Resin canals absent (Figure 11). Tracheid tangential and radial diameters measure 22 (5-40) µm and 23 (10-45) µm, respectively, in the earlywood, and 14 (7-22) µm and 7 (1-15) µm, respectively, in the latewood. Tracheid wall thickness is 3 (1-5) µm (thin walled) in the earlywood and 5 (1-12) µm (thick walled) in the latewood. Axial parenchyma common, diffuse and in short tangential bands both through the transition zone and in late latewood (Figure 11).
Transverse section (TS). Growth rings boundaries distinct with ring widths ranging from 0.4 to 1.2 mm; transition from earlywood to latewood almost abrupt, with 4-7 rows of latewood tracheids in KIR73 but gradual in KIR70. Resin canals absent (Figure 11). Tracheid tangential and radial diameters measure 22 (5-40) µm and 23 (10-45) µm, respectively, in the earlywood, and 14 (7-22) µm and 7 (1-15) µm, respectively, in the latewood. Tracheid wall thickness is 3 (1-5) µm (thin walled) in the earlywood and 5 (1-12) µm (thick walled) in the latewood. Axial parenchyma common, diffuse and in short tangential bands both through the transition zone and in late latewood (Figure 11).
Tangential longitudinal section (TLS). Rays uniseriate, homogenous, very rarely biseriate, mostly short, 5-15 (2-21) cells in height or 95 (22-347) μm high and 10 (4-18) µm in width (Figure 11).
Radial longitudinal section (RLS). Tracheid pitting on radial walls, uniseriate with abietinean radial pitting (i.e., spaced and uniseriate; sensu Philippe and Bamford, 2008)--round, not contiguous, relatively large with diameters of 16-19 μm. Poor preservation has mostly rendered only the outer borders of tracheid pits recognizable. Generally, two (1-3) pits per cross-field, large cupressoid and taxodioid. Vertical and oblique apertures observed in some pits (Figure 11). Horizontal cross-field pits were not observed.
Comments. The combination of wood characters including the presence of distinct growth rings, uniseriate bordered pits on radial walls and 1-2 (up to 3) large cupressoid and taxodioid pits per cross field place this material within the genus Podocarpus (Brezinova et al., 1994). Since identification to species based on wood anatomy alone is often inconclusive, we refer these specimens to Podocarpus. Fossil woods of putative affinity to the Podocarpaceae have been described by many authors and assigned to Podocarpoxylon Gothan, Protopodocarpoxylon Eckhold, Phyllocladoxylon Gothan, Mesembrioxylon Seward, Metapodocarpoxylon Duperon-Laudoueneix and Pons, Protophyllocladoxylon Kräusel, Circoporoxylon Kräusel and even Cupressinoxylon Göeppert (Pujana and Ruiz, 2017). Pujana and Ruiz (2017) have recently reviewed fossil Podocarpoxylon woods and conclude that although many supposed woods of the podocarpaceous type have been found in the Northern Hemisphere, the deficit of other podocarpaceous fossil remains substantiates the fact that this the fossil genus may include wood from taxa other than those of Podocarpaceae. Possible podocarpaceous woods have previously been identified: Süss and Velitzelos (2000) identified two Podocarpoxylon wood species from the lower Miocene of Lesbos, and from Thrace (Sakınç et al., 2007) and Akkemik (work in progress) have both identified wood samples as Podocarpoxylon. This suggests a possible podocarpaceous tree grew across this area during the early Miocene. The sample described here is the first record of Podocarpus in the wood flora of GVP.
Family FAGACEAE Dumortier, 1829
Genus QUERCUS Linnaeus, 1753
Quercus section Ilex sp.
Material. INL09, KIR07, KAR01, MEN02, MEN22 and MEN28.
Localities. Within the Beypazarı-Inozu valley (north side) (INL) and the Beypazarı-Mençeler valley (MEN) and near to the villages of Beypazarı-Nuhhoca (Kıraluç Site) (KIR) and Beypazarı-Karaşar (KAR).
Description. All samples are in situ and range from 4 to 12 cm in length and 4 to 8.5 cm in diameter.
 Transverse section (TS). Growth ring boundaries indistinct, wood diffuse porous. Vessel tangential diameter 82 (22-182) μm, and radial diameter 109 (33-206) µm; average vessel density is 22 (13-29) vessels per mm2. Tyloses common in wider vessels. Large multiseriate rays can clearly be seen with the naked eye. In well-preserved woods axial parenchyma is clearly visible arranged in short tangential bands in single rows (Figure 12).
Transverse section (TS). Growth ring boundaries indistinct, wood diffuse porous. Vessel tangential diameter 82 (22-182) μm, and radial diameter 109 (33-206) µm; average vessel density is 22 (13-29) vessels per mm2. Tyloses common in wider vessels. Large multiseriate rays can clearly be seen with the naked eye. In well-preserved woods axial parenchyma is clearly visible arranged in short tangential bands in single rows (Figure 12).
Tangential longitudinal section (TLS). Rays number 4-6 per mm and are of two distinct sizes. Narrow uniseriate and biseriate rays measure 334 (110-805) µm by 14 (4-26) μm in width; large multiseriate rays range from 10 to 20 cells, or 258 (67-411) μm, in width by >1 mm in height (Figure 12).
Radial longitudinal section (RLS). Perforation plates are simple. Rays are homocellular and composed of procumbent ray cells (Figure 12).
Comments. The combination of characters outlined above indicate that this wood type belongs to Quercus L. and in particular the Quercus section Ilex wood type given that all these fossil specimens are diffuse porous (as opposed to ring porous, characteristic of Quercus Sections Quercus and Cerris). Furthermore, macrofossils of Quercus section Ilex (Denk et al., 2017a, 2017c) and pollen (Yavuz-Işık, 2008) have also been found from the Güvem region.
This wood type, which may represent a lineage ancestral to the modern western Eurasia sclerophyllous oaks, has also been found at the Hoçaş site (Figure 1) (Akkemik et al., 2016). Akkemik et al. (2016) concluded that the presence of evergreen oak species found at a total of five fossil sites in GVP suggested that this taxon was not an uncommon element of the early Miocene vegetation. Further studies on modern oaks and sediments around the Mediterranean Basin (Denk and Grimm, 2009, 2010; Hubert et al., 2014; Denk et al., 2017c) suggest a Eurasian group of evergreen oaks, comprising about 30 species, that occurred across the Mediterranean region and extended into the southern foothills of the Himalayas and across East and Southeast Asia during the early Miocene. The material found on the GVP may belong to this Eurasian group.
Family OLEACEAE Hoffmannsegg and Link, 1809
Genus FRAXINUS Linnaeus, 1753
Fraxinus sp.
Material. AGU13.
Locality. Near the village of Beypazarı-Aşağıgüney (AGU).
Description. The following description is based on one piece of in situ material measuring 16 cm in length and 8 cm in diameter.
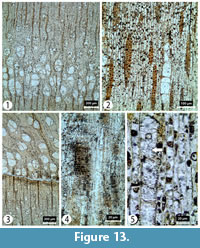 Transversal section (TS). Growth ring boundaries distinct, with ring widths ranging from 2.3 to 5 mm. Wood ring-porous. Earlywood vessels wide becoming narrower in the latewood. Tangential and radial diameters of vessel lumen 69 (26-116) μm and 90 (33-130) µm, respectively, in the earlywood, and in the latewood tangential and radial diameters measure 27 (13-46) 50 μm and 24 (11-52) µm, respectively. Vessel density 83 (67-104) vessels per mm2 with 26 (23-32) vessels per mm2 in earlywood and 57 (44-72) vessels in latewood. Vessels solitary and in radial multiples of up to three vessels. Paratracheal axial parenchyma almost lozenge-aliform especially in the latewood (Figure 13).
