

 Temperate to tropical palaeoclimates on the northwest margin of Europe during the middle Cenozoic
Temperate to tropical palaeoclimates on the northwest margin of Europe during the middle Cenozoic
Article number: 27.2.a43
https://doi.org/10.26879/1349
Copyright Palaeontological Association, August 2024
Author biographies
Plain-language and multi-lingual abstracts
PDF version
Appendices
Submission: 10 October 2023. Acceptance: 15 July 2024.
ABSTRACT
Globally, the middle Cenozoic (Oligocene to early Miocene, ~33.9–15.97 Ma) was characterized by a warmer, wetter climate than present. Reconstructing the climate of this stratigraphic interval helps us to better understand the implications of present and future anthropogenically-driven climate change in an Earth system with an established Antarctic ice mass and comparable pCO2 levels (400–700 ppm). Relative to mainland Europe, little palaeoclimate work has been done on the British Isles for this time interval. Compiled middle Cenozoic palynology records from across the British Isles were used to quantitatively reconstruct palaeoclimate, which was then used to define Köppen-Geiger signatures for each palynomorph assemblage. These reconstructions were used to show the presence of a temperate, dry-winter and hot-summer (Cwa) Köppen-Geiger climate type before 31.8 Ma, which was possibly a short-lived event driven by precessional (~26 k.y.) forcing. We attribute reconstructions with dry-winter Köppen-Geiger classifications to combined eccentricity-obliquity-precession (~405 k.y.) forcing, after the Eocene-Oligocene Transition. Declines in Mean Annual Temperature, in Chattian sections, are associated with the Svalbardella-2 and 3 North Sea cooling events. The late Oligocene warming event is shown to have produced tropical rainforest (Af) Köppen-Geiger classification types in the British Isles. Following early Miocene glaciation, a temperate, no-dry-season, warm-summer (Cfb) signature was reconstructed. We suggest the present-day climate of the northwest margin of Europe is comparable to the early Miocene palaeoclimate. Under increased pCO2 concentrations, based on projected twenty-first century anthropogenic warming scenarios, there is potential for wetter summers becoming more prevalent within the next century.
Jessica McCoy. Department of Engineering and Environmental Sciences, Northumbria University, Newcastle upon Tyne, UK. jessica.mccoy@northumbria.ac.uk
Martha E. Gibson. Department of Geology and Geography, West Virginia University, Morgantown, West Virginia, USA and PetroStrat, Parc Caer Seion, Conwy, UK. Martha.Gibson@petrostrat.com
Emma P. Hocking. Department of Engineering and Environmental Sciences, Northumbria University, Newcastle upon Tyne, UK. emma.hocking@northumbria.ac.uk
Jennifer M.K. O’Keefe. Department of Physics, Earth Science and Space Systems Engineering, Morehead State University, Morehead, Kentucky, USA. j.okeefe@moreheadstate.edu
James B. Riding. British Geological Survey, Keyworth, Nottingham, UK. jbri@bgs.ac.uk
Raymond Roberts. Natural Resources Wales, Chester Road, Flintshire, UK. raymond.roberts@cyfoethnaturiolcymru.gov.uk
Stewart Campbell. Natural Resources Wales, Chester Road, Flintshire UK. stewart.campbell@cyfoethnaturiolcymru.gov.uk
Geoffrey D. Abbott. School of Natural and Environmental Sciences, Newcastle University, Newcastle upon Tyne, UK. geoff.abbott@newcastle.ac.uk
Matthew J. Pound. Department of Engineering and Environmental Sciences, Northumbria University, Newcastle upon Tyne, UK. matthew.pound@northumbria.ac.uk
Key words: middle Cenozoic; British Isles; palaeobotany; palaeoclimate; palynology; Köppen-Geiger
Final citation: McCoy, Jessica, Gibson, Martha E., Hocking, Emma P., O’Keefe, Jennifer M.K., Riding, James B., Roberts, Raymond, Campbell, Stewart, Abbott, Geoffrey D., and Pound, Matthew J. 2024. Temperate to tropical palaeoclimates on the northwest margin of Europe during the middle Cenozoic. Palaeontologia Electronica, 27(2):a43.
https://doi.org/10.26879/1349
palaeo-electronica.org/content/2024/5275-mid-cenozoic-palaeoclimates-northwest-europe
Copyright: August 2024 Palaeontological Association.
This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
creativecommons.org/licenses/by/4.0
INTRODUCTION
Collapse of the Greenland ice sheet may occur within the next century, because of the weakening of ice shelf structure. This will produce a world with substantial ice sheets in the southern hemisphere only (Pattyn et al., 2018; Boers and Rypdal, 2021; Armstrong McKay et al., 2022; IPCC, 2022). The middle Cenozoic (Oligocene–early Miocene) was the last stratigraphic interval that had substantial ice solely in Antarctica (Wade and Pälike, 2004; Armstrong McKay et al., 2022). Following the formation of substantial ice in Antarctica during the Eocene–Oligocene Transition (Hutchinson et al., 2021), the Oligocene was an interval of fluctuating temperatures that included several warming intervals (such as the late Oligocene warming) and culminated in a major glaciation event around the Oligocene–Miocene boundary (Miller et al., 1991; Zachos et al., 1997; Wilson et al., 2008; Woods et al., 2014; Greenop et al., 2019).
There is a discrepancy in palaeoclimate reconstructions of the northwest margin of Europe during the Oligocene–Miocene time interval (Eldrett et al., 2009). The late Oligocene warming event (~25–24 Ma), in the North Atlantic marine record, corresponds with a reconstructed decline in pCO2 (~450–200 ppm; Zachos et al., 2001a; O’Brien et al., 2020). However, this has not been resolved with the known role of atmospheric pCO2 in global temperature change (Zachos et al., 2001a; Crowley and Berner, 2001; Scheffer et al., 2006). Here we provide new palaeoclimate reconstructions for the Oligocene to early Miocene from the United Kingdom and Ireland to document changes in terrestrial palaeoclimates during this interval. From the resulting reconstruction we present the magnitude of warming associated with the early Oligocene (Rupelian, 33.9–27.82 Ma) and the late Oligocene warming events, as well as the impacts of Early Miocene glaciation on this oceanographically sensitive terrestrial region.
BRIEF OVERVIEW OF PALAEOCLIMATES FROM THE EOCENE-OLIGOCENE TRANSITION TO THE EARLY MIOCENE
The Eocene–Oligocene Transition (circa 33.9 Ma) resulted in a global stepwise transition from greenhouse to icehouse conditions, within 0.4 Ma (Lear et al., 2000; Zachos et al., 2001a; Zachos and Kump, 2005; Katz et al., 2008; Zanazzi et al., 2007; Liu et al., 2009; Pound and Salzmann, 2017; Straume et al., 2022). A weakening of the North Atlantic overturning circulation and reductions in pCO2 (1000–400 ppm, O’Brien et al., 2020) led to reductions in temperatures in the Northern Hemisphere (Straume et al., 2022). A southward expansion of the Hadley Cell led to intensified tropical-rain regimes and temperatures (where winter temperatures exceeded 20°C) to the Southern Hemisphere, causing an intensification of the North Atlantic circulation. This in turn resulted in Northern Hemispheric cooling (Straume et al., 2022). The Eocene–Oligocene Transition cooling, associated with the positive (>1 ‰) δ18O excursion from the deep sea benthic foraminiferal record (Miller et al., 1991; Lear et al., 2000; Coxall et al., 2005; Wade et al., 2012; Brzelinski et al., 2023), led to the establishment of the present-day Antarctic ice sheet (Kennett and Shackleton, 1976; Ehrmann and Mackensen, 1992; Ivany et al., 2006; Galeotti et al., 2016). Other Eocene–Oligocene Transition-timed cooling-drivers included eustatic sea-level fall (Katz et al., 2008; Ladant et al., 2014; Miller et al., 2020); tectonics (Dupont-Nivet et al., 2007; Karagaranbafghi and Neubauer, 2018), and the opening of the Tasmanian Gateway, which strengthened the developing Antarctic Circumpolar Current (Kennett, 1977; Pound and Salzmann, 2017). Eocene–Oligocene Transition-driven cooling, particularly in high latitudes (Liu et al., 2009) contributed to increased Northern Hemispheric seasonality, with a reduction in high-latitude winter temperatures by 4–5°C (Ivany et al., 2000; Eldrett et al., 2009) and a northward shift of the Intertropical Convergence Zone (Hyeong et al., 2016; Straume et al., 2022).
Köppen-Geiger signatures assigned to Rupelian assemblages, from Central Europe, suggest that there was no overall dry-season (Pross et al., 1998, 2001; Kovar-Eder, 2016; Maxwell et al., 2016). Dry-winter Köppen-Geiger climate types, driven by the Asian monsoon system, likely developed throughout the Eocene in India (Shukla et al., 2014) and remained prevalent in Asian regions throughout the Oligocene (Srivastava et al., 2012; Miao et al., 2013; Lin et al., 2015; Farnsworth et al., 2019; Bhatia et al., 2021; Ling et al., 2021; Li et al., 2023). However, the palaeo spread of Podocarpium, often attributed to monsoonal-climate conditions, moved into Europe throughout the Cenozoic (Han et al., 2020). We suggest that the summer-monsoonal system may have been established sometime in Oligocene Europe, particularly in the Rupelian, where the Upper Rhine assemblage (Germany) reconstructed a temperate palaeoclimate with hot summers, and a slightly monsoonal signal (Cwb; Kovar-Eder, 2016).
From the onset of the Oligocene, there was a transition to a bipolar deep-water circulation, leading to the deep-water formation in the North Atlantic (Via and Thomas, 2006). This led to the development of a proto-Gulf Stream, which contributed to enhanced warming at high Northern Hemispheric latitudes (Via and Thomas, 2006).
Global cooling and periods of ice-sheet expansion and decline surrounding the Oligocene–Miocene Transition occurred in 100 k.y. cycles, likely driven by short-term eccentricity (100 k.y.), paced episodes (Zachos et al., 2001b; Lear et al., 2004; Pӓlike et al., 2006; Beddow et al., 2016; Liebrand et al., 2017; Greenop et al., 2019). Short-lived (~500 k.y.), orbitally forced glacial events, characterised by increases in the deep-sea foraminiferal δ18O record throughout the Oligocene and Miocene, were described as Oligocene isotope (Oi-) and Miocene isotope (Mi-) stages (Miller et al., 1991; Zachos et al., 2001b; Van Simaeys et al., 2005). Cooling events Oi-1a (~33.5 Ma, -1.3 %o) and Oi-2b, at the Rupelian–Chattian boundary (~27.82 Ma), and Mi-1 (~23.03 Ma) were driven by eccentricity minima (~405 k.y.) and low amplitude obliquity (1.2 m.y.) (Pӓlike et al., 2006; Jovane et al., 2006; Śliwińska and Heilmann-Clausen, 2011; Fenero et al., 2013). The influx of the Arctic-marker dinoflagellate cyst Svalbardella cooksoniae, in the North Atlantic, are associated with these cooling phases, and are labelled as Svalbardella events 3, 2, and 1, respectively (Van Simaeys et al., 2005; Śliwińska et al., 2010; Clausen et al., 2012; Śliwińska et al., 2014). Northern Hemispheric summer season warmth, throughout the Oligocene, was driven by Earth’s position at perihelion (De Vleeschouwer et al., 2017). The late Oligocene warming events in Central Europe occurred within an interval of increased late Oligocene (Chattian, 27.82–23.03 Ma) temperatures (Zachos et al., 2001a; De Man and Van Simaeys, 2004; Utescher et al., 2015; Moreno-Domínguez et al., 2021). The late Oligocene warming event was marked by the first appearance of Asterigerina-type foraminifera in North Atlantic assemblages, and by a rise in eustatic sea-level (Doppert and Neele, 1983; King, 1983, 1989; Van Simaeys, 2004).
A warming trend was interrupted by a return to global glacial conditions with the transition from the Oligocene–Miocene at 23.03 Ma (Zachos et al., 1997; Lear et al., 2000; Brzelinski et al., 2023). A positive excursion in benthic foraminiferal isotopic signatures suggested there was a reduction in mean global air temperatures, with a distinct gradient between the mid and high latitudes in air temperatures (Zachos et al., 2001b; Beddow et al., 2016; Liebrand et al., 2017; Torricelli et al., 2022). Following this cooling event, early Miocene European floral assemblages reconstructed well-drained riparian wetlands, with subtropical to tropical elements (Bessedik, 1984; Huguenery and Truc, 1976; Velitzelos et al., 2014). This Miocene warm palaeoclimate occurred when the summertime incoming solar radiation angle was at its maximum position (De Vleeschouwer et al., 2017).
To test whether these Central European reconstructions were consistent with the northwest margin of Europe, we set out to reconstruct the palaeoclimate of the British Isles, for the Oligocene to early Miocene. From our palaeoclimate reconstructions, we aim to assign Köppen-Geiger classes to study sites to illustrate palaeoclimate change during this interval of unipolar glaciation.
GEOLOGICAL SETTING OF THE BRITISH ISLES
The current climate of the British Isles (used here to refer to the islands that make up the countries of Ireland and the United Kingdom) is warm with relatively dry summer conditions. This climate is maintained by high pressure westerlies, and east-moving polar jet streams, which are strengthened between the winter to early spring months and create common fog and drizzle (Mayes, 1991, 1996; Hulme and Barrow, 1997; Hall et al., 2015; Methner et al., 2020). Westerlies were believed to have been established from the middle Miocene (Serravallian, 13.65–11.63 Ma) according to Quan et al. (2014) and Pound and Riding (2016). Given the relatively low topography of the northwest margin of Europe during the Miocene (Roberts, 1975), westerlies may have driven a stronger, more intense precipitation regime than those reconstructed for southern Europe (Henrot et al., 2010). The Gulf Stream contributes to the present-day warmth of the British Isles, and it regulates a mild winter regime; the Gulf Stream was established by at least the middle Miocene (Denk et al., 2013; Pound and McCoy, 2021). With the onset of anthropogenically-driven climate change, the future climate of the British Isles is expected to develop hotter summers and wetter winters (Pope et al., 2021).
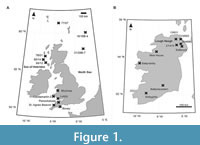 The British Isles have an abundance of onshore and offshore Oligocene siliciclastic deposits (Figure 1A-B). These are present in clay and lignite beds in the North Sea, the Republic of Ireland, Northern Ireland, Wales, and southern England (Figure 1A-B; Appendix 1). These units likely formed in shoreline environments, a result of the palaeo-shoreline being located ~75 km east during the Oligocene, relative to present-day (Murray, 1992; Walsh, 2001). Most of these strata lie along the Sticklepath-Lustleigh Fault that runs northwest from southwest England towards Northern Ireland and appears to have been a major influence on post-Eocene sediment accumulation (Tappin et al., 1994; King et al., 2016; Figure 1A-B). Therefore, the British Isles have a limited amount of onshore sediment of post-Eocene age available for palaeoclimate reconstruction. Offshore coring expeditions have retrieved abundant post-Eocene siliciclastic deposits (Wilson, 1996).
