

 Leaf trait data of two Miocene floras from eastern China and its palaeoclimate implications
Leaf trait data of two Miocene floras from eastern China and its palaeoclimate implications
Article number: 26.2.a23
https://doi.org/10.26879/1262
Copyright Paleontological Society, July 2023
Author biographies
Plain-language and multi-lingual abstracts
PDF version
Submission: 22 December 2022. Acceptance: 25 June 2023.
ABSTRACT
The study of Neogene palaeoclimate supports our understanding of effects and consequences of current climate changes. However, many aspects and details of Miocene climate development are still unclear. Fossil leaves are a valuable and rich source of palaeoclimate proxy data. In this contribution, two Miocene leaf assemblages from eastern China, the Toupi flora and the Shengxian flora, were studied with respect to palaeoclimate and leaf economics. Whereas the Shengxian flora is dated to the lower Tortonian, the Toupi flora dates to approximately the border between Burdigalian and Langhian. For palaeoclimate, Climate Leaf Analysis Multivariate Program (CLAMP) was applied. In addition, Leaf Mass per Area (LMA), an essential leaf trait strongly correlated to leaf longevity, was included and was calculated morphometrically. For both sites, the data indicated a principally warm and humid climate, with Mean Annual Temperature (MAT) values between 13 and 17 °C. Also, the LMA indicates evergreen vegetation for both sites, consistent with the identified fossils and palaeoclimate. For Shengxian, however, this study concludes there was a lower temperature in the cooler season. This might possibly indicate a stronger temperature seasonality for this site due to slight climate cooling and East Asian monsoon intensification from the middle to late Miocene.
Wenlong He. College of Geology and Environment, Xi’an University of Science and Technology, Xi'an 710054, China. State Key Laboratory of Palaeobiology and Stratigraphy, Nanjing Institute of Geology and Palaeontology, Chinese Academy of Sciences, Nanjing 210008, China. hegeosci@gmail.com
Anita Roth-Nebelsick. State Museum of Natural History Stuttgart, Rosenstein 1, Stuttgart 70191, Germany. anita.rothnebelsick@smns-bw.de
Bainian Sun. School of Earth Sciences & Key Laboratory of Mineral Resources of Western China (Gansu Province), Lanzhou University, Lanzhou 730000, China. bnsun@lzu.edu.cn
Keywords: Miocene; leaf traits; leaf mass per area; CLAMP; vegetation; palaeoclimate
Final citation: He, Wenlong, Roth-Nebelsick, Anita, and Sun, Bainian. 2023. Leaf trait data of two Miocene floras from eastern China and its palaeoclimate implications. Palaeontologia Electronica, 26(2):a23.
https://doi.org/10.26879/1262
palaeo-electronica.org/content/2023/3883-leaf-traits-of-miocene-floras
Copyright: July 2023 Paleontological Society.
This is an open access article distributed under the terms of Attribution-NonCommercial-ShareAlike 4.0 International (CC BY-NC-SA 4.0), which permits users to copy and redistribute the material in any medium or format, provided it is not used for commercial purposes and the original author and source are credited, with indications if any changes are made.
creativecommons.org/licenses/by-nc-sa/4.0/
INTRODUCTION
During the Cenozoic, major climate changes occurred, such as the PETM (Paleocene Eocene Thermal Maximum), the Early Eocene Climate Optimum or the Eocene/Oligocene transition with global cooling (Zachos et al., 2001, 2008; Westerhold et al., 2020). Also the Miocene witnessed substantial climate fluctuations, particularly the Middle Miocene Climatic Optimum and the Middle Miocene Climate transition with subsequent global cooling (Steinthorsdottir et al., 2021). For instance, previous studies have suggested that an arid region was formed in Northwest China during the Miocene, which might have been caused by the intensification of the Asian monsoon (Guo et al., 2002; Sun and Wang, 2005), further linked with the uplift of the Tibetan Plateau (An et al., 2001). Based on fossil plant data, however, Wang et al. (2021) proposed that an equable subtropical climate prevailed throughout China in the Miocene. Miao et al. (2022) suggested that Northwest China was drier than Southeast China, a gradient perhaps resulting from global cooling during the Miocene.