Transversal section (TS). Growth ring boundaries distinct, with ring widths ranging from 2.3 to 5 mm. Wood ring-porous. Earlywood vessels wide becoming narrower in the latewood. Tangential and radial diameters of vessel lumen 69 (26-116) μm and 90 (33-130) µm, respectively, in the earlywood, and in the latewood tangential and radial diameters measure 27 (13-46) 50 μm and 24 (11-52) µm, respectively. Vessel density 83 (67-104) vessels per mm2 with 26 (23-32) vessels per mm2 in earlywood and 57 (44-72) vessels in latewood. Vessels solitary and in radial multiples of up to three vessels. Paratracheal axial parenchyma almost lozenge-aliform especially in the latewood (Figure 13).
Tangential longitudinal section (TLS). Rays uniseriate to biseriate sometimes triseriate. Rays number ≥12 per mm (Figure 13) and measure 216 (58-345) µm in height and 23 (11-38) µm in width. Perforation plates simple. Fibres thin- to thick-walled (Figure 13).
Radial longitudinal section (RLS). Homocellular in some rays with all ray cells entirely procumbent others are heterocellular with procumbent body ray cells and square/upright marginals (Figure 13).
Comments. The combination of wood characters including ring porous vessels, distinct growth ring borders, clear transition from earlywood to latewood, sparse vessels in the latewood, rays 1-3 seriate with mostly procumbent body cells and slightly lozenge-aliform axial parenchyma shows closest anatomical similarity to Fraxinus L. (Akkemik and Yaman, 2012) and has thus been assigned to Fraxinus sp. Although Fraxinus is represented by four species, F. excelsior L., F. angustifolia Vahl., F. ornus L. and F. pallisae Wilmott. In Turkey today, this study represents the first record of Fraxinus wood material from the GVP (and from Turkey as a whole). Other macrofossil and palynological evidence including leaves (Kasaplıgil, 1977), fruits (Denk et al., 2017b) and pollen (Karayiğit et al., 1999) supports the presence of Fraxinus growing in the Güvem Basin at this time.
Family ROSACEAE Jussieu, 1789
Genus PRUNUS Linnaeus, 1753
Prunus sp.
Material. MEN31.
Locality. Beypazarı-Mençeler Valley (MEN).
Description. The following description is based on one piece of ex-situ material measuring 10 cm in length and 7 cm in width.
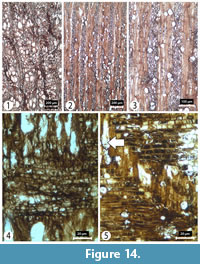 Transverse section (TS). Wood ring porous, sometimes more semi-ring porous, with distinct growth ring boundaries and ring widths ranging from 0.4 to 1 mm in width. Tangential and radial diameters of vessels measure 55 (32-75) µm and 82 (33-136) µm, respectively, in the earlywood, and 22 (8-43) µm and 30(15-50) µm, respectively, in latewood. Vessels solitary or in short radial files of 2-4. Vessel density 67 (55-78) per mm2 (Figure 14).
Transverse section (TS). Wood ring porous, sometimes more semi-ring porous, with distinct growth ring boundaries and ring widths ranging from 0.4 to 1 mm in width. Tangential and radial diameters of vessels measure 55 (32-75) µm and 82 (33-136) µm, respectively, in the earlywood, and 22 (8-43) µm and 30(15-50) µm, respectively, in latewood. Vessels solitary or in short radial files of 2-4. Vessel density 67 (55-78) per mm2 (Figure 14).
Tangential longitudinal section (TLS). Rays number 4-12 per mm and are clearly of two distinct sizes (Figure 14); wide rays 4-10 seriate, or 66 (51-78) µm in width, and >1 mm in height; narrow rays uniseriate (very rarely biseriate), or 7 (4-11) µm in width, and 117 (67-156) µm in height.
Radial longitudinal section (RLS). Rays heterocellular with procumbent body ray cells and one or more rows of upright and/or square marginal cells. Perforation plates simple. Intervessel pits are alternate or loosely opposite (Figure 14).
Comments. The combination of ring porous wood, relatively wide and long heterocellular rays and simple perforation plates suggest greatest affinity with Prunus. Today there are four species of Prunus growing in Turkey (Akkemik and Eminağaoğlu, 2014). Akkemik and Yaman (2012) described the wood anatomy of Prunus cocomilia Ten. from the Mediterranean region. The characters of this silicified wood exhibit strong anatomical similarity to the extant species but since species of Prunus are difficult to determine from wood anatomy alone this fossil type is assigned to Prunus sp. and represents the first record of fossilized Prunus wood from both the GVP and Turkey.
Family SALICACEAE Mirbel, 1815
Genus SALIX Linnaeus, 1753
Salix sp.
Material. KIR59, MEN30 and MEN34 measuring 5-7 cm in diameter and 9-13 cm high.
Localities. Near the village of Beypazarı-Nuhhoca (Kıraluç Site) (KIR) and in the Beypazarı-Mençeler Valley (MEN).
Description. This description is based on three pieces of in situ material measuring 9-13 cm in length by 5-7 cm in diameter.
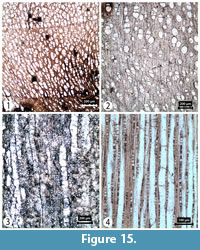 Transverse section (TS). Wood diffuse porous, sometimes semi-ring porous. Growth rings measure 1-11 mm wide with distinct or indistinct growth ring boundaries. Vessels solitary or in short radial files of 2-4, elliptic or angular when grouped. Vessel tangential and radial diameters measure 62 (20-165) µm and 75 (17-193) µm, respectively. Vessel density is 158 (81-252) per mm2 (Figure 15).
Transverse section (TS). Wood diffuse porous, sometimes semi-ring porous. Growth rings measure 1-11 mm wide with distinct or indistinct growth ring boundaries. Vessels solitary or in short radial files of 2-4, elliptic or angular when grouped. Vessel tangential and radial diameters measure 62 (20-165) µm and 75 (17-193) µm, respectively. Vessel density is 158 (81-252) per mm2 (Figure 15).
Tangential longitudinal section (TLS). Rays exclusively uniseriate, very rarely partly biseriate. Rays 272 (51-994) μm in height and 10 (4-23) μm in width, with ≥12 rays per mm (Figure 15).
Radial longitudinal section (RLS). Rays appear heterocellular with procumbent body ray cells and one to two upright and/or square marginal cells. Intervessel pits are alternate and loosely opposite. Perforation plates simple (Figure 15).
Comments. Salix L. wood can often be confused with that of Populus L., however, the clearly heterocellular rays in this silicified wood identifies this specimen as Salix and has therefore been assigned to Salix sp. Today Salix L. is represented by 33 species in Turkey favouring riparian environments.
Both Salix L. and Populus L. leaves and pollen have been described from GVP (Kasaplıgil, 1977; Denk et al., 2017b) and Salix / Populus wood has been identified from the Hoçaş site (Akkemik et al., 2016). These genera were also common in the Miocene across western Anatolia (Güner, 2016), and support the evidence for the presence of a localized riparian type vegetation occupying lacustrine environments (Toprak et al., 1996 and references cited therein) on the GVP.
Family SAPINDACEAE Jussieu, 1789
Genus ACER Linnaeus, 1753
Acer sp.
Material. INL02, INL06, KIR02, KIR13, KIR51, KIR52 and MEN29.
Localities. Valleys of Beypazarı-Inozu (north side) (INL) and Beypazarı-Mençeler (MEN), and near the village of Beypazarı-Nuhhoca (Kıraluç Site) (KIR).
Description. The following description is based on all seven pieces of ex-situ petrified wood samples measuring 6.5-10 cm in length by 4-9.5 cm in diameter although the preservation in the samples collected from Beypazarı-Nuhhoca Village (Kıraluç Site) (KIR) is poor.
