

 A new species of the antiarch Microbrachius from the Middle Devonian (Givetian) of Estonia
A new species of the antiarch Microbrachius from the Middle Devonian (Givetian) of Estonia
Article number: 28.2.a35
https://doi.org/10.26879/1525
Copyright Palaeontological Association, August 2025
Author biographies
Plain-language and multi-lingual abstracts
PDF version
Submission: 16 December 2024. Acceptance: 24 July 2025.
ABSTRACT
A new species of antiarch fish (Placodermi) Microbrachius longi sp. nov. is described from the Givetian (Middle Devonian) of Essi Farm in Estonia. The species consists of disarticulated plates that are suitably well preserved to distinguish the species from other species of Microbrachius. Characteristics include the large orbits and short cranium anterior of these orbits. The articulation structures for the pectorals is well preserved on the anterior ventrolateral plate allowing for its detailed description for the first time in this genus. This species also has reproductive claspers that show it reproduced by internal copulation. The species is compared with the type species Microbrachius dicki where a possible tail is described for the first time in this genus. The presence of Microbrachius in Estonia, Belarus, and Scotland adds to evidence of the interconnection between numerous locations in the Middle Devonian of Laurussia as demonstrated by numerous co-specific genera and species between these locations.
Michael J. Newman. Vine Lodge, Vine Road, Johnston, Haverfordwest, Pembrokeshire SA62 3NZ, UK. ichthyman@btinternet.com
Jan L. den Blaauwen. Willem Pijperstraat 22, 7425HS, Deventer, Netherlands. blaauwen-213142@kpnmail.nl
Keywords: new species; pectoral articulation; placoderm; internal copulation; Laurussia
https://zoobank.org/1F341F09-29CD-45A0-BD9E-63B2BCF5CE6F
Final citation: Mark-Kurik, Elga, Newman, Michael J., den Blaauwen, Jan L., and Jones, Roger. 2025. A new species of the antiarch Microbrachius from the Middle Devonian (Givetian) of Estonia. Palaeontologia Electronica, 28(2):a35.
https://doi.org/10.26879/1525
palaeo-electronica.org/content/2025/5613-microbrachius-from-the-middle-devonian-of-estonia
Copyright: August 2025 Palaeontological Association.
This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
creativecommons.org/licenses/by/4.0
INTRODUCTION
The late Estonian palaeontologist Elga Mark-Kurik (1928-2016) wrote a draft of this paper in the late 1980s, describing two new species of Microbrachius from Estonia and Belarus. Both these species are only known from disarticulated plates. Two species of the genus were then known, the Scottish type species Microbrachius dicki Traquair, 1888, where well-preserved articulated specimens are known, and the Chinese species Microbrachius sinensis Pan, 1984 where only a few isolated plates are known. In 1999 Microbrachius chuandongensis Wang and Zhang, 1999 from China was erected. A few articulated specimens are known of this species, but all the specimens are very poorly preserved with the restorations given by Wang and Zhang (1999, figures 2-4) overly interpreted. In 2015 the discovery of internal reproductive structures in both the Scottish and Estonian species (Long et al., 2015) revealed that antiarch placoderms reproduced by copulation. This implied that internal fertilization was the general condition in jawed fishes rather than external fertilization. In 2018 Microbrachius kedoae Mark-Kurik, Newman, Toom, and den Blaauwen, 2018, from Belarus was described. In this article we complete Mark-Kurik’s original manuscript and describe the Estonian species, along with some information on the tail of M. dicki.
MATERIAL AND METHODS
Only mechanical preparation of specimens was made. The illustrations of the individual plates were made by one of the authors (EMK) using pencil in the 1980s. Photographs of the Estonian material in Tallinn was taken with a Canon EOS 5DSR camera with finer details made using the apochromatic zoom system Leica Z16 APO. Photographs of the Scottish material was made using a Canon Digital Rebel 450D camera. Images and drawings were made using Adobe Photoshop.
Institutional Abbreviations
GIT - Institute of Geology at Tallinn University of Technology, Estonia. GSE - British Geological Survey collection, Edinburgh. NMS - National Museums Scotland.
HISTORY OF RESEARCH ON THE ESTONIAN MICROBRACHIUS
The general history of the genus Microbrachius was recently published by Mark-Kurik et al. (2018). In this paper we concentrate on the history of the Estonian species. On one day in July 1970 samples of siltstones and mudstones containing fossil remains were collected from an outcrop on the left bank of the Võhandu River near Essi farm in South-East Estonia. These were collected by Lia Nõmmisto, Andres Nõlvak, and Tiiu Märss. The fossil remains were so small they were not identified at the time. One of the authors (EMK) using a binocular microscope noted minute, disarticulated dermal plates of a genus she recognised as similar to Microbrachius dicki, a well-known species from the Middle Devonian of Scotland.
SYSTEMATIC PALAEONTOLOGY
Suborder BOTHRIOLEPIDOIDEI Miles, 1968
Family MICROBRACHIDAE Gross, 1965
Genus MICROBRACHIUS Traquair, 1888
Type species. Microbrachius dicki Traquair, 1888.
Diagnosis. As in Mark-Kurik et al., 2018, p. 4.
Species composition. Microbrachius dicki Traquair, 1888, Microbrachius sinensis Pan, 1984, Microbrachius chuandongensis Wang and Zhang, 1999, and Microbrachius kedoae Mark-Kurik, Newman, Toom, and den Blaauwen, 2018.
Microbrachius longi sp. nov.
Figure 1, Figure 2, Figure 3, Figure 4A, Figure 5A-F, Figure 6, Figure 7, Figure 8, Figure 9, Figure 10, Figure 11, Figure 12
zoobank.org/B7462E7B-022D-4205-9B3E-D45D5C41A715
2000 Microbrachius cf. dicki Traq.; Mark-Kurik, p. 317-319, fig. 4, tab. 2B, 3.
2003 Microbrachius sp.; Mark-Kurik and Nemliher, p. 38-40, tab. 1.
2009 Microbrachius sp.; Lebedev et al., p. 346, 348, 353, fig. 2P.
2014 Microbrachius; Mark-Kurik and Põldvere, p. 43
2015 Microbrachius sp.; Long et al., p. 196-197, figs. 1a-c, ED fig. 1.
2018 Microbrachius sp.; Mark-Kurik et al., p. 3-4, 12.
2019 Microbrachius dicki; Trinajstic et al., p. 211.
Holotype. GIT 628-7, a fused nuchal and post pineal plate in visceral aspect.
Etymology. Named after Professor John Long (Flinders University) who was the lead author of a paper (Long et al., 2015) that first described remains of this species.
Material. The material consists of around 25 dermal plates and around 30 fragments, which are all disarticulated except for the fused nuchal and post pineal plates. All the specimens are housed in the Institute of Geology at Tallinn University of Technology, Estonia with the registration prefix GIT 628. Examples of all the plates of Microbrachius longi were available to study except the premedian plate (Prm) and the postmarginal plate (Pmg) of the head. Also, no ventral plates of the head are known. Examples of all the trunk plates were available for study. The post trunk armour elements of the fish are unknown.
Locality and stratigraphic position. Outcrop near Essi farm (Essi müür) on the left bank of the Võhandu River, about 3 km downstream of Leevi village, South-East Estonia (Long et al. 2015, extended data figure 1). The outcrop is in the Abava Member of the Burtnieki Formation and is considered Middle Devonian (Givetian).