The British Isles have an abundance of onshore and offshore Oligocene siliciclastic deposits (Figure 1A-B). These are present in clay and lignite beds in the North Sea, the Republic of Ireland, Northern Ireland, Wales, and southern England (Figure 1A-B; Appendix 1). These units likely formed in shoreline environments, a result of the palaeo-shoreline being located ~75 km east during the Oligocene, relative to present-day (Murray, 1992; Walsh, 2001). Most of these strata lie along the Sticklepath-Lustleigh Fault that runs northwest from southwest England towards Northern Ireland and appears to have been a major influence on post-Eocene sediment accumulation (Tappin et al., 1994; King et al., 2016; Figure 1A-B). Therefore, the British Isles have a limited amount of onshore sediment of post-Eocene age available for palaeoclimate reconstruction. Offshore coring expeditions have retrieved abundant post-Eocene siliciclastic deposits (Wilson, 1996).
METHODS
Data Synthesis
 Seventy-eight published onshore and offshore Oligocene to early Miocene pollen assemblages (Figure 2), from the British Isles, were compiled into a presence-absence matrix (Appendix 2; King et al., 2016).
Seventy-eight published onshore and offshore Oligocene to early Miocene pollen assemblages (Figure 2), from the British Isles, were compiled into a presence-absence matrix (Appendix 2; King et al., 2016).
Details of each site, including coordinates, borehole names, lithology/sedimentology, and previous palynological reconstructions, are available in Appendix 1. The biostratigraphical ranges of dinoflagellate cysts, recovered from assemblages 16/16B-4, and 21/28B-7 by Wilson (1996), were reviewed and updated to reflect current biostratigraphical correlations (Wilson, 1996; Egger et al., 2016; King et al., 2016) of the concerned dinoflagellate cysts. Fossil form names were synonymised, where possible, following Stuchlik et al. (2001, 2002, 2009, 2014) and Utescher et al. (2024) (Appendix 3). Due to the morphological similarities between pollen-types belonging to the Cupressaceae family (Bortenschlager, 1990; Bouchal and Denk, 2020), genera belonging to the Cupressaceae family (e.g., Sequoiapollenites) were assigned “Cupressaceae” for their NLR.
The relict taxon Sciadopitys-type was removed from reconstructions. This is because their modern biogeography is limited in comparison to their Cenozoic environmental distribution, which was much broader (Mosbrugger et al., 1994; Figueiral et al., 1999; Utescher et al., 2014).
Palaeoclimate Reconstruction
The Climate Reconstruction SofTware (CREST, crestr R package) probability density function-based (pdf) model was used to reconstruct climate variables, based on the present-day distributions of contemporary taxa and the corresponding climatic tolerance ranges of assigned nearest-living relatives of pollen (Table 1; Chevalier et al., 2014; Chevalier, 2019; Chevalier, 2022). Modern floral distribution data was taken from the Global Biodiversity Information Facility (GBIF, 2020a-g). CREST-based outputs delivered an optimised value, based on the WorldClim2 climate dataset (Fick and Hijmans, 2017) with 50% and 95% uncertainty ellipses defined (Chevalier et al., 2014).
Full details of the CREST model, R code, and background can be found in Chevalier et al. (2014) and Chevalier (2022). To assign Köppen-Geiger climate-types to assemblages, CREST was used to reconstruct the Mean Annual Temperature (MAT), Mean Temperature of the Warmest Quarter (MTWQ), Mean Temperature of the Coldest Quarter (MTCQ), Mean Annual Precipitation (MAP), Precipitation of the Driest Quarter (MPDry); Precipitation of the Warmest Quarter and Precipitation of the Coldest Quarter (Fick and Hijmans, 2017).
Assignment of Köppen-Geiger classes, using output variables from CREST was adapted from the Köppen-Geiger assignment method (Beck et al., 2018). Tropical rainforest signatures (Af) were assigned to sites where MTCQ reconstructions exceeded 18°C, and where seasonal MAP, minus the Precipitation of the Driest Quarter, gave a positive value, thus indicating no dry season. Assemblages which reconstructed MTCQ values between 0–18°C were assigned a subtropical-temperate climate signature (C). Palaeoclimate types with a dry season were determined by differences between the Precipitation of the Warmest Quarter and the Precipitation of the Coldest Quarter, and their comparison to the annual rainfall, as adapted by Beck et al. (2018) (Table 2). Where reconstructions of the Precipitation of the Warmest Quarter exceeded the Precipitation of the Coldest Quarter by 10 mm, a dry-winter signal was assigned (Cw). Where the difference between the Precipitation of the Coldest and Warmest Quarter reconstructions did not exceed 10 mm, reconstructions were assigned a no-dry-season signal (Cf). Subtropical signatures were assigned hot summer (– a) and warm summer (–b) signals, where MTWQ reconstructions were >22°C and 12–22°C, respectively.
MPDry reconstructions were unsuccessful where particular families were included, i.e., Anacardiaceae; Asteraceae; Caprifoliaceae; Fabaceae; Lythraceae; Malvaceae; Poaceae and Solanaceae (Table 1). These taxa were removed from MPDry model runs. This was because their tolerance ranges, regarding MPDry, included 0 and so this resulted in a zero-based error being carried forward throughout model runs, resulted in an impossible division of a value by 0. As a result of the removal of these families, MPDry reconstructions may have been skewed towards wetter reconstructions.
RESULTS
Palaeoclimate Reconstruction
In text, palaeoclimate reconstructions are presented with their reconstructed optima, followed by their largest 50% confidence interval, marked after “±”. Model results, with 50% uncertainty ranges with differences from optima, are provided in Appendix 4.
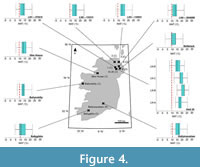
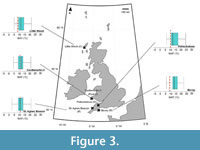 Mean annual temperature. Rupelian Mean Annual Temperature (MAT) reconstructions were lowest in the southernmost localities, i.e., the St. Agnes Beacon and Bovey Basin, where the optimal MAT reconstruction was 13.37 ±1.51°C (Figure 3). The Ballymacadam Rupelian assemblage reconstructed a higher MAT value of 16.38°C ±1.51°C (Figure 4). We note that overlaps in 50% statistical uncertainties occurred in reconstructions of 16/16B-4 (associations A–B); 88/12 (associations A–B) and 73/36 (D–E), other discussed MAT reconstructions throughout sequences below showed no overlap, and therefore showed statistically significant change (Appendix 4).
Mean annual temperature. Rupelian Mean Annual Temperature (MAT) reconstructions were lowest in the southernmost localities, i.e., the St. Agnes Beacon and Bovey Basin, where the optimal MAT reconstruction was 13.37 ±1.51°C (Figure 3). The Ballymacadam Rupelian assemblage reconstructed a higher MAT value of 16.38°C ±1.51°C (Figure 4). We note that overlaps in 50% statistical uncertainties occurred in reconstructions of 16/16B-4 (associations A–B); 88/12 (associations A–B) and 73/36 (D–E), other discussed MAT reconstructions throughout sequences below showed no overlap, and therefore showed statistically significant change (Appendix 4).
 Between associations A–B of site 73/36 (>31.8 Ma), reconstructed MATs increase from 13.37 (±1.51)–20.91 (±1.51)°C; no other MAT increases are reported from other Rupelian reconstructions (Figure 5). Between associations A–B, of well 16/16B-4 there is a reduction in MAT, from 19.40 (±1.51)–17.89 (±1.51)°C, where associations are assigned to 32.1–31.7 Ma (Figure 5). Significant, non-overlapping cooling from 20.91 (±1.51)–14.87 (±1.51)°C occurred from 32.1–31.7 Ma at site 21/28B-7, as shown by reconstructions from associations B–C of well 21/28B-7 (Figure 5).
Between associations A–B of site 73/36 (>31.8 Ma), reconstructed MATs increase from 13.37 (±1.51)–20.91 (±1.51)°C; no other MAT increases are reported from other Rupelian reconstructions (Figure 5). Between associations A–B, of well 16/16B-4 there is a reduction in MAT, from 19.40 (±1.51)–17.89 (±1.51)°C, where associations are assigned to 32.1–31.7 Ma (Figure 5). Significant, non-overlapping cooling from 20.91 (±1.51)–14.87 (±1.51)°C occurred from 32.1–31.7 Ma at site 21/28B-7, as shown by reconstructions from associations B–C of well 21/28B-7 (Figure 5). 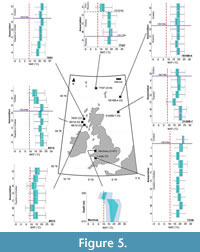 Similar reductions in MAT were reconstructed using assemblages from: associations A–B of site 88/12, (14.87 [±1.51]–11.86 [±1.51]°C, deposited >31.5 Ma); associations A–C of borehole 80/14, (22.41 [±0.00]–19.40 [±1.51]°C, deposited <31.8 Ma), and from associations D–E of borehole 73/36 (19.40 [±1.51]–17.89 [±0.00]°C, deposited <31.8 Ma) (Figure 5). Associations A–B of site 77/07 did not reconstruct cooling during this time (Figure 5).
Similar reductions in MAT were reconstructed using assemblages from: associations A–B of site 88/12, (14.87 [±1.51]–11.86 [±1.51]°C, deposited >31.5 Ma); associations A–C of borehole 80/14, (22.41 [±0.00]–19.40 [±1.51]°C, deposited <31.8 Ma), and from associations D–E of borehole 73/36 (19.40 [±1.51]–17.89 [±0.00]°C, deposited <31.8 Ma) (Figure 5). Associations A–B of site 77/07 did not reconstruct cooling during this time (Figure 5).
All Chattian MAT reconstructions exceed their present-day MAT values (Figure 3, Figure 4, Figure 5; Table 2). MAT reconstructions surrounding Lough Neagh produce similar optimal values, ranging from 13.37–14.87°C in boreholes 13/603, 13/611, 27/415, and 36/4680 (Figure 4). Associations B–D of Well-28 reconstructed a decline in MAT (22.42 [±0.00]–14.87 [±1.51]°C) that occurred between ~31.65–25.8 Ma (Figure 4). MAT reconstructions from associations A–G of 78/01 suggest MAT increased from 17.89 [±1.51]–22.41 [±1.51]°C without interruption between 33.9–24.8 Ma (Figure 5).
The Mochras Basal Red Unit reconstructed an increase in MAT, from 17.89 ±1.51°C to 23.92 ±1.51°C at depths 515.42 m and 408.43 m, respectively (Figure 5). Between the Mochras Basal Red Unit (408.43 m) and the Transitional Unit (368.80 m), reconstructions indicate a decline in MAT from 23.92 ±1.51°C to 14.87 ±1.51°C (Figure 5). Reconstructed MAT decline to 8.84 ±0.00°C at depth 368.20 m, then increase to 11.86 ±1.51°C at depth 324.61 m (Figure 5). The assemblage from Association E of 77/07 (<23.03 Ma) and the Castlemartin-3 assemblage (<23.03 Ma) reconstruct MATs of 11.86 (± 1.51)°C (Figure 3).
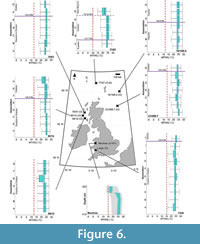 Mean temperature of the warmest quarter. All Chattian MTWQ reconstructions exceed the variables of their present-day location (Table 2). Sequences show no change to their optima reconstructions, from their Rupelian counterparts (Figure 6). Of the discussed changes in MTWQ below, we acknowledge overlap in uncertainty ranges in studied depths of the Mochras borehole, from in 77/07 (associations D–E).
Mean temperature of the warmest quarter. All Chattian MTWQ reconstructions exceed the variables of their present-day location (Table 2). Sequences show no change to their optima reconstructions, from their Rupelian counterparts (Figure 6). Of the discussed changes in MTWQ below, we acknowledge overlap in uncertainty ranges in studied depths of the Mochras borehole, from in 77/07 (associations D–E).
Chattian MTWQ reconstructions from associations C–D of borehole 77/07 remain unchanged from 24.62 (±0.00–1.41)°C prior to 37.1 Ma (Figure 6). MTWQ optimal reconstructed from the borehole 73/35 and borehole 73/58 assemblages overlap with the reconstructed optima from association A of borehole 73/36 with overlap occurring between, 33.9–31.8 Ma (Figure 6). Depths 515.42–408.43 m of the Mochras Basal Red Unit reconstructed a shared MTWQ of 26.03 ±0.00°C (Figure 6). Between the Basal Red Unit and the Transitional Unit of the Mochras borehole, reconstructed MTWQ decline to 24.62 ±1.41°C (Figure 6). MTWQ declines to 16.18 ±1.41°C at depth 368.20 m of the Transitional Unit (Figure 6). At depth 324.61 m, reconstructed MTWQ increase to 17.59 ±1.41°C (Figure 6). Reconstructed MTWQ declined from 24.62 ±1.41°C to 21.81 ±1.41°C between Association D–E of borehole 77/07 (Chattian to early Miocene) (Figure 6).
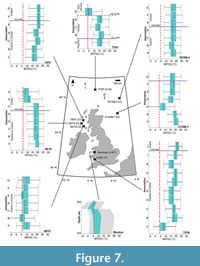 Mean temperature of the coldest quarter. At >31.8 Ma, Rupelian assemblages from associations A–B of borehole 80/14 reconstructs a decline in MTCQ from 21.00 (±0.00)–19.40 (±1.61)°C (Figure 7). Associations A–B of borehole 88/12, reconstruct a decline in MTCQ from 9.74 (±3.21)–4.92 (±1.61)°C (Figure 7). Associations B–C of well 21/28B-7 (~32.1–32 Ma) reconstruct a substantial decline in MTCQ values from 19.40 (±1.61)–8.14 (±1.61)°C; after association C (<32 Ma) MTCQ values increase to 16.18 (±1.61)°C, in association D (Figure 7). Overlap between 50% uncertainty ranges were observed in 80/14 (associations A–B), 88/12 (associations A–B), 77/07 (associations B–C) and between depths in the Mochras borehole (Appendix 4).
Mean temperature of the coldest quarter. At >31.8 Ma, Rupelian assemblages from associations A–B of borehole 80/14 reconstructs a decline in MTCQ from 21.00 (±0.00)–19.40 (±1.61)°C (Figure 7). Associations A–B of borehole 88/12, reconstruct a decline in MTCQ from 9.74 (±3.21)–4.92 (±1.61)°C (Figure 7). Associations B–C of well 21/28B-7 (~32.1–32 Ma) reconstruct a substantial decline in MTCQ values from 19.40 (±1.61)–8.14 (±1.61)°C; after association C (<32 Ma) MTCQ values increase to 16.18 (±1.61)°C, in association D (Figure 7). Overlap between 50% uncertainty ranges were observed in 80/14 (associations A–B), 88/12 (associations A–B), 77/07 (associations B–C) and between depths in the Mochras borehole (Appendix 4).
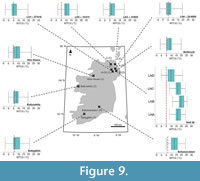
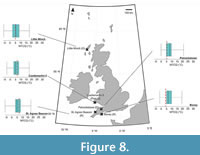 Coldest MTCQ reconstructions were noted for the reconstruction of the southernmost sites: the St. Agnes Beacon (4.92 ±1.60°C) and the Bovey Basin (6.53 ±1.61°C) (Figure 8). Assemblages from boreholes 13/603 and 36/4680 both reconstructed MTCQ values of 6.53 ±1.61°C, respectively (Figure 9). Assemblages from boreholes 27/415, 13/603, and 36/4680 reconstructed respective MTCQ values of 4.92 ± 1.60°C, 6.53 ± 1.61°C and 6.53 ±1.61°C (Figure 9). Well-28 reconstructed a step-like decline in reconstructed MTCQ, from 19.40 (±1.61) –9.75 (±3.22)°C between Associations B–D (Figure 9). Declines in MTCQ reconstructions were also seen in reconstructions taken from assemblages B–C of borehole 77/07 (31.7–26.8 Ma) where MTCQ temperatures declined from 16.18 (±1.61)–14.57 (±1.61)°C (Figure 7). Reconstructed MTCQ values decline from 19.40 (±3.21)–8.14 (1.61)°C for associations F–H of 80/14 (25.8–25.5 Ma) (Figure 7).