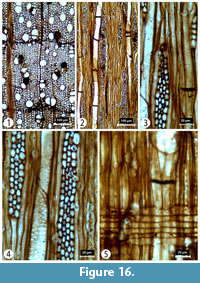 Transverse section (TS). Wood diffuse-porous. Ring widths 0.4-1.2 mm. Growth ring boundaries distinct and marked by 1-2 rows of marginal axial parenchyma cells. Tangential and radial diameters of vessels measure 55 (11-158) µm and 66 (14-181) µm, respectively. Vessels circular and predominantly solitary with radial multiples of 2-4 or more. Vessel density is 206 (92-343) per mm2. Axial parenchyma is both apotracheal diffuse and marginal. In some of the vessels dark deposits can clearly be seen. Fibers are generally thin walled (Figure 16).
Transverse section (TS). Wood diffuse-porous. Ring widths 0.4-1.2 mm. Growth ring boundaries distinct and marked by 1-2 rows of marginal axial parenchyma cells. Tangential and radial diameters of vessels measure 55 (11-158) µm and 66 (14-181) µm, respectively. Vessels circular and predominantly solitary with radial multiples of 2-4 or more. Vessel density is 206 (92-343) per mm2. Axial parenchyma is both apotracheal diffuse and marginal. In some of the vessels dark deposits can clearly be seen. Fibers are generally thin walled (Figure 16).
Tangential longitudinal section (TLS). Rays are of two distinct sizes: generally, uniseriate, sometimes biseriate, or multiseriate commonly 4-5 seriate. Rays measure 220 (18-656) μm in height by 25 (4-78) µm in width. Intervessel pitting alternate. Helical thickening commonly present on the walls of vessels (Figure 16).
Radial longitudinal section (RLS). Rays are homocellular and composed entirely of procumbent cells. Helical thickenings can also be seen in radial walls of the wood. Intervessel pitting alternate. Perforation plates simple (Figure 16).
Comments. This combination of wood features suggests that these samples belong to Acer. This genus is relatively common in the Miocene of Anatolia and is represented at five locations across the GVP with fossil wood having being recovered at Hoçaş and Kozyaka (Akkemik et al., 2016). Further micro- and macrofossils of Acer L. have been recovered from Güvem (Kasaplıgil, 1977; Denk et al., 2017b).
Today Acer is represented by 11 species in Turkey with Acer monspessulanum L. being one of the species adapted to the typically xeric conditions of the Mediterranean (Akkemik and Yaman, 2012). Since the wood features of Acer L. species in Turkey are very similar (Yaltırık, 1971) and cannot be distinguished by wood anatomy alone, this material is referred to Acer sp.
Family ULMACEAE Mirbel, 1815
Genus ULMUS Linnaeus, 1753
Ulmus sp.
Material. KIR14 and KIR65.
Localities. Beypazarı-Nuhhoca Village (Kıraluç Site) (KIR)
Description. These in situ samples range from 7-9 cm in length by 8-11 cm in diameter.
 Transverse section (TS). Wood ring porous; growth ring boundaries distinct; ring width 1-2 mm. Earlywood vessels are arranged in 2-3 rows, solitary or in radial multiples; latewood vessels arranged in tangential bands. Vessel tangential and radial diameters measure 70 (29-117) μm and 100 (50-169) μm, respectively, in the earlywood, and 21 (4-37) μm and 35 (20-46) μm, respectively, in the latewood. Vessel density is 11 (8-14) per mm2 in the earlywood and 180 (132-244) per mm2 in the latewood, overall vessel density is 191 (140-258) per mm2. Axial parenchyma scanty paratracheal and diffuse (Figure 17).
Transverse section (TS). Wood ring porous; growth ring boundaries distinct; ring width 1-2 mm. Earlywood vessels are arranged in 2-3 rows, solitary or in radial multiples; latewood vessels arranged in tangential bands. Vessel tangential and radial diameters measure 70 (29-117) μm and 100 (50-169) μm, respectively, in the earlywood, and 21 (4-37) μm and 35 (20-46) μm, respectively, in the latewood. Vessel density is 11 (8-14) per mm2 in the earlywood and 180 (132-244) per mm2 in the latewood, overall vessel density is 191 (140-258) per mm2. Axial parenchyma scanty paratracheal and diffuse (Figure 17).
Tangential longitudinal section (TLS). Rays 3-4 (up to 6) seriate measuring 34 (13-63) μm in width and 259 (120-445) μm in height (Figure 17).
Radial longitudinal section (RLS). All rays homocellular composed entirely of procumbent ray cells. Intervessel pitting alternate. Perforation plates simple (Figure 17).
Comments. The character combination of these two pieces of petrified woods (i.e., ring porous earlywood vessels, latewood vessels arranged in tangential continuous bands, rays 3-4 (up to 6) seriate, simple perforation plates and homocellular rays) identify this wood as Ulmus L.
Today four species of Ulmus are native to Turkey: Ulmus canescens Melville is adapted to xeric conditions within the Mediterranean Basin, the other three species (U. minor Mill., U. glabra Huds. and U. laevis Pall.) grow on more humid sites. However, regardless of their ecological preference, the wood anatomy of Ulmus species native to Turkey are all very similar, and it is not possible to separate the species on wood anatomy alone (Akkemik, 1995). Since the wood characters exhibited by the petrified woods studied here are very similar to all four extant species, we refer the fossil material to Ulmus sp.
Other micro and macrofossils of Ulmus have been identified from the Miocene of Turkey, namely woods from the Hoçaş site and the Kıraluç site to the northwest of Beypazarı-Nuhhoca Village (Akkemik et al., 2016), and leaves and pollen from both west Anatolia (Akgün et al., 2007; Akkiraz et al., 2011; Güner, 2016) and at Güvem (Kasaplıgil, 1977; Denk et al., 2017b). This record, therefore, extends the record of Ulmus L. in the local vegetation in this part of Turkey.
Genus ZELKOVA Spach, 1841
Zelkova sp.
Material. AGU15, AGU28 and KIR01.
Localities. Near the villages of Beypazarı-Aşağıgüney (AGU) and Beypazarı-Nuhhoca (Kıraluç Site) (KIR).
Description. The following description is based on three pieces of in situ wood measuring 9 cm in length by 6 cm in diameter.
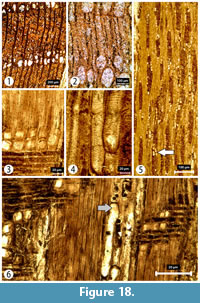 Transversal section (TS). Growth rings 0.5-2.5 mm in width, ring boundaries distinct with transition from earlywood to latewood abrupt. Wood ring porous (Figure 18). Earlywood vessels in 1-2 rows; vessel clusters common in late wood. Vessel tangential and radial diameters measure 76 (23-142) µm and 118 (55-182) µm, respectively, in the earlywood and 17 (6-36) µm and 19 (6-42) µm, respectively, in the latewood. Vessel density 325 (258-399) per mm2. Axial parenchyma unclear.
Transversal section (TS). Growth rings 0.5-2.5 mm in width, ring boundaries distinct with transition from earlywood to latewood abrupt. Wood ring porous (Figure 18). Earlywood vessels in 1-2 rows; vessel clusters common in late wood. Vessel tangential and radial diameters measure 76 (23-142) µm and 118 (55-182) µm, respectively, in the earlywood and 17 (6-36) µm and 19 (6-42) µm, respectively, in the latewood. Vessel density 325 (258-399) per mm2. Axial parenchyma unclear.
Tangential longitudinal section (TLS). Rays 1-6 seriate. Wide rays (3-6 seriate) generally 25-35 cells, or 379 (80-854) μm, with a maximum height of 51 cells; commonly >4 seriate or 41 (22-68) μm; narrow rays (1-2 seriate) 14 (6-28) µm in width and generally 11-23 cells or 213 (63-482) µm in height. Vascular/vasicentric tracheids with helical thickening present (Figure 18).
