

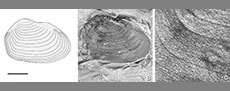
 Norestheria (Spinicaudata, Crustacea): Morphology and its stratigraphical and geographic significance
Norestheria (Spinicaudata, Crustacea): Morphology and its stratigraphical and geographic significance
Article number: 28.3.a48
https://doi.org/10.26879/1564
Copyright Palaeontological Association, October 2025
Author biographies
Plain-language and multi-lingual abstracts
PDF version
Submission: 13 April 2025. Acceptance: 7 October 2025.
ABSTRACT
A new species of the genus Norestheria, N. franconica n. sp., is recorded from the Hassberge Formation of the southern part of the Central European Basin. Norestheria franconica shows the characteristic morphology of the genus as exemplified by N. barnaschi, the type species of Norestheria. It predates the type species from the lower Norian, which has suggested as an important index fossil of the lower Norian of the Central European Basin, thus extending the stratigraphical and palaeogeographic range of the genus. It also sheds light on the phylogenetic lineages of mid- to late Triassic spinicaudatans.
Gerd Geyer. Lehrstuhl für Geodynamik und Geomaterialforschung, Institut für Geographie und Geologie, Bayerische Julius-Maximilians-Universität Würzburg, Am Hubland, 97074 Würzburg, Germany. gerd.geyer@uni-wuerzburg.de
Jürgen Sell. Museum Terra Triassica, Gerichtsgasse 22, 97717 Euerdorf, Germany. selljuergen@t-online.de
Keywords: Spinicaudata; new species; Carnian; Central European Basin
Final citation: Geyer, Gerd, and Sell, Jürgen. 2025. Norestheria (Spinicaudata, Crustacea): Morphology and its stratigraphical and geographic significance. Palaeontologia Electronica, 28(3):a48.
https://doi.org/10.26879/1564
palaeo-electronica.org/content/2025/5694-norestheria-spinicaudata-crustacea
Copyright: October 2025 Palaeontological Association.
This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
creativecommons.org/licenses/by/4.0
https://zoobank.org/EDA4CE70-D744-4027-9C9F-C78FD3B1A7A3
INTRODUCTION
Spinicaudatan “conchostracans” or clam shrimps are among the most valuable index fossils in the non-marine strata of the Triassic in the Central European Basin (earlier termed the “Germanic Basin”). A biostratigraphic framework for the otherwise barely subdividable Keuper beds of this basin has been attempted and established by a number of publications by Heinz Kozur (e.g., Kozur and Mock, 1993; Kozur and Weems, 2007, 2010; Kozur et al., 2013). Not surprisingly, the pioneerig work of Kozur required minor to moderate amendments in terms of adjusted ranges and taxonomic corrections of taxa.
The genus Norestheria was established by Kozur and Weems (2010) based on a single species, N. barnaschi Kozur and Weems, 2010, known from four specimens from the Morsleben 52A drill-core of eastern Niedersachsen, northern Germany. The specimens were found in strata of the Arnstadt Formation (sample 132 at depth m 162.9-163.0 m; =119.6 m above the base of the Arnstadt Formation), representing a late Alaunian (= middle Norian) age. Additional material of Norestheria was recorded by Kozur and Weems (2010) from drill-core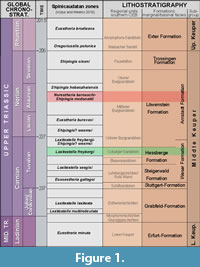 Malschenberg 1 of northern Baden-Württemberg, southwestern Germany, at depth m 60.0 and m 60.1-60.2, respectively, which represents the middle part of the Löwenstein Formation (= basal Stubensandstein 3) and thus the late Alaunian as well. This form was dealt with as Norestheria n. sp. These Norian occurrences led Kozur and Weems (2010) to name the genus Norestheria.
Malschenberg 1 of northern Baden-Württemberg, southwestern Germany, at depth m 60.0 and m 60.1-60.2, respectively, which represents the middle part of the Löwenstein Formation (= basal Stubensandstein 3) and thus the late Alaunian as well. This form was dealt with as Norestheria n. sp. These Norian occurrences led Kozur and Weems (2010) to name the genus Norestheria.
Kozur and Weems (2010) used Norestheria barnaschi as an index fossil for the Norestheria barnaschi-Shipingia macdonaldi Zone, which was not only used as age indicator within the Central European Basin, but also for correlation of the relevant strata into the basins of the Newark Subgroup of eastern North America (e.g., the Sanford Basin, North Carolina, and the Gettysburg Basin, Maryland and Pennsylvania).
In this study, we report another species of the genus Norestheria occurring in the Coburg Sandstone Member of the Hassberge Formation of northern Franconia, southern Germany. These strata represent the Tuvalian of the upper/late Carnian and thus predate the Norian occurrences of the genus Norestheria (Figure 1).
STUDY AREA, GEOLOGICAL SETTING AND ADDITIONAL FOSSILS
The spinicaudatans described herein were collected from the Coburg Sandstone member of the Hassberge Formation in the classical quarry area in northern Franconian, Bavaria. The situation has been described before in a number of studies (e.g., Geyer and Kelber, 2018) and is only shortly characterised below.