Diagnosis. Suture between the lateral plate and premedian plate short at 38% the length of the lateral plate; width of the lateral plate at the anterior end 18% the width of the headshield; postorbital crista of the headshield has a very shallow curve; large orbital fenestra at 47% the width and 39% the height of the headshield; the nuchal and postpineal plates wide at 44% the width of the headshield; wide longitudinal ridge in the obtected nuchal area of the nuchal plate; breadth/length index of the anterior median dorsal plate 102 to 110; prominent lateral corners on the posterior median dorsal plate; posterior ventral pit on the posterior median dorsal plate very shallow; ventral lamina 10% wider than the lateral lamina in the anterior ventro-lateral plate; anterolateral angle of subcephalic division of ventral lamina of anterior ventro-lateral plate irregular in shape and often crenulated; pars pedalis of the anterior ventro-lateral plate placed obliquely at an angle of 40 degrees to the longitudinal axis of the trunk armour; canal opening within the ventral half of the fossa articularis pectoralis in the anterior ventro-lateral plate.
Remarks. Microbrachius longi differs from Microbrachius dicki in having a much larger orbital fenestra, much shorter premedian plate, narrower lateral plates, and wider postpineal and nuchal plates. Postcranially, the lateral margins and posterior angle of the posterior median dorsal plate are more prominent. Additionally, the processus obstans of the anterior dorsolateral plate is more prominent, and the foramen axillare of the anterior ventrolateral plate is larger than M. dicki. M. longi differs from Microbrachius kedoae in having a narrower mixilateral plate. Of more importance is that the mixilateral plate overlaps the anterior median dorsal plate in M. longi, but the anterior median dorsal plate overlaps the mixilateral plate in M. kedoae. Another minor point of difference is that M. longi has conspicuous recurved denticles on the mesial side of the distil segment of the pectoral appendage, whereas M. kedoae has small, blunt denticles. Whilst Microbrachius chuandongensis is poorly known, the orbital fenestra is larger in M. longi and the postpineal and nuchal plates wider. Despite the poor preservation it seems that M. chuandongensis does not have tubercles on top of the ridges on the external surface of the anterior median dorsal plate like M. longi does. Microbrachius sinensis shares this character with M. chuandongensis, Further differences between M. longi and M. sinensis are that the lateral corners and posterior angle of the posterior median dorsal plate are much blunter in M. sinensis. The plate is also narrower in M. sinensis. Another difference is that the anterolateral angle of subcephalic division of ventral lamina of the anterior ventrolateral plate is irregular in shape in M. longi, smooth in M sinensis. Also, the foramen axillare is smaller in M. sinensis.
Description
Head plates. General features. No complete heads of Microbrachius longi are known with only are few isolated plates available for description (see below). No head plates are known in Microbrachius sinensis (Pan, 1984). Microbrachius chuandongensis (Wang and Zhang, 1999) is not well preserved, and only general characters can be ascertained. No articulated heads are known of Microbrachius kedoae (Mark-Kurik et al., 2018, figures 3-4) with only the lateral and paranuchal plates available for comparison. Microbrachius dicki (Hemmings 1978, figures 25-26) is the most complete species known, and comparisons with M. longi are mostly made with this species.
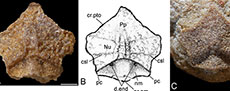
 Lateral plate (L). Four lateral plates were available to study, but only three were suitably preserved to warrant description. GIT 628-81 (Figure 1D) is a right, externally (dorsally) preserved plate, but is very incomplete. GIT 628-9 (Figure 1A-B) and GIT 628-79 (Figure 1C) are the best viscerally preserved plates. Both are left lateral plates. GIT 628-9 was figured by Long et al. (2015, extended figure 1B) but was figured upside down.
Lateral plate (L). Four lateral plates were available to study, but only three were suitably preserved to warrant description. GIT 628-81 (Figure 1D) is a right, externally (dorsally) preserved plate, but is very incomplete. GIT 628-9 (Figure 1A-B) and GIT 628-79 (Figure 1C) are the best viscerally preserved plates. Both are left lateral plates. GIT 628-9 was figured by Long et al. (2015, extended figure 1B) but was figured upside down.
The lateral plate is quite long and thin being about twice as long as wide. It is particularly thin at its anterior margin at 18% the width of the headshield. This compares to 25% in Microbrachius dicki (Hemmings, 1978). The suture between the lateral plate and premedian plate is short at 38% the length of the lateral plate. This is similar to Microbrachius kedoae at 37%, but very different Microbrachius dicki at 80% (Hemmings, 1978). The mesial edge curves around the orbital fenestra with the lateral edge reflecting this curve.
GIT 628-81 has rounded tubercles on its surface and part of the principle section of the infraorbital sensory line (ifc1) preserved. As the anterior and posterior ends of GIT 628-81 are incomplete little more can be said of the plate’s orientation to other plates of the head.
As the viscerally preserved lateral plates are more complete, more characters can be discerned. GIT 628-9 has a rod-like crenulation (rlc) at the anterior end like Microbrachius kedoae (Mark-Kurik et al., 2018, figures 3C-D). Microbrachius dicki conversely has a smooth anterior end (Hemmings, 1978, figure 26B). The mesial margin runs from the preorbital angle (cpr) to the postorbital angle (cpt) and is hollowed dorsally by the lateropremesial depression. This depression continues laterally into the lateropremesial ridge of the headshield (lpri). Lateral to this is a weakly developed premedian ridge (r). The preorbital angle has a distinct process. The antero-lateral angle of the otico-occipital depression of the headshield (pr.po) is a quite wide hollow entering the posterior edge. M. dicki also has a quite wide pr.po, but with a more pointed anterior termination. M. kedoae has a narrower pr.po compared to these two species.
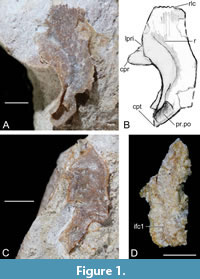
 Nuchal plate + postpineal plate (Nu+Pp). The nuchal plate is fused with the pineal plate and is quite a thick element like Microbrachius dicki (Hemmings 1978, figures 25, 26C). Six specimens were available for study. GIT 625-5 (Figure 2A-B) and GIT 625-6 (Figure 2C-D) are the best preserved externally (dorsally) and GIT 628-7 (Figure 2E-F) and GIT 628-8 (Figure 2G-H) the best preserved viscerally. The nuchal and postpineal are wide at 44% the width of the headshield. This compares to 33% in M. dicki (Hemmings, 1978).
Nuchal plate + postpineal plate (Nu+Pp). The nuchal plate is fused with the pineal plate and is quite a thick element like Microbrachius dicki (Hemmings 1978, figures 25, 26C). Six specimens were available for study. GIT 625-5 (Figure 2A-B) and GIT 625-6 (Figure 2C-D) are the best preserved externally (dorsally) and GIT 628-7 (Figure 2E-F) and GIT 628-8 (Figure 2G-H) the best preserved viscerally. The nuchal and postpineal are wide at 44% the width of the headshield. This compares to 33% in M. dicki (Hemmings, 1978).
Postpineal plate. The postpineal plate is rhomboidal in shape externally (dorsally), with the corners pointing longitudinally and laterally. Externally (dorsally) the breadth/length index ranges between 150 to 200 and is slightly convex horizontally. The anterior end has a pronounced point into the orbital fenestra. The plate has a shallower curve of the postorbital crista of the headshield (cr.pto) than Microbrachius dicki (Hemmings, 1978, figures 25, 26C). This is unsurprising as the orbital fenestra is larger in Microbrachius longi. The lateral corners of the pineal plate (pp.mc) sometimes extend laterally beyond the lateral corners of the nuchal plate (nu.mc). This is seen more in visceral aspect.
Viscerally, the posterior suture to the nuchal plate is concave rather than convex like in Microbrachius dicki. This means the postpineal plate is quite short compared to M. dicki and therefore, the median ridge (mr) is also much shorter.