Radial longitudinal section (RLS). Rays homocellular with entirely procumbent ray cells and heterocellular with mostly procumbent body ray cells and one row of upright and/or square marginal cells (Figure 18). Axial parenchyma in strands of 2-4 cells. Prismatic crystals present in enlarged axial parenchyma cells and in the upright cells of rays. Perforation plates simple.
Comments. The wood of Zelkova Spach. is very similar to that of Ulmus L. but these taxa can be distinguished by the presence of heterocellular rays with one or two rows of upright cells together with procumbent body ray cells and prismatic crystals in the axial parenchyma. These characters can be seen in the fossil wood described here and thus we have assigned this material to Zelkova sp.
The wood identified here is the first record of Zelkova in the fossil wood record of Turkey, although leaf fossils have been identified from both Güvem (Kasaplıgil, 1977; Denk et al., 2017b) and west Anatolia (Güner, 2016; Bouchal et al., 2016, 2017), and pollen has been identified from west Anatolia (Üçbaş Durak and Akkiraz, 2016) and central Anatolia (Akkiraz et al., 2011). Today Zelkova is represented by one species, Zelkova carpinifolia (Pall.) K. Koch., in the flora of Turkey.
DISCUSSION
Forest Associations Across the Galatian Volcanic Province
During the early Miocene the climate in western Eurasia was mild and equable, (Rögl, 1999) supporting the growth of rich forest vegetation in Northern Hemisphere as far north as the subarctic (Mai, 1995; Zachos et al., 2008; Denk et al., 2011, 2017a, 2017b, 2017c). Anatolia, along with the Lower Caucasus to the east, formed an island to the north of the eastern Mediterranean and to the south of the Eastern Paratethys (Popov et al., 2004). The Western Pontides Mountains to the north and the Taurus Mountains to the south surrounded the freshwater lakes and marshes that covered much of the Central Anatolian Plateau (Popov et al., 2004; Schildgen et al 2012a, 2012b). The GVP forms a region in the north-western part of this Central Anatolian Plateau. According to Tankut et al. (1998), the major phase of volcanic activity on the GVP at this time resulted in the deposition of the plant-bearing strata.
Modern analogue techniques (e.g., Utescher et al., 2007; Mustoe and Leopold, 2014) use the composition of floral assemblages present today and compare them with Cenozoic fossil assemblages to provide ecological data of the palaeoenvironments from which the fossils were recovered. Although we do not feel that our palaeoassemblages are complete enough to warrant undertaking such tests just yet, this assemblage approach can provide broad ideas pertaining to the environments in which the trees grew, especially given that these are geologically relatively young assemblages.
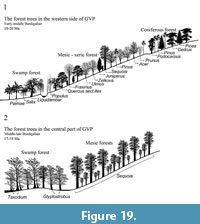 The current hypothesis that states the landscape across the GVP were relatively heterogeneous (Denk et al., 2017b) is supported by the results of the local (wood) flora at discrete sites across the GVP found in this study. The 10 sites on the western part of the GVP lie at an altitude of 1000-1400 m whereas those in the central part of the GVP lie at 1000-1250 m (Table 1). Since there is no evidence for any significant change in elevation as a result of tectonic processes since the early Miocene, we cannot use altitudinal differences to explain the different vegetation assemblages. Given that the sites located in the western part of the GVP are geographically distinct and possibly slightly younger than those in the western part (Figure 19 and Table 2), we will, however, consider these two regions separately in the context of previously published work.
The current hypothesis that states the landscape across the GVP were relatively heterogeneous (Denk et al., 2017b) is supported by the results of the local (wood) flora at discrete sites across the GVP found in this study. The 10 sites on the western part of the GVP lie at an altitude of 1000-1400 m whereas those in the central part of the GVP lie at 1000-1250 m (Table 1). Since there is no evidence for any significant change in elevation as a result of tectonic processes since the early Miocene, we cannot use altitudinal differences to explain the different vegetation assemblages. Given that the sites located in the western part of the GVP are geographically distinct and possibly slightly younger than those in the western part (Figure 19 and Table 2), we will, however, consider these two regions separately in the context of previously published work.
Forests of the central GVP. Güvem, located in the central part of the GVP, is one of the few precisely dated early Miocene assemblages of plant and animals. The age of this material is 20-18 Ma, or early Burdigalian, and many taxa are more typical of temperate and equatorial monsoon climates of SE Asia (van den Hoek Ostende, 2001). Denk et al. (2017b) undertook a major review of the plant material from this area using evidence from leaves, fruits and flower remains and published data of dispersed pollen and spores (Denk et al., 2017a, 2017c and reference cited therein). A total of 38 of the 40 known taxa identified from the GVP to date have been recorded from Güvem (Table 3). Denk et al. (2017b and reference cited therein) conclude that the landscape was varied and characterized by a mix of trees, shrubs, lianas, ferns and fern allies. Evergreen broadleaved forests comprising Ilex, Quercus and other Fagaceae types dominated the areas surrounding lowland lakes and across large parts of both lowland and uplands (Denk et al., 2017b). Restricted open areas probably supported Fabaceae and Poaceae (Yavuz-Isik, 2008; Denk et al., 2017b). Wetland biomes comprising swamp forest communities and wet meadows surrounded the lakes and rivers where taxodiaceous swamp elements (Glyptostrobus) and palms flourished (Kasapligil, 1977; Denk et al., 2017b). According to Schroeder (1998) and Denk et al. (2017b), the heterogeneous nature of the landscape is further reflected in well-drained lowland broadleaved evergreen forests, mesic mixed deciduous and coniferous upland forests with Sequoia, Fagus types and conifers (Cedrus and Cathaya Chub and Kuang), and the presence of laurel forests. In the pollen flora, a more widespread vegetation assemblage is usually recorded. Yavuz-Işık and Demirci (2009) identified pollen of Pinaceae, Ulmus, Salix, Quercus, Carpinus L., Carya Nutt. and Engelhardia Lesch. ex Blume, which are similar to our findings regarding the composition of the vegetation in the western part of GVP (see below). This suggests that floral associations did indeed vary geographically across this region at this time. These authors also conclude that across the Güvem region the climate was humid and warm temperate, with a somewhat drier season in the winter. This agrees with other studies that suggest the climate in Güvem during the early to mid-Miocene was warm-temperate with a mean annual temperature for the region of 17.2-20.8ºC (Benda and Muelenkamp, 1990; Akgün et al., 2007; Kayseri and Akgün, 2008; Yavuz-Işık, 2008).
At other sites across the central GVP region, research undertaken on the wood flora has been used to further knowledge of the prevailing local vegetation. In the central region of the GVP to the west of Güvem near Pelitcik, the Camlidere Fossil Forest, dated to 18.2-16.9 Ma, (Tankut et al., 1995) is thought to have been composed exclusively of ‘Taxodium’ and ‘Sequoia’ (Akkemik et al., 2009); 80 km to the southeast of Güvem, however, the coeval Güdül Fossil Forest comprised a riparian/wetland environment dominated by a ‘Glyptostrobus’ forest (Akkemik et al., 2017). Therefore, the wood flora adds support to the idea of local wet biome floral associations postulated by Kasapligil (1977) and later Denk et al. (2017b).