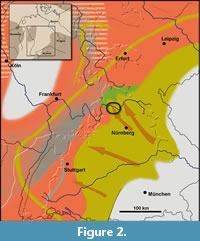 The Coburg Sandstone (German: Coburger Sandstein) forms the upper part of the Hassberge Formation in the siliciclastic marginal belt of the Keuper Basin. The succession features coarse- to fine-grained alluvial siliciclastics which are derived from the Vindelician and Bohemian High in the southeast. They form tongues that interfinger basinward with playa deposits composed of clay- and siltstones with intercalated thin dolostone beds of the central part of the Central European Basin (Figure 2). After an initial channel was formed, its filling by amalgamating, meandering channels led to sandstone beds of considerable thicknesses, accompanied by occasional slip-off slope and oxbow deposits (Hornung and Aigner, 2004).
The Coburg Sandstone (German: Coburger Sandstein) forms the upper part of the Hassberge Formation in the siliciclastic marginal belt of the Keuper Basin. The succession features coarse- to fine-grained alluvial siliciclastics which are derived from the Vindelician and Bohemian High in the southeast. They form tongues that interfinger basinward with playa deposits composed of clay- and siltstones with intercalated thin dolostone beds of the central part of the Central European Basin (Figure 2). After an initial channel was formed, its filling by amalgamating, meandering channels led to sandstone beds of considerable thicknesses, accompanied by occasional slip-off slope and oxbow deposits (Hornung and Aigner, 2004).
 Quarries in the Coburg Sandstone located in the Steigerwald and Hassberge regions expose successions of grey fluvial channel and sheet sandstone and intercalated varicoloured (light grey, greenish, yellow, and red) laminated lacustrine sedimentary rocks with small-scale cyclicity (Figure 3).
Quarries in the Coburg Sandstone located in the Steigerwald and Hassberge regions expose successions of grey fluvial channel and sheet sandstone and intercalated varicoloured (light grey, greenish, yellow, and red) laminated lacustrine sedimentary rocks with small-scale cyclicity (Figure 3).
Freyberg (1965) developed a subdivision of the member into twelve stratal units, which permit a quasi-chronological outline. Each stratal unit can be subdivided into single beds. A dominating more-or-less stratiform succession with intercalated playa-type deposits is punctuated by five major sandstone bodies of channel deposits, which are considerably incised into older strata. For additional data see, e.g., Freyberg (1965), Geyer (2002) and Geyer and Kelber (2018).
Fluvial siliciclastics occasionally yield remains of tetrapods, tetrapod tracks, fishes, and macroplant remains. The spinicaudatan conchostracans (Geyer and Kelber, 2018) are embedded in floodplain and lacustrine silt- and mudstones. These also yield insects (e.g., Kelber and Nitsch, 2005; Prokin et al., 2013; Fedorenko, 2014), notostracans (Trusheim, 1938; Kelber, 1998a, 1998b, 2021; Kelber and Nitsch, 2005), macroplant remains, palynomorphs, and various ichnofossils.
The spinicaudatans are mostly found as quite well-preserved bivalved carapaces which allows recognition of a considerable amount of morphological details.
LOCALITIES
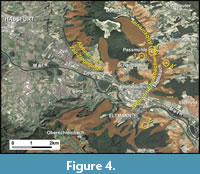 The spinicaudatans from the Coburg Sandstone member described herein have been collected from five quarries of the Ebelsbach-Eltmann quarry area (“Revier”; Freyberg 1965) between Schweinfurt and Bamberg, northern Bavaria, Germany (Figure 4).
The spinicaudatans from the Coburg Sandstone member described herein have been collected from five quarries of the Ebelsbach-Eltmann quarry area (“Revier”; Freyberg 1965) between Schweinfurt and Bamberg, northern Bavaria, Germany (Figure 4).
1. Roßberg North Quarry/Ankenbrand Quarry (no. 5930-3; Figure 4). Quarry 800 m east of Passmühle in the Ebelsbach valley and ca. 1 km west of Breitbrunn; quarry of the Fränkische Schleifsteinwerke GmbH/Brünner/Bamberger Natursteinwerke Glaser, formerly known as Ankenbrand Quarry.
Coordinates: 50° 00’ 55” N, 10° 40’ 55” E and R 44 05 670 H 55 44 210, respectively.
Fossils: Spinicaudatans occur in densely rhythmically laminated yellow to greenish floodplain mudstone/siltstones from 3.9 m above the lower ‘‘Werkstein’’ bed, occasionally associated with notostracans, fish remains, macroplants, and charcoal chunks (Kelber 1998b, 2007; Geyer and Kelber 2018).
2. Roßberg South Quarry/Ankenbrand Quarry (no. 5930-6; Figure 4). Abandoned quarry 200 m ENE of Schönbachsmühle in the Ebelsbach valley and ca. 1 km west of Breitbrunn; quarry of the Fränkische Schleifsteinwerke GmbH, formerly known as Ankenbrand Quarry.
Coordinates: 50° 00’ 40” N, 10° 40’ 45” E and R 44 05 420 H 55 42 720, respectively.
3. Quarry near Schönbachsmühle (no. 5930-1; Figure 4). Abandoned quarry 300 m west of Schönbachsmühle in the Ebelsbach valley.
Coordinates: 50° 00’ 40” N, 10° 40’ 15” E and R 44 04 830 H 55 42 540, respectively.
4. Quarry An der Finkenleite near Hermannsberg (no. 5930-4; Figure 3A, Figure 4). Quarry ca. 500 m SSW of Breitbrunn-Hermannsberg. Now abandoned and nearly completely filled.
Coordinates: 50° 00’ 25” N, 10° 41’ 50” E and R 44 06 720 H 55 41 970, respectively.
