

 Following their footsteps: Report of vertebrate fossil tracks from John Day Fossil Beds National Monument, Oregon, USA
Following their footsteps: Report of vertebrate fossil tracks from John Day Fossil Beds National Monument, Oregon, USA
Article number: 28.1.a11
https://doi.org/10.26879/1413
Copyright Paleontological Society, February 2025
Author biographies
Plain-language and multi-lingual abstracts
PDF version
Submission: 1 June 2024. Acceptance: 20 February 2025.
ABSTRACT
John Day Fossil Beds National Monument (JODA) preserves an exceptional geologic record stretching from the upper Eocene to the upper Miocene (50-6 Ma) of Oregon. These strata contain countless well-preserved vertebrate body fossils for which JODA is famous. While great emphasis has been placed on the study of the body fossils of JODA, comparatively little work has been done on vertebrate and invertebrate tracks recovered from the area. Four specimens with vertebrate tracks are known from JODA, which detail the behaviors of at least four separate types of trackmaker. These include avian and possible lacertiform reptile tracks from the Eocene upper Clarno Formation (50-39 Ma) and superimposed feliform tracks and a tridactyl ungulate track from the Oligocene Turtle Cove Member of the John Day Formation (31-26 Ma). We used commercially available software (Agisoft Metashape Professional) to produce 3-dimensional and digital elevation models (DEMs) of each track surface. The models allow digital preservation and remote study of the specimens, enable an enhanced resolution of surface features, and provide the means to acquire more accurate measurement of the tracks and trackways. The Clarno Formation tracks are the only evidence to suggest that both birds and lizards were present in the John Day Basin during the Eocene, providing a greater knowledge of faunal diversity at this time. The John Day Formation tracks bear resemblance to animals already known from the Turtle Cove Member from body fossils, such as false sabertooth cats (nimravids) and odd-toed ungulates (perissodactyls). Combined with the body fossil record, the JODA tracks provide important insights into the fauna, environment, and behaviors of taxa through the Eocene and Oligocene of Oregon.
Conner J. Bennett. Earth and Environmental Sciences, Utah Tech University, 225 S. University Ave., St. George, Utah 84770, USA. connerjbennett@gmail.com
Nicholas A. Famoso. John Day Fossil Beds National Monument, U.S. National Park Service, 32651 Highway 19, Kimberly OR, 97848, USA and Department of Earth Science, University of Oregon, 1272 University of Oregon Eugene, Oregon 97403, USA. nicholas_famoso@nps.gov
Daniel I. Hembree. Department of Earth and Planetary Sciences, University of Tennessee Knoxville, Knoxville, Tennessee 37996, USA. dhembre2@utk.edu
Keywords: Tracks; Clarno; John Day Formation; Oligocene; Eocene
Final citation: Bennett, Conner J., Famoso, Nicholas A., and Hembree, Daniel I. 2025. Following their footsteps: Report of vertebrate fossil tracks from John Day Fossil Beds National Monument, Oregon, USA. Palaeontologia Electronica, 28(1):a11.
https://doi.org/10.26879/1502
palaeo-electronica.org/content/2025/5469-john-day-fossil-tracks
Copyright: February 2025 Paleontological Society
This is an open access article distributed under the terms of Attribution-NonCommercial-ShareAlike 4.0 International (CC BY-NC-SA 4.0), which permits users to copy and redistribute the material in any medium or format, provided it is not used for commercial purposes and the original author and source are credited, with indications if any changes are made.
creativecommons.org/licenses/by-nc-sa/4.0/
INTRODUCTION
John Day Fossil Beds National Monument (JODA) in central and eastern Oregon is known for body fossils of upper Eocene to upper Miocene mammals which comprise a nearly continuous, 50 million-year record of mammalian evolution (Bestland et al., 1999; Hunt and Stepleton, 2004; Albright et al., 2008; Fremd, 2010; Samuels and Cavin, 2013; Maguire et al., 2018). This abundance of body fossils has caused other fossil types within the monument to remain relatively understudied. Among those are fossil tracks from the Eocene upper Clarno Formation (50-39 Ma) and the Oligocene Turtle Cove Member of the John Day Formation (31-26 Ma). While not prevalent in the John Day basin, fossil tracks provide data on trackmaker behaviors made during the life of the living organism, which cannot be defined in detail by using body fossils alone (Lockley and Hunt, 1995). When used in tandem with the body fossil record, tracks can also improve interpretations of faunal diversity, often providing evidence of ancient organisms in strata where body fossils are rare (Irmis, 2005). Key aspects of fossil tracks and associated trace fossils also provide information on ancient habitats (Hogue, 2018).
Fossil tracks similar to those at JODA have previously been described from the Eocene deposits of the Chuckanut Formation (49.9 Ma) of Washington (Mustoe, 2002) and the Green River Formation (56-38 Ma) of Utah, Wyoming, and Colorado (Moussa, 1968; Scott and Smith, 2015; Hogue, 2018; Lockley et al., 2021). The Chuckanut Formation contains very few body fossils, but a relative abundance of diverse fossil tracks which record the occurrence of several types of birds, turtles, plantigrade and digitigrade perissodactyl mammals. The Green River Formation is also well known for a diverse and abundant supply of fossil tracks (Scott and Smith, 2015) while also preserving numerous, well-preserved body fossils. By using the body fossil record and the fossil track record within the Green River Formation, further conclusions can be drawn concerning the overall diversity of organisms of the Eocene. While work has been done to describe the fossil tracks from both the Chuckanut and Green River formations, the JODA tracks have only been mentioned twice before, with both instances wrongly attributing the John Day Formation tracks to the Clarno Formation and neglecting to mention the actual Clarno Formation tracks entirely (Lockley and Hunt, 1995; Santucci et al., 1998). Due to this inaccurate attribution, it is necessary to properly describe the JODA tracks and attribute them to their proper formations.
In order to describe the JODA tracks, each was rendered as a photogrammetric model to identify surface features which are difficult to discern through visual study alone. Rendered models help to describe each sample and any possible behaviors they preserve, while also digitizing fossil track specimens for remote study (Lallensack et al., 2020). This provides easy access to the data each specimen holds while also assisting in monitoring the condition of the fossil tracks in the future (Lockley et al., 2022).