Such swamp forest elements were once widespread across the Northern Hemisphere. Today they are no longer found in Europe or the Mediterranean, but are restricted to the south-eastern USA and parts of East Asia (Mustoe and Leopold, 2014). Taxodium flourishes in regions characterized by poorly drained environments, mild winters, year round rainfall and mean annual precipitation levels of >1000 mm/yr (Hoagland et al., 1996). Today Sequoia is restricted to a 725 km strip along the Pacific Coast of North America in moist sheltered habitats where the climate is mild and frost free, and moisture levels remain relatively high all year (Eckenwalder, 2009). Glyptostrobus grows in pure stands across parts of Asia along swamp margins, on river floodplains and in deltas; the buttress base and pneumatophores evidence its adaptation to seasonal inundation (Eckenwalder, 2009). This association, as also suggested by the local wood flora in the central part of the GVP and the presence of palms (at Güvem), supports the hypothesis that the climate was warm temperate, mild and frost free, during the Burdigalian, and wet habitats existed across the region. Cooling of the climate during the later Miocene (Benda and Muelenkamp, 1990; Kayseri and Akgün, 2008; Yavuz-Işık, 2008) probably caused this swamp-forest association to ultimately disappear from the GVP.
Forests of western GVP. Fossil evidence for the vegetation across the western part of the GVP during the Burdigalian is more limited relative to the central region (Figure 19 and Table 2). Here fossil wood floras of Aquitanian-Burdigalian age are currently the main source of evidence and provide insights into the local, rather than a more regional, vegetation composition. Material from two sites in the very western edge of the GVP have already been studied: The Hoçaş Fossil Forest and the Kozyaka Fossil Forest close to the town of Seben. Wood from the Hoçaş site, dated to between 20 and 18 Ma (Burdigalian) (Akgün et al., 2002; Kayseri and Akgün, 2008; Yavuz-Işık, 2008; Akkemik et al., 2016), comprises palms, conifers (Cedrus, Picea, Juniperus,) and angiosperms (Salix/Populus, Acer, Quercus section Ilex and Liquidambar) and represents the most diverse local flora in this region with nine of a possible 40 taxa identified from the GVP. The Kozyaka Fossil Forest (considered to be coeval with the Hocas Fossil Forest; Akkemik et al., 2016) comprises a similar mixed flora of six taxa, namely the conifers Cedrus and Pinus, and the angiosperms Salix/Populus, Acer, evergreen Quercus and Ulmus.
The results of this study also suggest that relative to the central GVP the local flora in the western part of the GVP is more diverse (Table 3). Quercus section Ilex is the most geographically widespread taxon found at six of the nine sites followed closely by Salix/Populus and Acer found at five of the sites; Picea and Cedrus have been found at four; Zelkova, Ulmus, Juniperus, Podocarpus at two sites; Prunus, Liquidambar, Fraxinus, Sequoia and palms each only on one site.
The localities with the most diverse local floras are: The Kıraluç Site at Beypazarı-Nuhhoca Village (KAR) with seven taxa (Pinus, Podocarpus, Acer, Quercus section Ilex, Salix/Populus, Ulmus and Zelkova), Beypazarı-Aşağıgüney (AGU) with six taxa (Juniperus, Picea, Pinus, Fraxinus, Salix/Populus and Zelkova) and Beypazarı-Mençeler (MEN) with five taxa (Podocarpus, Acer, Prunus, Quercus section Ilex and Salix/Populus). The remaining localities have relatively few taxa identified to date: Beypazarı-Inozu valley (north side) (INL) had Cedrus, Acer and Quercus section Ilex ; Beypazarı Inozu valley (south side) (INO) had Cedrus and Sequoia ; Beypazarı-Karaşar Village (KAR) and Bolu-Kıbrısçık-Kuzca-Alanhümmet villages (KUZ) with only one taxon-- Quercus section Ilex and Pinus respectively (Figure 19, Table 2 and Table 3).
Once again, modern analogue techniques (Utescher et al., 2007; Mustoe and Leopold, 2014) can provide useful information regarding the paleoecology of the western part of the GVP during the Aquitanian-Burdigalian. From comparisons of the assemblages of wood material identified at the sites in the western part of the GVP with the extant flora that grows along the southwestern coasts of Turkey today, we suggest a possible three different forest types may have grown in this region.
A ‘riparian forest’ association. Today this riparian association is fairly widespread along water courses and includes elements such as Salix, Liquidambar, Alnus, Fraxinus, Ulmus, Platanus, Myrtus, growing together with palms (Phoenix theophrastii Greuter), Juniperus and Pinus (Efe, 1987). Since this association is restricted to water courses it is relatively intolerant of local conditions and can grow in wet, semi-dry and dry environments. This taxa combination has been identified from the HOC site along with Acer, Picea and Cedrus suggesting that the fossil ‘riparian forest’ flora may have been more taxonomically diverse than its modern counterpart.
A ‘mesic-xeric’ woodland association. Today in the Mediterranean region of Turkey, under a warm temperate climate with mean annual rainfall of c. 500-1000 mm/yr, a semi-dry mesic-xeric macchie-type association (Özalp, 2000) is commonly found growing around the extant riparian forest belt at relatively low altitudes (200-1000 m). For example, in south-western Turkey, species of Acer grow alongside several species of evergreen Quercus evergreen oaks (Quercus ilex L., Q. coccifera L. and Q. aucheri Jaub. and Spach) and deciduous oak species in xeric habitats of the Mediterranean region with Pinus and Juniperus (Yaltırık, 1971; Günal, 2011). During the early Miocene, evidence for Acer and Quercus can be found together in five of the nine sites in this part of the GVP (Table 3), sometimes with Pinus and one or more other species not found in the modern analogue. With more studies of these floras, the composition of this vegetational association and the degree of similarity to modern mesic-xeric woodlands will become clearer.
Upland mixed coniferous-angiosperm forest association. Today a conifer association comprising Pinus, Cedrus, Juniperus and Abies can be found at higher altitudes (1000-1800 m a.s.l.) (Yaltırık and Akkemik, 2012). These taxa, except Abies, are present in the fossil assemblages (e.g., KOZ, HOC, KUZ) and might suggest a more upland forest association of this type grew, together with Picea, Podocarpus, Cedrus, during the early Miocene. These conifer elements probably occurred alongside angiosperms (e.g., Acer, Quercus, Salix/Populus, Ulmus, and possibly Fraxinus and Zelkova). A link to this extant vegetation is certainly more tenuous at this stage, but the possibility does exist that drier habitats of the surrounding higher ground could have supported a more mixed conifer vegetation. Further work is needed to understand more fully the associations these taxa may have had across the GVP at this time.
The presence of Sequoia at the INO site probably reflects a habitat distinct from this association and more similar to the swamp forests in the central part of the GVP. However, the presence of Cedrus, more commonly found at higher altitudes today, alongside Sequoia may suggest a more diverse habitat and/or vegetation during the Miocene.
Taxodium and Podocarpus are not represented in the floristically diverse assemblage found at Güvem and constitute new records for the GVP. From nearest living relative studies Taxodium would also support a warm temperate environment with humid/damp growing conditions.
Paleoclimate studies undertaken by various workers suggest that a subtropical to warm-temperate climate prevailed over large parts of western Turkey (Güner, 2016), north-western central Anatolia and into Greece during the early Miocene (Velitzelos et al., 2014). The climate was characterized by mean annual temperature of 17.2-21.3ºC, a cold month mean of 6.2-13.3ºC, a warm month mean of 26.5-27.9℃ and a mean annual precipitation of c. 1000-1325 mm/yr and a driest month precipitation level between 40-60 mm (Akgün et al., 2007; Akkiraz et al., 2011). Although the wood associations currently do not provide us with absolute levels of temperature and precipitation at this stage and simply suggest a prevailing warm temperate climate, our findings are in support of the conclusions of Denk et al. (2017a, 2017b, 2017c) and other workers cited above.
CONCLUSION
The data from fossil wood found at 10 new fossil sites on the GVP (three localities in the central part and seven in the western part) provide a more detailed view of the vegetation associations that grew across the region during the early Miocene. The wood flora suggested evidence of taxonomically distinct local associations. The results provided further support for the current hypotheses for a warm temperate climate prevailing over a heterogeneous landscape comprising a riverine association, a mesic-xeric woodland association and possibly a mixed coniferous forest in the western part of the GVP and swamp forests associations located towards the central part of the GVP. Although this study brings the total of petrified wood genera to 15, studies of other macrofossils and pollen data suggest many more genera were present across the wider area of the GVP. More work is, therefore, needed before we can fully understand the degree of variation in the vegetation across the GVP.