Coldest MTCQ reconstructions were noted for the reconstruction of the southernmost sites: the St. Agnes Beacon (4.92 ±1.60°C) and the Bovey Basin (6.53 ±1.61°C) (Figure 8). Assemblages from boreholes 13/603 and 36/4680 both reconstructed MTCQ values of 6.53 ±1.61°C, respectively (Figure 9). Assemblages from boreholes 27/415, 13/603, and 36/4680 reconstructed respective MTCQ values of 4.92 ± 1.60°C, 6.53 ± 1.61°C and 6.53 ±1.61°C (Figure 9). Well-28 reconstructed a step-like decline in reconstructed MTCQ, from 19.40 (±1.61) –9.75 (±3.22)°C between Associations B–D (Figure 9). Declines in MTCQ reconstructions were also seen in reconstructions taken from assemblages B–C of borehole 77/07 (31.7–26.8 Ma) where MTCQ temperatures declined from 16.18 (±1.61)–14.57 (±1.61)°C (Figure 7). Reconstructed MTCQ values decline from 19.40 (±3.21)–8.14 (1.61)°C for associations F–H of 80/14 (25.8–25.5 Ma) (Figure 7).
 Lowest early Miocene MTCQ optimal reconstructions of 4.92 (±3.21)°C and 4.92 (±1.60)°C and 4.92 ±1.61°C were taken from the respective assemblages from Association E of borehole 77/07, the Castlemartin-3 borehole, and from depths 73–62.5 m of Little Minch (Figure 7–Figure 8; Table 2). A significant decline in optimal values from 16.18 (±1.61)°C to 4.92 (±3.22)°C from borehole 77/07 at Associations D–E (26.8–23.03 Ma) was reconstructed (Figure 7).
Lowest early Miocene MTCQ optimal reconstructions of 4.92 (±3.21)°C and 4.92 (±1.60)°C and 4.92 ±1.61°C were taken from the respective assemblages from Association E of borehole 77/07, the Castlemartin-3 borehole, and from depths 73–62.5 m of Little Minch (Figure 7–Figure 8; Table 2). A significant decline in optimal values from 16.18 (±1.61)°C to 4.92 (±3.22)°C from borehole 77/07 at Associations D–E (26.8–23.03 Ma) was reconstructed (Figure 7).
The Mochras Basal Red Unit reconstructs MTCQ values of 9.75 (±1.61)°C and 9.75 (±3.22)°C at depths 515.42 m and 408.43 m, respectively (Figure 7). MTCQ values decline to 6.53 (±3.21)°C at the Transitional Unit (368.80 m) then MTCQ reconstructions decline to 3.32 (±1.61)°C (at 368.20 m) (Figure 7). MTCQ reconstructions increase to 6.53 ±1.61°C (at 324.61 m) (Figure 7).
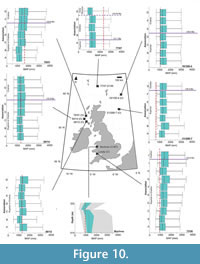 Mean annual precipitation. Overlaps in uncertainties are recorded in reconstructions, which compare 16/16B–4 (associations A–B); 80/14 (associations A–B); 77/07 (associations B–C); 78/01 (associations A–F); and between depths 515.42–408.43 m in the Mochras borehole (Appendix 4). The assemblages from associations A–B of well 21/28B-7 reconstruct a minor increase in MAP from 1195 (±227)–1422 (±284) mm (~33–32 Ma). MAP declines from 1308 [±228]–1251 [±284] mm, between associations A–B at site 80/14 (~31.8–31.5 Ma, Figure 10) A minor decline in MAP is reconstructed (1251 [±228]–1195 [±228] mm) between associations B–C of borehole 77/07 (31.7–26.8 Ma) (Figure 10). Associations A–F of borehole 78/01 reconstructed a minor increase in MAP (from associations A–F) from 1138 (±228)–1536 (±398) mm at >24.8 Ma (Figure 10). Association E of borehole 77/07 reconstructs a decline in MAP (to 1081±227 mm) (Figure 10).
Mean annual precipitation. Overlaps in uncertainties are recorded in reconstructions, which compare 16/16B–4 (associations A–B); 80/14 (associations A–B); 77/07 (associations B–C); 78/01 (associations A–F); and between depths 515.42–408.43 m in the Mochras borehole (Appendix 4). The assemblages from associations A–B of well 21/28B-7 reconstruct a minor increase in MAP from 1195 (±227)–1422 (±284) mm (~33–32 Ma). MAP declines from 1308 [±228]–1251 [±284] mm, between associations A–B at site 80/14 (~31.8–31.5 Ma, Figure 10) A minor decline in MAP is reconstructed (1251 [±228]–1195 [±228] mm) between associations B–C of borehole 77/07 (31.7–26.8 Ma) (Figure 10). Associations A–F of borehole 78/01 reconstructed a minor increase in MAP (from associations A–F) from 1138 (±228)–1536 (±398) mm at >24.8 Ma (Figure 10). Association E of borehole 77/07 reconstructs a decline in MAP (to 1081±227 mm) (Figure 10).
The Mochras Basal Red Unit reconstructs respective MAP values of 1308 ±171 mm and 1308 ±341 mm for depths 515.42 m and 408.43 m (Figure 10). MAP reconstructions decrease from 1138 ±228 mm to 739 ±114 mm (between depths 368.80–368.20 m) before then increasing to 1251 ±284 mm (324.61 m) (Figure 10).
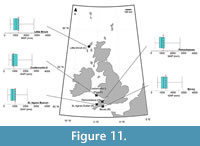 Higher MAP values, than those reconstructed from the Mochras borehole, were reconstructed from the Beacon Cottage Farm Outlier section of St. Agnes Beacon (796 mm ± 170 mm) to the Bovey Basin (1138 ±228 mm) to the Ballymacadam (1195 ±285 mm) study site (Figure 11–Figure 12).
Higher MAP values, than those reconstructed from the Mochras borehole, were reconstructed from the Beacon Cottage Farm Outlier section of St. Agnes Beacon (796 mm ± 170 mm) to the Bovey Basin (1138 ±228 mm) to the Ballymacadam (1195 ±285 mm) study site (Figure 11–Figure 12).
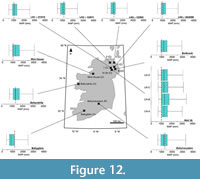 Mean precipitation of the driest month. Of discussed changes in MPDry reconstructions, we highlight that overlaps in uncertainties are recorded in 80/14 (associations A–C) and in 16/16B-4 (associations C–D) (Appendix 4). Associations A–C of well 21/28B-7 reconstructs a decline in MPDry from 68 (±11)–42 (±8) mm (~33–32 Ma) (Figure 13). Reconstructed MPDry from borehole 80/14 (associations A–C) reconstructs an increase in MPDry from 48 (±6)–54 (±8) mm (>31.8 Ma); reconstructions increase into site 80/14’s Chattian associations (Figure 13). Similar MPDry reconstructions, 42 (±8)–62 (±11) mm and 48 (±8)–56 (±11) mm, are reconstructed from associations C–F of well 21/28B–7 (~32–31.05 Ma) and for associations C–D of well 16/16B–4 (31.7–31.05 Ma), respectively (Figure 13).
Mean precipitation of the driest month. Of discussed changes in MPDry reconstructions, we highlight that overlaps in uncertainties are recorded in 80/14 (associations A–C) and in 16/16B-4 (associations C–D) (Appendix 4). Associations A–C of well 21/28B-7 reconstructs a decline in MPDry from 68 (±11)–42 (±8) mm (~33–32 Ma) (Figure 13). Reconstructed MPDry from borehole 80/14 (associations A–C) reconstructs an increase in MPDry from 48 (±6)–54 (±8) mm (>31.8 Ma); reconstructions increase into site 80/14’s Chattian associations (Figure 13). Similar MPDry reconstructions, 42 (±8)–62 (±11) mm and 48 (±8)–56 (±11) mm, are reconstructed from associations C–F of well 21/28B–7 (~32–31.05 Ma) and for associations C–D of well 16/16B–4 (31.7–31.05 Ma), respectively (Figure 13).
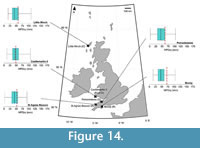
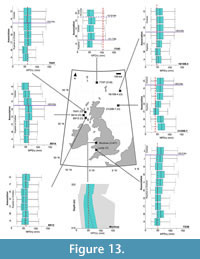 Early Miocene assemblages reconstruct MPDry values that range from 45 (±11) mm to 48 (±9) mm, based on respective reconstructions from the Castlemartin-3 and the 77/07 (Association E) boreholes (Figure 14). Association E of the 77/07 borehole, in comparison to its earlier-dated reconstructions in associations A–D, reconstructs a decline in MPDry to 48 ±9 mm (Figure 13).
Early Miocene assemblages reconstruct MPDry values that range from 45 (±11) mm to 48 (±9) mm, based on respective reconstructions from the Castlemartin-3 and the 77/07 (Association E) boreholes (Figure 14). Association E of the 77/07 borehole, in comparison to its earlier-dated reconstructions in associations A–D, reconstructs a decline in MPDry to 48 ±9 mm (Figure 13).
Tropical Taxa-Types
 Eight taxa-types classified to genus level, with subtropical-tropical signatures were present. Of these, Cycas is the sole conifer-type; the remaining seven genera are angiosperms (Figure 15). Anacolosa-types were not present in the northwest European palaeovegetation record during the Rupelian, whereas all other taxa-types were present (Figure 15). All tropical taxa-types were likely to have been present throughout the Chattian, and the warming events throughout the late Oligocene (Figure 15). Anacolosa-types were only recorded from the late Oligocene warming event to the Oligocene–Miocene Transition whereas Calamus-types were not recorded past the Rupelian (Figure 15). Following the LOWE, Mangifera-types were not recorded in the palaeovegetation record; Anacardiaceae, Brownlowia, and Cupania-types had an inferred relative abundance prior to the Oligocene–Miocene Transition (Figure 15). No tropical taxa were recorded in assemblages deposited throughout the early Miocene or post-Oligocene–Miocene Transition (>23.03 Ma) (Figure 15; Appendix 2).
Eight taxa-types classified to genus level, with subtropical-tropical signatures were present. Of these, Cycas is the sole conifer-type; the remaining seven genera are angiosperms (Figure 15). Anacolosa-types were not present in the northwest European palaeovegetation record during the Rupelian, whereas all other taxa-types were present (Figure 15). All tropical taxa-types were likely to have been present throughout the Chattian, and the warming events throughout the late Oligocene (Figure 15). Anacolosa-types were only recorded from the late Oligocene warming event to the Oligocene–Miocene Transition whereas Calamus-types were not recorded past the Rupelian (Figure 15). Following the LOWE, Mangifera-types were not recorded in the palaeovegetation record; Anacardiaceae, Brownlowia, and Cupania-types had an inferred relative abundance prior to the Oligocene–Miocene Transition (Figure 15). No tropical taxa were recorded in assemblages deposited throughout the early Miocene or post-Oligocene–Miocene Transition (>23.03 Ma) (Figure 15; Appendix 2).
Köppen-Geiger Classifications
 Köppen-Geiger classifications were defined from CREST-modelled optimised value reconstructions and ranged from temperate wet and warm summer types (Cfb) to tropical rainforest types (Af) (Table 2; Figure 16A–B). This range of Köppen-Geiger classifications suggests the palaeoclimate of the early Oligocene (Rupelian) to early Miocene of the northwest margin of Europe ranged from subtropical with dry winters and warm summers to tropical. This is notably different from the present-day temperate climate, which has warm summers (Cfb) across the British Isles and cold summers (Cfc) with more consistent rainfall throughout the year in the North Sea Region (Peel et al., 2007; Beck et al., 2018). Köppen-Geiger classification assignments are listed in Table 2 and presented in Figure 16A–B.
Köppen-Geiger classifications were defined from CREST-modelled optimised value reconstructions and ranged from temperate wet and warm summer types (Cfb) to tropical rainforest types (Af) (Table 2; Figure 16A–B). This range of Köppen-Geiger classifications suggests the palaeoclimate of the early Oligocene (Rupelian) to early Miocene of the northwest margin of Europe ranged from subtropical with dry winters and warm summers to tropical. This is notably different from the present-day temperate climate, which has warm summers (Cfb) across the British Isles and cold summers (Cfc) with more consistent rainfall throughout the year in the North Sea Region (Peel et al., 2007; Beck et al., 2018). Köppen-Geiger classification assignments are listed in Table 2 and presented in Figure 16A–B.
 References to assigned Köppen-Geiger types throughout this section, unless otherwise stated, refer to Table 2. The St. Agnes Beacon was the only Rupelian site to be assigned a subtropical-warm summer climate-type (Cfb). Association C of the 73/36 borehole (33.9-31.8 Ma), and depths 515.42–408.43 m of the Mochras borehole are assigned a temperate climate signature, with dry winters and hot summers (Cwa). The Petrockstowe Basin assemblage was assigned a temperate, dry-winter, warm-summer signal (Cwb). A tropical rainforest signature (Af) was assigned to the Rupelian reconstruction of associations A–B of borehole 80/14 and association B of well 21/28B-7.
References to assigned Köppen-Geiger types throughout this section, unless otherwise stated, refer to Table 2. The St. Agnes Beacon was the only Rupelian site to be assigned a subtropical-warm summer climate-type (Cfb). Association C of the 73/36 borehole (33.9-31.8 Ma), and depths 515.42–408.43 m of the Mochras borehole are assigned a temperate climate signature, with dry winters and hot summers (Cwa). The Petrockstowe Basin assemblage was assigned a temperate, dry-winter, warm-summer signal (Cwb). A tropical rainforest signature (Af) was assigned to the Rupelian reconstruction of associations A–B of borehole 80/14 and association B of well 21/28B-7.
Tropical rainforest Köppen-Geiger signatures were assigned to Rupelian–Chattian associations E–G of 78/01 and G and J of 73/36 (Figure 16A). A subtropical warm summer climate-type was assigned to the assemblage from association B of borehole 88/12. All other Rupelian, Rupelian–Chattian, and Chattian assemblages were assigned a temperate climate-type with no dry season and hot summers (Cfa). Tropical rainforest signatures were assigned to Chattian assemblages from associations E–G of 78/01, The Little Minch assemblage was assigned a temperate warm-summer (Cfa) Köppen-Geiger signature.
All early Miocene assemblages were assigned a temperate climate with warm-summers signature. Depths of the Mochras borehole (i.e., 515.42–408.43 m, 368.80 m and 368.20–324.61 m) were assigned the respective Köppen-Geiger classifications of Cwa, Cfa, and Cfb (Table 2).