Leaf traits are a rich source of climate information. Leaf Margin Analysis (Wolfe, 1979; Wing and Greenwood, 1993; Su et al., 2010) and the Climate Leaf Analysis Multivariate Program (CLAMP) (Wolfe, 1993; Yang et al., 2015) are two well-established methods for quantitative reconstruction of climate parameters. Leaf mass per area (LMA) represents an essential parameter in leaf economics and is strongly correlated with leaf longevity (Reich et al., 1997; Villar and Merino, 2001; Wright et al., 2004), thereby providing information on evergreen vs. deciduous vegetation (Royer et al., 2007; Royer et al., 2010; Roth-Nebelsick et al., 2017). LMA is considered a key parameter for the ecology and ecophysiology of modern floras (Wright et al., 2004; Ordoñez et al., 2009; Kröber et al., 2012; Chen and Xu, 2014). Reconstructing LMA from fossil leaf assemblages is possible by a morphometric method developed by Royer et al. (2007). Since leaf longevity depends on climate (Givnish, 2002), a combination of both methods, palaeoclimate reconstruction and fossil LMA data, can potentially deepen our understanding of palaeoenvironments.
In this study, leaf-based methods were applied to two fossil assemblages to contribute data and information with respect to climate development during the Miocene in eastern China. For this, the CLAMP was used for reconstructing palaeoclimate. For both sites, LMA was also calculated.
MATERIALS AND METHODS
Fossil Sites and Age
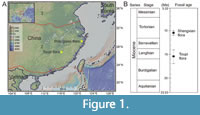 The Toupi fossil flora is located at Toupi town in Guangchang County, Jiangxi Province, eastern China (N 26.751 and E116.192; Figure 1A). The Toupi fossil flora was initially reported as evidence of Neogene strata in Jiangxi Province 40 years ago, and new fossil material has been collected and restudied in recent years (He and Wang, 2021). Fossiliferous strata belong to the Toupi Formation, which comprises mainly mudstone and shale. For the fossil-bearing sediments, pollen data indicate an age between 17 and 14 Ma (Figure 1B; Table 1; Sun and He, 1987; Ma et al., 2020; He and Wang, 2021). Most of this time interval belongs to the Miocene Climatic Optimum.
The Toupi fossil flora is located at Toupi town in Guangchang County, Jiangxi Province, eastern China (N 26.751 and E116.192; Figure 1A). The Toupi fossil flora was initially reported as evidence of Neogene strata in Jiangxi Province 40 years ago, and new fossil material has been collected and restudied in recent years (He and Wang, 2021). Fossiliferous strata belong to the Toupi Formation, which comprises mainly mudstone and shale. For the fossil-bearing sediments, pollen data indicate an age between 17 and 14 Ma (Figure 1B; Table 1; Sun and He, 1987; Ma et al., 2020; He and Wang, 2021). Most of this time interval belongs to the Miocene Climatic Optimum.
The Shengxian fossil flora is located at Zhejiang Province, eastern China (N 29.159 and E 121.240; Figure 1A). The fossiliferous strata belong to Shengxian Formation, which occurs at various sites of eastern Zhejiang, such as Jiahu Village in Tiantai County, Daluxia Village, Huangnitang Village and Gaotang Village in Ninghai County, and Xiananshan Village in Sanmen County (Li et al., 2014). The Shengxian Formation is composed of diatomite, mud shale and basalt interbeds, with a total of five sedimentary units. The fossil plants are preserved in the second sedimentary unit within the region (Li et al., 2014) and can be dated to 10.5 (±0.5) Ma according to geochemical data (Ho et al., 2003; Figure 1B), thereby falling within the cooling phase of the late Miocene, already a time of substantial polar ice sheet formation.
Fossil Materials
 At the present day, the local environment and geographical conditions of the Toupi and Shengxian fossil sites are similar, both are hilly landscapes with elevations of 170 m and 280 m and situated at latitudes of ca. N 27° and N 29° (Table 1). Both the Toupi and Shengxian areas support evergreen broad-leaved forest nowadays (Zhang et al., 2007 Figure1; Table 1).
At the present day, the local environment and geographical conditions of the Toupi and Shengxian fossil sites are similar, both are hilly landscapes with elevations of 170 m and 280 m and situated at latitudes of ca. N 27° and N 29° (Table 1). Both the Toupi and Shengxian areas support evergreen broad-leaved forest nowadays (Zhang et al., 2007 Figure1; Table 1).  Fossil plant materials from the Toupi Formation and the Shengxian Formation both include leaves, fruits and seeds. In total, 216 specimens from the Toupi flora and more than 1000 specimens from the Shengxian flora were checked for suitability for the present study. Only fossil woody dicots were selected for this study as the above described methods are limited to dicot leaves. Furthermore, complete or almost complete specimens were required for the measurements conducted in this study. Only few specimens, therefore, were suitable for the analysis. Various leaves are complete or almost complete and could be selected for measurement of leaf traits (Appendix 1 and Appendix 2). Finally, 22 and 31 species/morphotypes were identified for Toupi and Shengxian floras, respectively, with 36 specimens of Toupi and 68 specimens of Shengxian scored for CLAMP analysis, while LMA data could be determined for 14 specimens of Toupi and 26 for Shengxian.