5. Hahn Quarry southeast of Eltmann (no. 6030-1; Figure 3B, Figure 4). Quarry to the southeast of Eltmann, known as Hahn Quarry.
Coordinates: 49° 57’ 35” N, 10° 40’ 20” E and R 44 04 890 H 55 36 840, respectively.
Fossils: Spinicaudatans recovered from the basal 30 cm of blue grey claystone resting directly on the basal lower ‘‘Werkstein’’ level. Specimens occur with remains of macroplants, scattered insects, notostracans, and charcoal particles. For a detailed section of the quarry, see Blankmeister (1989).
SYSTEMATIC PALAEONTOLOGY
Most of the material used for this study is in a repository of the collection Mainfränkische Trias Euerdorf (acronym SMTE) in the Museum Terra Triassica in Euerdorf. Additional specimens are in the collection of Klaus-Peter Kelber, Würzburg (acronym SKW).
The taxonomy adopted here follows that of Chen and Shen (1985), Martin and Davis (2001), Scholze and Schneider (2015) and Geyer and Kelber (2018), respectively.
Subphylum CRUSTACEA Brünnich, 1772
Class BANCHIOPODA Latreille, 1817
Order SPINICAUDATA Linder, 1945
Suborder SPRINICAUDATA Linder 1945
[nom. transl. Tasch, 1969,
ex ‘‘tribe Spinicaudata’’ Linder, 1945]
Superfamily EOSESTHERIOIDEA Zhang and Chen in Zhang et al., 1976
[=Lioestherioidea Raymond, 1946 sensu Zhang et al., 1976]
Discussion. See discussion in Geyer and Kelber (2018).
“Family EUESTHERIIDAE Defretin-Lefranc, 1965”
Discussion. The family concepts of the Spinicaudata and particularly those within the Eosestherioidea are generally based on morphological features of the carapace, which are poorly understood in terms of evolutionary trends. Accordingly, no robust data exist that support the presently recognised families and their generic compositions (e.g., Hegna and Astrop, 2020). As an example for the resulting confusion, the genus Pseudestheria has been alternately assigned to four families in the past 20 years: Euestheriidae, Lioestheriidae, Pseudestheriidae, and Orthothemosiidae. The first three families are poorly defined and tied to type genera which are dependant on type species that have never been adequately figured (e.g., Poschmann et al., 2024). In particular, the family Euestheriidae is presently ill-defined (see, e.g., Fang and Liao, 2024; Poschmann et al., 2024). Part of the confusion arises from quite obvious differences in the specimens identified as Euestheria minuta (Alberti in Zieten, 1833), the type species of Euestheria Depéret and Mazeran (1912) exemplified their generic concept based on material identified as E. minuta from the Permian of Autun, France, which differs considerably from the specimens from the Lower Keuper beds of southern Germany, on which the species has been based. This brought Defretin-Lefranc (1965) to distinguish three groups with different surface ornaments (termed “structure lisse ou ponctuée”, “structure réticulée”, and “réticule-radiée et structure radiée”). Consequently, the species from the Permian of Autun represents a different genus than the species from the Lower Keuper beds of southern Germany. They could perhaps be assigned to Lioestheria Depéret and Mazeran, 1912 or may represent a different, not yet established genus (see also Sell, 2018).
Here, we assume that most of the spinicaudatans known from the Carnian through Norian beds of the Central European Basin and also of the Newark Subbasin reflect intricate morphological developments of a single clade, although this, in turn, does not suggest a geographical argument for the evolution of the group. We refer this clade provisionally to the family “Euestheriidae” although more data of analyses of morphological characters of the carapace as well as knowledge on soft-part anatomy would be needed to establish robust taxonomic units.
The genus Norestheria Kozur and Weems, 2010 has been placed under the family Shipingiidae Kozur and Weems, 2005 by Kozur and Weems (2010), but lacks the typical oval to elliptic, cycladiform shape of the carapace with a nearly straight dorsal margin, and an umbo in anterior, marginal or nearly marginal position.
Genus Norestheria Kozur and Weems, 2010
Type species. Norestheria barnaschi Kozur and Weems, 2010, by original designation; from the Arnstadt and Löwenstein formations, Alaunian, middle Norian, of the Central European Basin.
Diagnosis (emended herein). Genus of the Euestheriidae with dorsal margin straight to slightly convex, fairly short, grading into the anterior margin and a mostly indistinct posterodorsal corner; anterior margin more extended than posterior margin, in lower part with a long oblique region that changes its curvatures from barely curved in the lower and a slight curvature in its upper part; ventral margin poorly bent along its anterior and middle course, slightly convex in its posterior part, with an abrupt change in curvature creating a nearly angular course from the ventral to the anterior margin; transition from the ventral to the posterior margin along a gradual curvature. Growth bands variable, straight to nearly straight in the ventral and anteroventral sectors, but often more tilted and obliquely oriented in the lower part of the anterior quarter; microsculpture a dense reticulate pattern, partly accompanied by radial fringes and lirae.
Discussion. Kozur and Weems (2010, p. 403) presented a comprehensive diagnosis for the genus, which has the character of a description and does not pinpoint the quasi-apomorphic characters. The main feature that allows to recognize Norestheria barnaschi is the presence of a nearly arcuate course of the margin from the ventral to the anterior sector. The same feature can be seen in Norestheria franconica n. sp. as well as in the informal species Norestheria n. sp. suggested by Kozur and Weems (2010).