DISCUSSION
Our results show that the northwest margin of Europe was warmer throughout the Oligocene than palaeoclimates of Miocene (Figure 3, Figure 4, Figure 5, Figure 6, Figure 7, Figure 8, Figure 9 and Figure 15–Figure 16). Most Rupelian and Chattian-dated reconstructions were assigned a Köppen-Geiger classification of either a temperate palaeoclimate with hot summers, and no dry season (Cfa), or a tropical rainforest Köppen-Geiger signature, with no outstanding dry season (Af) (Figure 16A–B). Selected Chattian, if not possibly younger, samples from the Little Minch Basin (Evans et al., 1991) reconstructed a temperate palaeoclimate with warm summers (Cfb)- comparable to the present-day climate of the British Isles (Table 2). Reductions in summer temperatures, resulted in alternating warm and hot summer-types (Cfa-Cfb) between associations B-H of 80/14 and from the samples taken from the same 80/14 borehole, from Evans et al (1991), see Table 2. These alternations between hot and warm summer reconstructions between the two studies be associated with the pulses of interglacials, due to low eccentricity forcing (Pӓlike et al., 2006; Levy et al., 2019).
Tropical Rainforest Signals at 57.77°N
Forests in the Rupelian, and maybe earlier. Tropical rainforest Köppen-Geiger classifications (Af) are assigned to the assemblages from association A of 80/14, which was dated to a minimum age of 31.8 Ma, based on the temporal zonation of recorded dinoflagellate cysts (Wilson, 1996; Egger et al., 2016; King et al., 2016; Table 2). Our reconstructions suggest tropical rainforests were present during the Rupelian, if not earlier, at 57.77°N (Table 2; Figure 16A–B). However, this disagrees with Rupelian biome reconstructions of temperate mixed forests and conifer forests (Pound and Salzmann 2017). Assigning a late Eocene age, rather than Wilson’s (1996) previously assigned “Oligocene” age, for these assemblages, from association A of borehole 80/14, would allow our tropical reconstructions to be comparable with those from the Isle of Wight (latest Eocene) (Collinson and Hooker, 1987; Quicke, 1987; McCobb et al., 1998; Daley, 1989; Antropov et al., 2014; Krzemiński et al., 2019; Gorochov, 2019) amongst other tropical, western European Eocene reconstructions (Blondel, 2001). Warmer conditions are also correlative with high eustatic sea-levels in the Priabonian (Haq et al., 1987; Cande and Kent, 1995; Katz et al., 2008; Ladant et al., 2014).
A tropical rainforest-type is also reconstructed from the 21/28B-7 association A assemblage, the middle of which is dated to ~33–32.1 Ma (Egger et al., 2016; Figure 16; Table 2). Our reconstructions may suggest that the European tropical vegetation belt extended ~17.82° further north than previously proposed (Utescher et al., 2021). We suggest the possibility of the 21/28B-7 is likely older, given the middle of the assemblage was deposited within the timeframe of the Oi-2 cooling event, at 32.5 Ma, and so temperatures would have cooled, rather than have become more tropical (Wade and Pälike, 2004; Van Simaeys et al., 2005), furthermore, corresponding pCO2 levels were in decline (O’Brien et al., 2020; Zachos et al., 2001a). Should the assemblages from association B of 21/28B-7, note there was no overlap with the 50 % uncertainties between these depths, and so statistically significant changes occur outside the uncertainty, the transition from a tropical rainforest to a temperate hot-summer palaeoclimate, in later-deposited associations reconstructs a significant cooling event between possible Eocene and Oligocene horizons (Liu et al., 2009; Hutchinson et al., 2021).
Declines in MAT between associations A–C of 80/14 (3.01°C) and between associations B–C of 21/28B-7 (6.03°C), these reconstructed ranges fell outside of uncertainty ranges and so are consistent with ~4–6°C declines in annual air temperature reconstructions from the northwest margin of Europe at the Eocene–Oligocene Transition (Hren et al., 2013; Hutchinson et al., 2021; Table 2). These cores should be re-evaluated considering these palaeoclimate results may determine if a previously unknown sediment deposited throughout the Eocene–Oligocene Transition is present in the nearshore record of the British Isles.
Chattian rainforests. Chattian tropical rainforest (Af) Köppen-Geiger signatures are present in sequences 73/36 and 78/01 and are dated to 25.8–23.03 Ma, based on dinoflagellate cyst biostratigraphy (Wilson, 1996; Egger et al., 2016; King et al., 2016; Table 2). These reconstructions overlapped with the late Oligocene warming event (~26.3–23.7 Ma) in Europe (Liebrand et al., 2017; O’Brien et al., 2020; Brzelinski et al., 2023; Figure 16A). MTCQ optimal values increase above the Köppen-Geiger tropical threshold (Beck et al., 2018) in the Chattian associations F–G of borehole 73/36 (19.40 [±1.61]°C) and I–J (19.40 [±1.61]–21.01 [±1.61]°C) in a rhythmic pattern, resemblant of orbitally-induced forcing (Table 2). Increased MTCQ reconstructions coincide with peaks in the precession-cycle (~26 k.y.), and coeval warming events which occurred from 26.1–25.3 Ma (De Vleeschouwer et al., 2017). Earth’s position in perihelion during this time would have led to intense positive feedback associated with ablative regimes of sea-ice. This led to a reduced albedo effect, which enhanced deep water formation, leading to a warmer, more tropical, North Atlantic in the Chattian (Pälike et al., 2006; De Vleeschouwer et al., 2017; O’Brien et al., 2020).
High winter palaeotemperatures and tropical rainforest-type Northern Hemispheric palaeotemperature reconstructions throughout the Chattian are attributed to Earth being in eccentricity-driven perihelion from May through June (Zachos et al., 2001b; De Vleeschouwer et al., 2017). Large contrasts in winter temperature between higher and lower European latitudes can be attributed to a strengthening of the Hadley Cell (Mantsis et al., 2014). Under increased p CO2 scenarios, dynamic coupling would lead to a narrowing of the Hadley Cell, and in turn would cool Northern Hemispheric climates (Chemke et al., 2019). However, during relatively low pCO2 late Chattian (~300 ppm, O’Brien et al., 2020) periods, combined with periods of enhanced obliquity (41-k.y.) forcing (Xiao et al., 2010) more solar insolation would have been received at high Northern Hemisphere latitudes, causing a reverse in the meridional temperature gradient (Kang et al., 2019). We also note that sensitivity towards tropical climate types may also have been exacerbated by differences in orbital cycling, and subsequent eustatic sea-level fluctuations (Zachos et al., 2001a; Wade and Pӓlike, 2004), the opening and closing of seaways (Kennett, 1977; von der Heydt and Dijkstra, 2006), and aerosols (Bao et al., 2010), and therefore, further research is required.
This led to a reversal of the cross equatorial meridional transport gradient and contributed to increased heat transport to higher latitudes (Lobo and Bordoni, 2020) during winter, rather than in summer. Similar movement of heat occurred in low-obliquity model scenarios, which incorporated a present-day Earth system (Rind, 1998; Davis and Brewer, 2009; Mantsis et al., 2014).
Our tropical winter reconstructions highlight the possibility of an expansion of the Hadley Cell, and the establishment of tropical rainforests at 57°N, resulting from a reduction in temperature seasonality throughout northwest Europe. Tropical winter temperatures on the northwest margin of Europe were enhanced by advection from the tropical North Atlantic temperatures (25–26°C; O’Brien et al., 2020) resulting in a reduced latitudinal temperature gradient, compared to those reconstructed from Central Europe and the North Sea (Roth-Nebelsick et al., 2004; De Man and Van Simaeys, 2004). Our results reconstructed a decline in temperatures and precipitation seasonality, during a time prior to the latest Miocene, when cooling led to the development of before the Northern Hemispheric ice sheets (Herbert et al., 2016; Holbourn et al., 2018; Steinthorsdottir et al., 2021), and sea-ice, were established. Therefore, we suggest that in an Earth system without the Northern Hemispheric ice sheets, the northwest European palaeoclimate had amplified winter and mean annual temperatures (De Vleeschouwer et al., 2017). Such temperature change may be buffered by the existing Northern Hemispheric ice sheets in the present-day, reflected in modern temperatures.
Our results show the consistent preservation of the late Oligocene warming event and the correspondent relative abundance of tropical taxa-types (Figure 15) in the British palaeontological record.
Rupelian forests. We suggest that temperate palaeoclimate classifications, with hot summer signals (Cfa) reconstructed for later Rupelian associations B–F of 21/28B-7 and associations throughout 80/14, are more characteristic of the northwest European temperate palaeoclimates proposed by Pound and Salzmann (2017). Most of the northwestern European Rupelian reconstructions (associations A–B of 16/16B-4; associations C–F of 21/28B-7; association D of 88/12 and association E of 73/36 are most comparable to those taken from the Mittlerer Rupeleton Formation (Belgium), Mainz Basin, (Germany) and the Upper Rhine Graben (Germany) basins, for which a dominantly temperate, hot summer palaeoclimate with no dry season (Cfa) was reconstructed as well (Pross et al., 1998; Maxwell et al., 2016). All our new MATs, with various uncertainty ranges (8.14–19.40°C), are comparable to reconstructions from the Mittlerer Rupeleton Formation (16–18°C), if not cooler (Pross et al., 1998). These reductions in MAT reconstructions are attributable to the latitudinal difference between assemblages (Table 2).
Our new MAP reconstructions (Table 2; Appendix 4) are comparable to reconstruction ranges from the Mittlerer Rupeleton Formation, Mainz Basin, and the Upper Rhine Graben (1000–1300 mm and 1300–1700 mm) (Pross et al., 1998; Maxwell et al., 2016). These reconstructions suggest that the European palaeoclimatic record throughout the Rupelian was temperate-subtropical, and thus comparative to the biome reconstructions that show temperate mixed forests reaching 65.8°N (Pound and Salzmann, 2017; Utescher et al., 2021; Figure 16A–B).
Our results also show summer precipitation events, throughout the Rupelian, were slightly more intense on the northwest margin of Europe, in comparison to those in Central Europe (Table 2; Pross et al., 2001). This is demonstrated by our Rupelian temperate dry-winter Köppen-Geiger climate assignments (Cwa), at Association B of borehole 73/36 and at depths of 515.42–408.43 m of the Mochras borehole (Figure 16A), whereas central European Rupelian reconstructions, from the Mainz Basin (Germany) and Sava Basins (Slovenia) were assigned a no-dry-season Cfa Köppen-Geiger classification (Pross et al., 2001).
Rupelian Dry–Winter Palaeoclimates
Of all our studied intervals, Rupelian reconstructions suggested the largest ranges of seasonal precipitation differences and largest values in precipitation differences, being 4.58–27.30 mm (Table 2, uncertainties variable). Based on the reconstruction of dry-winter Köppen-Geiger types (Cw-) in the northwest European palaeoclimate record, we suggest that the British Isles had a slightly monsoonal-driven precipitation regime, during the Rupelian (Table 2). The Rupelian monsoonal palaeoclimate-type was determined from differences between the Precipitation of the Warmest Quarter and the Precipitation of the Coldest Quarter, and their comparison to the MAP, as adapted from Beck et al. (2018) (Table 2).
A shift in the precipitation regime, towards a temperate, dry-winter, hot-summer signal (Köppen Geiger signal of Cwa), as indicated by an increase in precipitation by 25.12–27.30 mm in summer, was assigned to Oligocene horizons 515.42 m and 408.43 m of the Mochras Basal Red Unit, and in Associations C of the 73/36 borehole (Figure 16A; Table 2; Appendix 4). Association C of the 73/36 borehole has a minimum age of 31.8 Ma, and so the associated assemblage was deposited either throughout the Rupelian age, or possibly Eocene (Table 2). Our Rupelian palaeoclimate dry-winter reconstructions are most like the dry-winter reconstructions from strata from the Upper Rhine Graben, Rauenberg, Germany dated to ~33.6 Ma (Kovar Eder, 2016). Our optima MAT reconstructions are comparable (19.40°C, uncertainty range variable) with those from Germany (19–24°C), though optima northwest European MTWQ reconstructions (24.62°C) were significantly cooler than German reconstructions (28–29°C) (Kovar Eder, 2016). Maximum northwest European MAP optima reconstructions are slightly lower, and thus were indicative of a drier palaeoclimate (1365 ±228 mm, Table 2) than highest MAP values reconstructed from Central European assemblages, of 1300–1700 mm (Kovar Eder, 2016) and 1500 mm (Mai, 1998), respectively. Like Central European signatures, Cwa-types on the northwest margin of Europe are characterised by slightly dry winters, where dry season precipitation does not fluctuate by more than 30 mm, suggesting monsoonal-types were most like Cwa-types in present-day North America near the Gulf of Mexico, presently 25–30°N (Köttek et al., 2006; Peel et al., 2007; Martinez et al., 2012). The northwest European monsoonal record was slightly drier, and cooler in comparison to Central European Rupelian moonsonal palaeoclimates (Kovar-Eder, 2016; Maxwell et al., 2016; Mai, 1995; 1998; Table 2). The temperate palaeoclimate with a dry-winter and hot-summer (Cwa) signal present at 31.8 Ma was not present throughout the remainder of the 73/36 associations (Table 2), and so its temporary appearance may have been the result of an orbitally forced pulse in the precipitation regime, given the Köppen-Geiger climate classification types vary between Cfa and Af in an alternating pattern (Table 2).
Northwest European dry-winter periods, throughout the Rupelian, are most consistent across the westernmost longitudes of the British Isles, highlighting that drier conditions are exacerbated by western processes, i.e., the contribution of the North Atlantic Sea to the terrestrial climate of northwest Europe (Figure 16A–B). Cooler and drier winter palaeoclimatic episodes in northernmost Rupelian Europe, compared to their slightly wetter, more southern European counterparts, suggest latitudinal differences in the winter precipitation regime that are like those driven by El Niño patterns in the present (Brönnimann, 2007; Koumoutsaris et al., 2008). Though, without concrete evidence regarding the timing of the establishment of the proto-El Niño’s Southern Oscillation (ENSO), it is not possible to attribute pulses of enhanced, almost monsoonal-driven, winter drying episodes to an ENSO strengthening.
Dry-winter Rupelian episodes also coincided with a eustatic shallowing throughout the Chattian (Cande and Kent, 1995; Wilson, 1996). These shallowing events lead to reduced sea-level, advection, and moisture transport in the North Atlantic, resulting in dry phases. Expansion of the Hadley Cell would have forced trade winds to subside and migrate tropical temperatures to lower latitudes, i.e., those in Central Europe (Gaemers and Van Hisbergh, 1978). Therefore, Europe’s northwest margin became prone to drier, cloud-free winters with warm to hot summer conditions, with pronounced differences in cross-seasonal precipitation regimes (Chou et al., 2001; Seidel et al., 2008; Bordoni and Schneider, 2008; Velitzelos et al., 2014).
Our results show that in the context of an Earth system without extensive Northern Hemispheric ice sheets, an increase in sea-surface temperature (O’Brien et al., 2020) corresponds to a short-lived (regarding a geological time scale) summer-wet climate, particularly on the westernmost side of the northwest margin of Europe. Reductions in temperature change across seasons, and high MTCQ temperatures on the northwest margin of Europe resulted from high pCO2 concentrations (~600–800 ppm, O’Brien et al., 2020). We suggest high pCO2 concentrations contributed to a weakening of the meridional thermal advection gradient at mid-high latitudes during the cold season, and this led to a reduction in seasonality at higher latitudes (Screen and Simmonds, 2010; Dai et al., 2019; Dai and Deng, 2021).