Fossil plant materials from the Toupi Formation and the Shengxian Formation both include leaves, fruits and seeds. In total, 216 specimens from the Toupi flora and more than 1000 specimens from the Shengxian flora were checked for suitability for the present study. Only fossil woody dicots were selected for this study as the above described methods are limited to dicot leaves. Furthermore, complete or almost complete specimens were required for the measurements conducted in this study. Only few specimens, therefore, were suitable for the analysis. Various leaves are complete or almost complete and could be selected for measurement of leaf traits (Appendix 1 and Appendix 2). Finally, 22 and 31 species/morphotypes were identified for Toupi and Shengxian floras, respectively, with 36 specimens of Toupi and 68 specimens of Shengxian scored for CLAMP analysis, while LMA data could be determined for 14 specimens of Toupi and 26 for Shengxian.
Leaf Traits Measurement
 For CLAMP, the general protocol was followed to collect the necessary leaf traits determined by the character definitions of the CLAMP datasets (http://clamp.ibcas.ac.cn/; Wolfe, 1993; Yang et al., 2011). Most of the selected fossil leaves were complete or almost complete in preservation, and it was possible to reconstruct these slightly fragmented leaves additionally to obtain quantitative data, such as lamina length (L), lamina width (W) and lamina area (A) by using ImageJ (Ferreira and Rasband, 2012; Traiser et al., 2018; Toumoulin et al., 2020; Figure 2; Appendix 3 and Appendix 4). For determination of LMA, leaf area and petiole width were measured and then LMA was calculated using the equation established by Royer et al. (2007):
For CLAMP, the general protocol was followed to collect the necessary leaf traits determined by the character definitions of the CLAMP datasets (http://clamp.ibcas.ac.cn/; Wolfe, 1993; Yang et al., 2011). Most of the selected fossil leaves were complete or almost complete in preservation, and it was possible to reconstruct these slightly fragmented leaves additionally to obtain quantitative data, such as lamina length (L), lamina width (W) and lamina area (A) by using ImageJ (Ferreira and Rasband, 2012; Traiser et al., 2018; Toumoulin et al., 2020; Figure 2; Appendix 3 and Appendix 4). For determination of LMA, leaf area and petiole width were measured and then LMA was calculated using the equation established by Royer et al. (2007):
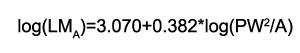
Climate Leaf Analysis Multivariate Program (CLAMP)
To obtain leaf-based palaeoclimate data for the Miocene Toupi and Shengxian, CLAMP was applied (Appendix 5 and Appendix 6). The Climate Leaf Analysis Multivariate Program (CLAMP) is a quantitative method to reconstruct the paleoclimate using woody dicotyledonous leaf architecture, including foliar characters such as margin, size, and shape. There is ample evidence that many leaf characters correlate with climate, and these are included in calibration datasets to reconstruct palaeoclimate. At present, five datasets are available for calibration (http://clamp.ibcas.ac.cn/), i.e., Physg3arcAZ, Physg3brcAZ, PhysgAsia1 (Jacques et al., 2011a), PhysgAsia2 (Khan et al., 2014) and PhysgGlobal378 (Yang et al., 2015). Among them, PhysgAsia1 is particularly suitable for monsoon climate because this dataset encompasses 45 sites from China (Jacques et al., 2011a) and was, therefore, used in this study. The leaf characters were scored with the template sheets, followed by a table with percentage values of the leaf characters (Appendix 5 and Appendix 6), and then the analysis was conducted online with suitable calibration (http://clamp.ibcas.ac.cn/).
RESULTS
Leaf Size
Leaf size is one of the traits used in CLAMP - albeit as categorical data - and represents a globally informative parameter. Therefore, these results were specifically considered here. In general, leaf size can be approximated by leaf length (L) and leaf width (W), and depends also on the leaf shape (Schrader et al., 2021). In CLAMP, leaf size is determined by the use of templates (http://clamp.ibcas.ac.cn/).
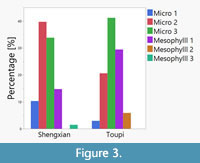 The results indicate that the average leaf size of the Toupi flora (2037.88 mm2) is larger than that of Shengxian flora (1033.35 mm2). The difference is statistically significant (t-test with log10-transformed data, p < 0.001) and also obvious from the CLAMP data (Figure 3). This difference between both considered sites is probably partially caused by the circumstance, that more broad-leaved taxa, such as Lithocarpus or Juglans, are present in the Toupi flora compared to the Shengxian flora.