GEOLOGIC SETTING
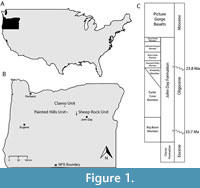 Four formations are recognized within JODA: Clarno (50-39 Ma), John Day (31-25 Ma), Mascall (16-12 Ma), and Rattlesnake (8-6 Ma) (Dillhoff et al., 2009). However, the only tracks recovered have come from the Clarno and John Day formations (Figure 1). The apparent absence of tracks from the Rattlesnake and Mascall formations is likely a result of either the lithology or the depositional environment of each formation. The Rattlesnake Formation is dominated by matrix-rich conglomerates and gravel deposits (Merriam et al., 1925; Samuels and Zancanella, 2011; Samuels and Cavin, 2013), which are not conducive to preservation of fossil tracks. The Mascall Formation possesses layered tuff beds and lacustrine deposits (Maguire et al., 2018), which may be conducive to the preservation of tracks. The apparent absence of tracks within the Mascall Formation may be due to either collection bias or the lacustrine environment represented by these deposits may have been too deep for terrestrial vertebrate tracks to be preserved.
Four formations are recognized within JODA: Clarno (50-39 Ma), John Day (31-25 Ma), Mascall (16-12 Ma), and Rattlesnake (8-6 Ma) (Dillhoff et al., 2009). However, the only tracks recovered have come from the Clarno and John Day formations (Figure 1). The apparent absence of tracks from the Rattlesnake and Mascall formations is likely a result of either the lithology or the depositional environment of each formation. The Rattlesnake Formation is dominated by matrix-rich conglomerates and gravel deposits (Merriam et al., 1925; Samuels and Zancanella, 2011; Samuels and Cavin, 2013), which are not conducive to preservation of fossil tracks. The Mascall Formation possesses layered tuff beds and lacustrine deposits (Maguire et al., 2018), which may be conducive to the preservation of tracks. The apparent absence of tracks within the Mascall Formation may be due to either collection bias or the lacustrine environment represented by these deposits may have been too deep for terrestrial vertebrate tracks to be preserved.
The upper section of the Clarno Formation is composed of mudstones capped by the Member A welded tuff of the Big Basin Member of the John Day Formation, whereas the lower section is composed of lahar deposits with interbedded shales and mudstones (Bestland et al., 1999; Wheeler and Manchester, 2002). The Clarno tracks, JODA 1535 and 1536, were found on pieces of interbedded shales from the lower section. These shales are stratigraphically below lahars, which causes deformation in these beds making it difficult to locate and identify tracks.
The John Day Formation is currently subdivided into seven members: Big Basin, Turtle Cove, Kimberly, Haystack Valley, Balm Creek, Johnson Canyon, and Rose Creek (Retallack et al., 2000; Hunt and Stepelton, 2004; Albright et al., 2008). Two of the specimens described in this study, JODA 251 and 283, are likely from the A/B Tuff of the Turtle Cove Member (29.75 Ma) (Albright et al., 2008). The exact locality of JODA 251 and 283 was not properly documented, leading to their stratigraphic context being inferred as the A/B Tuff. The Turtle Cove Member consists of about 400 m of predominantly volcaniclastic mudstones and volcanic tuffs that Albright et al. (2008) divided into 14 lithostratigraphic subunits (A-K2) with 10 dated tuffs including the Picture Gorge ignimbrite (PGI) - a super-volcanic event bed related to the Yellowstone Hotspot (Seligman et al., 2014). The PGI is between subunits F and G. The Turtle Cove fauna is assigned to the Whitneyan (Wh2) and Arikareen (Ar1 and Ar2) North American Land Mammal Ages (NALMAs).
METHODS
Vertebrate tracks were found within the monument on specimens JODA 1535, 1536, 251 and 283. JODA 1535 and 1536 were found in the Clarno Formation (50-39 Ma). JODA 251 and 283 were found in the Turtle Cove Member of the John Day Formation (31-25 Ma). The Clarno Formation tracks were collected through a Bureau of Land Management funded survey in 1987, and the Turtle Cove Member samples were donated to JODA by Larry McGraw in 1979, shortly after JODA was established in 1975.
Each JODA specimen was documented through rendered photogrammetric models using Agisoft Metashape Professional (version 1.7). Photographs were taken using a Nikon D810 digital camera and Nikon AF Micro NIKKOR 60 mm lens. Models were rendered from photographs taken of each specimen, using standard techniques outlined by Lallensack et al. (2022). Adequate image overlap, having roughly 60-70% overlap between images, is essential to provide Metashape with enough data to link together the entire model. In addition, tracks and tracksites require images to be captured perpendicular to the surface. Shadows within photographs will not register in the model, creating holes in the final product. Because of this, proper lighting of a surface is essential to capture as much detail as possible. Proper positioning and full capture of scale bars within images is required to ensure proper scaling information.
Model types used in this study include 3-Dimensional Mesh and Digital Elevation Models (DEMs). Each surface was rendered as a 3D Mesh model for remote study and accurate measurement. DEM models helped to visualize subtle shifts in surface elevation through color change, allowing small details to be observed. Screenshots taken from photogrammetric models were then measured in ImageJ (version 1.53t) to provide interdigital angle measurements using the techniques from Sarjeant et al. (2002). Pace data were measured using the ruler tool through Agisoft Metashape (version 1.8).
Institutional Abbreviations. JODA = John Day Fossil Beds National Monument, Kimberly, OR, USA; JDNM = Locality number for John Day Fossil Beds National Monument.
RESULTS
JODA 1536
 Material. JODA 1536 preserves two tracks (A1 and A2) from the Clarno Formation (JDNM-66, Hanson’s Bird Tracks) in the Clarno Unit (Wheeler County) of JODA (Figure 2; Table 1, Table 2, Table 3). Also present are four ovoid impressions clustered below A2, as well as several well-preserved sinusoidal trails (Figure 2B). 3D mesh models of JODA 1536 (A) part (https://doi.org/10.17602/M2/M454455) and JODA 1536 (B) counterpart (https://doi.org/10.17602/M2/M454460) are available through MorphoSource.
Material. JODA 1536 preserves two tracks (A1 and A2) from the Clarno Formation (JDNM-66, Hanson’s Bird Tracks) in the Clarno Unit (Wheeler County) of JODA (Figure 2; Table 1, Table 2, Table 3). Also present are four ovoid impressions clustered below A2, as well as several well-preserved sinusoidal trails (Figure 2B). 3D mesh models of JODA 1536 (A) part (https://doi.org/10.17602/M2/M454455) and JODA 1536 (B) counterpart (https://doi.org/10.17602/M2/M454460) are available through MorphoSource.