East-moving jet streams from the tropics may have been induced by a weakened and poleward expansion of the Hadley Cell (Fu et al., 2006; Previdi and Liepert, 2007; Lu et al., 2007; Sharmila and Walsh, 2018), leading to an increase in MTCQ reconstructions at Northern Hemispheric latitudes (Table 2). Our results show that under an increased pCO2 emission scenario, the northwest periphery of Europe may undergo a reduction in temperature seasonality due to increased winter temperatures (Dai and Deng, 2021).
Svalbardella-2 and 3 Events
Our results show that the Svalbardella-2 and 3 North Atlantic cooling events are present in the northwest European palaeorecord. Svalbardella cooksoniae-types were identified in association A of 73/36 (>31.8 Ma), overlapping with the Svalbardella-2 event (Wilson, 1996; Śliwińska and Heilmann-Clausen, 2011; Śliwińska et al., 2014; Appendix 2). Cooling phases throughout the Rupelian were reconstructed from reductions in MAT and MTWQ in 21/28B-7, 16/16B-4, 73/36, and 88/12 (Figure 5, Figure 6, Figure 7, Figure 8). We note that, based on 50% uncertainties, that MAT reductions from 21/28B-7 (Associations B–C) did not overlap (Appendix 4). Based on age re-assignment of associations A–B of 16/16B-4 and associations D–E of 80/14 to 32.8–30.2 Ma, declines in MAT (19.40 [±1.51]–17.89 [±1.51]°C, and 20.91 [±1.51]–19.40 [±1.51]°C, where 50 % uncertainty ranges overlap, respectively) may be associated with the Svalbardella-2 influx and cooling event in the North Atlantic (Pekar and Miller, 1996; Pekar et al., 2002; Śliwińska and Heilmann-Clausen, 2011; Figure 16).
Furthermore, associations D–E and F–G of 16/16B-4 were dated as ~27.1–26.3 Ma based on dinoflagellate cyst biostratigraphy (Egger et al., 2016; Table 2). Reductions in MAT between these associations D–E, of 19.40 (±1.51)–17.89 (±1.51)°C, may correlate with the timings of the Svalbardella-3 cooling event and the Middle Oligocene Glacial Event, at the Rupelian–Chattian boundary (Śliwińska et al., 2010; Clausen et al., 2012; Śliwińska et al., 2014). Thus, not only do we demonstrate that the Svalbardella -2 and 3 events are preserved in the palaeontological record of the British Isles, but that the Rupelian–Chattian boundary may also be preserved. Furthermore, should late Eocene age strata have been preserved, in associations A–B of 80/14, we suggest the possibility that the Svalbardella-1 and corresponding Oi-1a cooling events are also preserved, and had more pronounced cooling, than that observed in Svalbardella-3 reconstructions (Oi-2b), contrary to Śliwińska and Heilmann-Clausen’s (2011) proposal.
Our reconstructions, which overlap with the timing of the Svalbardella influx events, suggest that the palaeoclimate of the northwest margin of Europe may be susceptible to decreases in MAT during cooling phases in a climate scenario without extensive ice sheets in the Northern Hemisphere. Temperature declines, during Svalbardella-2 and 3, were not significant enough to cause changes in the Köppen-Geiger classification of a succession, based on reconstructed optimal values, or summer-type classification (Table 2; Figure 16A). However, our results show that despite the cooling phases present throughout the Oligocene–Miocene climate change much more sensitive, to increased winter temperatures (Table 2; Figure 16A–B).
The Early Miocene Palaeoclimate
Early Miocene conditions were cooler than those reconstructed for the Chattian. Whilst depths in the Lignite and Clay Unit of the Mochras borehole have been suggested to be early Miocene (Woodland, 1971; Herbert-Smith, 1979) we reconstruct reductions in MAT, MTCQ, and MAP, between depths 362.8–362.2 m (Table 2; Figure 5, Figure 7 and Figure 10). We attribute these reductions in palaeoclimate variables to the Oligocene–Miocene cooling event at Mi-1, 23.03 Ma (Zachos et al., 1997; Table 2). Reductions in our Mochras (optima) MAT of <2°C, previously, are associated with the Oligocene–Miocene boundary, and subsequent time towards the Middle Miocene (Śliwińska et al., 2014; Greenop et al., 2014). This resulted in the removal of tropical taxa from the vegetation record of the British Isles (Figure 15). An Early Miocene age was assigned to association E of 77/07 (Figure 16A–B), based on the calibration of its pollen and dinoflagellate cyst assemblages with 16/16B-4 (Wilson, 1996). The uppermost depths of association F of 16/16B-4 and the association D of 77/07 are assigned an age of 26.6 Ma (King et al., 2016; Egger et al., 2016). Wilson (1996) suggested that associations E of 77/07 were likely younger, based on the 1.52 m hiatus between associations D–E. Association E was assigned an Early Miocene age, based on its assignment of a temperate warm-summer Köppen-Geiger signature with no dry season (Cfb, Table 2). We associate this summer cooling with the onset of the δ18O maximum at the Oligocene–Miocene Transition, which leads to ice growth comparable, if not exceeding present-day volumes, and consequential cooling (Figure 16A–B; Liebrand et al., 2017; Miller et al., 2020; Steinthorsdottir et al., 2021).
This cooling phase was associated with long (400-k.y.) and short (100-k.y.) eccentricity cycling, before 16.9 Ma (Liebrand et al., 2016; Steinthorsdottir et al., 2021). Given the post-Oligocene-ages were assigned to the Castlemartin-3 assemblage, and to the Lignite and Clay Unit (368.8–368.2 m) of the Mochras borehole were also assigned a warm-summer signature (Cfb), we suggest these assemblages may also be early Miocene in age (Table 2; Woodland, 1971; McLean, 2002).
Given the Little Minch (Evans et al., 1991) assemblage, also reconstructed a Cfb signature, at depths 62.5–72 m, it is possible that these beds were deposited either throughout the Oligocene–Miocene Transition, or sometime throughout the Svalbardella-3 cooling phase. This is because our reconstructions, which overlapped with Svalbardella-3 (sequences Well 28, 88/12, 21/28B-7 and 16/16B-4), were used to reconstruct a temperate hot-summer climate with no dry season (Cfa), rather than a warm-summer (Cfb) Köppen-Geiger climate type. This warm-summer classification suggests the Little Minch (Evans et al., 1991) reconstructions are more likely early Miocene in age, given the consistency of Cfb-types in the Early Miocene age assemblages (Figure 16A). Sample 6 of Little Minch (Evans et al., 1991) was deposited above Association H of 80/14 (64.95–63.97 m), and so this core section is 25.5 Ma, at oldest (Wilson, 1996). Sample 6’s assigned Cfb signature is comparable with the early Miocene signature from association E of 77/07. This suggests that sample 6 (Evans et al., 1991) is also likely Early Miocene in age. The disappearance of Arecaceae from the record of Evans et al. (1991), from ~72 m, further supports this early Miocene age assessment (Appendix 2; Figure 16A).
All our early Miocene temperature and precipitation palaeoclimate variable reconstructions are consistently lower than those reconstructed from the early Miocene Gröbern assemblage (Roth-Nebelsick et al., 2004). Our optimised early Miocene MAT reconstructions <11.86 (±1.51)°C were lower than the Gröbern (Germany) assemblage’s MAT range, reconstructed using the co-existence approach, 14.1–20.8°C (Roth-Nebelsick et al., 2004). We show our optimised MTCQ reconstructions (4.92–6.53°C) overlap with the Gröbern Coldest Mean Month Temperature ranges of 4.7–13.3°C, as do our MAP reconstructions (853–1251 mm [variable uncertainty ranges]), with those reconstructed from Germany (897–1281 mm) (Roth-Nebelsick et al., 2004). However, our MTWQ reconstructions (17.59–21.81°C [variable uncertainty ranges]) are considerably cooler (25.7–28.1°C) than those from Gröbern (Roth-Nebelsick et al., 2004). CREST reconstructions are known to be cooler than co-existence approach reconstructions due to differences in input data and mathematical approach (Gibson et al., 2022). Furthermore, CREST and the co-existence approach use different modern climate databases, thus input data is different, resulting in inconsistencies between techniques (Mosbrugger and Utescher, 1997; Utescher et al., 2014; Grimm and Potts, 2016; Gibson et al., 2022). Our early Miocene assemblages reconstructed the smallest differences between summer and winter precipitation, of all studied intervals (-0.47–2.79 mm) (Table 2). The negligible, dry-summer seasonal difference in precipitation (<1 mm) suggests that following the Oligocene–Miocene Transition, there was a change in the precipitation regime. This likely resulted in the consistent, almost wholly equal precipitation distribution across our study sites (Table 2) up until the middle Miocene (Gibson et al., 2022). Our results reinforce the proposal that the precession (~26 k.y.) cycle had a bimodal impact on palaeoclimate (De Vleeschouwer et al., 2017), and so the northward (southward) migration of monsoonal-climate and slightly dry-winters bands associated with decreased (increased) precessional cycling was balanced (Ding et al., 2021). This resulted in an alternating (but small) fluctuation between positive and negative seasonal differences in precipitation, suggestive of a pulse-like fluctuation between early Miocene Köppen-Geiger types with very slight dry-summers and dry-winter signals (Table 2).
CONCLUSIONS
The Oligocene and Miocene sedimentary archives from the British Isles record orbitally induced climate change. Tropical rainforest types were reconstructed in short-lived pulses during the Chattian and the Rupelian/Priabonian. During the Chattian, the reconstructed tropical rainforest Köppen-Geiger signatures are coeval with the late Oligocene warming event and these rainforest types could have been a response to obliquity maxima forcing. Dry-winter Köppen-Geiger reconstructions, during the Rupelian, may have been driven by eccentricity-modulated pulses in the precession cycle (~26 k.y.) that lead to strengthened feedback between oceanic-atmospheric interactions.
The Svalbardella-2 and 3 cooling events are apparent in the palaeontological record of the British Isles and are represented by respective reductions in MAT (24.22–22.61°C) and MTCQ (23.22–17.59°C). The Oligocene–Miocene Transition may have been preserved in the Mochras and 77/07 boreholes, with early Miocene reconstructions showing a MAT reduction from 24.22 to 11.36°C, and a MTWQ reduction from 24.62 to 16.18°C: the latter leading to a transition from hot to warm summer palaeoclimate types due to summer cooling. After the Oligocene–Miocene Transition, differences in seasonal precipitation were almost negligible perhaps because of the hemispheric-bimodal influence of the precession-cycle after the Oligocene–Miocene Transition. We suggest that under similar pCO2 levels, the northwest edge of Europe may have wetter summers and possible tropical winter temperature because of anthropogenic climate change.
ACKNOWLEDGEMENTS
We would like to thank A.J. Suggitt for his help in resolving model errors in R. JM thanks NERC (NERC award identified NE/S007512/1) for funding ongoing research into the Cenozoic palynology of northwest Europe, through the ONEPlanet Doctoral Training Programme (OP 2170). MP, JO'K, and JM acknowledge funding from NSF/Geo- NERC project “Fungi in a Warmer World (FiaWW)” (NSF award #2015813 to JO’K and NERC award identifier NE/V01501X/1 to MP). MP and JM thank The Royal Society (IECR2202086). The authors thank J. Hue, M. Hyžný and J. Rumford for facilitating the manuscript through Palaeontologia Electronica. We are grateful to two anonymous reviewers for their comments which helped greatly in the development of our manuscript.
REFERENCES
Antropov, A.V., Belokobylskij, S.A., Compton, S.G., Dlussky, G.M., Khalaim, A.I., Kolyada, V.A., Kozlov, M.A., Perfilieva, K.S., and Rasnitsyn, A.P. 2013. The wasps, bees and ants (insecta: Vespida Hymenoptera) from the insect limestone (late Eocene) of the Isle of Wight UK. Earth and Environmental Science Transactions of the Royal Society of Edinburgh, 104(3-4):335–446.
https://doi.org/10.1017/S1755691014000103
Armstrong McKay, D.I., Staal, A., Abrams, J.F., Winkelmann, R., Sakschewski, B., Loriani, S., Fetzer, I., Cornell, S.E., Rockström, J., and Lenton, T.M. 2022. Exceeding 1.5°C global warming could trigger multiple climate tipping points. Science, 377(6611):1–10.
https://doi.org/10.1126/science.abn7950
Bao, H., Yu, S., and Tong, D.Q. 2010. Massive volcanic SO2 oxidation and sulphate aerosol deposition in Cenozoic North America. Nature, 465(7300):909–912.
Beck, H.E., Zimmermann, N.E., McVicar, T.R., Vergopolan, N., Berg, A., and Wood, E.F. 2018. Present and future Köppen-Geiger climate classification maps at 1-km resolution. Scientific Data, 5(1):1–12.
https://doi.org/10.1038/s41597-020-00616-w
Beddow, H.M., Liebrand, D., Sluijs, A., Wade, B.S., and Lourens, L.J. 2016. Global change across the Oligocene–Miocene transition high-resolution stable isotope records from IODP Site U1334 (equatorial Pacific Ocean). Paleoceanography, 31(1):81–97.
https://doi.org/10.1002/2015PA002820
Bessedik, M. 1984. The early Aquitanian and upper Langhian-lower Serravallian environments in the northwestern Mediterranean region. Paléobiologie Continentale, 14(2):153–179.
Bhatia, H., Srivastava, G., Spicer, R.A., Farnsworth, A., Spicer, T.E., Mehrotra, R.C., Paudayal, K.N., and Valdes, P. 2021. Leaf physiognomy records the Miocene intensification of the South Asia Monsoon. Global and Planetary Change, 196:103365.
https://doi.org/10.1016/j.gloplacha.2020.103365
Boers, N. and Rypdal, M. 2021. Critical slowing down suggests that the western Greenland ice sheet is close to a tipping point. Proceedings of the National Academy of Sciences, USA, 118(21):e2024192118.
https://doi.org/10.1073/pnas.2024192118
Bordoni, S. and Schneider, T. 2008. Monsoons as eddy-mediated regime transitions of the Tropical Overturning Circulation. Nature Geoscience, 1(8):515–519.
https://doi.org/10.1038/ngeo248
Bortenschlager, S. 1990. Aspects of pollen morphology in the Cupressaceae. Grana, 29(2):129–138.
Bouchal, J.M. and Denk, T. 2020. Low taxonomic resolution of papillate Cupressaceae pollen former Taxodiaceae impairs their applicability for palaeo-habitat reconstruction. Grana, 59(1):70–93.
Boulter, M.C. 1980. Irish Tertiary plant fossils in a European context. Journal of Earth Sciences:1–11.