The results indicate that the average leaf size of the Toupi flora (2037.88 mm2) is larger than that of Shengxian flora (1033.35 mm2). The difference is statistically significant (t-test with log10-transformed data, p < 0.001) and also obvious from the CLAMP data (Figure 3). This difference between both considered sites is probably partially caused by the circumstance, that more broad-leaved taxa, such as Lithocarpus or Juglans, are present in the Toupi flora compared to the Shengxian flora.
Leaf Margin
Most leaves are untoothed in both the Toupi and the Shengxian flora with percentage values of 77.27 % and 67.74 %, respectively. Furthermore, the toothed leaves all are regular. The Toupi flora has five toothed taxa, with one distantly toothed, and no round or compound teeth. Ten taxa are toothed in the Shengxian flora, three distantly toothed, and three compound teeth, but no round ones.
Leaf Mass per Area (LMA)
Based on the preserved or partly preserved petioles of the Toupi and Shengxian fossil leaves, together with the data for lamina area, the following LMA values were found: 101.51 g/m2 for the Toupi flora and 91.93 g/m2 for the Shengxian flora (Table 2). In both cases, the data indicate an evergreen vegetation type (Villar and Merino, 2001; Royer et al., 2007).
CLAMP Results
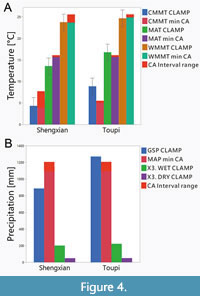 Based on the collected leaf traits data (Appendix 3, Appendix 4, Appendix 5 and Appendix 6), an online CLAMP analysis with the PhysgAsia1 calibration was conducted (http://clamp.ibcas.ac.cn/). The results are given in Table 3 and Figure 4, which also include data from a former palaeoclimate study for comparison. Note that CLAMP MAT results are associated with uncertanties of about plus or minus 2 °C, and the other climate variables with somewhat more.
Based on the collected leaf traits data (Appendix 3, Appendix 4, Appendix 5 and Appendix 6), an online CLAMP analysis with the PhysgAsia1 calibration was conducted (http://clamp.ibcas.ac.cn/). The results are given in Table 3 and Figure 4, which also include data from a former palaeoclimate study for comparison. Note that CLAMP MAT results are associated with uncertanties of about plus or minus 2 °C, and the other climate variables with somewhat more.
These climate parameters show warm and humid climates for both Toupi and Shengxian floras. The lower Miocene MAT for Shengxian (13.6 ℃), when compared to Toupi (16.7 ℃), is mainly caused by a lower coldest mean month temperature (CMMT). With respect to warmest mean month temperature (WMMT), both sites are almost the same. The difference between the coldest and warmest month temperatures (DT) amounts to 15.5 ℃ for Toupi and 19.8 ℃ for Shengxian.
Compared to the current climate, the palaeoclimate of both Miocene sites is somewhat cooler. At present day, MAT amounts to 18.5 ℃ for Toupi and 16.4 ℃ for Shengxian (Table 3 and Table 4, Figure 4). Also, the DT is higher under current conditions, amounting to 24.1 ℃ for Toupi and 23 ℃ for Shengxian (Table 4). Humidity values indicate a quite humid climate which includes, however, a season with lower precipitation. The data indicate a somewhat lower precipitation in Shengxian compared to Toupi (Figure 4; Table 3 and Table 4).
DISCUSSION
Leaf Traits
The data indicate a larger leaf size for the Toupi flora compared to the Shengxian flora. This difference might be caused by various factors, such as soil conditions, temperature, precipitation and geographical conditions. Generally, leaf size is positively correlated with temperature and water availability (Wright et al., 2017). Also, leaf size tends to be positively correlated with nutrient availability (McDonald et al., 2003). Palaeo-humidity values obtained in this study are, however, quite similar for both sites (Figure 4; Table 3). Also, results of previous climate reconstructions based on the Coexistence Approach (CA) indicate similar humidity conditions for both sites (Yao et al., 2011; Figure 4). In contrast to these previous CA studies (which will be further considered, see below), CLAMP data indicate stronger temperature seasonality for the Shengxian flora, compared to the Toupi flora. There are also more deciduous taxa present in the Shengxian flora, like Betula and Ulmus (Table 5). Possibly, the smaller leaf size of the Shengxian flora may, therefore, be at least partially caused by lower temperature, but other unknown factors may have also been influential.