Nuchal plate. The nuchal plate is widest at the lateral corners. The breadth/length index is quite variable ranging from 156 to 169 dorsally and 134 to 149 viscerally. The postero-lateral corners (pc) are not as prominent as the lateral corners. The posterior median process of headshield is blunt (pr.nm). The obtected nuchal area (nm) lacks ornamentation and has a very wide longitudinal ridge at the centre. Microbrachius dicki does not appear to have a similar ridge (Hemmings 1978). Either side of this ridge towards the anterior end of the obtected nuchal area is the opening of canal for endolymphatic duct (d.end). The openings are about twice as widely spaced as in M. dicki due to the greater width of the nuchal plate. This ridge continues anteriorly into the ornamented area of the nuchal and postpineal plates where it bifurcates (Figure 2A-B). A wide longitudinal hollow separates the two lateral ridges. These ridges conjoin anteriorly in the centre of the pineal plate. On the tops of these ridges is a row of tubercles. A pair of lateral ridges are also present in M. dicki (Hemmings, 1978), but they are contained within the postpineal plate and have a different structure. A faint horizontal central sensory line groove (csl) crosses the middle of the ornamented part of the nuchal plate but is difficult to discern centrally (Figure 2A-B).
On the visceral side of the nuchal plate, the supraoptic groove of headshield (sog) is quite broad (Figure 2E-H) and of similar dimensions to Microbrachius dicki (Hemmings, 1978). The internal opening of canal for endolymphatic duct (d.end1) are widely spaced like M. dicki. Laterally, there are deep embayments with a sub horizontal ridge near the centre. This ridge separates the areas of overlap for the paranuchal plate (cf.Pn) and the lateral plate (cf.L).
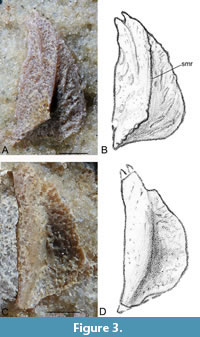 Submarginal plate (SM). Two submarginal plates were available for study. GIT 628-32 (Figure 3A-B) is a right submarginal plate in dorso-lateral (external) aspect, and GIT 628-33 (Figure 3C-D) is a left submarginal plate in visceral aspect. Submarginal plates are unknown in the other species of Microbrachius, although Hemmings (1978) considered Microbrachius dicki had a quite narrow plate at the lateral edges of the headshield.
Submarginal plate (SM). Two submarginal plates were available for study. GIT 628-32 (Figure 3A-B) is a right submarginal plate in dorso-lateral (external) aspect, and GIT 628-33 (Figure 3C-D) is a left submarginal plate in visceral aspect. Submarginal plates are unknown in the other species of Microbrachius, although Hemmings (1978) considered Microbrachius dicki had a quite narrow plate at the lateral edges of the headshield.
Dorso-laterally, the mesial edge is quite straight and would have been sutured to the lateral edges of the skull roof. There is no evidence of the infraspiracular notch (cf. Stensiö, 1948, figure 32). The anterior end is pointed and bifurcated. The ventral margin is strongly convex posteriorly. In the cross section the plate is roof shaped with a longitudinal median ridge (smr) that bends towards the posterior mesial corner near the posterior margin of the plate. The ornamentation consists of short, narrow ridges.
Viscerally, the plate has a smooth surface with a longitudinal depression beneath the median ridge on the external surface. Morphologically the submarginal plate resembles the submarginal plates of asterolepidoids, such as Sherbonaspis (Young and Gorter, 1981, figure 14) or Pterichthyodes (Hemmings, 1978, figures 5E-I, plate 3, figure 1, referred to as an extralateral plate).
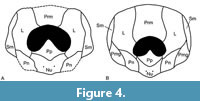 Whilst only three plates of the headshield are known, comparison with the better known Microbrachius dicki allows a tentative reconstruction of the headshield of Microbrachius longi to be made (Figure 4A). Compared to Hemmings’ (1978, figure 36) reconstruction of M. dicki (reproduced in Figure 4B), the orbital fenestra of the headshield is large at 47% of the width and 39% of the height. The corresponding figures for M. dicki are 39% and 27%, respectfully. This makes the orbital fenestra about three times as large as M. dicki. Also, the premedian plate is about 60% longer than M. dicki. Finally, the anterior part of the lateral plate is narrower than M. dicki with the nuchal and postpineal plates also wider.
Whilst only three plates of the headshield are known, comparison with the better known Microbrachius dicki allows a tentative reconstruction of the headshield of Microbrachius longi to be made (Figure 4A). Compared to Hemmings’ (1978, figure 36) reconstruction of M. dicki (reproduced in Figure 4B), the orbital fenestra of the headshield is large at 47% of the width and 39% of the height. The corresponding figures for M. dicki are 39% and 27%, respectfully. This makes the orbital fenestra about three times as large as M. dicki. Also, the premedian plate is about 60% longer than M. dicki. Finally, the anterior part of the lateral plate is narrower than M. dicki with the nuchal and postpineal plates also wider.
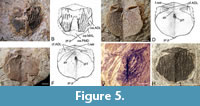 Thorax. Anterior median dorsal plate (AMD). Seven anterior median dorsal plates were available to study with GIT 628-1 (Figure 5A-B) the best preserved externally (dorsally). GIT 628-2 (Figure 5C-D) and GIT 628-4 (Figure 5E-F) are the best preserved viscerally preserved plates.
Thorax. Anterior median dorsal plate (AMD). Seven anterior median dorsal plates were available to study with GIT 628-1 (Figure 5A-B) the best preserved externally (dorsally). GIT 628-2 (Figure 5C-D) and GIT 628-4 (Figure 5E-F) are the best preserved viscerally preserved plates.
The anterior median dorsal plate is a short, broad, and ovoid plate with a breadth/length index of between 102 to 110. This compares with a breadth/length index of 100 to 120 for Microbrachius dicki (Hemmings, 1978, figure 30), 120 to 128 for Microbrachius kedoae (Mark-Kurik et al., 2018, figure 5), and around 100 for both Microbrachius sinensis (Pan, 1984, figure 1A, plate 1, figures 1-3) and Microbrachius chuandongensis (Wang and Zhang, 1999, figure 6). The posterior margin is convex in profile with a pronounced posterior median process (pr.p). The anterior margin is also convex in profile with the lateral edges fairly straight with no obvious lateral corners. M. dicki (Hemmings, 1978), M. kedoae (Mark-Kurik et al., 2018), and M. sinensis (Pan, 1984) have similar profiles. The situation in M. chuandongensis (Wang and Zhang, 1999) is difficult to gauge as the material is so poorly preserved and little more can be said of the plate (but see below).
Externally (dorsally) the anterior median dorsal plate of Microbrachius longi is overlapped by the mixilateral plates (MxL), the posterior median dorsal plate (PMD), and the posterior part of the anterior dorsolateral plates (ADL). The areas of overlap for the mixilateral plates (oa.MxL) are short, thin embayments, with the areas of overlap for the anterior dorsolateral plates (oa.ADL) being a little deeper. The area of overlap for the posterior median dorsal plate (oa.PMD) is a large embayment that consists of most of the posterior edge of the anterior median dorsal plate. M. dicki (Hemmings, 1978) and M. sinensis (Pan, 1984) have similar areas of overlap.
The ornamentation consists of quite thick, sinuous, longitudinal ridges with rounded tubercles on top. Microbrachius dicki has a similar ornamentation (Hemmings, 1978). However, there is some variability in this species as some of the ornamental ridges can have a similar thickness to Microbrachius longi such as GSE 13338 (Figure 5G), but other specimens such as GSE 13333 (labelled B139i in Hemmings, 1978, figure 30E) (Figure 5H) can have much narrower ridges. This may indicate that the ornamentation in Microbrachius is quite variable, but a greater number of specimens would be required to see if this was the same for M. longi. Microbrachius sinensis (Pan, 1984) and Microbrachius chuandongensis (Wang and Zhang, 1999) have a similar ornamentation, but no tubercles on top of the ridges. Externally preserved anterior median dorsal plates are not known in Microbrachius kedoae. M. longi also has a wide smooth area surrounded by a pair of ridges in the middle of the anterior median dorsal plate. These ridges emanate from the posterior median dorsal plate and join about two thirds along the anterior median dorsal plate in a broad arch. These paired ridges do not have tubercles on top of them. M dicki and M. sinensis have a similar pair of ridges, although in the latter they join more anteriorly.