Boulter, M.C. and Craig, D.L. 1979. A middle Oligocene pollen and spore assemblage from the Bristol Channel. Review of Palaeobotany and Palynology, 28(3-4):259–272.
https://doi.org/10.1016/0034-6667(79)90028-9
Brönnimann, S. 2007. Impact of El Niño-Southern Oscillation on European climate. Reviews of Geophysics, 45(3):1–28.
https://doi.org/10.1029/2006RG000199
Brzelinski, S., Bornemann, A., Liebrand, D., van Peer, T.E., Wilson, P.A., and Friedrich, O. 2023. Large obliquity-paced Antarctic ice-volume fluctuations suggest melting by atmospheric and ocean warming during late Oligocene. Communications Earth and Environment, 4(1):222. https://doi.org/10.1038/s43247-023-00864-9
Cande, S.C. and Kent, D.V. 1995. Revised calibration of the geomagnetic polarity timescale for the late Cretaceous and Cenozoic. Journal of Geophysical Research: Solid Earth, 100(B4):60936095.
https://doi.org/10.1029/94JB03098
Chaanda, M.S. 2016. Cenozoic terrestrial palaeoenvironemtal change an investigation of the Petrockstowe and Bovey basins southwest United Kingdom. PhD Thesis, University of Plymouth, Plymouth, UK.
Chemke, R., Polvani, L.M., and Deser, C. 2019. The effect of Arctic Sea ice loss on the Hadley circulation. Geophysical Research Letters, 46(2):963–972.
https://doi.org/10.1029/2018GL081110
Chevalier, M. 2019. Enabling possibilities to quantify past climate from fossil assemblages at a global scale. Global and Planetary Change, 175:27–35.
https://doi.org/10.1016/j.gloplacha.2019.01.016
Chevalier, M. 2022. crestr: an R package to perform probabilistic climate reconstructions from palaeoecological datasets. Climate of the Past, 18(4):821–844.
https://github.com/mchevalier2/crestr
https://doi.org/10.5194/cp-18-821-2022
Chevalier, M., Cheddadi, R., and Chase, B.M. 2014. CREST (Climate REconstruction SofTware): a probability density function (PDF)-based quantitative climate reconstruction method. Climate of the Past, 10(6):2081–2098.
https://doi.org/10.5194/cp-10-2081-2014
Chou, C., Neelin, J.D., and Su, H. 2001. Ocean-atmosphere-land feedbacks in an idealized monsoon. Quarterly Journal of the Royal Meteorological Society, 127(576):1869–1891.
https://doi.org/10.1002/qj.49712757602
Clausen, O.R., Śliwińska, K.K., and Gołędowski, B. 2012. Oligocene climate changes controlling forced regression in the eastern North Sea. Marine and Petroleum Geology, 29(1):1–14.
https://doi.org/10.1016/j.marpetgeo.2011.10.002
Collinson, M.E. and Hooker, J.J. 1987. Vegetational and mammalian faunal changes in the early Tertiary of southern England, p. 259–303. In Friis, E.M., Chaloner, W.G., and Crane, P.R. (eds.), The origins of angiosperms and their biological consequences. Cambridge University Press, Cambridge.
Coxall, H.K., Wilson, P.A., Pälike, H., Lear, C.H., and Backman, J. 2005. Rapid stepwise onset of Antarctic glaciation and deeper calcite compensation in the Pacific Ocean. Nature, 433(7021):53–57.
Crowley, T.J. and Berner, R.A. 2001. CO2 and climate change. Science, 292(5518):870–872.
https://doi.org/10.1126/science.1061664
Dai, A., Luo, D., Song, M., and Liu, J. 2019. Arctic amplification is caused by sea-ice loss under increasing CO2. Nature Communications, 10(1):1–13.
https://doi.org/10.1038/s41467-018-07954-9
Dai, A. and Deng, J. 2021. Arctic amplification weakens the variability of daily temperatures over northern middle-high latitudes. Journal of Climate, 34(7):2591–2609.
https://doi.org/10.1175/JCLI-D-20-0514.1
Daley, B. 1989. Silica pseudomorphs from the Bembridge limestone (upper Eocene) of the Isle of Wight, southern England and their palaeoclimatic significance. Palaeogeography, Palaeoclimatology, Palaeoecology, 69:233–240.
https://doi.org/10.1016/0031-0182(89)90166-1
Davis, B.A. and Brewer, S. 2009. Orbital forcing and role of the latitudinal insolation/temperature gradient. Climate Dynamics, 32:143–165.
https://doi.org/10.1007/s00382-008-0480-9
De Man, E. and Van Simaeys, S. 2004. Late Oligocene warming event in the southern North Sea Basin benthic foraminifera as paleotemperature proxies. Netherlands Journal of Geosciences, 83(3):227–239.
https://doi.org/10.1017/S0016774600020291
De Vleeschouwer, D., Vahlenkamp, M., Crucifix, M., and Pälike, H. 2017. Alternating Southern and Northern Hemisphere climate response to astronomical forcing during the past 35 my. Geology, 45(4):375–378.
https://doi.org/10.1130/G38663.1
Denk, T., Grimm, G.W., Grímsson, F., and Zetter, R. 2013. Evidence from Köppen signatures of fossil plant assemblages for effective heat transport of Gulf Stream to subarctic North Atlantic during Miocene cooling. Biogeosciences, 10(12):7927–7942.
https://doi.org/10.5194/bg-10-7927-2013
Ding, Z., Huang, G., Liu, F., Wu, R., and Wang, P. 2021. Responses of global monsoon and seasonal cycle of precipitation to precession and obliquity forcing. Climate Dynamics, 56:3733–3747.
https://doi.org/10.1007/s00382-021-05663-6
Doppert, J.W.C. and Neele, N.G. 1983. Biostratigraphy of marine Paleogene deposits in the Netherlands and adjacent areas. Mededelingen Rijks Geologische Dienst, 37(2):–79.
Dupont-Nivet, G., Krijgsman, W., Langereis, C.G., Abels, H.A., Dai, S., and Fang, X. 2007. Tibetan plateau aridification linked to global cooling at the Eocene-Oligocene transition. Nature, 445(7128):635–638.
https://doi.org/10.1038/nature05516
Egger, L.M., Śliwińska, K.K., Van Peer, T.E., Liebrand, D., Lippert, P.C., Friedrich, O., Wilson, P.A., Norris, R.D., and Pross, J. 2016. Magnetostratigraphically-calibrated dinoflagellate cyst bioevents for the uppermost Eocene to lowermost Miocene of the western North Atlantic (IODP Expedition 342, Paleogene Newfoundland sediment drifts). Review of Palaeobotany and Palynology, 234:159–185.
https://doi.org/10.1016/j.revpalbo.2016.08.002
Ehrmann, W.U. and Mackensen, A. 1992. Sedimentological evidence for the formation of an east Antarctic ice sheet in Eocene/Oligocene time. Palaeogeography, Palaeoclimatology, Palaeoecology, 93(1-2):85–112.
https://doi.org/10.1016/0031-0182(92)90185-8
Eldrett, J.S., Greenwood, D.R., Harding, I.C., and Huber, M. 2009. Increased seasonality through the Eocene to Oligocene transition in northern high latitudes. Nature, 459(7249):969–973.
https://doi.org/10.1038/nature08069
Evans, D., Hallsworth, C., Jolley, D.W., and Morton, A.C. 1991. Late Oligocene terrestrial sediments from a small basin in the Little Minch. Scottish Journal of Geology, 27(1):33–40.
https://doi.org/10.1144/sjg27010033
Farnsworth, A., Lunt, D.J., Robinson, S.A., Valdes, P.J., Roberts, W.H., Clift, P.D., Markwick, P., Su, T., Wrobel, N., Bragg, F., and Kelland, S.J. 2019. Past east Asian monsoon evolution controlled by paleogeography, not CO2. Science Advances, 5(10):eaax1697.
https://doi.org/10.1126/sciadv.aax1697
Fenero, R., Cotton, L., Molina, E., and Monechi, S. 2013. Micropalaeontological evidence for the late Oligocene Oi-2b global glaciation event at the Zarabanda section Spain. Palaeogeography, Palaeoclimatology, Palaeoecology, 369:1–13.
https://doi.org/10.1016/j.palaeo.2012.08.020
Fick, S.E. and Hijmans, R.J. 2017. WorldClim 2 new 1-km spatial resolution climate surfaces for global land areas. International Journal of Climatology, 37(12):4302–4315.
https://doi.org/10.1002/joc.5086
Fitzgerald, J.A. 1999. Pollen and spore assemblages from the Oligocene Lough Neagh Group Vol. 1, Published PhD Thesis, University of Sheffield, Sheffield, UK.
Figueiral, I., Mosbrugger, V., Rowe, N.P., Ashraf, A.R., Utescher, T., and Jones, T.P. 1999. The Miocene peat-forming vegetation of northwestern Germany an analysis of wood remains and comparison with previous palynological interpretations. Review of Palaeobotany and Palynology, 104(3-4):239–266.
https://doi.org/10.1016/S0034-6667(98)00059-1
Fu, Q., Johanson, C.M., Wallace, J.M., and, Reichler, T. 2006. Enhanced mid-latitude tropospheric warming in satellite measurements. Science, 312(5777):1179.
https://doi.org/10.1126/science.1125566
Gaemers, P.A. and van Hinsbergh, V.W.M. 1978. Rupelian middle Oligocene fish otoliths from the clay pit'De Vlijt'near Winterswijk the Netherlands. Scripta Geologica, 46:1–77.
Galeotti, S., DeConto, R., Naish, T., Stocchi, P., Florindo, F., Pagani, M., Barrett, P., Bohaty, S.M., Lanci, L., Pollard, D., and Sandroni, S. 2016. Antarctic ice sheet variability across the Eocene-Oligocene boundary climate transition. Science, 352(6281):76–80.
GBIF. 2020a. Cycadopsida occurrence data, downloaded 24 September 2020. GBIF Database.
https://doi.org/10.15468/dl.sfjzxu
GBIF. 2020b. Gnetopsida occurrence data, downloaded 24 September 2020. GBIF Database.
https://doi.org/10.15468/dl.h2kjnc
GBIF. 2020c. Liliopsida occurrence data, downloaded 24 September 2020. GBIF Database.
https://doi.org/10.15468/dl.axv3yd
GBIF. 2020d. Lycopodiopsida occurrence data, downloaded 24 September 2020. GBIF Database.
https://doi.org/10.15468/dl.ydhyhz
GBIF. 2020e. Magnoliopsida occurrence data, downloaded 24 September 2020. GBIF Database.
https://doi.org/10.15468/dl.ra49dt
GBIF. 2020f. Pinopsida occurrence data, downloaded 24 September 2020. GBIF Database.
https://doi.org/10.15468/dl.x2r7pa
GBIF. 2020g. Polypodiopsida occurrence data, downloaded 24 September 2020. GBIF Database.
https://doi.org/10.15468/dl.87tbp6
Gorochov, A.V. 2019. Crickets and grasshoppers Orthoptera from the latest Eocene of the Isle of Wight southern England UK. Earth and Environmental Science Transactions of the Royal Society of Edinburgh, 110(3-4):317–329.
https://doi.org/10.1017/S1755691018000889
Greenop, R., Foster, G.L., Wilson, P.A., and Lear, C.H. 2014. Middle Miocene climate instability associated with high-amplitude CO2 variability. Paleoceanography, 29(9):845–853.
https://doi.org/10.1002/2014PA002653
Greenop, R., Sosdian, S.M., Henehan, M.J., Wilson, P.A., Lear, C.H., and Foster, G.L. 2019. Orbital forcing, ice volume, and CO2 across the Oligocene–Miocene transition. Paleoceanography and Paleoclimatology, 34(3):316–328.
https://doi.org/10.1029/2018PA003420
Grimm, G.W. and Potts, A.J. 2016. Fallacies and fantasies the theoretical underpinnings of the coexistence approach for palaeoclimate reconstruction. Climate of the Past, 12(3):611–622. https://doi.org/10.1016/j.revpalbo.2016.07.001
Hall, R., Erdélyi, R., Hanna, E., Jones, J.M., and Scaife, A.A. 2015. Drivers of North Atlantic polar front jet stream variability. International Journal of Climatology, 35(8):1697–1720.
Han, F., Yang, T., Zhang, K., Hou, Y., and Song, B. 2020. Early Oligocene Podocarpium Leguminosae from Qaidam Basin and its paleoecological and biogeographical implications. Review of Palaeobotany and Palynology, 282:104309.
https://doi.org/10.1016/j.revpalbo.2020.104309
Haq, B.U., Hardenbol, J., and Vail, P.R. 1987. Chronology of fluctuating sea levels since the Triassic. Science, 235(4793):1156–1167.
https://doi.org/10.1002/joc.4121
Henrot, A.J., François, L., Favre, E., Butzin, M., Ouberdous, M., and Munhoven, G. 2010. Effects of CO2, continental distribution topography and vegetation changes on the climate at the middle Miocene a model study. Climate of the Past, 6(5):675–694.
https://doi.org/10.5194/cp-6-675-2010
Herbert-Smith, M. 1971. The Palynology of the Mochras borehole. PhD Thesis, University College of Wales, Aberystwyth, UK.
Herbert-Smith, M. 1979. The Age of the Tertiary Deposits of the Llanbedr Mochras Farm borehole as determined from palynological studies, Institute of Geological Sciences Report 78/24:15–29.
Herbert, T.D., Lawrence, K.T., Tzanova, A., Peterson, L.C., Caballero-Gill, R., and Kelly, C.S. 2016. Late Miocene global cooling and the rise of modern ecosystems. Nature Geoscience, 9(11):843–847.
https://doi.org/10.1038/ngeo2813
Holbourn, A.E., Kuhnt, W., Clemens, S.C., Kochhann, K.G., Jöhnck, J., Lübbers, J., and Andersen, N. 2018. Late Miocene climate cooling and intensification of southeast Asian winter monsoon. Nature Communications, 9(1):1584.
https://doi.org/10.1038/s41467-018-03950-1
Hren, M.T., Sheldon, N.D., Grimes, S.T., Collinson, M.E., Hooker, J.J., Bugler, M., and Lohmann, K.C. 2013. Terrestrial cooling in Northern Europe during the Eocene-Oligocene transition. Proceedings of the National Academy of Sciences USA, 110(19):7562–7567.
https://doi.org/10.1073/pnas.1210930110
Hugueney, M. and Truc, G. 1976. Decouvertes recentes de mammiferes et de mollusquesdans des formations d'age oligocene terminal et aquitanien du SE de la France comparaison avec les gisements deja connus dans la meme region. Geobios, 9(3):359–362.
https://doi.org/10.1016/S0016-6995(76)80041-1
Hulme, M. and Barrow, E.M. 1997. Climates of the British Isles present, past and future. Psychology Press Routledge, London.