ACKNOWLEDGMENTS
This study was supported by the Research Fund of Istanbul University Project Number: YÖP-41738 and 22800.
REFERENCES
Adıyaman, Ö., Chorowicz, J., Arnaud, O.C., Gündoğdu, M.N., and Gourgaud, A. 2001. Late Cenozoic tectonics and volcanism along the North Anatolian Fault: new structural and geochemical data. Tectonophysics, 338:135-165. https://doi.org/10.1016/S0040-1951(01)00131-7
Akbaş, B., Sevin, M., and Aksay, A. 2002. [1/100.000 Scale Geological Map of Turkey No. 40 Bolu - H27 Sections of MTA Geological Survey Department]. MTA Geological Survey Department, Ankara, Turkey. [In Turkish]
Akgün, F., Akay, E., and Erdoğan, B. 2002. Tertiary terrestrial to shallow marine deposition in central Anatolia: a palynological approach. Turkish Journal of Earth Sciences, 11:127-160.
Akgün, F., Kayseri, M.S., and Akkiraz, M.S. 2007. Paleoclimatic evolution and vegetational changes during the Late Oligocene-Miocene period in western and central Anatolia (Turkey). Palaeogeography, Palaeoclimatology, Palaeoecology, 253:56-90. https://doi.org/10.1016/j.palaeo.2007.03.034
Akkemik, Ü. 1995. The morphological characteristics of native elm (Ulmus L.) taxa. Review of the Faculty of Forestry, University of Istanbul, 45(A-2):93-115.
Akkemik, Ü., Acarca N.N., and Hatipoğlu M. 2017. The first Glyptostroboxylon from the Miocene of Turkey. IAWA Journal, 38(4):561-570. https://doi.org/10.1163/22941932-20170181
Akkemik, Ü., Arslan, M., Poole, I., Tosun, S., Köse, N., Karlıoğlu Kılıç, N., and Aydın, A. 2016. Silicified woods from two previously undescribed early Miocene forest sites near Seben, northwest Turkey. Review of Palaeobotany and Palynology, 235:31-50. https://doi.org/10.1016/j.revpalbo.2016.09.012
Akkemik, Ü. and Eminağaoğlu, Ö. 2014. [ Prunus L.], p. 369-373. In Akkemik, Ü. (ed.), [Trees and Shrubs of Turkey]. OGM Publications, Ankara. [In Turkish]
Akkemik, Ü., Köse, N., and Poole, I. 2005. Sequoioiodae (Cupressaceae) woods from the upper Oligocene of European Turkey (Thrace). Phytologia Balcanica, 11(2):119-131.
Akkemik, Ü. and Sakınç, M. 2013. Sequoioxylon petrified woods from the Mid to Late Oligocene of Thrace (Turkey). IAWA Journal, 34(2):177-182. https://doi.org/10.1163/22941932-00000015
Akkemik, Ü, Türkoğlu, N., Poole, I., Çiçek, İ., Köse, N., and Gürgen, G. 2009. Woods of a Miocene petrified forest near Ankara, Turkey. Turkish Journal of Agriculture and Forestry, 33:89-97.
Akkemik, Ü. and Yaman, B. 2012. Wood Anatomy of Eastern Mediterranean Species. Kessel Publishing House, Remagen, Germany.
Akkiraz, M.S., Akgün, F., Utescher, T., Bruch, A.A., and Mosbrugger, V. 2011. Precipitation gradients during the Miocene in Western and Central Turkey as quantified from pollen data. Palaeogeography, Palaeoclimatology, Palaeoecology, 304:276-290. https://doi.org/10.1016/j.palaeo.2010.05.002
Altun, İ.E., Kadınkız, G., and Aksay, A. 2002. [ 1/100.000 Scale Geological Map of Turkey No. 41 Bolu - H28 Sections of MTA Geological Survey Department ]. MTA Geological Survey Department Ankara, Turkey. [In Turkish]
Barefoot A.C. and Hankins F.W. 1982. Identification of Modern and Tertiary Woods. Clarendon Press, Oxford.
Bartling, F.T. 1830. Ordines Naturales Plantarum, eorumque Characteres et Affinitates. Sumtibus Dieterichianis, Göttingen.
Benda, L. and Muelenkamp, J.E. 1990. Biostratigraphic correlations in the Eastern Mediterranean Neogene, 9. Sporomorph associations and event stratigraphy of the Eastern Mediterranean, Newsletters on Stratigraphy, 23(1):1-10.
Biondi, E. and Brugiapaglia, E. 1991. Taxodioxylon gypsceum in the fossil forest of Dunarobba (Umbria, Central Italy. Flora Mediterranea, 1:111-120.
Bouchal, J.M., Grímsson, F., Zetter, R., and Denk, T. 2016. The middle Miocene palynoflora and palaeoenvironments of Eskihisar (Yatağan Basin, southwestern Anatolia): A combined LM and SEM investigation. Botanical Journal of the Linnean Society, 182:14-79. https://doi.org/10.1111/boj.12446
Bouchal, J.M., Mayda, S., Akgün, F., Grímsson, F., Zetter, R., and Denk, T. 2017. Miocene palynofloras of the Tınaz lignite mine, Muğla, southwest Anatolia: Taxonomy, palaeoecology and local vegetation change. Review of Palaeobotany and Palynology, 243:1-36. https://doi.org/10.1016/j.revpalbo.2017.02.010
Brezinova, D., Holy, F., Kuzvartova, A., and Kvacek, Z. 1994. A silicified stem of Podocarpoxylon helmstedtianum Gottwald, 1966 from the Palaeogene site Kuclin (NW Bohemia). Journal of the Czech Geological Society, 39(2-3):221-234.
Denk, T. and Grimm, G.W. 2009. Significance of pollen characteristics for infrageneric classification and phylogeny in Quercus (Fagaceae). International Journal of Plant Sciences, 170:926-940. https://doi.org/10.1086/600134
Denk, T. and Grimm, G.W. 2010. The oaks of western Eurasia: traditional classifications and evidence from two nuclear markers. Taxon, 59:351-366
Denk, T., Grimm, G.W., Manos, P.S., Deng, M., and Hipp, A. 2017a. An updated infrageneric classification of the oaks: review of previous taxonomic schemes and synthesis of evolutionary patterns, p. 13-38. In Gil-Peregrin, E., Peguero-Pina, J.J., and Sancho-Knapik, D. (eds.), Oaks Physiological Ecology. Exploring the Functional Diversity of Genus Quercus. Tree Physiology 7. Springer Nature, Cham, Switzerland.
Denk, T., Grímsson, F., Zetter, R., and Símonarson, L.A. 2011. A brief review of palaeobotanical research in Iceland, p. 31-43. In Landman, N.H. and Harries, P. (eds.), Late Cainozoic Floras of Iceland. 15 Million Years of Vegetation and Climate History in the Northern North Atlantic. Topics in Geobiology 35. Springer, Dordrecht.
Denk, T., Güner, T.H., Kvaček, Z., and Bouchal, M.J. 2017b. The early Miocene flora of Güvem (Central Anatolia, Turkey): A window into early Neogene vegetation and environments in the Eastern Mediterranean. Acta Palaeobotanica (monograph), 57:237-338. https://doi.org/10.1515/acpa-2017-0011
Denk, T., Velitzelos, D., Güner, T., Bouchal, J.M., Grimsson, F., and Grimm, G.W. 2017c. Taxonomy and palaeoecology of two widespread western Eurasian Neogene sclerophyllous oak species: Quercus drymeja Unger and Q. mediterranea Unger. Review of Palaeobotany and Palynology, 241:98-128. https://doi.org/10.1016/j.revpalbo.2017.01.005
Dernbach, U., Herbst, R., Jung, W., Selmeier, A., Schaarschmidt, F., and Velitzelos, E. 1996. Ulrich Dernbach's Petrified Forests: The World's Most Beautiful Petrified Forests. D'oro Verlag Publishing, Heppenheim.