Viscerally, the anterolateral corners of the anterior median dorsal plate overlap the anterior dorsolateral plates (cf.ADL). The area consists of deep embayments that form an extended indentation following the course of the postlevator thickening (alr). Microbrachius dicki (Hemmings, 1978) and Microbrachius kedoae (Mark-Kurik et al., 2018) have a similar structure. One note of difference viscerally in M. kedoae is that the anterior median dorsal plate overlaps the mixilateral plate rather than the other way around. The postlevator thickening of Microbrachius longi continues to the lateral edges in the form of thin ridges. This is also seen in M. kedoae, but not the other species where this area is known. Anterior of the postlevator thickening is the levator fossa (f.retr). A fairly deep, anterior ventral pit (pt1) is located on a north/south axis in the anterior third of the plate. Microbrachius sinensis (Pan, 1984) and M. kedoae have a similar pit, but Microbrachius dicki does not (Hemmings, 1978).
 Posterior median dorsal plate (PMD). 11 posterior median dorsal plates were available for study. The holotype GIT 628-3 (Figure 6A-B) and GIT 628-11 (Figure 6C-D) are the best preserved in dorsal (external) aspect and GIT 628-34 (Figure 6E-F) and GIT 628-10 (Figure 6G-H) the best preserved in visceral aspect. The overall shape of the plate is similar to Microbrachius dicki (Hemmings, 1978, figure 29), Microbrachius kedoae (Mark-Kurik et al., 2018, figure 6) and Microbrachius sinensis (Pan, 1984, figure 1B, plate 1, figure 4). The plate is poorly preserved in Microbrachius chuandongensis and therefore, not comparable.
Posterior median dorsal plate (PMD). 11 posterior median dorsal plates were available for study. The holotype GIT 628-3 (Figure 6A-B) and GIT 628-11 (Figure 6C-D) are the best preserved in dorsal (external) aspect and GIT 628-34 (Figure 6E-F) and GIT 628-10 (Figure 6G-H) the best preserved in visceral aspect. The overall shape of the plate is similar to Microbrachius dicki (Hemmings, 1978, figure 29), Microbrachius kedoae (Mark-Kurik et al., 2018, figure 6) and Microbrachius sinensis (Pan, 1984, figure 1B, plate 1, figure 4). The plate is poorly preserved in Microbrachius chuandongensis and therefore, not comparable.
The plate is short and broad, being broadest at the lateral corners (l). These are much more pronounced and sharper than in other Microbrachius species, other than Microbrachius kedoae. The plate has a breadth/length index of 97-103, compared to about 100 in Microbrachius dicki and 76 in Microbrachius sinensis. Mark-Kurik et al. (2018) were unable to give a breadth/length index for the plate for Microbrachius kedoae. At the posterior end of the plate is a pronounced posterior angle (pa). This angle is similar to M. kedoae and M. sinensis, but M. dicki has a blunter angle. The lateral edges of the plate are gently curved between the posterior angle and the lateral corners. At the anterior margin is a pronounced anterior angle (aa). Lateral to this angle are the anterolateral angles (aal) which are connected to the lateral corners by a straight lateral margin.
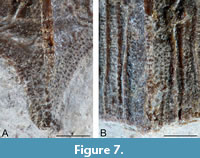 The posterior median dorsal plate is heavily ornamented on its exterior (dorsal) surface. A prominent dorsal median ridge (dmr) connects the anterior and posterior angles. This ridge is topped with a row of rounded tubercles that become more elongated posteriorly (Figure 6A-D, Figure 7A). The ridge reaches an apex at the posterior dorsal angle (pda) where it meets an indistinct lateral ridge that stretches from the lateral corners. Either side of this ridge are the dorsal sensory line grooves (dlg). When these grooves reach the lateral ridge, they curve postero-laterally. Lateral to these grooves are four to six longitudinal ridges (Figure 6A-D, Figure 7B) that reach as far as the lateral ridge or stop shortly after. They are not necessarily symmetrical in number either side of the plate. These ridges consist of elongated longitudinal tubercles. Smaller tubercles are present between these ridges. Posterior to the lateral ridge, are rows of small rounded tubercles that follow the lateral edge of the plate.
The posterior median dorsal plate is heavily ornamented on its exterior (dorsal) surface. A prominent dorsal median ridge (dmr) connects the anterior and posterior angles. This ridge is topped with a row of rounded tubercles that become more elongated posteriorly (Figure 6A-D, Figure 7A). The ridge reaches an apex at the posterior dorsal angle (pda) where it meets an indistinct lateral ridge that stretches from the lateral corners. Either side of this ridge are the dorsal sensory line grooves (dlg). When these grooves reach the lateral ridge, they curve postero-laterally. Lateral to these grooves are four to six longitudinal ridges (Figure 6A-D, Figure 7B) that reach as far as the lateral ridge or stop shortly after. They are not necessarily symmetrical in number either side of the plate. These ridges consist of elongated longitudinal tubercles. Smaller tubercles are present between these ridges. Posterior to the lateral ridge, are rows of small rounded tubercles that follow the lateral edge of the plate.
Overall, the ornamentation is similar to Microbrachius dicki, but the longitudinal ridges are narrower and more numerous in M. dicki (Hemmings, 1978; Long, 2015). Microbrachius kedoae has far fewer longitudinal ridges, but the only known specimen that shows the dorsal ornamentation is very small and it could be a juvenile character (Mark-Kurik et al., 2018). Likewise, the lack of tubercles in M. kedoae might also be a juvenile character. Microbrachius sinensis has an ornamentation consisting of numerous, thin smooth ridges (Pan 1984). The preservation is not good enough to tell if they continued posterior of the median ridge, or any other details of the external (dorsal) ornamentation.
Viscerally, the posterior median dorsal plate of Microbrachius longi overlaps the anterior median dorsal plate (cf.AMD) at the anterior end. This area of overlap has a midline longitudinal groove. This area of overlap continues laterally to form areas that overlap the two mixilateral plates (cf.MxL). These two areas are moderately narrow longitudinally but form a deep embayments just anterior of a prominent crista transversalis interna posterior (crtp). Viscerally there is very little to distinguish between the known species of Microbrachius, including Microbrachius kedoae where only the anterior end is known. However, in M. longi a very shallow posterior ventral pit (pt2) is located midway between these embayments contrasting with the deeper pits seen in Microbrachius dicki (Hemmings, 1978) and Microbrachius sinensis (Pan, 1984). The posterior marginal area (pma) is quite long, but this might be quite a variable character like M. dicki (Hemmings, 1978).
 Anterior dorsolateral plate (ADL). Five anterior dorsolateral plates were available for study, with GIT 628-11 (Figure 8A-B), a right plate and GIT 628-14 (Figure 8C-D), a left plate the best externally preserved specimens. GIT 628-13 (Figure 8E-F), a left plate, is the only specimen preserved viscerally. GIT 628-11 is particularly interesting as it shows two puncture marks on the dorsal lamina (dlm), which have been interpreted as bite marks from a predator (Lebedev et al., 2009). The illustration of the plate (Figure 8B) does not show these bite marks to allow better description of the plate. The plate is a sub-rectangular shaped plate of variable length. This variable length makes a breadth/length index an unreliable character. The plate is widest at the anterior border. This border is wide to accommodate the large head. Comparisons can only be made with Microbrachius dicki (Hemmings, 1978, figures 31A-C) as there are no specimens of this plate known in Microbrachius kedoae (Mark-Kurik et al., 2018) and Microbrachius sinensis (Pan, 1984). Microbrachius chuandongensis (Wang and Zhang, 1999) is too poorly preserved to warrant comparison.