Another significant character claimed for Norestheria barnaschi is the rapid change in the direction of the growth bands from the anterior quarter to the rest of the shell exterior. The authors emphasize that this change of the tilting is seen in Norestheria n. sp. as well. However, Kozur and Weems (2010) admitted that this feature may be enhanced by deformation. Indeed, some of the specimens of N. franconica partly present this change in inclination of the growth ridges (e.g., SMTE 5930-4-576), whereas it seems not to be developed in others (e.g., SMTE 5930-1-198) so that we tend to assume that the major expression of this feature must be ascribed to a (resulting) dorsoventral compaction (or rather compression) of the valves which happens to occur in the Keuper beds during early diagenetic effects oblique to the general bedding planes.
Among the similar and certainly closely related genera is Laxitextella Kozur, 1982, which has a similar microsculpture, but this ornamentation does not become larger in the anterior sector of the valves. Furthermore, Laxitextella is distinguished by is its “normal” ovoidal outline of the valves and has a stronger curvature in the umbonal area.
Euestheria Depéret and Mazeran, 1912 had developed into a waste bin for Triassic clam shrimps but underwent a number of considerable shifts in taxonomy and nomenclature during the last decades (see discussion above and also in Geyer and Kelber, 1998). Nonetheless, generic concepts still differ, and imperfect preservation of several critical species led to persisting confusion. Species such as “ Euestheria” kozuri Geyer and Kelber, 1998 (from the Coburg Sandstone member of the Hassberge Formation) and two unnamed forms from the same stratigraphic interval appear to represent transitional forms that bracket the gap between the early representatives of “Euestheria” (such as E. minuta Alberti in Zieten, 1833; but see discussion above) and Laxitextella. In developing a small-scale reticulate ornament between the growth lines but retain a more protruding umbo and lack the angulation seen at the valve margin of Norestheria.
The relatively conspicuous change in curvature of the margin and the corresponding growth ridges creating a nearly angular course from the ventral to the anterior margin is quite unique in Triassic spinicaudatans from the northern hemisphere. However, the family Ulugkemiidae includes other clam-shrimps from the Devonian through Permian of the Pre-Baltic and Pre-Uralian region of Russia and the Triassic of Argentina with abrupt changes in the curvature of growth-bands, such as Beligum Novozhilov, 1958, Ulugkemia Novozhilov, 1958, and Triasulugkemia Gallego and Melchor, 2000. However, those genera show only anteroventral and posteroventral indentations or solely an anteroventral indentation depicted by the growth ridges in nearly regularly ovate carapaces - quite different in character from the course of the carapace margin in Norestheria.
Norestheria franconica n. sp.
Figure 5, Figure 6, Figure 7
zoobank.org/58F35DDD-9658-4CA7-A269-DC0243EBB5D2
v. Euestheria? sp. nov. A; Geyer and Kelber, 2018, p. 13-15, fig. 8A, C.
Etymology. A reference to the geographic occurrence of the species in the Franconian region, southern Germany.
Holotype. Specimen SKW ELT-C-12a, carapace composed of two slightly rotated valves, described and figured as Euestheria? sp. nov. A in Geyer and Kelber (2018, fig. 8A, 8C).
Type locality. Hahn Quarry, SE of Eltmann, Franconia.
Type stratum. Coburg Sandstone member, Hassberge Formation; Laxitextella freybergi Zone, late Tuvalian, upper/late Carnian.
Paratypes. 48 specimens, all from Coburg Sandstone member of the Hassberge Formation; 5 specimens from Roßberg North Quarry (SMTE 5930-3-8, -39, -45, -64a, -121), all from stratal unit 2; 6 specimens from quarry near Schönbachsmühle (SMTE 5930-1-197, -198a, -199Ia/-199IIa, -199Ib, -200, -201), all from stratal unit 10; single specimen from Roßberg South Quarry (SMTE 5930-6-40); 14 specimens from Finkenleite quarry near Hermannsberg (SMTE 5930-4-576a, -576b, -577a, -577b, -582a, -582b, -582c, -585, -588I, -589a, -599a, -599b, -673a, -673b), all from stratal unit 2; 41 specimens from Hahn quarry near Eltmann (SMTE 6030-1-120Ia/-120IIa, -120Ib/120IIb, -120Ic/-120IIc, -210Ia/-210IIa, -218I/-218II, -254a, -254b, -255a, -255b, -255c, -283, -390a, -390b, -496Ia/-496IIa, -496Ib. -496Ic, -496Id, -500a, -500b, -512a, -517a, -517b, -517c, -517d, -528, -531, -544, -547, -623, -625, -626a, -626b, -626*, -628, -638a, -638b, -640I/-640II, -641, -657, -659a, -659b), all from stratal unit 5. Tentatively assigned to N. franconica: SKW ELT-C-12b (from the same slab as the holotype).
Diagnosis. Species of Norestheria with moderately large carapace (ca. 2 to 3 mm long in adult individuals); umbonal area small; dorsal margin with subangular posterodorsal edge; anterior margin with slightly curved upper section, lower section almost straight, obliquely directed; growth bands up to ca. 25, directed steeply oblique to axis in the anterior sector.