Hutchinson, D.K., Coxall, H.K., Lunt, D.J., Steinthorsdottir, M., De Boer, A.M., Baatsen, M., von der Heydt, A., Huber, M., Kennedy-Asser, A.T., Kunzmann, L., and Ladant, J.B. 2021. The Eocene-Oligocene transition: a review of marine and terrestrial proxy data, models and model-data comparisons. Climate of the Past, 17(1):269–315.
https://doi.org/10.5194/cp-17-269-2021
Hyeong, K., Kuroda, J., Seo, I., and, Wilson, P.A. 2016. Response of the Pacific inter-tropical convergence zone to global cooling and initiation of Antarctic glaciation across the Eocene Oligocene Transition. Scientific Reports, 6(1):30647.
https://doi.org/10.1038/srep30647
IPCC. 2022. Climate change 2022 impacts adaptation, and vulnerability. In Pörtner, H.-O., Roberts, D.C., Tignor, M., Poloczanska, E.S., Mintenbeck, K., Alegría, A., Craig, M., Langsdorf, S., Löschke, S., Möller, V., Okem, A., and Rama, B. (eds.), Contribution of working group II to the sixth assessment report of the Intergovernmental Panel on Climate Change. Cambridge University Press, Cambridge, UK and New York, USA.
https://doi.org/10.1017/9781009325844
Ivany, L.C., Patterson, W.P., and Lohmann, K.C. 2000. Cooler winters as a possible cause of mass extinctions at the Eocene/ Oligocene boundary. Nature, 407(6806):887–890.
https://doi.org/10.1038/35038044
Ivany, L.C., Van Simaeys, S., Domack, E.W., and Samson, S.D. 2006. Evidence for an earliest Oligocene ice sheet on the Antarctic Peninsula. Geology, 34(5):377–380.
https://doi.org/10.1130/G22383.1
Jovane, L., Florindo, F., Sprovieri, M., and Pälike, H. 2006. Astronomic calibration of the late Eocene early Oligocene Massignano section central Italy. Geochemistry Geophysics Geosystems, 7(7).
https://doi.org/10.1029/2005GC001195
Jowsey, N.L., Parkin, D.L., Slipper, I.J., Smith, A.P.C., and, Walsh, P.T. 1992. The geology and geomorphology of the Beacon Cottage Farm Outlier St Agnes Cornwall. Geological Magazine, 129(1):101–121.
https://doi.org/10.1017/S0016756800008153
Kang, W., Cai, M., and Tziperman, E. 2019. Tropical and extratropical general circulation with a meridional reversed temperature gradient as expected in a high obliquity planet. Icarus, 330:142–154.
https://doi.org/10.1016/j.icarus.2019.04.028
Katz, M.E., Miller, K.G., Wright, J.D., Wade, B.S., Browning, J.V., Cramer, B.S., and Rosenthal, Y. 2008. Stepwise transition from the Eocene greenhouse to the Oligocene icehouse. Nature Geoscience, 1(5):329–334.
https://doi.org/10.1038/ngeo179
Kennett, J.P. 1977. Cenozoic evolution of Antarctic glaciation the circum-Antarctic Ocean and their impact on global paleoceanography. Journal of Geophysical Research, 82(27):3843–3860.
https://doi.org/10.1029/jc082i027p03843
Kennett, J.P. and Shackleton, N.J. 1976. Oxygen isotopic evidence for the development of the psychrosphere 38 my ago, Nature, 260(513).
King, C. 1983. Cainozoic micropalaeontological biostratigraphy of the North Sea. Report Institute of Geological Sciences 82(7):1-40.
King, C. 1989. Cenozoic of the North Sea, p. 418–489. In Jenkins, D.G. and Murray, J.W. (eds.), Stratigraphical atlas of fossil foraminifera. British Micropalaeontological Society Series Ellis Horwood Limited, Chichester.
King, C., Gale, A.S., and Barry, T.L. 2016. A revised correlation of Tertiary rocks in the British Isles and adjacent areas of NW Europe, The Geological Society of London Special Reports No. 27, London.
https://doi.org/10.1144/SR27
Koumoutsaris, S., Bey, I., Generoso, S., and Thouret, V. 2008. Influence of El Niño-Southern Oscillation on the interannual variability of tropospheric ozone in the northern midlatitudes. Journal of Geophysical Research Atmospheres, 113(D19).
https://doi.org/10.1029/2007JD009753
Kovar-Eder, J. 2016. Early Oligocene plant diversity along the Upper Rhine Graben the fossil flora of Rauenberg Germany. Acta Palaeobotanica, 56(2):329–440.
https://doi.org/10.1515/acpa-2016-0011
Krzemiński, W., Blagoderov, V., Azar, D., Lukashevich, E., Szadziewski, R., Wedmann, S., André, N.E.L., Collomb, F.M., Waller, A., and Nicholson, D.B. 2019. True flies insecta: Diptera from the late Eocene insect limestone Bembridge Marls of the Isle of Wight England UK. Earth and Environmental Science Transactions of the Royal Society of Edinburgh, 110(3-4):495–554.
https://doi.org/10.1017/S1755691018000464
Ladant, J.B., Donnadieu, Y., Lefebvre, V., and Dumas, C. 2014. The respective role of atmospheric carbon dioxide and orbital parameters on ice sheet evolution at the Eocene-Oligocene transition. Paleoceanography, 29(8):810–823.
https://doi.org/10.1002/2013PA002593
Lear, C.H., Elderfield, H., and Wilson, P.A. 2000. Cenozoic deep-sea temperatures and global ice volumes from Mg/Ca in benthic foraminiferal calcite. Science, 287(5451):269–272.
https://doi.org/10.1126/science.287.5451.269
Lear, C.H., Rosenthal, Y., Coxall, H.K.. and, Wilson, P.A. 2004. Late Eocene to early Miocene ice sheet dynamics and the global carbon cycle. Paleoceanography, 19(4).
https://doi.org/10.1029/2004PA001039
Levy, R.H., Meyers, S.R., Naish, T.R., Golledge, N.R., McKay, R.M., Crampton, J.S., DeConto, R.M., De Santis, L., Florindo, F., Gasson, E.G., and Harwood, D.M. 2019. Antarctic ice-sheet sensitivity to obliquity forcing enhanced through ocean connections. Nature Geoscience, 12(2):132–137.
https://doi.org/10.1038/s41561-018-0284-4
Li, M., Wan, S., Colin, C., Jin, H., Zhao, D., Pei, W., Jiao, W., Tang, Y., Tan, Y., Shi, X., and Li, A. 2023. Expansion of C4 plants in South China and evolution of East Asian monsoon since 35 Ma black carbon records in the northern South China Sea. Global and Planetary Change, 223:104079.
https://doi.org/10.1016/j.gloplacha.2023.104079
Liebrand, D., Beddow, H.M., Lourens, L.J., Pälike, H., Raffi, I., Bohaty, S.M., Hilgen, F.J., Saes, M.J., Wilson, P.A., Van Dijk, A.E., and Hodell, D.A. 2016. Cyclostratigraphy and eccentricity tuning of the early Oligocene through early Miocene 30.1-17.1 Ma Cibicides mundulus stable oxygen and carbon isotope records from Walvis Ridge Site 1264. Earth and Planetary Science Letters, 450:392–405.
https://doi.org/10.1016/j.epsl.2016.06.007
Liebrand, D., de Bakker, A.T., Beddow, H.M., Wilson, P.A., Bohaty, S.M., Ruessink, G., Pälike, H., Batenburg, S.J., Hilgen, F.J., Hodell, D.A., and Huck, C.E. 2017. Evolution of the early Antarctic ice ages. Proceedings of the National Academy of Sciences USA, 114(15):3867–3872.
https://doi.org/10.1073/pnas.1615440114
Lin, X., Wyrwoll, K.H., Chen, H., Zhang, X., Fu, K., Sun, M., Yu, X., and Shen, Z. 2015. An active east Asian monsoon at the Oligocene–Miocene boundary evidence from the Sikouzi Section, Northern China. The Journal of Geology, 123(4):355–367.
https://doi.org/10.1086/682278
Ling, C.C., Ma, F.J., Dong, J.L., Zhou, G.H., Wang, Q.J., and Sun, B.N. 2021. A mid-altitude area in southwestern China experienced a humid subtropical climate with subtle monsoon signatures during the early Oligocene evidence from the Ningming flora of Guangxi. Palaeogeography, Palaeoclimatology, Palaeoecology, 579:110601.
https://doi.org/10.1016/j.palaeo.2021.110601
Liu, Z., Pagani, M., Zinniker, D., DeConto, R., Huber, M., Brinkhuis, H., Shah, S.R., Leckie, R.M., and Pearson, A. 2009. Global cooling during the Eocene Oligocene climate transition. Science, 323(5918):1187–1190.
https://doi.org/10.1126/science.1166368
Lobo, A.H. and Bordoni, S. 2020. Atmospheric dynamics in high obliquity planets. Icarus, 340:113592.
https://doi.org/10.1016/j.icarus.2019.04.028
Lu, J., Vecchi, G.A., and Reichler, T. 2007. Expansion of the Hadley cell under global warming. Geophysical Research Letters, 34(6):L06805.
https://doi.org/10.1029/2006GL028443
Mai, D.H. 1995. Tertiӓre Vegetationgeschichte Europas. Fischer, Jena:691.
Mai, D.H. 1998. Contribution to the flora of the middle Oligocene Calau Beds in Brandenburg, Germany. Review of Palaeobotany and Palynology, 101(1-4):43–70.
https://doi.org/10.1016/S0034-6667(97)00069-9
Mantsis, D.F., Lintner, B.R., Broccoli, A.J., Erb, M.P., Clement, A.C., and Park, H.S. 2014. The response of large-scale circulation to obliquity-induced changes in meridional heating gradients. Journal of Climate, 27(14): 5504-5516.
https://doi.org/10.1175/JCLI-D-13-00526.1
Martinez, C.J., Maleski, J.J., and Miller, M.F. 2012. Trends in precipitation and temperature in Florida USA. Journal of Hydrology, 452:259–281.
https://doi.org/10.1016/j.jhydrol.2012.05.066
Maxwell, E.E., Alexander, S., Bechly, G., Eck, K., Frey, E., Grimm, K., Kovar-Eder, J., Mayr, G., Micklich, N., Rasser, M., and Roth-Nebelsick, A. 2016. The Rauenberg fossil Lagerstätte Baden-Württemberg, Germany a window into early Oligocene marine and coastal ecosystems of Central Europe. Palaeogeography, Palaeoclimatology, Palaeoecology, 463:238–260.
https://doi.org/10.1016/j.palaeo.2016.10.002
Mayes, J.C. 1991. Regional airflow patterns in the British Isles. International Journal of Climatology, 11(5):473–491.
Mayes, J. 1996. Spatial and temporal fluctuations of monthly rainfall in the British Isles and variations in the mid-latitude westerly circulation, International Journal of Climatology A Journal of the Royal Meteorological Society, 16(5):585–596.
https://doi.org/10.1002/(SICI)1097-0088(199605)16:5<585::AID-JOC24>3.0.CO;2-%23
McCobb, L.M.E., Duncan, I.J., Jarzembowski, E.A., Stankiewicz, B.A., Wills, M.A., and Briggs, D.E.G. 1998. Taphonomy of the insects from the insect bed Bembridge Marls late Eocene Isle of Wight England. Geological Magazine, 135(4):553–563.
https://doi.org/10.1017/S0016756898001204
McLean, D. 2002. The nature age and origin of the Flimston Clay Pembrokeshire palynological and mineralogical analyses. Countyside Council for Wales Report No. 0019:1–26.
Methner, K., Campani, M., Fiebig, J., Löffler, N., Kempf, O., and Mulch, A. 2020. Middle Miocene long-term continental temperature change in and out of pace with marine climate records. Scientific Reports, 10(1):7989.
https://doi.org/10.1038/s41598-020-64743-5
Miao, Y., Wu, F., Herrmann, M., Yan, X., and Meng, Q. 2013. Late early Oligocene east Asian summer monsoon in the NE Tibetan Plateau evidence from a palynological record from the Lanzhou Basin, China. Journal of Asian Earth Sciences, 75:46–57.
https://doi.org/10.1016/j.jseaes.2013.07.003
Miller, K.G., Wright, J.D., and Fairbanks, R.G. 1991. Unlocking the icehouse Oligocene–Miocene oxygen isotopes eustasy and margin erosion. Journal of Geophysical Research: Solid Earth, 96(B4):6829–6848.
https://doi.org/10.1029/90JB02015
Miller, K.G., Browning, J.V., Schmelz, W.J., Kopp, R.E., Mountain, G.S., and Wright, J.D. 2020. Cenozoic sea-level and cryospheric evolution from deep-sea geochemical and continental margin records. Science advances, 6(20):eaaz1346.
https://doi.org/10.1126/sciadv.aaz1346
Moreno-Domínguez, R., Postigo-Mijarra, J.M., and Barrón, E. 2021. Palaeoclimatic reconstruction for the late Oligocene La Val fossil site Estadilla Huesca Spain based on CLAMP and LMA. Palaeogeography, Palaeoclimatology, Palaeoecology, 567: 110302.
https://doi.org/10.1016/j.palaeo.2021.110302
Mosbrugger, V., Gee, C.T., Belz, G., and Ashraf, A.R. 1994. Three-dimensional reconstruction of an in-situ Miocene peat forest from the Lower Rhine Embayment northwestern Germany new methods in palaeovegetation analysis. Palaeogeography, Palaeoclimatology, Palaeoecology, 110(3-4):295–317.
https://doi.org/10.1016/0031-0182(94)90089-2
Mosbrugger, V. and Utescher, T. 1997. The coexistence approach, a method for quantitative reconstructions of Tertiary terrestrial palaeoclimate data using plant fossils. Palaeogeography, Palaeoclimatology, Palaeoecology, 134(1-4):61–86.
https://doi.org/10.1016/S0031-0182(96)00154-X
Murray, J.W. 1992. Palaeogene and Neogene. Geological Society, London, Memoirs, 13(1):141–147.
O’Brien, C.L., Huber, M., Thomas, E., Pagani, M., Super, J.R., Elder, L.E., and Hull, P.M. 2020. The enigma of Oligocene climate and global surface temperature evolution. Proceedings of the National Academy of Sciences USA, 117(41):25302–25309.
https://doi.org/10.1073/pnas.2003914117
Pälike, H., Norris, R.D., Herrle, J.O., Wilson, P.A., Coxall, H.K., Lear, C.H., Shackleton, N.J., Tripati, A.K., and Wade, B.S. 2006. The heartbeat of the Oligocene climate system. Science, 314(5807):1894–1898.
https://doi.org/10.1126/science.1133822
Pattyn, F., Ritz, C., Hanna, E., Asay-Davis, X., DeConto, R., Durand, G., Favier, L., Fettweis, X., Goelzer, H., Golledge, N.R., and Kuipers Munneke, P. 2018. The Greenland and Antarctic ice sheets under 1.5°C global warming. Nature Climate Change, 8(12):1053–1061.
https://doi.org/10.1038/s41558-018-0305-8
Peel, M.C., Finlayson, B.L., and McMahon, T.A. 2007. Updated world map of the Köppen-Geiger climate classification. Hydrology and Earth System Sciences, 11(5):1633–1644.
https://doi.org/10.5194/hess-11-1633-2007
Pekar, S. and Miller, K.G. 1996. New Jersey Oligocene icehouse sequences ODP Leg 150X correlated with global δ18O and Exxon eustatic records. Geology, 24(6):567–570.
https://doi.org/10.1130/0091-7613(1996)024<0567:NJOISO>2.3.CO;2
Pekar, S.F., Christie-Blick, N., Kominz, M.A., and Miller, K.G. 2002. Calibration between eustatic estimates from backstripping and oxygen isotopic records for the Oligocene. Geology, 30(10):903–906.
https://doi.org/10.1130/0091-7613(2002)030<0903:CBEEFB>2.0.CO;2
Pope, J.O., Brown, K., Fung, F., Hanlon, H.M., Neal, R., Palin, E.J., and Reid, A. 2021. Investigation of future climate change over the British Isles using weather patterns. Climate Dynamics, 58(9-10):2405–2419.
https://doi.org/10.1007/s00382-021-06031-0
Pound, M.J. and Riding, J.B. 2016. Palaeoenvironment, palaeoclimate and age of the Brassington Formation Miocene of Derbyshire UK. Journal of the Geological Society, 173(2):306–319.
https://doi.org/10.1144/jgs2015-050
Pound, M.J. and Salzmann, U. 2017. Heterogeneity in global vegetation and terrestrial climate change during the late Eocene to early Oligocene transition. Scientific Reports, 7(1):43386.
https://doi.org/10.1038/srep43386
Pound, M.J. and McCoy, J. 2021. Palaeoclimate reconstruction and age assessment of the Miocene flora from the Trwyn y Parc solution pipe complex of Anglesey Wales UK. Palynology, 45(4):697–703.
https://doi.org/10.1080/01916122.2021.1916636
Previdi, M. and Liepert, B.G. 2007. Annular modes and Hadley cell expansion under global warming. Geophysical Research Letters, 34(22):L22701.
https://doi.org/10.1029/2007GL031243
Pross, J., Bruch, A., and Kvaček, Z. 1998. Paläoklima-Rekonstruktion für den Mittleren Rupelton Unter-Oligozän des Mainzer Beckens auf der Basis mikro und makrobotanischer Befunde. Mainzer Geowissenschaftliche Mitteilungen., 27:79–92.