Anterior dorsolateral plate (ADL). Five anterior dorsolateral plates were available for study, with GIT 628-11 (Figure 8A-B), a right plate and GIT 628-14 (Figure 8C-D), a left plate the best externally preserved specimens. GIT 628-13 (Figure 8E-F), a left plate, is the only specimen preserved viscerally. GIT 628-11 is particularly interesting as it shows two puncture marks on the dorsal lamina (dlm), which have been interpreted as bite marks from a predator (Lebedev et al., 2009). The illustration of the plate (Figure 8B) does not show these bite marks to allow better description of the plate. The plate is a sub-rectangular shaped plate of variable length. This variable length makes a breadth/length index an unreliable character. The plate is widest at the anterior border. This border is wide to accommodate the large head. Comparisons can only be made with Microbrachius dicki (Hemmings, 1978, figures 31A-C) as there are no specimens of this plate known in Microbrachius kedoae (Mark-Kurik et al., 2018) and Microbrachius sinensis (Pan, 1984). Microbrachius chuandongensis (Wang and Zhang, 1999) is too poorly preserved to warrant comparison.
Most of the characters of the anterior dorsal-lateral plate are in the external surface. There is a very well-developed processus obstans (pro) unlike in Microbrachius dicki where it is hardly prominent at all. The main lateral line groove (lcg) is conspicuous and runs subparallel to the lateral margin of the plate. Near the processus obstans the groove turns a right angle to the articular fossa (f.art). The anterior part of the main lateral line grove is not prominent in M. dicki, and it seems to peter out before it gets near to the anterior edge of the plate. The dorsolateral ridge of the trunk armour (dlr) is prominent unlike in M. dicki and ends posteriorly at a prominent posterior angle. This ridge separates the dorsal lamina and the lateral lamina (llm) with the former being about three times to about half the breadth as the latter. This is similar to M. dicki. A very small area is overlapped by the anterior ventrolateral plate (oa.AVL) lateral to the processus obstans. Most of the contact between the anterior dorsolateral and anterior ventrolateral plates in Microbrachius longi is by a simple suture whereas in M. dicki the whole contact is a simple suture. The anterior median dorsal plate overlaps the antero-lateral mesial corner of the anterior dorsolateral plate creating a sinuous overlap area (oa.AMD). This area has a central sub-horizontal ridge. The anterior dorsolateral plate has an ornamentation of sinuous longitudinal ridges with quite large, rounded tubercles on top, a bit like M. dicki, although the ridges tend to be less longitudinal as they follow the curvature of the edges of the plate.
The visceral side of the anterior dorsolateral plate is smooth with a wide longitudinal valley under the dorsolateral ridge of the trunk armour mentioned above. Posteriorly the plate overlaps the mixilateral plate creating a triangular area of overlap (cf.MxL). Emanating from this area is a median, narrow embayment going a short way anteriorly. The anterior dorsolateral plate also overlaps the anterior median dorsal plate on its mesial edge creating a small semi-circular area of overlap (cf.AMD). The anterior edge of the plate has a prominent, smooth lateral division of crista transversalis interna anterior (cit).
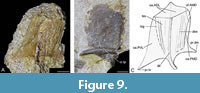 Mixilateral plate (MxL). Eight mixilateral plates were available for study, but most were fragmentary with no known right plates. GIT 628-16 (Figure 9A) a plate in external aspect and GIT 628-64 (Figure 9B) a plate in visceral aspect are the best-preserved plates GIT 628-16 is sufficiently complete to allow a reconstruction of the external armour to be made (Figure 9C).
Mixilateral plate (MxL). Eight mixilateral plates were available for study, but most were fragmentary with no known right plates. GIT 628-16 (Figure 9A) a plate in external aspect and GIT 628-64 (Figure 9B) a plate in visceral aspect are the best-preserved plates GIT 628-16 is sufficiently complete to allow a reconstruction of the external armour to be made (Figure 9C).
The breadth/length index is estimated to be 90. This compares to 123 to 132 in Microbrachius dicki (Hemmings, 1978, figures 31D-G), 101 in Microbrachius kedoae (Mark-Kurik et al., 2018, figure 7). The plate is unknown in Microbrachius sinensis (Pan, 1984) and the breadth/length index of 83 in Microbrachius chuandongensis (Wang and Zhang, 1999, figure 4) is only a rough estimate as the plate is poorly preserved. The plate is mostly slender, but with a broad posterior margin with prominent median dorsal (pr.dm) and posteroventral (pr.tv) processes.
Externally, the dorsal (dlm) and lateral laminas (llm) are roughly equal in size like M. dicki. This contrasts with Microbrachius kedoae where the dorsal lamina is significantly larger. The mixilateral plate is overlapped by the anterior dorsolateral plate creating a large triangular overlap area (oa.ADL) covering the whole anterior margin of the plate. The lateral margin of mixilateral plate is overlapped by the posterior ventrolateral plate, but the preservation is not good, so the area overlapped (oa.PVL) is indistinct. Most of the dorsal margin of the mixilateral plate is overlapped by the posterior median dorsal plate creating a long, thin embayment overlap area (oa.PMD), except for the anterior lateral corner of the plate which is not overlapped. A prominent median dorso-lateral ridge (dlr) is present that ends posteriorly with a prominent posterior process. Anteriorly this ridge enters the area overlapped by the anterior dorsolateral plate creating a slight ridge than ends with a pronounced anterior corner. On the lateral side of the median dorso-lateral ridge is the main lateral line groove (lcg). The ornamentation is not well preserved but appears to consist of smooth longitudinal ridges.
The visceral side of the mixilateral plate is very smooth. A horizontal, quite thick, crista transversalis interna posterior (cr.tp) is present near the posterior border. The anterior dorsal corner of the mixilateral plate overlaps the anterior median dorsal plate (cf.AMD) creating a smooth slight embayment. This feature is also seen in Microbrachius dicki. This contrasts with Microbrachius kedoae as the anterior median dorsal plate overlaps the mixilateral plate.
 Anterior ventrolateral plate (AVL). Of the 12 AVL plates available for study, GIT 628-18 (Figure 10A-B) and GIT 628-22 (Figure 10C-E) are the best preserved in an external aspect. GIT 628-20 (Figure 10F), GIT 628-23 (Figure 10G), and GIT 628-19 (Figure 10H) whilst incomplete were the best-preserved plates in visceral aspect allowing a reconstruction of the visceral side (Figure 10I).
Anterior ventrolateral plate (AVL). Of the 12 AVL plates available for study, GIT 628-18 (Figure 10A-B) and GIT 628-22 (Figure 10C-E) are the best preserved in an external aspect. GIT 628-20 (Figure 10F), GIT 628-23 (Figure 10G), and GIT 628-19 (Figure 10H) whilst incomplete were the best-preserved plates in visceral aspect allowing a reconstruction of the visceral side (Figure 10I).
The anterior ventrolateral plate is ovoid in shape with a breadth/length index of about 63. This compares to between 65 to 85 for Microbrachius dicki (Hemmings, 1978, figure 32), about 60 for Microbrachius sinensis (Pan, 1984, figure 1C, plate 2) and about 75 for Microbrachius chuandongensis (Wang and Chang, 1999). However, M. chuandongensis is poorly preserved and no more comparison can be made with this species. Only a small portion of a plate in visceral aspect of Microbrachius kedoae (Mark-Kurik, 2018, figure 8) is known. The ventral lamina (vlm) is 10% wider than the lateral lamina (llm), compared to around 60% for Microbrachius dicki and 40% for M. sinensis. The posterior end of the ventrolateral plate has a pronounced ventrolateral angle (vcl). Leading from this angle is a prominent ventrolateral ridge of trunk armour (vlr) that reaches the anterior edge of the plate. A posterodorsal angle (pdc) is located on the edge of the lateral lamina. The anterolateral angle of subcephalic division of ventral lamina (c.al) whilst prominent is irregular in shape and often crenulated contra to M. dicki and M sinensis which have smooth profiles. On the mesial edge of the left anterior ventrolateral plate is a triangular area overlapped by the right anterior ventrolateral plate (oa.AVL).