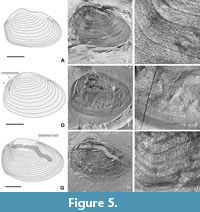 Description. Carapace moderately large, length up to 8 mm and height up to 5.5 mm, slightly convex. Umbonal area distinct, relatively small, marginal to slightly supramarginal, located submedially. Dorsal margin very short to short, straight to faintly, length 45 to 55% maximum length of carapace; posterodorsal corner blunt and indistinct. Posterior margin clearly less high than anterior margin, its middle section strongly curved, posterodorsal section straight to slightly curved, lower part gently bent (e.g., Figure 5B, E, H). Anterior margin consists of distinctly curved upper section, anteroventral section poorly curved and directed distinctly oblique, with steeply inclined ventralward; anterior and ventral margins connected at a blunt corner, which becomes increasingly more strongly curved during ontogeny and thus develops into a distinct semi-angular transition from the ventral to the anterior margin (e.g., Figure 6C, D, G, J). Ventral margin with moderate curvature; its anterior and middle section slightly curved, posterior sector more strongly curvilinear. Growth bands distinct, variable in number (from ca. 10 to 25); with sharp face towards margin of the carapace, subevenly developed throughout most of the valve, but growth bands more steeply inclined in the anterior quarter of the valve; growth lines thin, moderately prominent, smooth strips, confined on both sides by a sharp, shallow groov
Description. Carapace moderately large, length up to 8 mm and height up to 5.5 mm, slightly convex. Umbonal area distinct, relatively small, marginal to slightly supramarginal, located submedially. Dorsal margin very short to short, straight to faintly, length 45 to 55% maximum length of carapace; posterodorsal corner blunt and indistinct. Posterior margin clearly less high than anterior margin, its middle section strongly curved, posterodorsal section straight to slightly curved, lower part gently bent (e.g., Figure 5B, E, H). Anterior margin consists of distinctly curved upper section, anteroventral section poorly curved and directed distinctly oblique, with steeply inclined ventralward; anterior and ventral margins connected at a blunt corner, which becomes increasingly more strongly curved during ontogeny and thus develops into a distinct semi-angular transition from the ventral to the anterior margin (e.g., Figure 6C, D, G, J). Ventral margin with moderate curvature; its anterior and middle section slightly curved, posterior sector more strongly curvilinear. Growth bands distinct, variable in number (from ca. 10 to 25); with sharp face towards margin of the carapace, subevenly developed throughout most of the valve, but growth bands more steeply inclined in the anterior quarter of the valve; growth lines thin, moderately prominent, smooth strips, confined on both sides by a sharp, shallow groov e (Figure 6D). Microsculpture consists of reticulate pattern (e.g., Figure 6C, F, L), with meshes of ca. 5 µm on the growth bands of the juvenile part of the valves, maximum size ca. 25 to 30 or 35 µm on growth bands of the adult part of the valves (depending on their relative position on the valves).
e (Figure 6D). Microsculpture consists of reticulate pattern (e.g., Figure 6C, F, L), with meshes of ca. 5 µm on the growth bands of the juvenile part of the valves, maximum size ca. 25 to 30 or 35 µm on growth bands of the adult part of the valves (depending on their relative position on the valves).
The growth lines are thin smooth strips, which can be seen in well preserved specimens to be confined on both sides by a sharp, shallow groove. The microsculpture between the growth lines consists of a dense reticulate network that is often quite irregularly developed and accompanied by radial fringes and lirae in some areas of the valves and in a considerable number of specimens. Dorsoventral compression (see under genus) certainly intensified these lirae.
Remarks. Norestheria barnaschi Kozur and Weems, 2010 has a microsculpture that consists of a dense and delicate network of radial lirae. The size of the meshes changes rapidly at approximately one-quarter of the valve length into larger elements. Kozur and Weems (2010, fig. A5,5 and A5,6) figured a second, informal new species of Norestheria from the Löwenstein Formation, which has a microsculpture that consists only of dense and fine meshes which also changes abruptly into a coarse reticulate pattern at the turning line of curvature of the growth lines. The microsculpture in N. franconica consists of a reticulate pattern that does not change the widths of the meshes into anterior direction.
Modern spinicaudatans have been reported to show three reproductive strategies, i.e. dioecy, hermaphroditism and parthenogenesis (e.g., Weeks et al., 2007, 2008; Brantner et al., 2013). Female spinicaudatan individuals may carry up to several hundreds of resting eggs, or cysts, in a subumbilical area between the back and the carapace. These cysts are quite resistant against drought and other factors as in all branchiopods, and the embryos remain able to hatch after many years even under extreme conditions (such as long dry periods, extreme heat or strong frost). Some specimens from the Hahn quarry and also from the quarry near Schönbachsmühle were preserved with clusters of densely packed cysts located in the dorsal part of the carapace (Figure 7).
 All cysts were found inside the carapace (Figure 7). This suggests that they were still attached to the body during the moment of embedding. The cysts are now mostly flattened and preserved quite dimly. Their surfaces were affected by structures of the carapace and are thus further deformed. However, they appear relatively globular and with a smooth surface and carbon staining. Some of the cysts with a preservation that appear to reliably reflect the original size and morphology have been measured and are approximately 100 μm in diameter (thus of similar size as documented for other clam shrimps by Poschmann et al., 2024). The number of cysts on the samples varies between 190 und 290.
All cysts were found inside the carapace (Figure 7). This suggests that they were still attached to the body during the moment of embedding. The cysts are now mostly flattened and preserved quite dimly. Their surfaces were affected by structures of the carapace and are thus further deformed. However, they appear relatively globular and with a smooth surface and carbon staining. Some of the cysts with a preservation that appear to reliably reflect the original size and morphology have been measured and are approximately 100 μm in diameter (thus of similar size as documented for other clam shrimps by Poschmann et al., 2024). The number of cysts on the samples varies between 190 und 290.