Description. The tracks are small, with a mean length of 21.2 mm and a mean width of 15.8 mm. The mean angles between digits II-III, III-IV, and II-IV are 28.9°, 30.5°, and 62.2° respectively. The distance between the two tracks is approximately 40.2 mm. Both A1 and A2 lack webbing between digits. Digit claw impressions on A1 are pointed and slightly oblique to the digit direction. A1 is more detailed than A2 because digit claws and pads are difficult to discern in A2.
The ovoid impressions have a short axis between 2-3.5 mm in diameter and a long axis between 2-5 mm in diameter, of which the long axis is parallel to the orientation of A1 and A2 (Figure 2B). Three of the ovoid impressions are clustered roughly 5 mm from digit II of A2, with the smallest of the ovoid impressions outlying this concentration, approximately 18.3 mm from digit II of A2.
The trails have smooth surfaces, possess a meandering sinusoidal morphology, have very shallow depths, and lengths ranging from 1.60-5.25 cm (Figure 2B). The trails are abundant along the margins of the specimen, and less common toward the center.
JODA 1535
 Material. JODA 1535 preserves three possible tracks (L1, L2, L3) from the Clarno Formation (JDNM-66, Hansen’s Bird Tracks) in the Clarno Unit (Wheeler County) of JODA (Figure 3; Table 1, Table 2, Table 3). 3D mesh models of JODA 1535 (A) part (https://doi.org/10.17602/M2/M454449) and JODA 1535 (B) counterpart (https://doi.org/10.17602/M2/M454444) are available on MorphSource.
Material. JODA 1535 preserves three possible tracks (L1, L2, L3) from the Clarno Formation (JDNM-66, Hansen’s Bird Tracks) in the Clarno Unit (Wheeler County) of JODA (Figure 3; Table 1, Table 2, Table 3). 3D mesh models of JODA 1535 (A) part (https://doi.org/10.17602/M2/M454449) and JODA 1535 (B) counterpart (https://doi.org/10.17602/M2/M454444) are available on MorphSource.
Description. L1 preserves the most detail of the three tracks, with five claw-like impressions. Digit V of L1 is significantly smaller and located distally to digits I-IV; digit III is the longest of the impressions (Table 1, Table 2, Table 3). Digit V is oriented almost perpendicular to the long axis of L1, while the remaining digits are roughly parallel to the long axis of L1. The toe claw impressions of L1 curve proximally toward the trackmaker, showing drag marks from motion of the foot during locomotion.
The remaining two impressions are incomplete; L2 only includes three digit impressions (likely from digits II-IV), while L3 preserves only partial impressions of digits I-V. The morphology of L2 and L3 differs from that of L1, in that each digit impression is roughly equidistant and oriented perpendicular to the long axis of the tracks. It is important to note that both L2 and L3 may represent a different trackmaker. No pace data could be collected from L2 and L3 due to the incomplete nature of these tracks.
JODA 283
 Material. JODA 283 preserves four tracks (F1, F2, F3, F4), with F1-F2 and F3-F4 each representing a pair of superimposed impressions, likely from the A/B Tuff of the Turtle Cove Member of the John Day Formation (JDNM-8, Sheep Rock) in the Sheep Rock Unit (Grant County) of JODA (Figure 4, Table 1, Table 2, Table 3). A 3D mesh of JODA 283 is available on MorphoSource (https://doi.org/10.17602/M2/M454313).
Material. JODA 283 preserves four tracks (F1, F2, F3, F4), with F1-F2 and F3-F4 each representing a pair of superimposed impressions, likely from the A/B Tuff of the Turtle Cove Member of the John Day Formation (JDNM-8, Sheep Rock) in the Sheep Rock Unit (Grant County) of JODA (Figure 4, Table 1, Table 2, Table 3). A 3D mesh of JODA 283 is available on MorphoSource (https://doi.org/10.17602/M2/M454313).
Description. The tracks have a mean length of 24.5 mm and a mean width of 30 mm. Mean interdigital angles for digits II-III are 19.7°, III-IV 25.2°, and IV-V 32.2°. Pace data between F1 and F3 is 94.4 mm, while F2 and F4 is 95.3 mm. Each track is composed of one center pad impression and four ovoid digit impressions forming a semicircle in front of the center pad. The long axis of the digital pads points outward away from the center of the track and the ends of each digital pad lack digit claw impressions. The digital pads of F1 and F3 are slightly obscured due to the overprinting from F2 and F4.
JODA 251
 Material. JODA 251 preserves two overlapping tracks, one of which (F5) is superimposed over the other (F6), as well as a single partial track (P1) (Figure 5, Figure 6; Table 1, Table 2, Table 3). This slab is likely from the A/B Tuff of the Turtle Cove Member of the John Day Formation (JDNM-8, Sheep Rock) in the Sheep Rock Unit (Grant County) of JODA. A 3D mesh of JODA 251 is available on MorphoSource (https://doi.org/10.17602/M2/M454308).
Material. JODA 251 preserves two overlapping tracks, one of which (F5) is superimposed over the other (F6), as well as a single partial track (P1) (Figure 5, Figure 6; Table 1, Table 2, Table 3). This slab is likely from the A/B Tuff of the Turtle Cove Member of the John Day Formation (JDNM-8, Sheep Rock) in the Sheep Rock Unit (Grant County) of JODA. A 3D mesh of JODA 251 is available on MorphoSource (https://doi.org/10.17602/M2/M454308).
 Description. The morphology and size of F5 and F6 are similar to the tracks (F1-F4) from JODA 283. F5 has a length of 29.9 mm and a width of 29.3 mm. The width of F6 is 26.5 mm, however, the length cannot be accurately determined due to overprinting by F5. Interdigital angles for F5 are distorted; the only measurable angle is 23.2° between digits IV-V. In F6 the interdigital angles between digits II-III is 18.9°, III-IV is 23.4°, and IV-V is 34.4°.
Description. The morphology and size of F5 and F6 are similar to the tracks (F1-F4) from JODA 283. F5 has a length of 29.9 mm and a width of 29.3 mm. The width of F6 is 26.5 mm, however, the length cannot be accurately determined due to overprinting by F5. Interdigital angles for F5 are distorted; the only measurable angle is 23.2° between digits IV-V. In F6 the interdigital angles between digits II-III is 18.9°, III-IV is 23.4°, and IV-V is 34.4°.