The leaf mass per area (LMA) provides significant information with respect to leaf economics and leaf longevity (LL) (Villar and Merino, 2001; Wright et al., 2004; Royer et al., 2007), with an LL of > 12 months considered as evergreen (Givnish, 2002). The average LMA of the Toupi flora, with an LMA of 101.51 g/m2, is higher than that of the Shengxian flora (91.93 g/m2) (Table 2). These values are typical for evergreen vegetation (Mediavilla et al., 2008).
Further information on the vegetation of both sites is provided by a study using the Integrated Plant Record (IPR) analysis, indicating that the Toupi flora represents a broad - leaved evergreen forest, while the Shengxian flora was a subhumid sclerophyllous forest (Jacques et al., 2011b). This vegetation type of the Shengxian flora appears, however, to be in contrast with the LMA results, because a higher LMA is expected for a sclerophyllous flora, compared to a broadleaved flora. The reason might be that the IPR analyses conducted by Jacques et al. (2011b) were based on pollen data rather than on leaf macro fossils. In fact, applying the IPR analysis to macro fossils of the Shengxian flora also indicates an evergreen broad-leaved forest vegetation. The pollen data possibly included vegetation, which originated not directly from the Shengxian site but from a larger catchment area (due to pollen transport) and therefore from other habitats.
Palaeoclimate
Based on plenty of fossils, the Shengxian flora has been studied in various fields, such as plant evolution, palaeoclimate and stratigraphy (Li et al., 2014). Previous studies indicated appropriate subtropical climate with cool winters in the Shengxian region during late Miocene (Yao et al., 2011; Li et al., 2014). In terms of the Toupi fossil flora, the pollen data and the plant composition also supported a warm and moist climate during the middle Miocene (Sun and He, 1987; He and Wang, 2021).
The fossil floras of Toupi and Shengxian were considered in previous studies on regional palaeoclimate by using CA (Yao et al., 2011; Wang et al., 2021). However, these CA results have indicated almost identical MAT (15.7 - 16.1℃) for both the Toupi flora and Shengxian flora (Yao et al., 2011; Table 4). Meanwhile, CA results indicating an equable subtropical climate throughout China in the Miocene (Wang et al., 2021) were questioned by Miao et al. (2022).
The CLAMP results obtained in the present study indicate a cooler MAT (13.6 ℃) for Shengxian, caused by a certain seasonality as CLAMP provided a lower CMMT for Shengxian compared to Toupi (Table 4).
At the present day, a dry-cold winter and warm-humid summer monsoon climate prevails in eastern China, and apparently since the late Oligocene (An, 2000; Sun and Wang, 2005; Wu et al., 2022). The results of the present study may possibly indicate a moderate intensification of seasonality from the middle Miocene to late Miocene, i.e, the DT increased from 15.5 ℃ (Toupi) at the middle Miocene to 19.8 ℃ (Shengxian) at the late Miocene. That would mean the East Asian monsoon may have intensified during that time. This assumption is partly supported by Sun and Wang (2005) and recent data (Yang et al., 2021). Monsoonal development during the Miocene is, however, complex and a controversial topic. The results of the present study are based on two fossil leaf assemblages, and more sites should be studied to obtain information on climate development in eastern China during the Miocene.
CONCLUSIONS
In this study, leaf trait data from two Miocene floras of China were obtained. The palaeoclimate proxies derived by these leaf traits indicate climate cooling during the Middle Miocene Climate transition due to higher temperature seasonality with possible relevance for East Asian monsoon development. The well-preserved leaves also allowed to determine leaf mass per area data indicating evergreen vegetation for both sites. More leaf-based studies of palaeoenvironment and palaeoclimate for this region are desirable.
ACKNOWLEDGEMENTS
This study supported by State Key Laboratory of Palaeobiology and Stratigraphy (Nanjing Institute of Geology and Palaeontology, CAS) (Nos. 213105, 213115), and the National Science Foundation of China (41972010). W. He was supported by China Scholarship Council (CSC grant 202108610118) during a visit to the State Museum of Natural History Stuttgart. Our thanks to anonymous referees and the handling editor, M. Pole, for their comments that helped improve this manuscript.
REFERENCES
An, Z. 2000. The history and variability of the East Asian paleomonsoon climate. Quaternary Science Reviews, 19:171-187.
https://doi.org/10.1016/S0277-3791(99)00060-8
An, Z., Kutzbach, J.E., Prell, W.L., and Porter, S.C. 2001. Evolution of Asian monsoons and phased uplift of the Himalaya-Tibetan plateau since Late Miocene times. Nature, 411:62-66.
https://doi.org/10.1038/35075035
Chen, Y. and Xu, Z. 2014. Review on research of leaf economics spectrum. Chinese Journal of Plant Ecology, 38:1135-1153. (In Chinese, with English abstract)
Ding, S. 2011. Reconstruction of paleovegetation and paleoclimate of the late Miocene flora in Tiantai, Zhejiang, China. Unpublished PhD Thesis, Lanzhou University, Lanzhou, China.