A superficially similar phenomenon can be seen in specimen SMTE 5930-4-577a (Figure 6B). However, the apparent egg-bearing area in this specimen results from the insertion of quartz grains. These have diameters in the range of approximately 150 to 200 μm.
Norestheria franconica is found in two different forms, which clearly suggests the presence, or absence of a brood chamber. This dimorphism led to a size of the carapace which is distinctly higher at the female specimens (height/length ratio 0.72 to 0.77) than at male specimens (height/length ratio 0.64 to 0.68) (Table 1).
STRATIGRAPHIC SIGNIFICANCE AND CONCLUSIONS
As mentioned above, Kozur and Weems (2010) used Norestheria barnaschi as an index fossil for their Norestheria barnaschi-Shipingia mcdonaldi Zone, introduced as a distinct biozone for the Alaunian, middle Norian, replacing the lower part of the Shipingia hebaozaiensis Zone of Kozur and Weems (2007). That zone is based on the (locally!) common occurrence of Norestheria barnaschi and Shipingia mcdonaldi, with Euestheria sp. being the only other spinicaudatan known from that zone. Kozur and Weems (2010) referred to the Malschenberg 1 borehole (Etzold and Franz, 2005) as the “type locality” for the zone.
The occurrences of Norestheria barnaschi in the Morsleben 52A and in the Malschenberg 1 drill-cores ties the basinal playa facies of the Central European Basin (= Arnstadt Formation) with its marginal arenitic facies (Löwenstein Formation). Shipingia macdonaldi was identified from occurrences without Norestheria in an intermediate facies belt (“Steinmergelkeuper”) of Thuringia, central Germany, from the Wachsenburg Sandstone (= middle Arnstadt Formation).
Shipingia mcdonaldi Kozur and Weems, 2010 was originally determined as being early representatives of Shipingia hebaozaiensis Shen, 1976. Kozur and Weems (2010) reinterpreted these specimens as representing a different species. The type material comes from the Heidlersberg Member of the Gettysburg Formation of the Newark Subgroup, but the species was also identified from the Wachsenburg Sandstone (Arnstadt Formation) and the Stubensandstein 3 (Löwenstein Formation) of the southern Central European Basis. This enabled Kozur and Weems (2010) to identify the Norestheria barnaschi-Shipingia mcdonaldi Zone not within the Central European Basin but also in the Newark Subgroup of eastern the United States, i.e. from the Heidlersburg Member of the Gettysburg Formation of the Gettysburg Basin, Pennsylvania, and from the Sanford Formation of the Sanford Subbasin, Deep River Basin, North Carolina.
This study features another, closely related species of Norestheria, N. franconica n. sp., from the Tuvalian of the upper/late Carnian of the southern part of the Central European Basin. These strata clearly belong to the Laxitextella freybergi Zone as described and discussed in some in detail by Geyer and Kelber (2018). These separated occurrences again undermine a more or less tacit supposition that most of the Carnian through Rhaetian “conchostracan” zones reflect phylogenetic developments within the spinicaudatans. As in the case of the genera Laxitextella and Shipingia, stratigraphic occurrences do not fit with the assumed ancestor-descendant relationships. More material will be needed to settle the momentarily largely accepted, but possibly overaccurate biozonation for the Central European Basin.
ACKNOWLEDGEMENTS
We thank T.A. Hegna (Fredonia, NY) and an anonymous reviewer for constructive reviews of the manuscript.
REFERENCES
Bachmann, G.H. and Kozur, H.W. 2004. The Germanic Triassic: correlations with the international scale, numerical ages and Milankovitch cyclicity. Hallesches Jahrbuch für Geowissenschaften, B 26:17-62.
Berger, H.A.C. 1854. Die Keuperformation mit ihren Conchylien in der Gegend von Coburg beschrieben. Neues Jahrbuch für Mineralogie, Geognosie, Geologie und Petrefakten-Kunde, 1854:408-414.
Beutler, G. and Nitsch, E. 2005. Paläogeographischer Überblick. In Beutler, G., Hauschke, N., Nitsch, E. and Vath, U. (eds.), Stratigraphie von Deutschland IV. Keuper. Courier Forschungsinstitut Senckenberg, 253:15-30.
Blankmeister, B. 1989. Grüner, Weißer und Gelber Mainsandstein. Dokumentation und Materialkartierung im Umfeld einiger Abbaue bei Bamberg (N-Bayern). Unpublished diploma thesis. Institut für Geologie, Ruhr-Universität Bochum, Bochum, 61 pp.
Brantner, J.S., Ott, D.W., Duff, J., Sanoamuang, L.-o., Simhachalam, G.P., Subash Babu, K.H., and Weeks, S.C. 2013. Androdioecy and hermaphroditism in five species of clam shrimps (Crustacea: Branchiopoda: Spinicaudata) from India and Thailand. Invertebrate Biology, 132(1): 27-37.
https://doi.org/10.1111/ivb.12012
Brünnich, M.T. 1772. Zoologiae fundamenta praelectionibus aca demicis accomodata. Grunde i Dyrelaeren, 254 pp. Fridericus Christianus Pelt, Hafniae et Lipsiae [Copenhagen and Leipzig].
Defretin-Lefranc, S. 1965. Étude et révision de Phyllopodes Conchostracés en provenance d’U.R.S.S. Annales de la Société Géologique du Nord, 85:15-48.