P1 consists of three ovoid pad impressions lacking a central pad. The width of P1 is approximately 119 mm, however, the length of the track cannot be determined due to the absence of an impression of the back of the foot. The interdigital angles between digits II-III is 24.1° and between digits III-IV is 18.6°. The three digits are rounded and ovoid, with no digit claw impressions evident. Digit II and III are noticeably larger than digit IV. Digit IV is also spaced farther from III than digit II is from III.
DISCUSSION
Avian Tracks and Feeding Behavior (JODA 1536)
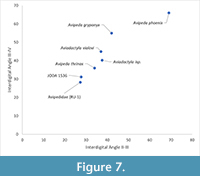 The tracks present on JODA 1536 are interpreted as the first avian fossils from the Eocene of JODA. In order to better understand the possible ichnotype, the divarication angles of A1 and A2 were compared to those of similar ichnotypes including Aviadactyla vialovi, Aviadactyla isp., Avipeda gryponyx, Avipeda phoenix, Avipeda thrinax, and Avipedidae (WWU Site RU-1; Mustoe, 2002) (Figure 7). Of these, A1 and A2 are most similar to the ichnogenus Avipeda because of the absence of webbing within the tracks, the narrow digit divarication angles, and the apparent absence of digit I (Sarjeant and Langston, 1994). However, the lack of further avian track material from JODA, coupled with the incomplete nature of A2, makes it impossible to definitively attribute the ichnogenus of these avian tracks. JODA 1536 is not attributed to ichnotaxa such as Rivavipes, Fuscinapeda, and Charadriipeda, because JODA 1536 possesses slender digits and no evidence of webbing (Sarjeant and Langston, 1994; Mustoe 2002; Mustoe, 2012). The absence of hallux impressions does not allow for JODA 1536 to be attributed to ichnotaxa such as Gruipeda and Ardeipeda.
The tracks present on JODA 1536 are interpreted as the first avian fossils from the Eocene of JODA. In order to better understand the possible ichnotype, the divarication angles of A1 and A2 were compared to those of similar ichnotypes including Aviadactyla vialovi, Aviadactyla isp., Avipeda gryponyx, Avipeda phoenix, Avipeda thrinax, and Avipedidae (WWU Site RU-1; Mustoe, 2002) (Figure 7). Of these, A1 and A2 are most similar to the ichnogenus Avipeda because of the absence of webbing within the tracks, the narrow digit divarication angles, and the apparent absence of digit I (Sarjeant and Langston, 1994). However, the lack of further avian track material from JODA, coupled with the incomplete nature of A2, makes it impossible to definitively attribute the ichnogenus of these avian tracks. JODA 1536 is not attributed to ichnotaxa such as Rivavipes, Fuscinapeda, and Charadriipeda, because JODA 1536 possesses slender digits and no evidence of webbing (Sarjeant and Langston, 1994; Mustoe 2002; Mustoe, 2012). The absence of hallux impressions does not allow for JODA 1536 to be attributed to ichnotaxa such as Gruipeda and Ardeipeda.
Originally, the ovoid impressions on JODA 1536 were considered raindrop impressions. Raindrop impressions are found widely dispersed over a bedding surface, oftentimes with impressions occurring within each other (Metz, 1981). The JODA 1536 ovoid impressions are localized to the region near A2 and do not occur within each other. Because the JODA 1536 ovoid impressions lack these general aspects of raindrop impressions, we interpret the numerous ovoid impressions near A2 as beak impressions of the avian trackmaker. This feeding behavior is shared between numerous modern shorebird species which consume various worms, insects, crustaceans, and molluscs on and within the substrate (Van de Kam, 2017). Invertebrate traces attributed to Cochlichnus (Figure 2) occurring on JODA 1536 provide evidence of possible prey for the trackmaker and suggest the tracks were made along a lakeshore environment likey in very shallow water (Hogue, 2018). Previously described “foot-stirring” and beak-probing behaviors have been identified from the Cretaceous (Lockley et al., 2009; Falk et al., 2010). Erickson (1967) also described a bird feeding trace from the Eocene of Utah from a larger, web-footed bird. Zonneveld et. al. (2024) also recently described numerous circular to ovoid impressions in proximity to avian trackways from the Eocene Tanjung Formation of Indonesia. Due to the commonality between modern shorebird feeding behavior and that inferred from JODA 1536, this feeding process is successful in sourcing a variety of food types and has been utilized by shorebirds for millions of years. As there are few examples of feeding traces of birds from the Eocene, JODA 1536 provides an important understanding of Eocene shorebird feeding behaviors.
Avian and invertebrate traces are likely to occur within a near-shore, lacustrine environment. The lithology of the JODA 1536 sample slab supports this assumption, as it has been classified as a section of interbedded shale within the otherwise lahar-rich geology of the Clarno Formation (Wheeler and Manchester, 2002). Because shale deposits are not dominant within the Clarno, JODA 1536 is therefore important in possessing traces, which would have otherwise been impossible to preserve within the more dominant lahar deposits. Therefore, the lithology and fossil traces of the JODA 1536 slab provide a greater understanding of this Eocene ecosystem, suggesting interaction between avian and invertebrate taxa along a nearshore, lacustrine environment.
Reptile Tracks (JODA 1535)
The morphology of L1 in JODA 1535 is similar to tracks produced by reptiles, especially those of extant lizards (Kubo, 2010). This is significant because few Eocene reptilian ichnotaxa have been described. Unfortunately, the incomplete preservation of the JODA 1535 tracks prevents attribution to any specific ichnotaxon.
Sarjeant and Langston (1994) described a trackway from the Chadronian (Late Eocene) of Texas, which they attributed to the ichnogenus Chelonipus and interpreted as being produced by an amphibious turtle. Mustoe (2019) described similar Chelonipus trackways from the Lower Eocene of Washington as well as the new ichnogenus Anticusuchipes which they attributed to a crocodilian trackmaker. Lizard-like tracks are also known from Eocene deposits but have not been described in any detail. For example, the Green River Formation of Utah, Colorado, and Wyoming has one documented trackway (Curry, 1957) and one isolated track (Lockley et al., 2021) with morphologies similar to L1.
Track L1 in JODA 1535 possesses thin digits with pronounced, sharp digit claw impressions. This eliminates dactyloidiform, helodermatiform, iguaniform, and crocodyliform traces from attribution to JODA 1535. The claw marks, thin digits, and digit orientation (I-V) of L1 are more similar to Neosauroides which are interpreted as lacertiform tracks from the Lower Cretaceous of Korea (Kim et al., 2017, 2019). These features have also been described from tracks produced by various extant lizards in neoichnological experiments (Kubo 2010). The absence of similar Eocene track descriptions makes JODA 1535 an essential sample to base future research into Eocene lacertiform fossil tracks. If correctly attributed, JODA 1535 would be the first lacertiform fossil from JODA.