The area for articulation of the pectoral appendage is well preserved on some of the anterior ventrolateral plates. This area is very poorly preserved in Microbrachius dicki (Hemmings, 1978) and difficult to interpret in Microbrachius sinensis (Pan, 1984) with areas of breakage obscuring detail. The processus brachialis (pbr) in Microbrachius longi is long with a bulbous end. A very small funnel pit (fp) is located near the end of the processus brachialis. The foramen axillare (f.ax) is quite large like Microbrachius kedoae (Mark-Kurik et al., 2018), but unlike Microbrachius dicki (Hemmings, 1978) and Microbrachius sinensis (Pan, 1984) where they are about half the size. Anterior of this and the processus brachialis is a moderately long margo limitans (m.lim) forming a semi-circular shape. This is broken by a narrow pars pedalis (pe). The ventral half of the fossa articularis pectoralis (f.art.v) is longer than the dorsal half of the fossa articularis pectoralis (f.art.d). This is due to the pars pedalis being placed obliquely at an angle of 40 degrees to the longitudinal axis of the trunk armour.
Within the dorsal half of the fossa articularis pectoralis is a large opening for a rostrocaudal canal (c.rc). Whilst this canal is not preserved in other species of Microbrachius, it is known in other antiarchs such as Bothriolepis (e.g., Young and Guorui, 1992).
The external ornamentation consists of small rounded tubercles forming long lateral ridges. The ridges become much shorter and sinuous anteriorly. Microbrachius sinensis has a similar ornamentation, but the preservation is not good, and it is not certain if the ridges were composed of tubercles (Pan, 1984). Microbrachius dicki has an ornamentation of small tubercles, but the preservation is not good, and it is uncertain if they form ridges or not (Hemmings, 1978).
The anterior ventrolateral plate is poorly preserved viscerally, and only general characters can be given of the left plate. The posterior edge of the left plate overlaps the left posterior ventrolateral plate forming a deep embayment (cf.PVL). A prominent, horizontal, and narrow crista transversalis anterior (cit) is located about a quarter distance from the anterior edge of the plate. The dorsal edge of the left plate overlaps the right anterior dorsolateral plate (cf.ADL). The lateral edge is not well preserved, and it is not possible to determine the nature of the contact with the median ventral plate. No real differences in the anterior ventrolateral plates can be seen between Microbrachius longi, Microbrachius dicki (Hemmings, 1978), Microbrachius kedoae (Mark-Kurik et al., 2018), and Mircobrachius sinensis (Pan, 1984), although the plates in the first three species are poorly preserved.
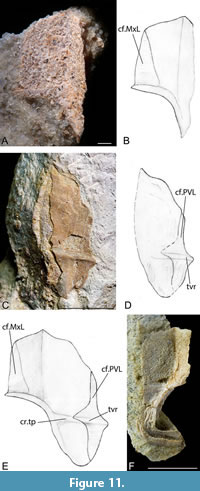 Posterior ventrolateral plate (PVL). Nine posterior ventrolateral plates were available, but only plates in visceral aspect were suitably preserved to warrant description. GIT 628-17 (Figure 11A-B) and GIT 628-25 (Figure 11C-D) whilst incomplete left plates do allow a reconstruction of the left plate in visceral aspect to be made (Figure 11E). GIT 628-24 (Figure 11F) is the best-preserved right plate but is very fragmentary. The breadth/length index of Microbrachius longi at its widest point is 63. This compares to between 50 to 73 for Microbrachius dicki (Hemmings, 1978, figure 34) and 53-67 for Microbrachius kedoae (Mark-Kurik et al., 2018, figure 9). The plate is unknown in Microbrachius sinensis (Pan, 1984) and very speculatively reconstructed in Microbrachius chuandongensis (Wang and Zhang, 1999). The significant variability in the breadth/length index of some species makes this not a useful character to compare between species.
Posterior ventrolateral plate (PVL). Nine posterior ventrolateral plates were available, but only plates in visceral aspect were suitably preserved to warrant description. GIT 628-17 (Figure 11A-B) and GIT 628-25 (Figure 11C-D) whilst incomplete left plates do allow a reconstruction of the left plate in visceral aspect to be made (Figure 11E). GIT 628-24 (Figure 11F) is the best-preserved right plate but is very fragmentary. The breadth/length index of Microbrachius longi at its widest point is 63. This compares to between 50 to 73 for Microbrachius dicki (Hemmings, 1978, figure 34) and 53-67 for Microbrachius kedoae (Mark-Kurik et al., 2018, figure 9). The plate is unknown in Microbrachius sinensis (Pan, 1984) and very speculatively reconstructed in Microbrachius chuandongensis (Wang and Zhang, 1999). The significant variability in the breadth/length index of some species makes this not a useful character to compare between species.
The anterior and posterior portions of the posterior ventrolateral are broadly equal in length. This is similar to Microbrachius kedoae (Mark-Kurik et al., 2018, figure 9) but contrasts with Microbrachius dicki where the anterior portion is significantly longer. The posterior ventrolateral plate overlaps the mixilateral plate creating a triangular area of overlap (cf.MxL). A well-developed horizontal crista transversalis interna posterior (cr.tp) is located at the lower third of the plate. The left posterior ventrolateral plate overlaps the right ventrolateral plate creating a deep, long embayment on the mesial edge (cf.PVL). On the left plate, the crista transversalis interna posterior continues into an area overlapping the right posterior ventrolateral plate as a transverse lateral ridge (tvr).
GIT 628-24 is especially important as it has a male reproductive clasper attached to the posterior part of the plate. Long et al. (2015, figures 1a-c) provided a thorough description of this specimen, where they describe the clasper as a deeply grooved, hook-shaped dermal bone. The deep groove was for the transfer of sperm. Long et al. (2015) describe a similar structure on both the left and right (so paired structures) posterior ventrolateral plates in Microbrachius dicki. We postulate that Microbrachius longi also had paired claspers. It is reasonable to assume that this character is common to all species of Microbrachius and maybe all antiarchs (Long et al., 2015). However, due to the lack of evidence of bony claspers in other antiarchs, they may have been composed of cartilage.
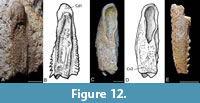 Pectoral appendages. Five partial pectoral appendages were available for study. Of most use were two right proximal appendages including GIT 628-27 (Figure 12A-B) that shows the lateral side well and GIT 628-28 (Figure 12C-D) that shows the mesial side well. GIT 628-29 (Figure 12E) is the only distil segment available for study.
Pectoral appendages. Five partial pectoral appendages were available for study. Of most use were two right proximal appendages including GIT 628-27 (Figure 12A-B) that shows the lateral side well and GIT 628-28 (Figure 12C-D) that shows the mesial side well. GIT 628-29 (Figure 12E) is the only distil segment available for study.