Ferreira, T. and Rasband, W. 2012. ImageJ user guide. ImageJ/Fiji, 1:155-161.
Givnish, T.J. 2002. Adaptive significance of evergreen vs. deciduous leaves: solving the triple paradox. Silva Fennica, 36:703-743.
https://doi.org/10.14214/sf.535
Guo, Z.T., Ruddiman, W.F., Hao, Q.Z., Wu, H.B., Qiao, Y.S., Zhu, R.X., Peng, S.Z., Wei, J.J., Yuan, B.Y., and Liu, T.S. 2002. Onset of Asian desertification by 22 Myr ago inferred from loess deposits in China. Nature, 416:159-163.
https://doi.org/10.1038/416159a
He, W. and Wang, X. 2021. A Miocene flora from the Toupi Formation in Jiangxi Province, southeastern China. Palaeoworld, 30:757-769. https://doi.org/10.1016/j.palwor.2020.12.006
He, Y. 2017. Geologic time and ecological environment of the Shengxian Formation flora from eastern Zhejiang and volcanic activity influence. Unpublished PhD Thesis, Lanzhou University, Lanzhou, China.
Ho, K.S., Chen, J.C., Lo, C.H., and Zhao, H.L. 2003. 40Ar - 39Ar dating and geochemical characteristics of late Cenozoic basaltic rocks from the Zhejiang-Fujian region, SE China: eruption ages, magma evolution and petrogenesis. Chemical Geology, 197:287-318.
https://doi.org/10.1016/S0009-2541(02)00399-6
Jacques, F.M.B., Shi, G., and Wang, W. 2011b. Reconstruction of Neogene zonal vegetation in South China using the Integrated Plant Record (IPR) analysis. Palaeogeography, Palaeoclimatology, Palaeoecology, 307:272-284.
https://doi.org/10.1016/j.palaeo.2011.05.025
Jacques, F.M., Su, T., Spicer, R.A., Xing, Y., Huang, Y., Wang, W., and Zhou, Z. 2011a. Leaf physiognomy and climate: are monsoon systems different? Global and Planetary Change, 76:56-62.
https://doi.org/10.1016/j.gloplacha.2010.11.009
Khan, M.A., Spicer, R.A., Bera, S., Ghosh, R., Yang, J., Spicer, T.E., Guo, S., Su, T., Jacques, F., and Grote, P.J. 2014. Miocene to Pleistocene floras and climate of the Eastern Himalayan Siwaliks, and new palaeoelevation estimates for the Namling-Oiyug Basin, Tibet. Global and Planetary Change, 113:1-10.
https://doi.org/10.1016/j.gloplacha.2013.12.003
Kröber, W., Böhnke, M., Welk, E., Wirth, C., and Bruelheide, H. 2012. Leaf trait-environment relationships in a subtropical broadleaved forest in South-East China. PLoS ONE, 7:e35742.
https://doi.org/10.1371/journal.pone.0035742
Li, X., Sun, B., Xiao, L., Ding, S., He, W., and Du, B. 2014. Stratum characteristics of the Neogene Shengxian Formation in Zhejiang Province and its related fossil studies. Journal Lanzhou University (Natural Science), 50:145-153. (In Chinese, with English abstract)
Ma, F., Ling, C., Ou-Yang, M., Yang, G., Shen, X., and Wang, Q. 2020. Plant-insect interactions from the Miocene (Burdigalian-Langhian) of Jiangxi, China. Review of Palaeobotany and Palynology, 275:104176.
https://doi.org/10.1016/j.revpalbo.2020.104176
McDonald, P., Fonseca, C., McC, J., and Westoby, M. 2003. Leaf-size divergence along rainfall and soil-nutrient gradients: is the method of size reduction common among clades? Functional Ecology, 17(1):50-57.
https://doi.org/10.1046/j.1365-2435.2003.00698.x
Mediavilla, S., Garcia-Ciudad, A., Garcia-Criado, B., and Escudero, A. 2008. Testing the correlations between leaf life span and leaf structural reinforcement in 13 species of European Mediterranean woody plants. Functional Ecology, 22:787-793.