Feliform Tracks (JODA 283, 251)
We attribute JODA 283 and 251 (F5-F6) to a feliform trackmaker because of the presence of four spheroid to ovoid digits with a semicircular center pad and the absence of claw impressions, a trait primarily possessed in felids which discerns them from canid tracks (Remeika, 2001). Feliform body fossils are well known from the John Day Formation and include the nimravids Dinaelurus, Dinictis, Hoplophoneus (= Eusmilus), Nimravus, and Pogonodon, and the palaeogalid Palaeogale (Fremd, 2010; Barrett, 2016; Barrett et al., 2021; Famoso and Orcutt, 2022; Barrett and Hopkins, 2024). While matching the JODA feliform tracks to any one of these taxa is impossible, the small size of the tracks compares well with the nimravid Hoplophoneus (= Eusmilus) cerebralis, a small, bobcat-sized nimravid of the Oligocene John Day Formation (Barrett, 2016).
Currently, four ichnogenera belong to the morphofamily Felipedidae: Felipeda (Panin and Avram, 1962), Mitsupes (Rodríguez-de la Rosa and Guzmán-Gutiérrez, 2012), Pantheraichnus (Diedrich, 2011), and Pycnodactylopus (Sarjeant et al., 2002). JODA 283 and JODA 251 (F5-F6) tracks closely resemble the ichnogenus Felipeda based on the symmetrical nature of the center pad, the overall “U” shape of the center pad, and the ratio of digit width to length exhibited in the JODA feliform tracks (specifically digits III and IV) (Anton et al., 2004; Agnolin et al., 2018). The ichnogenus Mitsupes is characterized by large center pads in the shape of a “U”, but asymmetrically oriented to the digits, a trait not shared with the symmetrical center pads from the JODA feliform tracks. The ichnogenus Pantheraichnus possess center pads more plantigrade than those of the JODA tracks, in some cases being partially or completely ovoid in shape, as opposed to the “U” shape exhibited by the JODA feliform tracks (Diedrich, 2011). The ichnogenus Pycnodactylopus is a broad, overall ovoid track with digits having widths two-thirds their length, much thicker than the digit width ratio of the JODA tracks (approximately one-third).
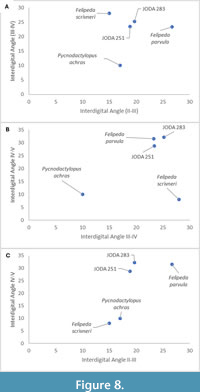 To better understand how JODA 283 and JODA 251 (F5-F6) compared to other feliform ichnotaxa, we compared the interdigital angles of the JODA impressions to the ichnospecies Felipeda parvula, Felipeda scrivneri, and Pycnodactylopus achras (Figure 8 A-B) (Sarjeant et al., 2002). Of the ichnospecies compared, JODA 283 and JODA 251 (F5-F6) are most similar to Felipeda parvula. However, while similar to the ichnogenus Felipeda, the superimposition of the feliform tracks in JODA 283 and JODA 251 (F5-F6) makes this attribution difficult, and therefore assignment to a single ichnogenus is provisional.
To better understand how JODA 283 and JODA 251 (F5-F6) compared to other feliform ichnotaxa, we compared the interdigital angles of the JODA impressions to the ichnospecies Felipeda parvula, Felipeda scrivneri, and Pycnodactylopus achras (Figure 8 A-B) (Sarjeant et al., 2002). Of the ichnospecies compared, JODA 283 and JODA 251 (F5-F6) are most similar to Felipeda parvula. However, while similar to the ichnogenus Felipeda, the superimposition of the feliform tracks in JODA 283 and JODA 251 (F5-F6) makes this attribution difficult, and therefore assignment to a single ichnogenus is provisional.
Odd-Toed Ungulate Track (JODA 251)
The three round impressions on JODA 251 (P1) resemble known Miocene perissodactyl track types (Vyalov, 1966; Sarjeant and Langston, 1994; Mustoe and Hopkins, 2012; Vera and Krapovickas, 2022). Numerous perissodactyl trackmakers are known from the John Day Formation, including equids such as Miohippus annectens and Mesohippus sp. (Fremd, 2010; Famoso, 2017), rhinocerotids such as Diceratherium (Albright et al., 2008) and tapirids such as Nexuotapirus (Dingus, 1990; Fremd, 2010).
Ichnotaxa likely made by rhinocerotids include Rhinoceripeda (Vyalov, 1966), Palaeotheriipus (Ellenberger, 1980), and Dicerotinichnus (Guerin and Demathieu, 1993). The primary components of rhinocerotid tracks include three small, rounded digit impressions with occasional preservation of a large central pad that corresponds to the sole of the foot (Sarjeant and Langston, 1994). The absence of a center pad within P1 of JODA 251 does not affect determination of the possible ichnotaxon, as some rhinocerotid tracks can lack this feature (Costeur et al., 2009). This absence may suggest that P1 is an undertrack, causing only the front digits to be preserved. However, the digit impressions from P1 include an overall greater length to width ratio in digits II and III, an aspect which is not shared in rhinocerotid impressions (Vyalov, 1966; Ellenberger, 1980; Guerin and Demathieu, 1993).
An ichnogenus associated with equids includes Hippipeda (Vyalov, 1966). This ichnogenus was attributed to hoofed ungulates after Vyalov (1965) amended the ichnogenus from the previous name, Hippichnis. These equid tracks lack digit impressions, composed solely of a convexly arched center pad similar to the hoof of the trackmaker (Lucas, 2007). Miohippus, a functionally three-toed taxon with a center pad, is smaller than a rhino or tapir (Famoso, 2017, McHorse et al., 2017, Janis and Bernor, 2019, McHorse et al., 2019). The presence of only digit impressions from P1 does not conclusively rule out equid ichnotaxa, but this track does not match the described morphology of known ichnotaxa, as Hippichnis does not possess digit impressions.