The proximal segments are box-like and long and slender as is typical for Microbrachius dicki and Microbrachius kedoae (e.g., Hemmings, 1978, figure 35; Mark-Kurik et al., 2018, figure 10). The socket has a rounded anterior end but is quite elongated. Both margins of the proximal segment are denticulated. The dorsal central plate 1 (Cd1) is determinable on GIT 628-27 and the triangular end of the ventral central plate 2 (Cv2) determinable on GIT 628-28. However, it is not possible to discern any of the other individual plates. Both margins of the proximal segment are denticulated, unlike M. dicki and M. kedoae which have smooth margins. The ornamentation of the lateral side consists of longitudinal ridges composed of rounded tubercles. M. kedoae has a similar ornamentation on both the lateral and mesial side of the proximal segment, unlike Microbrachius longi which has more randomly placed tubercles on the mesial side. M. dicki has an ornamentation of closely packed tubercles. Only inner moulds of proximal segments are known in Microbrachius sinensis (Pan, 1984, figures 1D-E, plate 1, figures 5-6) and Microbrachius chuandongensis (Wang and Chang, 1999, figure 5), none of which provide points of comparison with Microbrachius longi.
GIT 628-29 is incomplete and broken in places with no individual plates discernible. Rounded tubercles can be seen on the lateral side of the segment. A single row of evenly space recurved denticles is present on the mesial margin. This character is shared with Microbrachius dicki. Microbrachius kedoae differs by having small, blunt conical denticles. The ornamentation is not well preserved but appears to consist of randomly placed denticles somewhat reminiscent to M. dicki and M. kedoae.
DISCUSSIONS AND CONCLUSIONS
The vertebrate assemblage at the Essi Farm location is reasonably diverse. The fauna includes the heterostracans Psammolepis abavica Mark-Kurik, 1965 (in Obruchev and Mark-Kurik, 1965) (e.g., GIT 262-6) and Psammosteus bergi (Obruchev, 1943) (e.g., GIT 680-380). Another antiarch Asterolepis essica Lyarskaya, 1981 is also present (e.g., GIT 498-1) as well as an undescribed species of the arthrodire Watsonosteus (e.g., GIT 728-13). Finally, a species of the porolepiform Laccognathus sp. (e.g., GIT 365-284) is also present in the fauna. This assemblage is interesting as the genera Microbrachius and Watsonosteus are found in the Middle Devonian Orcadian Basin of Scotland. It had been recognised for some time that there was a faunal connection between the Baltic sequences and the Orcadian Basin (Ahlberg et al., 1999; Mark-Kurik, 2000; Newman and Trewin, 2008; Mark-Kurik and Põldvere, 2012). Recently, co-specific fauna between the two areas have been recognised (Newman et al., 2015; Newman et al., 2017; Burrow et al., 2016; den Blaauwen et al., 2019; Burrow et al., 2020a; Burrow et al., 2020b; Newman et al., 2021). The acanthodian stem chondrichthyan fishes have been shown to be particularly cosmopolitan (Plax and Newman, 2021). Microbrachius longi occurs in the Abava Member of the Burtnieki Regional Stage of the Baltic region along with the arthrodire placoderm Watsonosteus sp. mentioned above. These two genera also occur together in the John o’ Groats Sandstone Group in Caithness and its lateral equivalent, the Eday Group in Orkney. These two groups are correlated with the Burtnieki Regional Stage (Long et al., 2015; Burrow et al., 2020a; Plax and Newman, 2021) and thus are of the same age. Whilst the Microbrachius species are different, the Watsonosteus species may be the same, i.e., Watsonosteus fletti Miles and Westoll, 1963, the Scottish species. This is because another arthrodire species Coccosteus cuspidatus Miller, 1841 ex Agassiz MS is present in both areas at the same horizon. Clearly, the Estonian Watsonosteus species needs to be described to determine the situation.
 Since the Microbrachius dicki monograph by Hemmings (1978) new features have been discovered about this species. The most important is the discovery of sexual characters by Long et al. (2015). Unfortunately, we are still ignorant of the plates of the ventral side of the head shield. What is almost always lacking is the tail, which is surprising as the preservation is usually excellent, e.g., the embryos of Watsonosteus fletti discovered by Newman et al. (2021). Hemmings (1978, plate 8, figure 1) figured GSE 13339 (referred to as GSE B507i by Hemmings), which she thought might be a detached tail. However, on close examination we consider this to be a weathering feature. A newly discovered specimen NMS G.2025.9.1 from Weems Castle in South Ronaldsay in Scotland (British Grid reference ND 433888) may have a preserved tail (Figure 13). The possible tail is preserved as a carbonised smear with no outline of its original shape, or any microscopic detail. The slight raised ridge along its length is a preparation glue line. It could be that this smear is the remains of body fluids that escaped the body cavity during diagenesis. However, this is even more unlikely to be preserved than a tail. We felt it is prudent to mention this specimen as its ephemeral nature may explain why the tail is not preserved in deposits of otherwise excellent preservation. Another explanation is that M. dicki did not have a tail, but we feel this is anatomically unlikely.
Since the Microbrachius dicki monograph by Hemmings (1978) new features have been discovered about this species. The most important is the discovery of sexual characters by Long et al. (2015). Unfortunately, we are still ignorant of the plates of the ventral side of the head shield. What is almost always lacking is the tail, which is surprising as the preservation is usually excellent, e.g., the embryos of Watsonosteus fletti discovered by Newman et al. (2021). Hemmings (1978, plate 8, figure 1) figured GSE 13339 (referred to as GSE B507i by Hemmings), which she thought might be a detached tail. However, on close examination we consider this to be a weathering feature. A newly discovered specimen NMS G.2025.9.1 from Weems Castle in South Ronaldsay in Scotland (British Grid reference ND 433888) may have a preserved tail (Figure 13). The possible tail is preserved as a carbonised smear with no outline of its original shape, or any microscopic detail. The slight raised ridge along its length is a preparation glue line. It could be that this smear is the remains of body fluids that escaped the body cavity during diagenesis. However, this is even more unlikely to be preserved than a tail. We felt it is prudent to mention this specimen as its ephemeral nature may explain why the tail is not preserved in deposits of otherwise excellent preservation. Another explanation is that M. dicki did not have a tail, but we feel this is anatomically unlikely.
The presence of three species of Microbrachius in Laurussia indicates a widely connected continent via seaways and rivers (Plax and Newman, 2021, figure 8). This is further enhanced by the recent discoveries in the uppermost Givetian of Svalbard (Newman et al., 2019; 2021) and the Middle Devonian of Germany (Otto, 1999; Newman et al., 2020). The species of Microbrachius from China are not well known, with the available material poorly preserved. If they are indeed species of Microbrachius then it suggests faunal connection not only within Laurussia but also much further afield.
ACKNOWLEDGEMENTS
The authors would like to thank Ursula Toom (Tallinn University of Technology) and Stig Walsh (National Museum of Scotland) for access to specimens in their care. We would also like to thank Gennady Baranov (Tallinn University of Technology) for photographic services. We also thank two anonymous for their insightful comments that greatly improve the quality of this paper.
REFERENCES
Ahlberg, P.E., Ivanov, A., Luksevics, E., and Mark-Kurik, E. 1999: Middle and Upper Devonian Correlation of the Baltic Area and Scotland based on fossil fishes., p. 6-8. In Luksevics, E., Stinkulis, G., and Kalnina, L. (eds.), The Fourth Baltic Stratigraphical Conference, Problems and methods of Modern Regional Stratigraphy.