Dietrich, A. 1824. Flora der Gegend um Berlin. G.C. Nauck Publishing, Berlin.
Dumortier, B.C.J. 1829. Analyse des familles des plantes, avec l'indication des principaux genres qui s'y rattachent. Tournay. Imprimerie de J. Casterman, Aine.
Eckenwalder, J.E. 2009. Conifers of the World. Timber Press, Portland.
Efe, A. 1987. Morphological and palynological studies on Liquidambar orientalis Mill. in Turkey. Review of Faculty of Forestry, University of Istanbul, 37 (A-2): 84-114. [In Turkish with English abstract]
Eliçin, G. 1977. [ Researches on Anatomical Features and Distributions of Native Juniper Taxa in Turkey. Istanbul University, Forestry Faculty Publications, No. 2327/232 ]. Istanbul University, Istanbul. [In Turkish]
Endlicher, S. 1847. Synopsis Coniferarum. Sangalli, Aput Scheitlin & Zollikofer, Sangalli.
Fahn, A., Werker, E., and Baas, P. 1986. Wood Anatomy and Identification of Trees and Shrubs from Israel and Adjacent Regions. The Israel Academy of Science and Humanities, Jerusalem.
Greguss, P. 1955. Xylotomische Bestimmung der Heute Lebenden Gymnospermen, Academia Kiado, Budapest.
Greguss, P. 1967. Fossil Gymnosperm Woods in Hungary, from Permian to Pliocene. Academia Kiado, Budapest.
Günal, N. 2011. [Holm oak (Quercus ilex) 's geographical distribution in Turkey, and its ecological and floristic features. Physical geography surveys: Systematic and regional]. Turkish Geography Institute, 6:267-278. [In Turkish]
Güner, T. 2016. Yatağan Basin (Western Anatolia) Miocene Flora and Vegetation. Unpublished PhD Thesis, Istanbul University Institute of Science, Istanbul, Turkey. [In Turkish]
Hoagland, B.W., Sorrels, L.R., and Glenn, S.M. 1996. Woody species composition of floodplain forests of the Little River, McCurtain and LeFlore Counties, Oklahoma. Proceedings of Oklahoma Academy of Science, 76:23-29.
Hoffmannsegg, J.C. and Link, H.F. 1809. Flore Portugaise, Vol. 1. Charles Fréderic Amelang, Berlin.
Hubert, F., Grimm, G.W., Jousselin, E., Berry, V., Franc, A., and Kremer, A. 2014. Multiple nuclear genes stabilize the phylogenetic backbone of the genus Quercus. Systematics and Biodiversity, 12:405-423. https://doi.org/10.1080/14772000.2014.941037
IAWA Committee. 1989. IAWA List of microscopic features for hardwood identification by an IAWA Committee. International Association of Wood Anatomists Journal, 10:219-332.
IAWA Committee. 2004. IAWA list of microscopic features for softwood identification. International Association of Wood Anatomists Journal, 25:1-70. https://doi.org/10.1163/22941932-90000349
Jacquiot, C. 1955. Atlas d’anatomie des Bois des Coniferes, Press of Centre Technique du bois et du Fonds Forestier National, Paris.
Jussieu, A.L. 1789. Genera Plantarum. Historiae Naturalis Classica, Tomus XXXV. Wheldon and Wesley, Ltd., New York.
Karayiğit, A.I., Akgün, F., Gayer, R.A., and Temel, A. 1999. Quality, palynology, and palaeoenvironmental interpretation of the Ilgın lignite, Turkey. International Journal of Coal Geology, 38:219-236. https://doi.org/10.1016/s0166-5162(98)00015-9
Kasaplıgil, B. 1977. [A late Tertiary conifer-hardwood forest from the vicinity of Güvem village, near Kızılcahaman, Ankara]. Journal of MTA, 88:94-102. [In Turkish]
Kayseri, M.S. and Akgün, F. 2008. Palynostratigraphic, palaeovegetational and palaeoclimatic investigations on the Miocene deposits in Central Anatolia (Çorum Region and Sivas Basin), Turkish Journal of Earth Sciences, 17:361-403.
Kazancı, N. 2012. Geological background and three vulnerable geosites of Kızılcahamam-Çamlıdere Project in Ankara, Turkey. Geoheritage, 4:249-261. https://doi.org/10.1007/s12371-012-0064-2
Keller, J., Jung, D., Eckhardt, F.J., and Kreuzer, H. 1992. Radiometric ages and chemical characterization of the Galatean Andesite Massif, Pontus, Turkey. Acta Vulcanologica, 2:267-276.
Koçyiğit, A., Winchester, J.A., Bozkurt, E., and Holland, G. 2003. Sarackoy volcanic suite: Implications for the subductional phase of arc evolution in the Galatean arc complex, Ankara, Turkey. Geological Journal, 38:1-14. https://doi.org/10.1002/gj.921
Linnaeus, C. 1753. Species Plantarum. lmpensis Laurentii Salvii, Stockholm.
Mai, D.H. 1995. Tertiäre Vegetationsgeschichte Europas. Methoden und Ergebnisse. Gustav Fischer Verlag, Jena, Stuttgart, and New York.
Mirbel, C.F.B. 1815. Eléments de Physiologie Végétale et de Botanique. Magimel Publishing. Paris.
Mustoe, G.E. and Leopold, E.B. 2014. Paleobotanical evidence for post-Miocene uplift of the Cascade Range. Canadian Journal of Earth Sciences, 51:809-824. https://doi.org/10.1139/cjes-2013-0223
Özalp, G. 2000. [Sclerophyllous trees and macchie]. Review of Faculty of Forestry, University of Istanbul, 50(2-A):131-155. [In Turkish]
Persoon, C.H. 1807. Synopsis Plantarum, seu Enchiridion botanicum, Complectens Enumerationem Systematicam Specierum Hucusque Cognitarum. Pars secunda. Parisiis Lutetiorum: apud Bibliopolas Treuttel et Würtz, et Tübingen, apud J. G. Cottam. Treuttel and Würtz; Tübingen.
Popov, S.V., Rögl, F., Rozanov, A.Y., Steininger, F.F., Shcherba, I.G., and Kovac, M. 2004. Lithological-Paleogeographic Maps of Paratethys-10 Maps Late Eocene to Pliocene. Courier Forschungsinstitut Senckenberg, Band 250. E. Schweizerbart’sche Verlagsbuchhandlung (Nägela u. Obermiller), Stuttgart.
Pujana, R.R. and Ruiz, D.P. 2017. Podocarpoxylon Gothan reviewed in the light of a new species from the Eocene of Patagonia. IAWA Journal, 38(2):220-244. https://doi.org/10.1163/22941932-20170169
Richard, L.C. M. 1810. Note sur les plantes dites conifers. Annales du Museum National d’Histoire Naturelle, 16:296-299.
Rögl, F. 1999. Mediterranean and Paratethys. Facts and hypotheses of an Oligocene to Miocene paleogeography (short overview). Geologica Carpathica, 50(4):339-349.
Rudolphi, F. 1830. Systema orbis vegetabilium. Gryphiae, (Publisher’s locality unknown).