Pross, J., Bruch, A.A., Mosbrugger, V., and Kvacek, Z. 2001. Palaeogene pollen and spores as a tool for quantitative palaeoclimate reconstructions the Rupelian Oligocene of central Europe. Conference Proceedings of the IX International Palynological Congress, Houston, TX, USA, p. 299–310.
Quan, C., Liu, Y.S.C., Tang, H., and Utescher, T. 2014. Miocene shift of European atmospheric circulation from trade wind to westerlies. Scientific Reports, 4(1):1–6.
https://doi.org/10.1038/srep05660
Quicke, D.L. 1987. The old-world genera of braconine wasps Hymenoptera: Braconidae. Journal of Natural History, 21(1):43–157.
https://doi.org/10.1080/00222938700770031
Rind, D. 1998. Latitudinal temperature gradients and climate change. Journal of Geophysical Research: Atmospheres, 103(D6):5943–5971.
Roberts, D.G. 1975. Marine geology of the Rockall Plateau and Trough. Philosophical Transactions of the Royal Society of London Series A Mathematical and Physical Sciences, 278(1285):447–509.
https://doi.org/10.1098/rsta.1975.0033
Roth-Nebelsick, A., Utescher, T., Mosbrugger, V., Diester-Haass, L., and Walther, H. 2004. Changes in atmospheric CO2 concentrations and climate from the late Eocene to early Miocene palaeobotanical reconstruction based on fossil floras from Saxony Germany. Palaeogeography, Palaeoclimatology, Palaeoecology, 205(1-2):43–67.
https://doi.org/10.1016/j.palaeo.2003.11.014
Scheffer, M., Brovkin, V., and Cox, P.M. 2006. Positive feedback between global warming and atmospheric CO2 concentration inferred from past climate change. Geophysical research letters, 33(10):L10702.
https://doi.org/10.1029/2005GL025044
Scotese, C. R. and Wright, N. 2018. PALEOMAP Paleodigital Elevation Models (PaleoDEMS) for the Phanerozoic PALEOMAP Project. Available at:
https://www.earthbyte.org/paleodem-resourcescotese-and-wright-2018/
Screen, J.A. and Simmonds, I. 2010. The central role of diminishing sea ice in recent Arctic temperature amplification. Nature, 464(7293):1334–1337.
Seidel, D.J., Fu, Q., Randel, W.J., and Reichler, T.J. 2008. Widening of the tropical belt in a changing climate. Nature Geoscience, 1(1):21–24.
https://doi.org/10.1038/ngeo.2007.38
Sharmila, S. and Walsh, K.J.E. 2018. Recent poleward shift of tropical cyclone formation linked to Hadley cell expansion. Nature Climate Change, 8(8):730–736.
https://doi.org/10.1038/s41558-018-0227-5
Shukla, A., Mehrotra, R.C., Spicer, R.A., Spicer, T.E., and Kumar, M. 2014. Cool equatorial terrestrial temperatures and the South Asian Monsoon in the early Eocene evidence from the Gurha Mine Rajasthan India. Palaeogeography, Palaeoclimatology, Palaeoecology, 412:187–198.
https://doi.org/10.1016/j.palaeo.2014.08.004
Simms, M.J. and Boulter, M.C. 2000. Oligocene cave sediments in County Cork implications for reconstructing the Tertiary landscape of southwest Ireland. Proceedings of the Geologists' Association, 111(4):363–372.
https://doi.org/10.1016/S0016-7878(00)80092-9
Śliwińska, K.K., Clausen, O.R., and Heilmann-Clausen, C. 2010. A mid-Oligocene cooling Oi-2b reflected in the dinoflagellate record and in depositional sequence architecture an integrated study from the eastern North Sea Basin. Marine and Petroleum Geology, 27(7):1424–1430.
https://doi.org/10.1016/j.marpetgeo.2010.03.008
Śliwińska, K.K. and Heilmann-Clausen, C. 2011. Early Oligocene cooling reflected by the dinoflagellate cyst Svalbardella cooksoniae. Palaeogeography, Palaeoclimatology, Palaeoecology, 305(1-4):138–149.
https://doi.org/10.1016/j.palaeo.2011.02.027
Śliwińska, K.K., Dybkjær, K., Schoon, P.L., Beyer, C., King, C., Schouten, S., and Nielsen, O.B. 2014. Paleoclimatic and paleoenvironmental records of the Oligocene–Miocene transition central Jylland Denmark. Marine Geology, 350:1–15.
https://doi.org/10.1016/j.margeo.2013.12.014
Srivastava, G., Spicer, R.A., Spicer, T.E., Yang, J., Kumar, M., Mehrotra, R., and Mehrotra, N. 2012. Megaflora and palaeoclimate of a late Oligocene tropical delta Makum Coalfield Assam evidence for the early development of the South Asia Monsoon. Palaeogeography, Palaeoclimatology, Palaeoecology, 342:130–142.
https://doi.org/10.1016/j.palaeo.2012.05.002
Steinthorsdottir, M., Coxall, H.K., De Boer, A.M., Huber, M., Barbolini, N., Bradshaw, C.D., Burls, N.J., Feakins, S.J., Gasson, E., Henderiks, J., and Holbourn, A.E. 2021. The Miocene the future of the past. Paleoceanography and Paleoclimatology, 36(4):2020PA004037.
https://doi.org/10.1029/2020PA004037
Straume, E.O., Nummelin, A., Gaina, C., and Nisancioglu, K.H. 2022. Climate transition at the Eocene-Oligocene influenced by bathymetric changes to the Atlantic-Arctic oceanic gateways. Proceedings of the National Academy of Sciences, USA, 119(17):e2115346119.
https://doi.org/10.1073/pnas.2115346119
Stuchlik, L., Ziembińska-Tworzydło, M., Kohlman-Adamska, A., Grabowska, I., Ważyńska, B., Słodkowska, B., and Sadowska, A. 2001.Volume 1 - Spores. In Stuchlik, L. (ed.), Atlas of pollen and spores of the Polish Neogene, W. Szafer Institute of Botany, Polish Academy of Sciences, Kraków, Poland.
Stuchlik, L., Ziembińska-Tworzydło, M., Kohlman-Adamska, A., Grabowksa, I., Ważyńska, H., Słodkowska, B., and Sadowska, A. 2002. Volume 2 - Gymnosperms. In series: Stuchlik, L. (ed.) Atlas of pollen and spores of the Polish Neogene, W. Szafer Institute of Botany, Polish Academy of Sciences, Kraków, Poland.
Stuchlik, L., Ziembińska-Tworzydło, M., Kohlman-Adamska, A., Grabowska, I., Słodkowska, B., Ważyńska, H., and Sadowska, A. 2009. Volume 3 - Angiosperms (1). In Stuchlik, L. (ed.), Atlas of pollen and spores of the Polish Neogene, W. Szafer Institute of Botany, Polish Academy of Sciences, Kraków, Poland.
Stuchlik, L., Ziembińska-Tworzydło, M., Kohlman-Adamska, A., Grabowska, I., Słodkowska, B., Worobiec, E., and Durska, E. 2014. Volume 4 - Angiosperms (2) In Stuchlik, L. (ed.), Atlas of pollen and spores of the Polish Neogene, W. Szafer Institute of Botany, Polish Academy of Sciences, Kraków, Poland.
Tappin, D.R., Chadwick, R.A., Jackson, A.A., Wingfield, R.T.R., and Smith, N.J.P. 1994. The geology of Cardigan Bay and the Bristol Channel British Geological Survey. HMSO, London.
The Mathworks Inc. 2021. MATLAB version 9.10.1649659 (R2021a) Update 1, Mapping Toolbox Version 5.1The MathWorks Inc 2021, Natick, Massachusetts.
Torricelli, S., Menichetti, E., and Knezaurek, G. 2022. Climate-driven dinoflagellate cyst stratigraphy of the Oligocene-lower Miocene turbidite succession of the Lower Congo Basin offshore NW Angola. Journal of African Earth Sciences, 192:104536.
https://doi.org/10.1016/j.jop.2022.03.004
Utescher, T., Bruch, A., and Mosbrugger, V. 2024. Documentation and Data (Version 2024), records accessed 2023. The Palaeoflora Database (Data set). Zenodo.
https://doi.org/10.5281/zenodo.10881069
Utescher, T., Bruch, A.A., Erdei, B., François, L., Ivanov, D., Jacques, F.M.B., Kern, A.K., Mosbrugger, V., and Spicer, R.A. 2014. The coexistence approach theoretical background and practical considerations of using plant fossils for climate quantification. Palaeogeography, Palaeoclimatology, Palaeoecology, 410:58–73.
https://doi.org/10.1016/j.palaeo.2014.05.031
Utescher, T., Erdei, B., François, L., Henrot, A.J., Mosbrugger, V., and Popova, S. 2021. Oligocene vegetation of Europe and western Asia diversity change and continental patterns reflected by plant functional types. Geological Journal, 56(2):628–649.
https://doi.org/10.1002/gj.3830
Van Simaeys, S. 2004. The Rupelian–Chattian boundary in the North Sea Basin and its calibration to the international timescale. Netherlands Journal of Geosciences, 83(3):241–248.
https://doi.org/10.1017/S0016774600020308
Van Simaeys, S., Munsterman, D., and Brinkhuis, H. 2005. Oligocene dinoflagellate cyst biostratigraphy of the southern North Sea Basin. Review of Palaeobotany and Palynology, 134(1-2):105–128.
https://doi.org/10.1016/j.revpalbo.2004.12.003
Velitzelos, D., Bouchal, J.M., and Denk, T. 2014. Review of the Cenozoic floras and vegetation of Greece. Review of Palaeobotany and Palynology, 204:56–117.
https://doi.org/10.1016/j.revpalbo.2014.02.006
Via, R.K. and Thomas, D.J. 2006. Evolution of Atlantic thermohaline circulation early Oligocene onset of deep-water production in the North Atlantic. Geology, 34(6):441–444.
https://doi.org/10.1130/G22545.1
Von der Heydt, A. and Dijkstra, H.A. 2006. Effect of ocean gateways on the global ocean circulation in the late Oligocene and early Miocene. Paleoceanography, 21(1):PA1011.
https://doi.org/10.1029/2005PA001149
Wade, B.S. and Pälike, H. 2004. Oligocene climate dynamics. Paleoceanography, 19(4): PA4019.
https://doi.org/10.1029/2004PA001042
Wade, B.S., Houben, A.J., Quaijtaal, W., Schouten, S., Rosenthal, Y., Miller, K.G., Katz, M.E., Wright, J.D., and Brinkhuis, H. 2012. Multiproxy record of abrupt sea-surface cooling across the Eocene-Oligocene transition in the Gulf of Mexico. Geology, 40(2):159–162.
https://doi.org/10.1130/G32577.1
Walsh, P. 2001. The Palaeogeography of the southern half of the British Isles and adjacent Continental Shelf at the Palaeogene/Neogene (g/n) boundary and its subsequent modification: a reconsideration, PhD thesis, Wydawnictwo Uniwersytetu Śląskiego, Katowice, Poland.
Walsh, P.T., Atkinson, K., Boulter, M.C., and Shakesby, R.A. 1987. The Oligocene and Miocene outliers of west Cornwall and their bearing on the geomorphological evolution of Oldland Britain. Philosophical Transactions of the Royal Society of London Series A Mathematical and Physical Sciences:211–245.
Watts, W.A. 1962. Early Tertiary pollen deposits in Ireland. Nature, 193(4815):600.
Wilkinson, G.C. and Boulter, M.C. 1980. Oligocene pollen and spores form the western part of the British Isles. Palaeontographica Abteilung B, 175B(1-3):27–83.
Wilkinson, G.C., Bazley, R.A.B., and Boulter, M.C. 1980. The geology and palynology of the Oligocene Lough Neagh clays Northern Ireland. Journal of the Geological Society, 137(1):65–75.
https://doi.org/10.1144/gsjgs.137.1.0065
Wilson, G.S., Pekar, S.F., Naish, T.R., Passchier, S., and DeConto, R. 2008. The Oligocene Miocene boundary Antarctic climate response to orbital forcing. Developments in Earth and Environmental Sciences, 8:369–400.
https://doi.org/10.1016/S1571-9197(08)00009-8
Wilson, S.J. 1996. High resolution comparative palynostratigraphy and palaeoecology of Oligocene sequences in the terrestrial basins of the western British Isles and the marine North Sea Basin. PhD thesis, University of Sheffield, Sheffield, UK.
Woodland, A.W. 1971. The Llanbedr (Mochras Farm) borehole - Report 71/18, Institute of Geological Sciences, HMSO, London.
Woods, M.A., Vandenbroucke, T.R.A., Williams, M., Riding, J.B., De Schepper, S., and Saabe, K. 2014. Complex response of dinoflagellate cyst distribution patterns to cooler early Oligocene oceans. Earth-Science Reviews, 138:215–230.
https://doi.org/10.1016/j.earscirev.2014.02.004
Xiao, G.Q., Abels, H.A., Yao, Z.Q., Dupont-Nivet, G., and Hilgen, F.J. 2010. Asian aridification linked to the first step of the Eocene-Oligocene climate Transition (EOT) in obliquity-dominated terrestrial records Xining Basin China. Climate of the Past, 6(4):501–513.
https://doi.org/10.5194/cp-6-501-2010
Zachos, J.C., Flower, B.P., and Paul, H. 1997. Orbitally paced climate oscillations across the Oligocene /Miocene boundary. Nature, 388(6642):567–570.
https://doi.org/10.1038/41528
Zachos, J.C., Pagani, M., Sloan, L., Thomas, E., and Billups, K. 2001a. Trends, rhythms, and aberrations in global climate 65 Ma to present. Science, 292(5517):686–693.
https://doi.org/10.1126/science.1059412
Zachos, J.C., Shackleton, N.J., Revenaugh, J.S., Palike, H., and Flower, B.P. 2001b. Climate response to orbital forcing across the Oligocene–Miocene boundary. Science, 292(5515):274–278. https://doi.org/10.1126/science.1058288
Zachos, J.C. and Kump, L.R. 2005. Carbon cycle feedbacks and the initiation of Antarctic glaciation in the earliest Oligocene. Global and Planetary Change, 47(1):51–66.
https://doi.org/10.1016/j.gloplacha.2005.01.001
Zanazzi, A., Kohn, M.J., MacFadden, B.J., and Terry, D.O. 2007. Large temperature drop across the Eocene-Oligocene transition in central North America. Nature, 445(7128):639–642.
https://doi.org/10.1038/nature05551