Burrow, C., den Blaauwen, J., and Newman, M. 2020a. A redescription of the three longest known species of the acanthodian Cheiracanthus from the Middle Devonian of Scotland. Palaeontologia Electronica, 23:1-43.
https://doi.org/10.26879/1035
Burrow, C., Newman, M., and den Blaauwen, J. 2020b. Cheiracanthid acanthodians from the lower fossil fish bearing horizons (Eifelian, Middle Devonian) of the Orcadian Basin, Scotland. Scottish Journal of Geology, 57:1-13.
https://doi.org/10.1144/sjg2020-006
Burrow, C.J., den Blaauwen, J.L., Newman, M.J., and Davidson, R.G. 2016. The diplacanthid fishes (Acanthodii, Diplacanthiformes, Diplacanthidae) from the Middle Devonian of Scotland. Palaeoelectronica, 19.1.10A:1-83.
https://doi.org/10.26879/601
den Blaauwen, J.L., Newman, M.J., and Burrow, C.J. 2019. A new cheiracanthid acanthodian from the Middle Devonian (Givetian) Orcadian Basin of Scotland and its biostratigraphic and biogeographical significance. Scottish Journal of Geology, 55:166-177.
https://doi.org/10.1144/sjg2018-023
Hemmings, S.K. 1978. The Old Red Sandstone antiarchs of Scotland: Pterichthyodes and Microbrachius. Palaeontographical Society Monographs, 551:1-64.
https://doi.org/10.1080/25761900.2022.12131737
Lebedev, O.A., Mark-Kurik, E., Karatajūtė-Talimaa, V.N., Lukševičs, E., and Ivanov, A. 2009. Bite marks as evidence of predation in early vertebrates. Acta Zoologica, 90:344-356.
https://doi.org/10.1111/j.1463-6395.2008.00344.x
Long, J.A., Mark-Kurik, E., Johanson, Z., Lee, M.S.Y., Young, G.C., Zhu, M., Ahlberg, P.E., Newman, M., Jones, R., den Blaauwen, J., Choo, B., and Trinajstic, K. 2015. Copulation in antiarch placoderms and the origin of gnathostome internal fertilization. Nature, 517:196-199.
https://doi.org/10.1038/nature13825
Lyarskaya, L.A. 1981. Панцирные рыбы девона Прибалтики. Asterolepididae (Аrmoured Fishes from Devonian of the Baltic States. Asterolepididae). Zinatne, Riga. (In Russian with an English summary)
Mark-Kurik Е. 2000. The Middle Devonian fishes of the Baltic States (Estonia, Latvia) and Belarus. Courier Forschungsinstitut Senckenberg, 223:309-324.
Mark-Kurik, E. and Nemliher, J. 2003. Once more on the Middle Devonian Abava Beds, p. 38-41. In Plado, J. and Puura, I. (eds.), Eesti geoloogide neljas ülemaailmne kokkutulek. Eesti geoloogia uue sajandi künnisel. Konverentsi materjalid ja ekskursioonijuht (The 4th World Meeting of Estonian Geologists. Estonian Geology in the beginning of the new century. Conference materials and excursion guide). Eesti Geoloogia Selts, Tartu Ülikooli geoloogia instituut, Tartu. (In Estonian)
Mark-Kurik, E. and Põldvere, A. 2012. Devonian stratigraphy in Estonia: current state and problems. Estonian Journal of Earth Sciences, 61:33-47. https://doi.org/10.317/earth.2012.1.03
Mark-Kurik, E., Newman, M.J., Toom, U., and den Blaauwen, J. L. 2018. A new species of the antiarch Microbrachius from the Middle Devonian (Givetian) of Belarus. Estonian Journal of Earth Sciences, 67:3-13. https://doi.org/10.3176/earth.2017.22
Miles, R.S. and Westoll, T.S. 1963. Two new genera of coccosteid Arthrodira from the Middle Old Red Sandstone of Scotland, and their stratigraphical distribution. Transactions of the Royal Society of Scotland, 65:179-208. https://doi.org/10.1017/S0080456800012527
Miller, H. 1841. Old Red Sandstone or new walks in an old field. John Johnstone, Edinburgh.
Newman, M.J. and Trewin, N.H. 2008. Discovery of the arthrodire genus Actinolepis (class Placodermi) in the Middle Devonian of Scotland. Scottish Journal of Geology, 44:83-88.
https://doi.org/10.1144/sjg44010083
Newman, M.J., Mark-Kurik, E., den Blaauwen, J.L., and Zupiņš, I. 2015. Scottish Middle Devonian fishes in Estonia. Scottish Journal of Geology, 51:141-147.
https://doi.org/10.1144/sjg2014-006
Newman, M.J., den Blaauwen, J.L., and Tuuling, T. 2017. Middle Devonian coccosteid (Arthrodira, Placodermi) biostratigraphy of Scotland and Estonia. Scottish Journal of Geology, 53:63-69.
https://doi.org/10.1144/sjg-2016-012
Newman, M.J., Burrow, C.J., and den Blaauwen, J.L. 2019. The Givetian vertebrate fauna from the Fiskeklofta Member (Mimerdalen Subgroup), Svalbard. Part I. Stratigraphic and faunal review. Part II. Acanthodii. Norwegian Journal of Geology, 99:1-16. https://doi.org/10.17850/njg99-1-01.
Newman, M.J., Burrow, C.J., and den Blaauwen, J.L. 2020. A new species of ischnacanthiform acanthodian from the Givetian of Mimerdalen, Svalbard. Norwegian Journal of Geology, 99:619-631.
https://doi.org/10.17850/njg99-4-05
Newman, M.J., Burrow, C.J., den Blaauwen, J.L., and Giles, S. 2021. A new actinopterygian Cheirolepis jonesi nov. sp. from the Givetian of Spitsbergen, Svalbard. Norwegian Journal of Geology, 101:1-14.
https://doi.org/10.17850/njg101-1-3
Newman, M.J., den Blaauwen, J.L., and Burrow, C.J. 2021. Two newly identified cheiracanthid acanthodians from the Mey Flagstone Formation (Givetian, Middle Devonian) of the Orcadian Basin, Scotland. Scottish Journal of Geology, 57:1-18. https://doi.org/10.1144/sjg2020-009
Newman, M.J., den Blaauwen, J., Burrow, C., and Jones, R. 2021. Earliest evidence (Middle Devonian, Givetian) of vertebrate embryos in the fossil record. Palaeontology, 64: 21-30.
Obruchev, D.V. 1943. Yoglinia n. g., последний представитель птераспид из среднего девона Ленинградской области (Yoglinia n. g., latest pteraspid from the Middle Devonian of the Leningrad District). Doklady Akademii Nauk SSSR, 41:41-43. (In Russian)
Obruchev, D.V. and Mark-Kurik, E. Yu. 1965. Псаммостеиды (Agnatha, Psammosteidae) девона СССР (Devonian Psammosteids (Agnatha, Psammosteidae) of the USSR). Institute of Geology, Academy of Sciences of the Estonian SSR, Tallinn.([In Russian with an English summary)
Otto, M. 1999. New finds of vertebrates in the Middle Devonian Brandenberg Group (Sauerland, Northwest Germany). Paläontologische Zeitschrift, 73:113-131.
https://doi.org/10.1007/BF02987986
Pan, J. 1984. A new species of Microbrachius from the Middle Devonian of Yunnan. Vertebrata PalAsiatica, 22:8-13. (In Chinese with an English abstract)
Plax, D.P. and Newman, M. 2021. Middle Devonian acanthodians from Belarus - new data and interregional biostratigraphy. Acta Geologica Polonica:71, 393-414. https://doi.org/10.24425/agp.2020.134568
Stensiö, E.A. 1948. On the Placodermi of the Upper Devonian of East Greenland. II, Antiarchi: subfamily Bothriolepinae. Palaeozoologica Groenlandica, 2:1-662.
Trinajstic, K., Boisvert, C., Long, J.A., and Johanson, Z. 2019. Evolution of vertebrate reproduction, p. 207-226. In Johanson, Z., Underwood, C., and Richter, M. (eds.), Evolution and development of fishes. Cambridge University Press. https://doi.org/10.1017/9781316832172.013
Wang, J.-Q. and Zang, G.R. 1999. New material of Microbrachius from the Lower Devonian of Qujing, Yunnan, China. Vertebrata PalAsiatica, 37:200-211. (In Chinese with an English summary)
Young, G.C. and Gorter, J.D. 1981. A new Fish fauna of the Middle Devonian age from the Taemas/Wee Jasper region of New South Wales. Bureau of Mineral Resources, Geology and Geophysics Bulletin, 209:83-147.