Depéret, C. and Mazeran, P. 1912. Les Estheria du Permien d’Autun. Bulletin de la Société d’Histoire Naturelle d’Autun, 25:165-174.
Fang, S. and Liao, H. 2024. A worldwide annotated checklist of fossil (Devonian-Cretaceous) species of the clam shrimp genus Euestheria (Branchiopoda: Diplostraca: Spinicaudata). Life, 14(1438):1-25.
https://doi.org/10.3390/life14111438
Fedorenko, D.N. 2014. Wings of Coptoclavidae (Coleoptera, Adephaga) from the Keuper (Triassic) of Franconia, Germany. Paleontologicheskiy Zhurnal’, 2014(6):32-36; and Paleontological Journal, 48:605-609.
https://doi.org/10.1134/S0031030114060069 (in Russian and English, respectively)
Freyberg, B. von 1965. Der Coburger Bausandstein (Mittlerer Keuper) von Zeil-Ebelsbach als Beispiel einer epikontinentalen Schichtfolge. Erlanger Geologische Abhandlungen, 58:1-60.
Gallego, O.F. and Melchor, R.N. 2000. La familia Ulugkemiidae Novozhilov, 1958 (Conchostraca) en el Triásico de Argentina: implicancias paleobiogeográficas. Ameghiniana, 37:47-58.
Geyer, G. 2002. Geologie von Unterfranken und angrenzenden Regionen. Fränkische Landschaft, Arbeiten zur Geographie von Franken, 2:1-588. Klett-Perthes, Gotha.
Geyer, G. and Kelber, K.-P. 2018. Spinicaudata (“Conchostraca”, Crustacea) from the Middle Keuper (Upper Triassic) of the southern Germanic Basin, with a review of Carnian-Norian taxa and suggested biozones. Paläontologische Zeitschrift, 92:1-34.
https://doi.org/10.1007/s12542-017-0363-7
Hegna, T.A. and Astrop, T.I. 2020. The fossil record of the clam shrimp (Crustacea; Branchiopoda). Zoological Studies, 59, Article 43:1-9.
https://doi.org/10.6620/zs.2020.59-43
Hornung, J. and Aigner, T. 2004. Sedimentäre Architektur und Poroperm-Analyse fluviatiler Sandsteine: Fallbeispiel Coburger Sandstein, Franken. Hallesches Jahrbuch für Geowissenschaften, B, Beiheft 18:121-138.
Jones, R. 1862. A Monograph of the Fossil Estheriae. VIII + 134 pp. Palaeontographical Society, London.
Kelber, K.-P. 1998a. New triopsids (Crustacea, Notostraca) from the Upper Triassic of Franconia, Germany. Epicontinental Triassic International Symposium, Halle/Saale, Germany, September 21-23, 1998. Hallesches Jahrbuch für Geowissenschaften, Beiheft 5:85.
Kelber, K.-P. 1998b. Triops cancriformis (Crustacea, Notostraca): Ein bemerkenswertes Fossil aus der Trias Mitteleuropas, p. 383-394. In Hauschke, N. and Wilde, V. (eds.), Trias. Eine ganz andere Welt. Friedrich Pfeil, München.
Kelber, K.-P. 2021. Triopsiden (Crustacea, Notostraca) im Mittleren Keuper (Hassberge-Formation) Frankens, p. 455-462. In Hauschke, N., Franz, M. and Bachmann, G. (eds.), Trias. Aufbruch in das Erdmittelalter, 2. Verlag Dr. Friedrich Pfeil, München.
Kelber, K.-P. and Nitsch, E. 2005. Paläoflora und Ablagerungsräume im unterfränkischen Keuper (Exkursion H am 1. April 2005). Jahresberichte und Mitteilungen des Oberrheinischen Geologischen Vereins, Neue Folge, 87:217-253.
Kozur, H. 1982. Beiträge zur Taxonomie und stratigraphischen Auswertung der untertriassischen Conchostracen. Geologisch Paläontologische Mitteilungen Innsbruck, 11:355-398.
Kozur, H.W. and Mock, R. 1993. The importance of conchostracans for the correlation of continental and marine beds, p. 261-266. In Lucas, S.G. and Morales, M. (eds.), The nonmarine Triassic. Bulletin of the New Mexico Museum of Natural History and Science, 3.
Kozur, H.W. and Weems, R.E. 2007. Upper Triassic conchostracan biostratigraphy of the continental rift basins of eastern North America: its importance for correlating Newark Supergroup events with the Germanic Basin and the international geologic time scale, p. 137-188. In Lucas, S.G. and Spielmann, J.A. (eds.), The Global Triassic. Bulletin of the New Mexico Museum of Natural History and Science, 41.
Kozur, H.W. and Weems, R.E. 2010. The biostratigraphic importance of conchostracans in the continental Triassic of the northern hemisphere, p. 315-417. In Lucas, S.G. (ed.), The Triassic Timescale. Geological Society Special Publications, 334. The Geological Society of London, London.
https://doi.org/10.1144/SP334.13
Kozur, H.W., Franz, M., and Bachmann, G.H. 2013. Shipingia weemsi n. sp., a biostratigraphically important conchostracan species from the uppermost Carnian and lowermost Norian of Central Europe, p. 325-330. In Tanner, L.H., Spielmann, J.A. and Lucas, S.G. (eds.). The Triassic System. Bulletin of the New Mexico Museum of Natural History and Science, 61.