https://doi.org/10.1111/j.1365-2435.2008.01453.x
Miao, Y., Xiang, M., Zhang, T., and Warny, S. 2022. Comment on “An equable subtropical climate throughout China in the Miocene based on palaeofloral evidence” by Qing Wang, Yan Li David K. Ferguson, Wen-Bo Mo, and Nan Yang, Earth-Science Reviews, [Earth-Science Reviews 218 (2021) 103649]. Earth-Science Reviews:104020.
https://doi.org/10.1016/j.earscirev.2022.104020
Ordoñez, J.C., Van Bodegom, P.M., Witte, J.P.M., Wright, I.J., Reich, P.B., and Aerts, R. 2009. A global study of relationships between leaf traits, climate and soil measures of nutrient fertility. Global Ecology and Biogeography, 18:137-149.
https://doi.org/10.1111/j.1466-8238.2008.00441.x
Reich, P.B., Walters, M.B., and Ellsworth, D.S. 1997. From tropics to tundra: global convergence in plant functioning. Proceedings of the National Academy of Sciences, 94:13730-13734.
https://doi.org/10.1073/pnas.94.25.13730
Roth-Nebelsick, A., Grein, M., Traiser, C., Moraweck, K., Kunzmann, L., Kovar-Eder, J., Kvaček, J., Stiller, S., and Neinhuis, C. 2017. Functional leaf traits and leaf economics in the Paleogene--A case study for Central Europe. Palaeogeography, Palaeoclimatology, Palaeoecology, 472:1-14.
https://doi.org/10.1016/j.palaeo.2017.02.008
Royer, D.L., Miller, I.M., Peppe, D.J., and Hickey, L.J. 2010. Leaf economic traits from fossils support a weedy habit for early angiosperms. American Journal of Botany, 97:438-445.
https://doi.org/10.3732/ajb.0900290
Royer, D.L., Sack, L., Wilf, P., Lusk, C.H., Jordan, G.J., Niinemets, U., Wright, I.J., Westoby, M., Cariglino, B., and Coley, P.D. 2007. Fossil leaf economics quantified: calibration, Eocene case study, and implications. Paleobiology, 33:574-589.
https://doi.org/10.1666/07001.1
Schrader, J., Shi, P., Royer, D.L., Peppe, D.J., Gallagher, R.V., Li, Y., Wang, R., and Wright, I.J. 2021. Leaf size estimation based on leaf length, width and shape. Annals of Botany, 128:395-406.
https://doi.org/10.1093/aob/mcab078
Steinthorsdottir, M., Coxall, H., De Boer, A., Huber, M., Barbolini, N., Bradshaw, C., Burls, N., Feakins, S., Gasson, E., and Henderiks, J. 2021. The Miocene: the future of the past. Paleoceanography and Paleoclimatology, 36:e2020PA004037.
https://doi.org/10.1029/2020PA004037
Su, T., Xing, Y.W., Liu, Y.S., Jacques, F.M., Chen, W.Y., Huang, Y.J., and Zhou, Z.K. 2010. Leaf margin analysis: a new equation from humid to mesic forests in China. Palaios, 25:234-238.
https://doi.org/10.2110/palo.2009.p09-129r
Sun, X.J. and He, Y.M. 1987. Neogene spore-pollen assemblages from Jiangxi province, China. Botanical Research, 3:83-120. (In Chinese with English abstract)
Sun, X.J. and Wang, P.X. 2005. How old is the Asian monsoon system? Palaeobotanical records from China. Palaeogeography, Palaeoclimatology, Palaeoecology, 222:181-222.
https://doi.org/10.1016/j.palaeo.2005.03.005
Toumoulin, A., Kunzmann, L., Moraweck, K., and Sack, L. 2020. Reconstructing leaf area from fragments: testing three methods using a fossil paleogene species. American Journal of Botany, 107:1786-1797.
https://doi.org/10.1002/ajb2.1574
Traiser, C., Roth-Nebelsick, A., Grein, M., Kovar-Eder, J., Kunzmann, L., Moraweck, K., Lange, J., Kvaček, J., Neinhuis, C., and Folie, A. 2018. MORPHYLL: A database of fossil leaves and their morphological traits. Palaeontologia Electronica, 21: e1T.
https://doi.org/10.26879/773
Villar, R. and Merino, J. 2001. Comparison of leaf construction costs in woody species with differing leaf life-spans in contrasting ecosystems. New Phytologist, 151:213-226.
https://doi.org/10.1046/j.1469-8137.2001.00147.x
Wang, Q., Li, Y., Ferguson, D.K., Mo, W.-B., and Yang, N. 2021. An equable subtropical climate throughout China in the Miocene based on palaeofloral evidence. Earth-Science Reviews, 218:103649.