Tapirid ichnogenera include Apoxypus (Sarjeant and Langston, 1994) and Oplidctylapes (Mustoe and Hopkins, 2012). Each of these ichnotaxa have four-digit manus and three-digit pes impressions; pes digit impressions have overall greater length to width ratios, as do digits II and III of P1. This similar ratio suggests a tapirid trackmaker is more likely than other perissodactyls. It is important to note, however, that P1 is likely an undertrack, causing distortion of the digits and absence of the center section of the track. If attributed as tapiriform, JODA 251 (P1) would be one of only three tapirid fossils from the John Day Formation. However, the incomplete nature of P1 and the lack of similar tracks in the region provides insufficient evidence to attribute this track to a known ichnogenus.
CONCLUSIONS
Fossil tracks found within JODA detail significant data in helping to understand the fauna of both the Eocene and Oligocene of Oregon. While most of the JODA samples are incomplete and can be impossible to accurately assign to known ichnogenera, these trace fossils aid in understanding the behaviors and diversity of fauna during their respective periods. JODA 1536 is especially significant, as it records the behavior of Eocene lacustrine shorebirds. In addition, the presence of possible lacertiform traces from JODA 1535 adds to our understanding of known fauna within the Eocene of Oregon. Environmental conditions from the Clarno shales can be inferred from JODA 1535 and 1536, which suggest a nearshore, lacustrine area with low flow velocity. The presence of many invertebrate traces, small shorebird feeding traces, and potential small lizard traces all are suggestive of organisms which would likely inhabit such an environment. For these reasons, the JODA traces can serve to identify aspects of the upper Eocene environment of Oregon. The traces from JODA 283 and 251 (F5-F6) provide a link between feliform trace and body fossil taxa within JODA. In addition, JODA 251 (P1) may further support the presence of tridactyl trackmaker within the Oligocene of Oregon. While future study of the formations within JODA may present further fossil track material, the current JODA tracks provide important insights into the fauna, environment, and behaviors of taxa through the Eocene and Oligocene of Oregon.
ACKNOWLEDGEMENTS
Work was conducted by CJB while a Scientist-In-Parks Intern at JODA. We acknowledge C. Schierup and J. Cavin (JODA) for assistance with the specimens and associated records and archives. We would also like to thank J. Wood (NPS) for assistance with photogrammetry. Funding for this project was provided by the National Park Service. We acknowledge that these specimens were found at sites on the traditional territory of the Confederated Tribes of the Umatilla Indian Reservation (Cayuse, Umatilla, and Walla Walla) and on the traditional territory of the Confederated Tribes of Warm Springs (Wasco, Warm Springs, and Paiute) and the Burns Paiute Tribe.
REFERENCES
Agnolin, F.L., Chimento, N.R., Campo, D.H., Magnussen, M., Boh, D., and De Cianni, F. 2018. Large carnivore footprints from the Late Pleistocene of Argentina. Ichnos, 26(2):119–126.
https://doi.org/10.1080/10420940.2018.1479962
Albright III, L., Woodburne, M., Fremd, T., Swisher III, C., MacFadden, B., and Scott, G. 2008. Revised chronostratigraphy and biostratigraphy of the John Day Formation (Turtle Cove and Kimberly Members), Oregon, with implications for updated calibration of the Arikareen North American Land Mammal Age. The Journal of Geology, 116:211–237.
https://doi.org/10.1086/587650
Ando, H., Koike, W., Koda, Y., and Okumura, Y. 2010. Large mammalian fossil footprints discovered from the Miocene Asakawa Formation in Diago Town, Ibaraki. Bulletin of Ibaraki Nature Museum, 13:1–21.
Anton, M., Lopez, G., and Santamaria, R. 2004. Carnivore trackways from the Miocene site of Salinas de Anana (Alava, Spain). Ichnos, 11(3-4):371–384.
https://doi.org/10.1080/10420940490444951
Barrett, P.Z. 2016. Taxonomic and systematic revisions to the North American Nimravidae (Mammalia, Carnivora). PeerJ, 4:e1658.
https://doi.org/10.7717/peerj.1658
Barrett, P.Z., Hopkins, S.S.B., and Price, S.A. 2021. How many sabertooths? Reevaluating the number of carnivoran sabertooth lineages with total evidence Baysian techniques and a novel origin of the Miocene Nimravidae. Journal of Vertebrate Paleontology, 41(1).
https://doi.org/10.1080/02724634.2021.1923523
Barrett, P.Z. and Hopkins, S.S.B. 2024. Mosaic evolution underlies feliform morphological disparity. Proceedings of the Royal Society B, 291:20240756.
https://doi.org/10.1098/rspb.2024.0756
Bestland, E., Hammond, P., Blackwell, D., Kays, M., Retallack, G., and Stimac, J. 1999. Geologic framework of the Clarno Unit, John Day Fossil Beds National Monument, Central Oregon. Oregon Geology, 61(1):3–19.
Costeur, L., Balme, C., and Legal, S. 2009. Early Oligocene mammal tracks from Southeastern France. Ichnos, 16(4):257–267.
https://doi.org/10.1080/10420940902953197
Curry, H.D. 1957. Fossil tracks of Eocene vertebrates, southwestern Uinta Basin, Utah. Guidebook to the Geology of the Uinta Basin. Intermountain Association of Petroleum Geologists, 42–47.
Diedrich, C. 2011. Late Pleistocene steppe lion Panthera leo spelaea (Goldfuss, 1810) footprints and bone records from open air sites in northern Germany - Evidence of hyena-lion antagonism and scavenging in Europe. Quaternary Science Reviews, 30:1883–1906.
https://doi.org/10.1016/j.quascirev.2011.03.006
Dillhoff, R.M., Dillhoff, T.A., Dunn, R.E., Myers, J.A., and Strömberg, C.A. 2009. Cenozoic paleobotany of the John Day Basin, central Oregon. Geological Society of America Field Guide, 15:135–164.
https://doi.org/10.1130/2009.fld015(07)
Dingus, L. 1990. Systematics, stratigraphy, and chronology for mammalian fossils (Late Arikareean to Hemingfordian) from the uppermost John Day Formation, Warm Springs, Oregon. PaleoBios, 12(47/48):1–24.
Ellenberger, P. 1980. Sur les impreintes de pas de gros mammiferes de I'Eocene superieur de Garrigues Ste Eulalie (Gard): Paleovertebrata Montpellier. Memoire Jubilaire R. Lavocat, 37–77. (In French)
Erickson, B. 1967. Fossil bird tracks from Utah. Museum Observer, 5:1–7.