Latreille, P.A. 1817. Le Règne Animal. Tome III, Contenant les Crustacés, les Arachnides et les Insectes, 1-653. A. Bedin, Paris.
Linder, F. 1945. Affinities within the Branchiopoda with notes on some dubious fossils. Arkiv för Zoologi, 37A: 1-28.
Martin, J.W., and Davies, G.E. 2001. An updated classification of the recent Crustacea. Natural History Museum of Los Angeles County, Science Series, 39:1-124.
Nitsch, E. 2005. Zyklostratigraphie des Keupers, p. 106-133. In Beutler, G., Hauschke, N., Nitsch, E., and Vath, U. (eds.), Deutsche Stratigraphische Kommission, Stratigraphie von Deutschland IV. Keuper. Courier Forschungsinstitut Senckenberg, 253.
Novozhilov, N.I. 1958. Recueil d’articles sur les phylopodes conchostracès. Annales de la Service d’Information Géologique du Bureau de Recherches Géologiques, Géophysiques, et Minières, 6:1-135.
Poschmann, M.J., Hegna, T.A., Astrop, T.I., and Hoffmann, R. 2024. Revision of Lower Devonian clam shrimp (Branchiopoda, Diplostraca) from the Rhenish Massif (Eifel, SW-Germany), and the early colonization of non-marine palaeoenvironments. Palaeobiodiversity and Palaeoenvironments, 104:535-569.
https://doi.org/10.1007/s12549-023-00597-9
Prokin, A.A., Makarov, K.V., Ponomarenko, A.G., and Bashkuev, A.S. 2013. New beetle larvae (Coleoptera: Coptoclavidae, Caraboidea, Polyphaga) from the Upper Triassic of Germany. Russian Entomological Journal, 22:259-274.
Raymond, R.E. 1946. The genera of fossil Conchostraca--an order of bivalved Crustacea. Bulletin of the Museum of Comparative Zoology, 96:217-311.
Reible, P. 1962. Die Conchostraken (Branchiopoda, Crustacea) der germanischen Trias. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, 114:169-244.
Richter, S., Olesen, J., and Wheeler, W.C. 2007. Branchiopod phylogeny - a combined analysis using morphology and six molecular loci. Cladistics, 23:301-336.
https://doi.org/10.1111/j.1096-0031.2007.00148.x
Schlirf, M., Uchman, A., and Kümmel, M. 2001. Upper Triassic (Keuper) non-marine trace fossils from the Haßberge area (Franconia, south-eastern Germany). Paläontologische Zeitschrift, 75:71-96.
https://doi.org/10.1007/BF03022599
Scholze, F. and Schneider, J.W. 2015. Improved methodology of ‘conchostracan’ (Crustacea: Branchiopoda) classification for biostratigraphy. Newsletters on Stratigraphy, 48:287-298.
https://doi.org/10.1127/nos/2015/0065
Seegis, D. 1999. Die Wirbellosen-Fauna des Keupers: Zusammensetzung und ökologische Aussagemöglichkeiten, p. 371-382. In Hauschke, N. and Wilde, V. (eds.), Trias. Eine ganz andere Welt. Friedrich Pfeil, München.
Sell, J. 2018. Die Euestheriidae (Conchostraca) des Oberen Muschelkalks und Unteren Keupers von Unterfranken. Semana, 33:55-90.
Tasch, P. 1969. Branchiopoda, p. R128-R191. In Moore, R.C. (ed.), Treatise on Invertebrate Paleontology. Part R, Arthropoda 4, Volume 1. Geological Society of America, Boulder, CO, and University of Kansas Press, Lawrence, KS.
Trusheim, F. 1938. Triopsiden (Crust. Phyll.) aus dem Keuper Frankens. Paläontologische Zeitschrift, 19:198-216.
Weeks, S.C., Chapman, E.G., Rogers, D.C., Senyo, D.M., and Hoeh, W.R. 2009. Evolutionary transitions among dioecy, androdioecy and hermaphroditism in limnadiid clam shrimp (Branchiopoda: Spinicaudata). Journal of Evolutionary Biology, 22:1781-1799.
https://doi.org/10.1111/j.1420-9101.2009.01813.x
Weeks, S.C., Sanderson, T.F., Zofkova, M., and Knott, B. 2008. Breeding systems in the clam shrimp family Limnadiidae (Branchiopoda, Spinicaudata). Invertebrate Biology, 127:336-349.
https://doi.org/10.1111/j.1744-7410.2008.00130.x
Weems, R.E. and Lucas, S.G. 2015. A revision of the Norian conchostracan zonation in North America and its implications for Late Triassic North American tectonic history, p. 303-318. In Sullivan, R.M. and Lucas, S.G. (eds.), Fossil Record 4. New Mexico Museum of Natural History and Science Bulletin, 67.
Zhang, W.T. (‘‘Chang’’), Chen, P.J. and Shen, Y.B. 1976. Fossil Conchostraca of China, 1-325. Science Press, Beijing. (in Chinese)
Zieten, C.H.V. 1833. Die Versteinerungen Württembergs, oder naturgetreue Abbildungen der in den vollständigsten Sammlungen, namentlich der in dem Kabinet des Oberamts-Arzt Dr. Hartmann befindlichen Petrefacten, mit Angabe der Gebirgs-Formationen, in welchen dieselben vorkommen und der Fundorte, Teil 12. i-ii, 89-102. E. Schweizerbart’sche Verlagsbuchhandlung, Stuttgart.