https://doi.org/10.1016/j.earscirev.2021.103649
Westerhold, T., Marwan, N., Drury, A.J., Liebrand, D., Agnini, C., Anagnostou, E., Barnet, J.S., Bohaty, S.M., De Vleeschouwer, D., and Florindo, F. 2020. An astronomically dated record of Earth’s climate and its predictability over the last 66 million years. Science, 369:1383-1387.
https://doi.org/10.1126/science.aba6853
Wing, S.L. and Greenwood, D.R. 1993. Fossils and fossil climate: the case for equable continental interiors in the Eocene. Philosophical Transactions of the Royal Society of London. Series B: Biological Sciences, 341:243-252.
https://doi.org/10.1098/rstb.1993.0109
Wolfe, J.A. 1979. Temperature parameters of humid to mesic forests of eastern Asia and relation to forests of other regions of the northern hemisphere and Australasia. USGS Professional Papers, 1106 (1979):1-37.
Wolfe, J.A. 1993. A method of obtaining climatic parameters from leaf assemblages. US Government Printing Office, USA.
Wright, I.J., Dong, N., Maire, V., Prentice, I.C., Westoby, M., Díaz, S., Gallagher, R.V., Jacobs, B.F., Kooyman, R., and Law, E.A. 2017. Global climatic drivers of leaf size. Science, 357:917-921.
https://doi.org/10.1126/science.aal4760
Wright, I.J., Reich, P.B., Westoby, M., Ackerly, D.D., Baruch, Z., Bongers, F., Cavender-Bares, J., Chapin, T., Cornelissen, J.H., and Diemer, M. 2004. The worldwide leaf economics spectrum. Nature, 428:821-827.
https://doi.org/10.1038/nature02403
Wu, F., Fang, X., Yang, Y., Dupont-Nivet, G., Nie, J., Fluteau, F., Zhang, T., and Han, W. 2022. Reorganization of Asian climate in relation to Tibetan Plateau uplift. Nature Reviews Earth & Environment, 3:684-700.
https://doi.org/10.1038/s43017-022-00331-7
Xiao, L., Wu, Z., Guo, L., Li, X., Ji, D., Xia, X., Wang, J., Liang, J., and Sun, N. 2022. Late Miocene Leaves and Endocarps of Choerospondias (Anacardiaceae) from Zhejiang, Eastern China: Implications for Paleogeography and Paleoclimate. Biology, 11:1399.
https://doi.org/10.3390/biology11101399
Yang, C., Dang, H., Zhou, X., Zhang, H., Wang, X., Wang, Y., Qiao, P., Jiang, X., and Jian, Z. 2021. Upper ocean hydrographic changes in response to the evolution of the East Asian monsoon in the northern South China Sea during the middle to late Miocene. Global and Planetary Change, 201:103478.
https://doi.org/10.1016/j.gloplacha.2021.103478
Yang, J., Spicer, R.A., Spicer, T.E., and Li, C.-S. 2011. ‘CLAMP Online’: a new web-based palaeoclimate tool and its application to the terrestrial Paleogene and Neogene of North America. Palaeobiodiversity and Palaeoenvironments, 91:163-183.
https://doi.org/10.1007/s12549-011-0056-2
Yang, J., Spicer, R.A., Spicer, T.E., Arens, N.C., Jacques, F.M., Su, T., Kennedy, E.M., Herman, A.B., Steart, D.C., and Srivastava, G. 2015. Leaf form-climate relationships on the global stage: an ensemble of characters. Global Ecology and Biogeography, 24:1113-1125.
https://doi.org/10.1111/geb.12334
Yao, Y.F., Bruch, A.A., Mosbrugger, V., and Li, C.S. 2011. Quantitative reconstruction of Miocene climate patterns and evolution in Southern China based on plant fossils. Palaeogeography, Palaeoclimatology, Palaeoecology, 304:291-307.
https://doi.org/10.1016/j.palaeo.2010.04.012
Zachos, J.C., Dickens, G.R., and Zeebe, R.E. 2008. An early Cenozoic perspective on greenhouse warming and carbon-cycle dynamics. Nature, 451:279-283.
https://doi.org/10.1038/nature06588
Zachos, J., Pagani, M., Sloan, L., Thomas, E., and Billups, K. 2001. Trends, rhythms, and aberrations in global climate 65 Ma to present. Science, 292:686-693.
https://doi.org/10.1126/science.1059412
Zhang, X., Sun, S., and Yong, S. 2007. Vegetation Map of the People’s Republic of China (1:1000000). Geological Publishing House, Beijing, China.