Falk, A., Hasiotis, S., and Martin, L. 2010. Feeding traces associated with bird tracks from the Lower Cretaceous Haman Formation, Republic of Korea. Palaios, 25:730–741.
https://doi.org/10.2110/palo.2010.p10-057r
Famoso, N.A. 2017. Statistical analysis of dental variation in the Oligocene equid Miohippus (Mammalia, Perissodactyla) of Oregon. Journal of Paleontology, 91(5):1060–1068.
https://doi.org/10.1017/jpa.2017.42
Famoso, N.A. and Orcutt, J.D. 2022. First occurrences of Palaeogale von Meyer, 1846 in the Pacific Northwest, United States. Geodiversitas, 44(14):427–436.
https://doi.org/10.5252/geodiversitas2022v44a14
Filhol, H. 1877. Recherches sur les Phosphorites du Quercy. Étude des fossiles qu'on y rencontre et spécialement des mammifères. Annales des sciences géologiques, 1–340. (In French)
Fremd, T.J. 2010. Guidebook: SVP Field Symposium 2010, John Day Basin Field Conference, John Day Fossil Beds National Monument (and Surrounding Basin) Oregon, USA. Society of Vertebrate Paleontology, McLean, Virginia.
Guerin, C. and Demathieu, G. 1993. Empreintes et pistes de Rhinocerotidae (Mammalia, Perissodactyla) du gisement pliocène terminal de Laetoli (Tanzanie). Geobios, 26(4):497–513. (In French)
Häntzschel, W. 1975. Trace fossils and problematica, p. 1–269. In Teichert, C. (ed.), Treatise on Invertebrate Paleontology, Part W. Miscellanea, Supplement I. Geological Society of America and University of Kansas Press, Lawrence.
Hogue, J. 2018. Ichnotaxonomy of the Eocene Green River Formation, Soldier Summit and Spanish Fork Canyon, Unita Basin, Utah: interpreting behaviors, lifestyles, and erecting the Cochlichnus ichnofacies. Unpublished PhD Thesis. University of Kansas, Lawrence, Kansas, USA.
Hunt, R.M. and Stepleton, E. 2004. Geology and paleontology of the upper John Day beds, John Day River Valley, Oregon: lithostratigraphic and biochronologic revision in the Haystack Valley and Kimberly areas (Kimberly and Mt. Misery quadrangles). Bulletin of the American Museum of Natural History, 282:1–90.
https://doi.org/10.1206/0003-0090(2004)282<0001:GAPOTU>2.0.CO;2
Irmis, R.B. 2005. A review of the vertebrate fauna of the Lower Jurassic Navajo Sandstone in Arizona. Mesa Southwest Museum Bulletin, 11:55–71.
Janis, C.M. and Bernor, R.L. 2019. The evolution of equid monodactyly: a review including a new hypothesis. Frontiers in Ecology and Evolution, 7:119.
https://doi.org/10.3389/fevo.2019.00119
Kim, K.S., Lockley, M.G., Lim, J.D., Pinuela, L., and Xing, L. 2017. First report of lacertiform (lizard) tracks from the Cretaceous of Asia. Cretaceous Research, 69:62–70.
https://doi.org/10.1016/j.cretres.2016.08.013
Kim, K.S., Lim, J.D., Lockley, M.G., Kim, D.H., Pinuela, L., and Yoo, J.S. 2019. Largest Cretaceous lizard track assemblage, new morphotypes and longest trackways comprise diverse components of an exceptional Korean Konservat-Lagerstatten ichnofauna. Scientific Reports, 9(1):1–12.
https://doi.org/10.1038/s41598-019-49442-0
Kubo, T. 2010. Extant lizard tracks: variation and implications for paleoichnology. Ichnos, 17(3):187–196.
https://doi.org/10.1080/10420940.2010.502500
Lallensack, J., Buchwitz, M., and Romillo, A. 2022. Photogrammetry in ichnology: 3D model generation, visualization, and data extraction. Journal of Paleontological Techniques, 22:1–18.
https://doi.org/10.31223/X5J30D
Lockley, M. and Hunt, A.P. 1995. Dinosaur tracks and other fossil footprints of the western United States. Columbia University Press, New York.
Lockley, M.G., Chin, K., Houck, K., Matsukawa, M., and Kukihara, R. 2009. New interpretations of Ignotornis, the first-reported Mesozoic avian footprints: implications for the paleoecology and behavior of an enigmatic Cretaceous bird. Cretaceous Research, 30(4):1041–1061.
https://doi.org/10.1016/j.cretres.2009.04.001
Lockley, M.G., Kim, K.S., Lim, J.D., and Romilio, A. 2021. Bird tracks from the Green River Formation (Eocene) of Utah: ichnotaxonomy, diversity, community structure and convergence. Historical Biology, 33(10):2085–2102.
https://doi.org/10.1080/08912963.2020.1771559
Lockley, M.G., Noffke, N., Buntin, C., Breithaupt, B., Matthews, N., and LaMontagne, J. 2022. Dinosaur tracks, microbial mat, and applied photogrammetry: A holistic approach to ichnology and geoheritage at Dinosaur Ridge, Colorado, USA, p. 1–16. In Mahan, K.H. and Carpenter, L. (eds.), Field Excursions in the Front Range and Wet Mountains of Colorado for GSA Connects 2022. Geological Society of America, Boulder, USA.
https://doi.org/10.1130/2022.0064(01)
Lucas, S. 2007. Cenozoic vertebrate footprint ichnotaxa named by O.S. Vyalov in 1965 and 1966. New Mexico Museum of Natural History and Science Bulletin, 42:113–148.
Maguire, K.C., Samuels, J.X., and Schmitz, M.D. 2018. The fauna and chronostratigraphy of the middle Miocene Mascall type area, John Day Basin, Oregon, USA. PaleoBios, 35:1–51.
https://doi.org/10.5070/P9351037578
McHorse, B.K., Biewener, A.A., and Pierce, S.E. 2017. Mechanics of evolutionary digit reduction in fossil horses (Equidae). Proceedings of the Royal Society B: Biological Sciences, 284(1861): 20171174.
https://doi.org/10.1098/rspb.2017.1174
McHorse, B.K., Biewener, A.A., and Pierce, S.E. 2019. The evolution of a single toe in horses: causes, consequences, and the way forward. Integrative and Comparative Biology, 59(3):638–655.
https://doi.org/10.1093/icb/icz050
Merriam, J.C., Stock, C., and Moody, C.L. 1925. The Pliocene Rattlesnake Formation and fauna of eastern Oregon, with notes on the geology of the Rattlesnake and Mascall deposits. Publications, Carnegie Institution of Washington, 347:43–92.