Sakınç, M., Aras, A., Yaltırak, C., Batı, Z., and Çağatay, N., 2007. [Silicified Trees in Tertiary of Thrace: Paleoflora, Paleoclimatology, Paleogeography. Scientific and Technical Research Council of Turkey Project Number: 103Y137]. Research Council of Turkey, İstanbul. [In Turkish]
Schildgen, T.F., Cosentino, D., Bookhagen, B., Niedermann, S., Yıldırım, C., Echtler, H., Wittmann, H., and Strecker, M.R. 2012a. Multi-phased uplift of the southern margin of the Central Anatolian Plateau, Turkey: A record of tectonic and upper mantle processes. Earth and Planetary Science Letters, 317-318:85-95. https://doi.org/10.1016/j.epsl.2011.12.003
Schildgen, T.F., Cosentino, D., Caruso, A., Buchwaldt, R., Yıldırım, C., Bowring, S.A., Rojay, B., Echtler, H., and Strecker, M.R. 2012b. Surface expression of eastern Mediterranean slab dynamics: Neogene topographic and structural evolution of the southwest margin of the Central Anatolian Plateau, Turkey. Tectonics, 31:TC2005. https://doi.org/10.1029/2011TC003021
Schroeder, G.F. 1998. Lehrbuch der Pflanzengeographie. Quelle & Meyer, Wiesbaden.
Schweingruber, F.H. 1990. Anatomy of European Woods. Verlag Paul Haupt, Bern and Stuttgart.
Schweingruber, F.H. 1993. Trees and Wood in Dendrochronology. Springer-Verlag, Berlin and Heidelberg.
Sen, S., Delfino, M., and Kazanci, N. 2017. Çeştepe, a new early Pliocene vertebrate locality in Central Anatolia and its stratigraphic context. Annales de Paléontologie, 103:149-163. https://doi.org/10.1016/j.annpal.2017.01.004
Süss, H. and Velitzelos, E. 2000. Zwei neue fossile Hölzer der Formgattung Podocarpoxylon Gothan aus tertiären Schichten der Insel Lesbos, Griechenland. Feddes Repertorum, 111:135-149. https://doi.org/10.1002/fedr.20001110304
Süzen, M.L. and Türkmenoglu, A.G. 2000. Lacustrine mineral facies and implications for estimation of palaeoenvironmental parameters: Neogene intervolcanic Pelitçik Basin (Galatean Volcanic Province), Turkey. Clay Minerals, 35(3):461-475. https://doi.org/10.1180/000985500546882
Tankut, A., Satır, M., Güleç, N., and Toprak, V. 1995. [Petrogenesis of Galatia Volcanoes, TÜBİTAK Project No. YBAG-0059]. TÜBİTAK, Ankara. [In Turkish]
Tankut, A., Wilson, M., and Yihunie, T. 1998. Geochemistry and tectonic setting of Tertiary volcanism in the Güvem area, Anatolia, Turkey. Journal of Volcanology and Geothermal Research, 85:285-301. https://doi.org/10.1016/s0377-0273(98)00060-2
Tatlı, S. 1975. [Geology and Geothermal Energy Possibilities of Kizilcahamam East Area]. MTA Report, No. 5749. MTA Publication, Ankara. [In Turkish]
Tidwell, W.D. 1998. Common Fossil Plants of Western North America. Smithsonian Institution Press, Washington and London.
Toprak, V., Savasçın, Y., Güleç, N., and Tankut, A. 1996. Structure of the Galatia volcanic province. International Geology Review, 38:747-758. https://doi.org/10.1080/00206819709465358
Trew, C.J. 1757. Cedrorum Libani Historia: Earumque Character Botanicus Cum Illo Laricis, Abietis Pinique Comparatus. Accedit Brevis Disquis. Impensis W. Schwarzkopfii, Norimbergae. [In Latin]
Türkecan, A., Hepşen, N., Papuk, I., Akbuş, B., Dinçel, A., Karataş, S., Özgür, İ., Akay, E., Bedi, Y., Sevin, M., Mutlu, G., Sevin, D., Ünay, E., and Saraç, G. 1991. Petrology of Geology and Volcanic Rocks of Seban-Gerede (Bolu) -Güdül-Beypazarı (Ankara) and Çerkeş-Orta-Kurşunlu (Çankırı) Regions (Köroğlu Mountains). MTA Report No. 9193. MTA Publication, Ankara. [In Turkish]
Üçbaş Durak, S.D. and Akkiraz, M.S. 2016. Kalkım-Gönen Havzasındaki Kömürlü Tortulların Paleoekolojik Özellikleri, Kuzeybatı Anadolu. Geology Bulletin of Turkey, 59(3):239-258.
Uhl, D., Dolezych, M., and Böhme M. 2014. Taxodioxylon-like charcoal from the Late Miocene of western Bulgaria. Acta Palaeobotanica, 54(1):101-111. https://doi.org/10.2478/acpa-2014-0004
Utescher, T., Djordjevic-Milutinovic, D., Bruch, A., and Mosbrugger, V. 2007. Palaeoclimate and vegetation change in Serbia during the last 30 Ma. Palaeogeography, Palaeoclimatology, Palaeoecology, 253:157-168. https://doi.org/10.1016/j.palaeo.2007.03.037
Van den Hoek Ostende, L.W. 2001. A revised generic classification of the Galericini (Insectivora, Mammalia) with some remarks on their palaeobiogeography and phylogeny. Geobios, 34 (6): 681-695. https://doi.org/10.1016/S0016-6995(01)80029-2
Vassio, E., Martinetto, E., Dolezych, M., and Van der Burgh, J. 2008. Wood anatomy of the Glyptostrobus europeus “whole-plant” from a Pliocene forest of Italy. Review of Palaeobotany and Palynology, 151:81-89. https://doi.org/10.1016/j.revpalbo.2008.02.006
Velitzelos, D., Bouchal, J.M., and Denk, T. 2014. Review of the Cenozoic floras and vegetation of Greece. Review of Palaeobotany and Palynology, 204:56-117. https://doi.org/10.1016/j.revpalbo.2014.02.006
Vosoughi Moradi, A., Sarı, A., and Akkaya, P. 2016. Geochemistry of the Miocene oil shale (Hançili Formation) in the Çankırı-Çorum Basin, Central Turkey: Implications for Paleoclimateconditions, source-area weathering, provenance and tectonic setting. Sedimentary Geology, 341:289-303. https://doi.org/10.1016/j.sedgeo.2016.05.002
Wheeler, E.A. 2011. InsideWood - a web resource for hardwood anatomy. IAWA Journal, 32(2):199-211. https://doi.org/10.1163/22941932-90000051
Wilson, M., Tankut, A., and Guleç, N. 1997. Tertiary volcanism of the Galatia Province, NW Central Anatolia, Turkey. Lithos, 42:105-121. https://doi.org/10.1016/S0024-4937(97)00039-X
Yaltırık, F. 1971. [Morphological and Anatomical Investigations on native maple (Acer L.) taxa in Turkey. Istanbul University Forest Faculty Publications, No. 179]. Istanbul University Forest Faculty, İstanbul, Turkey. [In Turkish]
Yaltırık, F. and Akkemik, Ü. 2012. [Native Conifers of Turkey]. General Directorate of Forestry Publications, İstanbul. [In Turkish]
Yavuz-Işık, N. 2008. [Palynology of Lower Miocene lacustrine sediments of the Güvem Basin (Central Anatolia)], p. 337. In Genç, Y. and Kuzucu, C. (eds.), Proceedings of the 61st Geological Congress of Turkey. MTA, Ankara. [In Turkish]
Yavuz-Işık, N. and Demirci, C. 2009. Miocene spores and pollen from Pelitçik Basin, Turkey-environmental and climatic implications. Comptes Rendus Palevol, 8 (5):437-446. https://doi.org/10.1016/j.crpv.2009.02.002
Zachos, J.C., Dickens, G.R., and Zeebe, R.E. 2008. An early Cenozoic perspective on greenhouse warming and carbon-cycle Dynamics. Nature, 451:17. https://doi.org/10.1038/nature06588