Metz, R. 1981. Why not raindrop impressions? Journal of Sedimentary Research, 51(1):265–268.
https://doi.org/10.1306/212F7C66-2B24-11D7-8648000102C1865D
Moussa, M.T. 1968. Fossil tracks from the Green River Formation (Eocene) near Soldier Summit, Utah. Journal of Paleontology, 42(6):1433–1438.
Mustoe, G.E. 2002. Eocene bird, reptile, and mammal tracks from the Chuckanut Formation, northwest Washington. Palaios, 17(4):403–413.
https://doi.org/10.1669/0883-1351(2002)017<0403:EBRAMT>2.0.CO;2
Mustoe, G.E. 2019. Lower Eocene footprints from northwest Washington, USA. Part 1: Reptile Tracks. Geosciences, 9(7):321.
https://doi.org/10.3390/geosciences9070321
Mustoe, G. and Hopkins, D. 2012. Mammal and bird tracks from the Eocene Puget Group, Northwest Washington, USA. Ichnos, 20:36–42.
https://doi.org/10.1080/10420940.2012.759953
Panin, N. and Avram, E. 1962. Noi urme de vertebrate in Miocenul Subcarpatilor Rominesti, Studii si Cercetari de Geologie, 7:455–484. (In Romanian)
Remeika, P. 1999. Identification, stratigraphy, and age of Neogene vertebrate footprints from the Vallecito-Fish Creek basin, Anza-Borrego Desert State Park, California. San Bernadino County Museum Association Quarterly, 46(2):37–46.
Remeika, P. 2001. The Fish Creek Canyon ichnofauna: a Pliocene (Blancan) vertebrate footprint assemblage from Anza-Borrego Desert State Park, California. Proceedings of the 6th Fossil Resource Conference, Geological Resources Division, Lakewood, Colorado, p. 55.
Retallack, G.J., Bestland, E.A., and Fremd, T.J. 2000. Eocene and Oligocene paleosols of central Oregon. Geological Society of America Special Papers, 344:1–192.
https://doi.org/10.1130/0-8137-2344-2.1
Robinson, P.T., Brem, G.F., and McKee, E.H. 1984. John Day Formation of Oregon: a distal record of early Cascade volcanism. Geology, 12(4):229–232.
Rodríguez-de la Rosa, R. and Guzmán-Gutiérrez, J. 2012. Huellas de aves y mamíferos del Neógeno de Jalisco, México. Paleontología Mexicana, 62:147–158. (In Spanish)
Samuels, J.X., and Cavin, J. 2013. The earliest known fisher (Mustelidae), a new species from the Rattlesnake Formation of Oregon. Journal of Vertebrate Paleontology, 33(2):448–454.
Samuels, J.X. and Zancanella, J. 2011. An early Hemphillian occurrence of Castor (Castoridae) from the Rattlesnake Formation of Oregon. Journal of Paleontology, 85(5):930–935.
https://doi.org/10.1666/11-016.1
Santucci, V.L., Hunt, A.P., and Lockley, M.G. 1998. Fossil vertebrate tracks in National Park Service areas. Dakoterra, 5:107–114.
Sarjeant, W. and Langston, W. 1994. Vertebrate footprints and invertebrate traces from the Chadronian (Late Eocene) of Trans-Pecos Texas. Texas Memorial Museum Bulletin, 36:1–86.
Sarjeant, W., Reynolds, R., and Kissell-Jones, M. 2002. Fossil creodont and carnivore footprints from California, Nevada, and Wyoming. Between the Basins: Exploring the Western Mojave and Southern Basin and Range Province California State University, Fullerton, 37–50.
Scott, J. and Smith, M. 2015. Trace fossils of the Eocene Green River lake basins, Wyoming, Utah, and Colorado, p. 313–350. In Smith, M.E. and Carroll, A.R. (eds.), Stratigraphy and Paleolimnology of the Green River Formation, Western USA. Springer Dordrecht, Heidelberg.
https://doi.org/10.1007/978-94-017-9906-5
Seligman, A.N., Bindeman, I.N., McClaughry, J., Stern, R.A., and Fisher, C. 2014. The earliest low and high δ18 O caldera-forming eruptions of the Yellowstone plume: implications for the 30-40 Ma Oregon calderas and speculations on plume-triggered delaminations. Frontiers in Earth Science, 2.
https://doi.org/10.3389/feart.2014.00034
Van de Kam, J., Ens., B., Piersma, T., and Zwarts, L. 2017. Shorebirds: An Illustrated Behavioral Ecology. Brill, Leiden.
Vera, R.B. and Krapovickas, V. 2022. Paleobiology and paleoecology of ungulates from the Miocene of South America based on an ichnological analysis. Palaeogeography, Palaeoclimatology, Palaeoecology, 606:111247.
https://doi.org/10.1016/j.palaeo.2022.111247
Vyalov, O. 1965. Stratigrafiya Neogenovykh molass Predkarpatskovo progriba. Akademiya Nauk Ukrainskoy SSR Institut Geologii I Geokhimii Goryuchikh, 165. (In Russian)
Vyalov, O. 1966. Sledy zhiznedeyatelnosti organizmov i ikh paleontologicheskoe znacheniye. Academiya Nauk Ukrainskoy SSR Institut Geologii I Geokhimii Goryuchikh Iskopayemykh, 219. (In Russian)
Wheeler, E.A. and Manchester, S.R. 2002. Woods of the middle Eocene nut beds flora, Clarno Formation, Oregon, USA. International Association of Wood Anatomists at the Nationaal Herbarium Nederland, 3.
https://doi.org/10.1600/036364404774195665
Xing, L., Belvedere, M., Buckley, L., Falk, A., Lockley, M., Klein, H., Abassi, N., Zhang, X., and Tang, Y. 2014. First record of bird tracks from Paleogene of China (Guangdong Province). Palaeogeography, Palaeoclimatology, Palaeoecology, 414:415–425.
https://doi.org/10.1016/j.palaeo.2014.08.031
Zonneveld, J.P., Zaim, Y., Aswan, A., Ciochon, R.L., Smith, T., Head, J., Wilf, P., and Bloch, J.I. 2024. Avian foraging on an intertidal mudflat succession in the Eocene Tanjung Formation, Asem Asem Basin, South Kalimantan, Indonesian Borneo. Palaios, 39:67–96.
https://doi.org/10.2110/palo.2023.004
